
Investigating the Effects of Atrial Natriuretic Peptide on the Maternal Endothelium to Determine Potential Implications for Preeclampsia

 , , ,
, , , 
Abstract
1. Introduction
2. Results
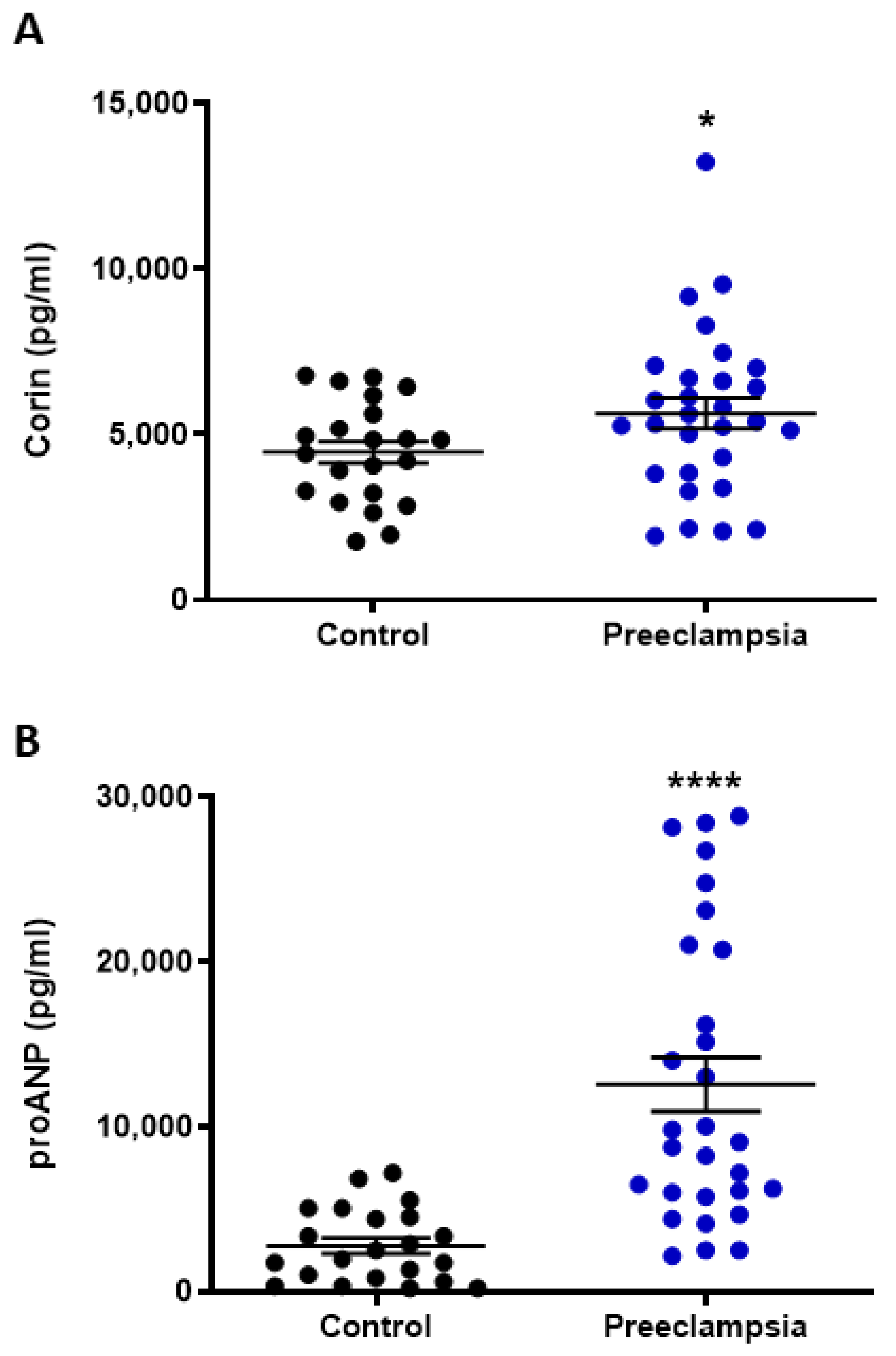
2.1. Circulating Levels of Corin and NT-proANP Increased with Preeclampsia
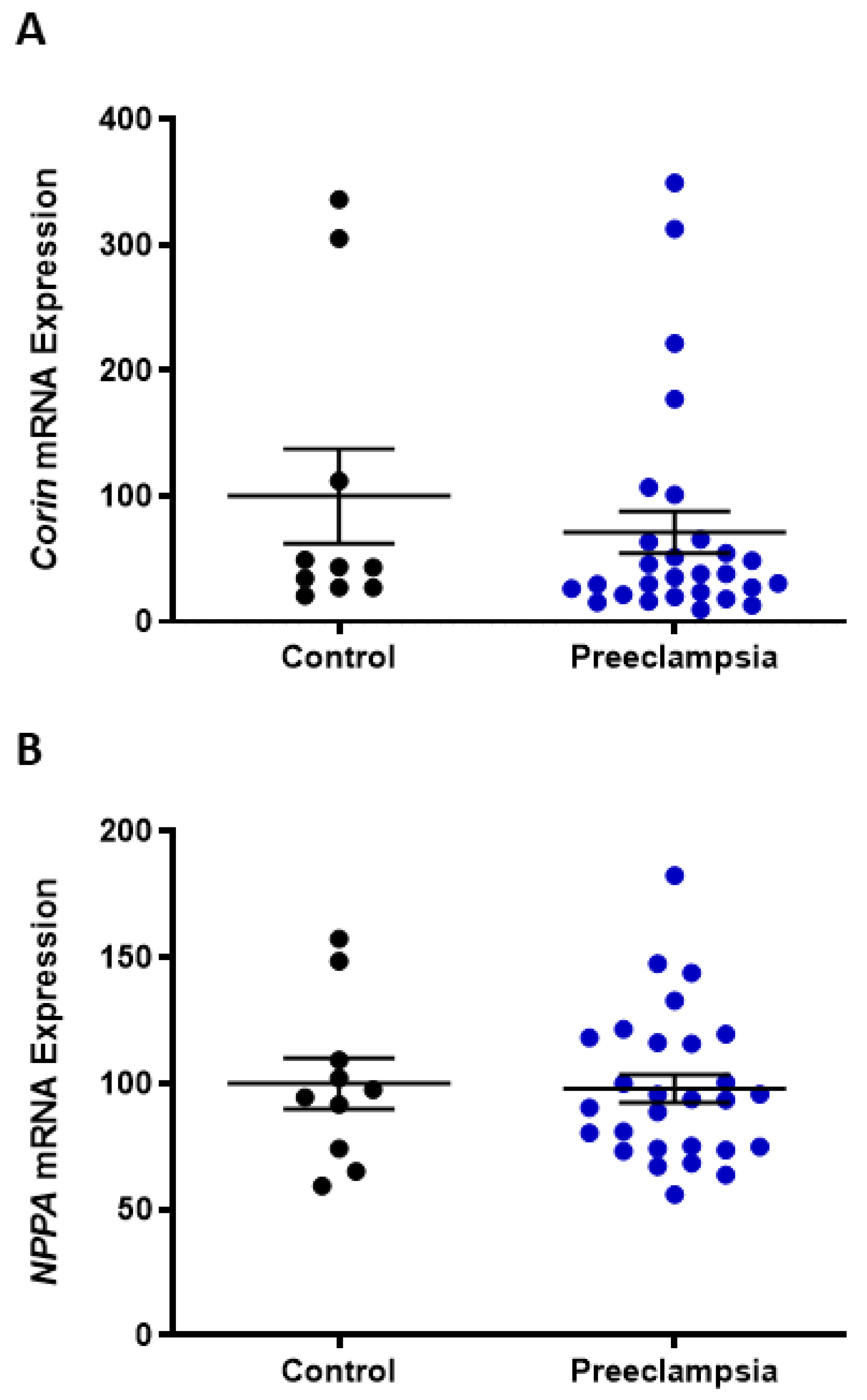
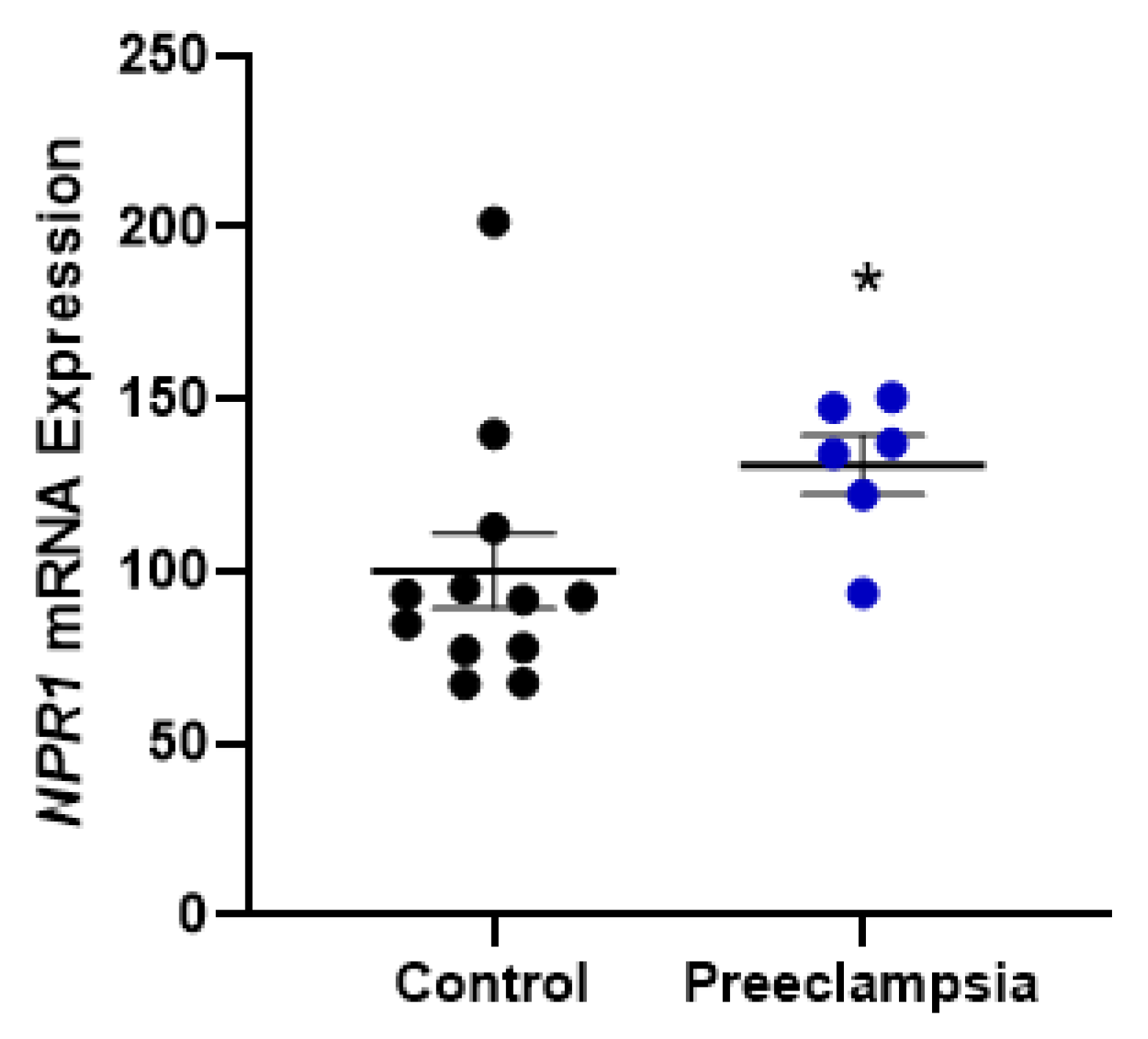
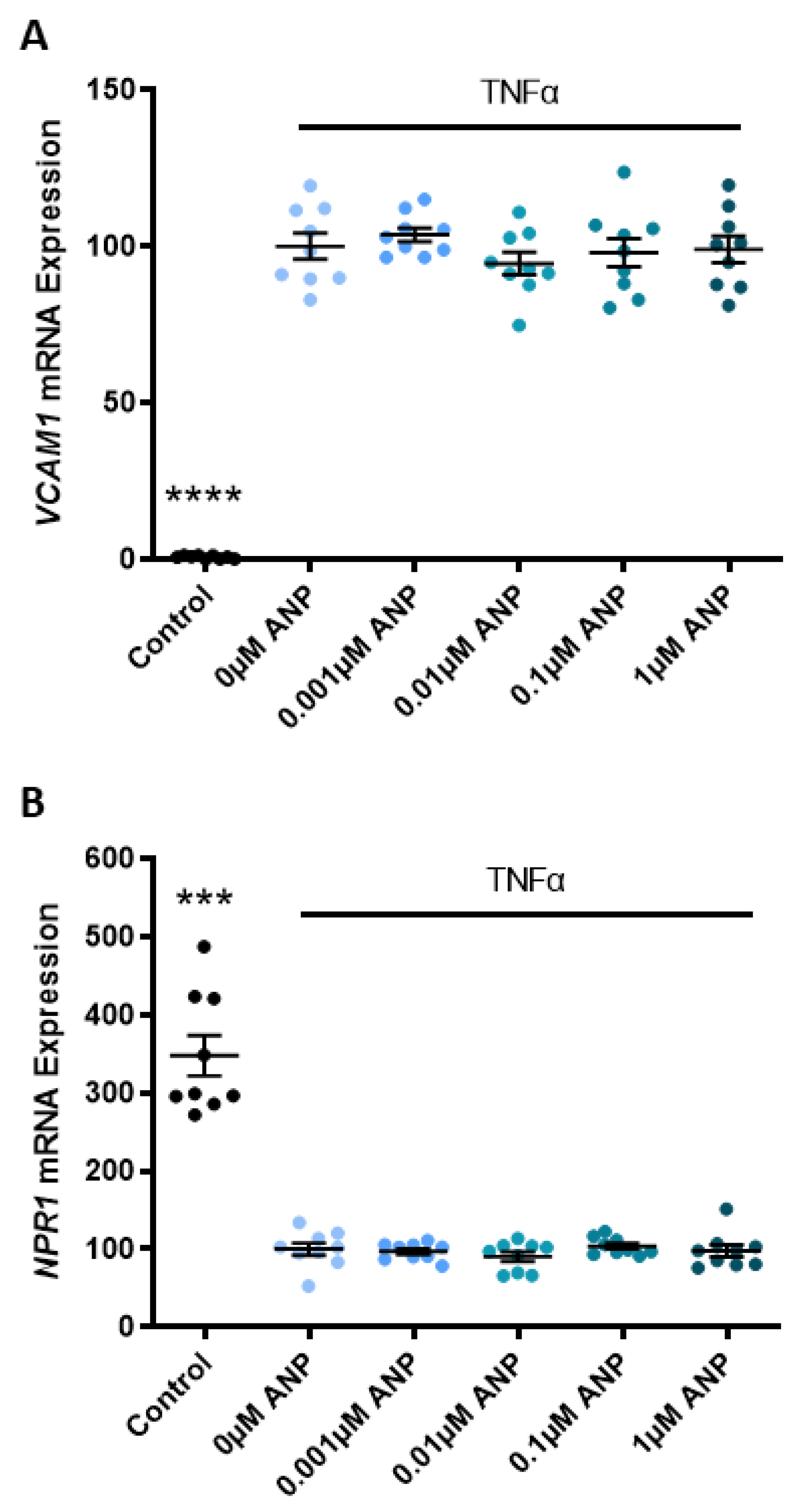
2.2. Omental Artery Expression of ANP Receptor, Natriuretic Peptide Receptor A (NPR1), Significantly Increased with Preeclampsia
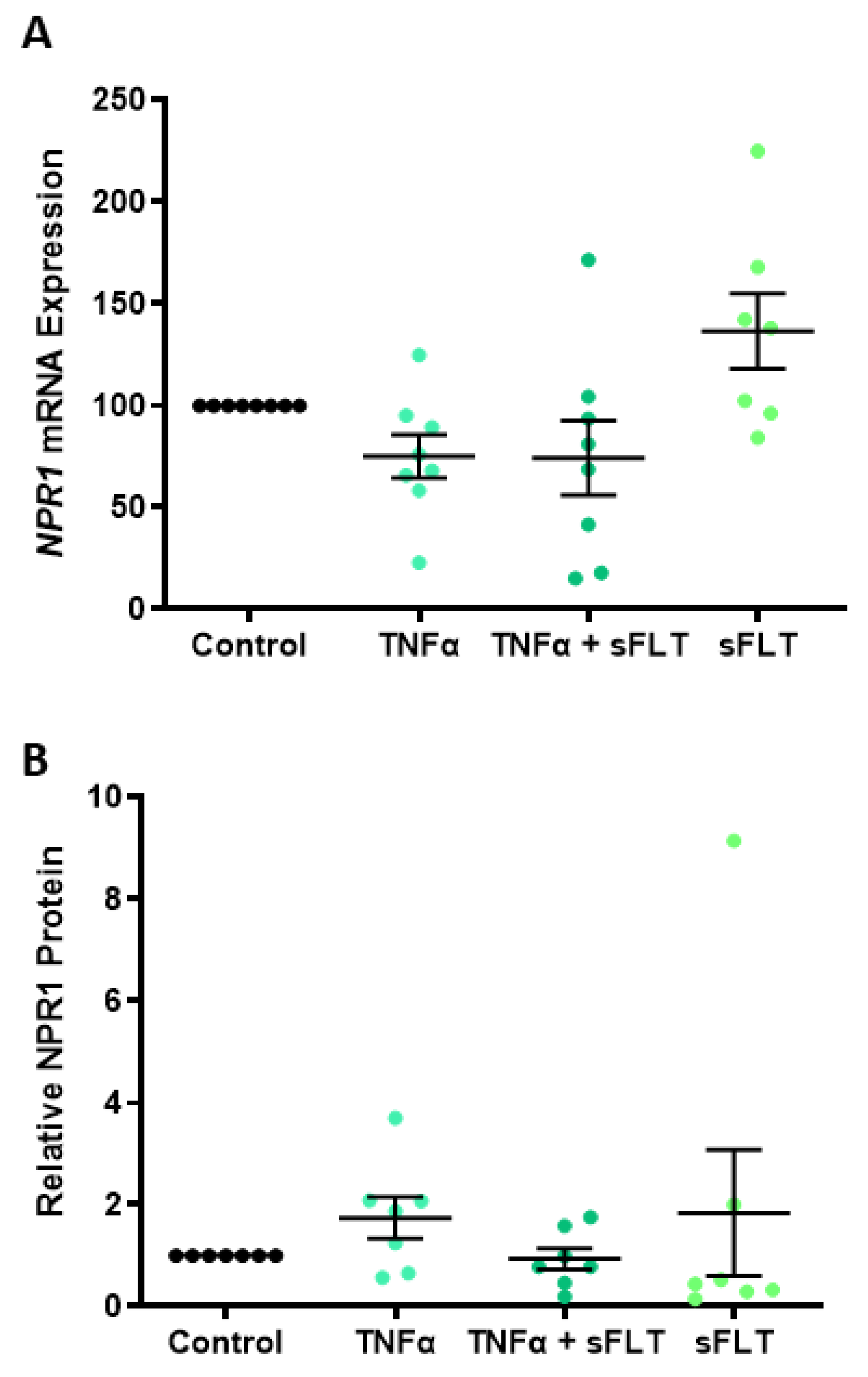
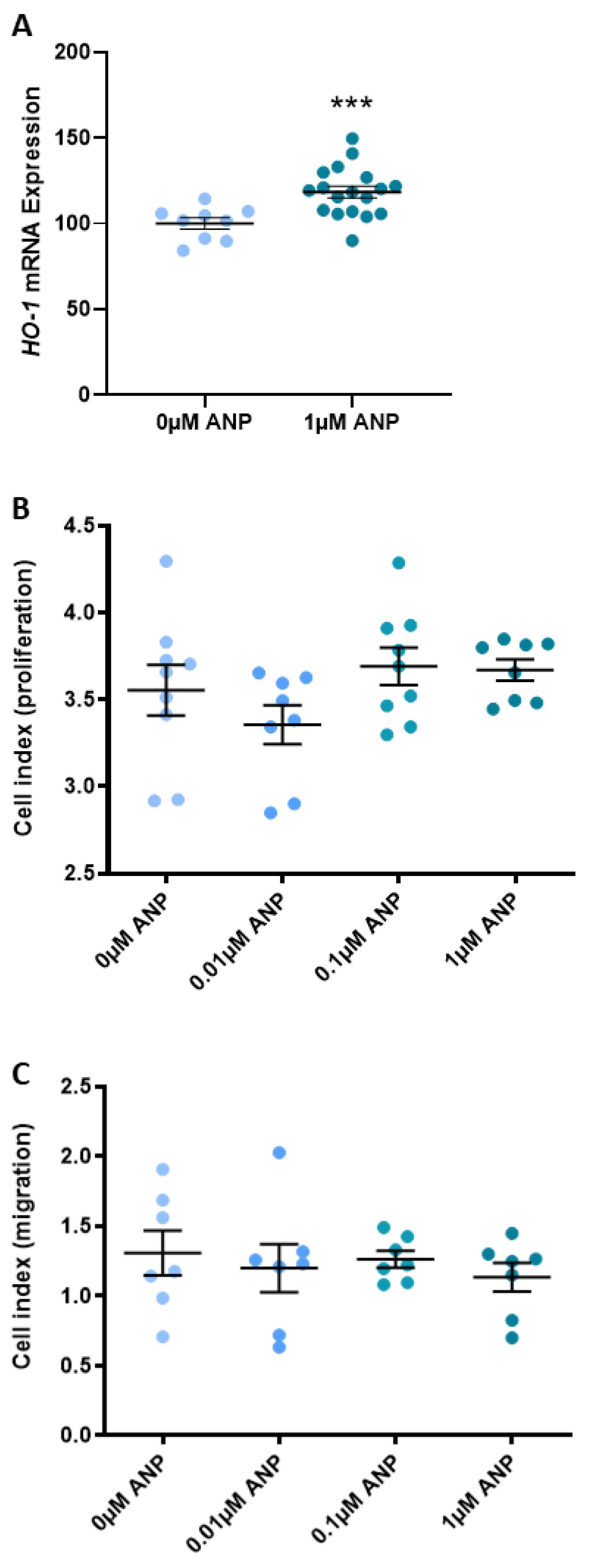
2.3. ANP Increases Heme Oxygenase (HO)-1 Expression but Does Not Affect Human Umbilical Vein Endothelial Cell (HUVEC) Proliferation or Migration
3. Discussion
3.1. Circulating Corin and Pro-ANP Are Elevated in Preeclamptic Pregnancies
3.2. Omental Artery Expression of the ANP Receptor, NPR1, Is Elevated in Preeclamptic Pregnancies
3.3. ANP Enhances Endothelial Production of HO-1 but Does Not Affect Proliferation, Migration, or Endothelial Dysfunction
4. Materials and Methods
4.1. Tissue Collection
4.2. Blood and Placenta Processing
4.3. Isolation and Treatment of Omental Arteries
4.4. Isolation and Treatment of Primary HUVECs
4.5. ELISA Analysis
4.6. Quantitative RT-PCR Analysis
4.7. Western Blot Analysis
4.8. xCELLigence
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wallis, A.B.; Saftlas, A.F.; Hsia, J.; Atrash, H.K. Secular trends in the rates of preeclampsia, eclampsia, and gestational hypertension, United States, 1987–2004. Am. J. Hypertens. 2008, 21, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Duley, L. The global impact of pre-eclampsia and eclampsia. Semin. Perinatol. 2009, 33, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Porreco, R.P.; Barkey, R. Peripartum intensive care. J. Matern. Fetal. Neonatal. Med. 2010, 23, 1136–1138. [Google Scholar] [CrossRef]
- Goldenberg, R.L.; Rouse, D.J. Prevention of premature birth. N. Engl. J. Med. 1998, 339, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Sircar, M.; Thadhani, R.; Karumanchi, S.A. Pathogenesis of preeclampsia. Curr. Opin. Nephrol. Hypertens. 2015, 24, 131–138. [Google Scholar] [CrossRef]
- El-Sayed, A.A.F. Preeclampsia: A review of the pathogenesis and possible management strategies based on its pathophysiological derangements. Taiwan. J. Obstet. Gynecol. 2017, 56, 593–598. [Google Scholar] [CrossRef]
- Irgens, H.U.; Reisaeter, L.; Irgens, L.M.; Lie, R.T. Long term mortality of mothers and fathers after pre-eclampsia: Population based cohort study. BMJ 2001, 323, 1213–1217. [Google Scholar] [CrossRef]
- Smith, G.C.; Pell, J.P.; Walsh, D. Pregnancy complications and maternal risk of ischaemic heart disease: A retrospective cohort study of 129,290 births. Lancet 2001, 357, 2002–2006. [Google Scholar] [CrossRef]
- Ray, J.G.; Vermeulen, M.J.; Schull, M.J.; Redelmeier, D.A. Cardiovascular health after maternal placental syndromes (CHAMPS): Population-based retrospective cohort study. Lancet 2005, 366, 1797–1803. [Google Scholar] [CrossRef]
- Wilson, B.J.; Watson, M.S.; Prescott, G.J.; Sunderland, S.; Campbell, D.M.; Hannaford, P.; Smith, W.C. Hypertensive diseases of pregnancy and risk of hypertension and stroke in later life: Results from cohort study. BMJ 2003, 326, 845. [Google Scholar] [CrossRef]
- Wikström, A.K.; Haglund, B.; Olovsson, M.; Lindeberg, S.N. The risk of maternal ischaemic heart disease after gestational hypertensive disease. BJOG Int. J. Obstet. Gynaecol. 2005, 112, 1486–1491. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, L.; Casas, J.P.; Hingorani, A.D.; Williams, D.J. Pre-eclampsia and risk of cardiovascular disease and cancer in later life: Systematic review and meta-analysis. BMJ 2007, 335, 974. [Google Scholar] [CrossRef] [PubMed]
- Hermes, W.; Van Kesteren, F.; De Groot, C.J. Preeclampsia and cardiovascular risk. Minerva Ginecol. 2012, 64, 281–292. [Google Scholar] [PubMed]
- Harskamp, R.E.; Zeeman, G.G. Preeclampsia: At risk for remote cardiovascular disease. Am. J. Med. Sci. 2007, 334, 291–295. [Google Scholar] [CrossRef]
- Degrelle, S.A.; Chissey, A.; Stepanian, A.; Fournier, T.; Guibourdenche, J.; Mandelbrot, L.; Tsatsaris, V. Placental Overexpression of Soluble CORIN in Preeclampsia. Am. J. Pathol. 2020, 190, 970–976. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, Q. Role of corin and atrial natriuretic peptide in preeclampsia. Placenta 2013, 34, 89–94. [Google Scholar] [CrossRef]
- Dong, N.; Niu, Y.; Chen, Y.; Sun, S.; Wu, Q. Function and regulation of corin in physiology and disease. Biochem. Soc. Trans. 2020, 48, 1905–1916. [Google Scholar] [CrossRef]
- Wu, Q. The serine protease corin in cardiovascular biology and disease. Front. Biosci. J. Virtual Libr. 2007, 12, 4179–4190. [Google Scholar] [CrossRef]
- Yu, R.; Han, X.; Zhang, X.; Wang, Y.; Wang, T. Circulating soluble corin as a potential biomarker for cardiovascular diseases: A translational review. Clin. Chim. Acta Int. J. Clin. Chem. 2018, 485, 106–112. [Google Scholar] [CrossRef]
- Wu, P.; Haththotuwa, R.; Kwok, C.S.; Babu, A.; Kotronias, R.A.; Rushton, C.; Zaman, A.; Fryer, A.A.; Kadam, U.; Chew-Graham, C.A.; et al. Preeclampsia and Future Cardiovascular Health: A Systematic Review and Meta-Analysis. Circ. Cardiovasc. Qual. Outcomes 2017, 10, e003497. [Google Scholar] [CrossRef]
- Yan, W.; Sheng, N.; Seto, M.; Morser, J.; Wu, Q. Corin, a mosaic transmembrane serine protease encoded by a novel cDNA from human heart. J. Biol. Chem. 1999, 274, 14926–14935. [Google Scholar] [CrossRef] [PubMed]
- Potter, L.R.; Yoder, A.R.; Flora, D.R.; Antos, L.K.; Dickey, D.M. Natriuretic peptides: Their structures, receptors, physiologic functions and therapeutic applications. Handb. Exp. Pharmacol. 2009, 191, 341–366. [Google Scholar] [CrossRef]
- Armaly, Z.; Assady, S.; Abassi, Z. Corin: A new player in the regulation of salt-water balance and blood pressure. Curr. Opin. Nephrol. Hypertens. 2013, 22, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.J.; Jardine, A.G. Atrial natriuretic peptide: Water and electrolyte homeostasis. Bailliere’s Clin. Endocrinol. Metab. 1989, 3, 431–450. [Google Scholar] [CrossRef] [PubMed]
- Goetze, J.P.; Bruneau, B.G.; Ramos, H.R.; Ogawa, T.; de Bold, M.K.; de Bold, A.J. Cardiac natriuretic peptides. Nat. Rev. Cardiol. 2020, 17, 698–717. [Google Scholar] [CrossRef]
- Dong, N.; Zhou, T.; Zhang, Y.; Liu, M.; Li, H.; Huang, X.; Liu, Z.; Wu, Y.; Fukuda, K.; Qin, J.; et al. Corin mutations K317E and S472G from preeclamptic patients alter zymogen activation and cell surface targeting. J. Biol. Chem. 2014, 289, 17909–17916. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, W.; Dong, N.; Lou, J.; Srinivasan, D.K.; Cheng, W.; Huang, X.; Liu, M.; Fang, C.; Peng, J.; et al. Role of corin in trophoblast invasion and uterine spiral artery remodelling in pregnancy. Nature 2012, 484, 246–250. [Google Scholar] [CrossRef]
- Liu, Y.; Hu, J.; Yu, Q.; Zhang, P.; Han, X.; Peng, H. Increased serum soluble corin in mid pregnancy is associated with hypertensive disorders of pregnancy. J. Women’s Health 2015, 24, 572–577. [Google Scholar] [CrossRef]
- Zaki, M.A.; El-Banawy Sel, D.; El-Gammal, H.H. Plasma soluble corin and N-terminal pro-atrial natriuretic peptide levels in pregnancy induced hypertension. Pregnancy Hypertens. 2012, 2, 48–52. [Google Scholar] [CrossRef]
- Miyazaki, J.; Nishizawa, H.; Kambayashi, A.; Ito, M.; Noda, Y.; Terasawa, S.; Kato, T.; Miyamura, H.; Shiogama, K.; Sekiya, T.; et al. Increased levels of soluble corin in pre-eclampsia and fetal growth restriction. Placenta 2016, 48, 20–25. [Google Scholar] [CrossRef]
- Kaitu’u-Lino, T.J.; Ye, L.; Tuohey, L.; Dimitriadis, E.; Bulmer, J.; Rogers, P.; Menkhorst, E.; Van Sinderen, M.; Girling, J.E.; Hannan, N.; et al. Corin, an enzyme with a putative role in spiral artery remodeling, is up-regulated in late secretory endometrium and first trimester decidua. Hum. Reprod. 2013, 28, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, S.; Lou, J.; Li, H.; Liu, M.; Dong, N.; Wu, Q. Atrial natriuretic peptide promotes uterine decidualization and a TRAIL-dependent mechanism in spiral artery remodeling. J. Clin. Investig. 2021, 131, e151053. [Google Scholar] [CrossRef] [PubMed]
- Staff, A.C.; Fjeldstad, H.E.; Fosheim, I.K.; Moe, K.; Turowski, G.; Johnsen, G.M.; Alnaes-Katjavivi, P.; Sugulle, M. Failure of physiological transformation and spiral artery atherosis: Their roles in preeclampsia. Am. J. Obstet. Gynecol. 2022, 226, S895–S906. [Google Scholar] [CrossRef]
- Baird, R.C.; Li, S.; Wang, H.; Naga Prasad, S.V.; Majdalany, D.; Perni, U.; Wu, Q. Pregnancy-Associated Cardiac Hypertrophy in Corin-Deficient Mice: Observations in a Transgenic Model of Preeclampsia. Can. J. Cardiol. 2019, 35, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Abassi, Z.; Kinaneh, S.; Skarzinski, G.; Cinnamon, E.; Smith, Y.; Bursztyn, M.; Ariel, I. Aberrant corin and PCSK6 in placentas of the maternal hyperinsulinemia IUGR rat model. Pregnancy Hypertens. 2020, 21, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gu, X.; Zhang, Y.; Dong, N.; Wu, Q. Corin: A Key Mediator in Sodium Homeostasis, Vascular Remodeling, and Heart Failure. Biology 2022, 11, 717. [Google Scholar] [CrossRef]
- Gu, Y.; Thompson, D.; Xu, J.; Lewis, D.F.; Morgan, J.A.; Cooper, D.B.; McCathran, C.E.; Wang, Y. Aberrant pro-atrial natriuretic peptide/corin/natriuretic peptide receptor signaling is present in maternal vascular endothelium in preeclampsia. Pregnancy Hypertens. 2018, 11, 1–6. [Google Scholar] [CrossRef]
- Veerbeek, J.H.; Hermes, W.; Breimer, A.Y.; van Rijn, B.B.; Koenen, S.V.; Mol, B.W.; Franx, A.; de Groot, C.J.; Koster, M.P. Cardiovascular disease risk factors after early-onset preeclampsia, late-onset preeclampsia, and pregnancy-induced hypertension. Hypertension 2015, 65, 600–606. [Google Scholar] [CrossRef]
- Fraser, R.; Whitley, G.S.; Johnstone, A.P.; Host, A.J.; Sebire, N.J.; Thilaganathan, B.; Cartwright, J.E. Impaired decidual natural killer cell regulation of vascular remodelling in early human pregnancies with high uterine artery resistance. J. Pathol. 2012, 228, 322–332. [Google Scholar] [CrossRef]
- Sunderland, N.S.; Thomson, S.E.; Heffernan, S.J.; Lim, S.; Thompson, J.; Ogle, R.; McKenzie, P.; Kirwan, P.J.; Makris, A.; Hennessy, A. Tumor necrosis factor alpha induces a model of preeclampsia in pregnant baboons (Papio hamadryas). Cytokine 2011, 56, 192–199. [Google Scholar] [CrossRef]
- Cindrova-Davies, T.; Sanders, D.A.; Burton, G.J.; Charnock-Jones, D.S. Soluble FLT1 sensitizes endothelial cells to inflammatory cytokines by antagonizing VEGF receptor-mediated signalling. Cardiovasc. Res. 2011, 89, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, Z.; He, M.; Zhou, T.; Niu, Y.; Sun, S.; Li, H.; Zhang, C.; Zhang, S.; Liu, M.; et al. Krüppel-like factor 17 upregulates uterine corin expression and promotes spiral artery remodeling in pregnancy. Proc. Natl. Acad. Sci. USA 2020, 117, 19425–19434. [Google Scholar] [CrossRef]
- Pouta, A.M.; Vuolteenaho, O.J.; Laatikainen, T.J. An increase of the plasma N-terminal peptide of proatrial natriuretic peptide in preeclampsia. Obstet. Gynecol. 1997, 89, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Boroń, D.; Kornacki, J.; Gutaj, P.; Mantaj, U.; Wirstlein, P.; Wender-Ozegowska, E. Corin-The Early Marker of Preeclampsia in Pregestational Diabetes Mellitus. J. Clin. Med. 2022, 12, 61. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Larson, M.G.; Keyes, M.J.; Levy, D.; Benjamin, E.J.; Vasan, R.S. Association of plasma natriuretic peptide levels with metabolic risk factors in ambulatory individuals. Circulation 2007, 115, 1345–1353. [Google Scholar] [CrossRef]
- Knappe, S.; Wu, F.; Madlansacay, M.R.; Wu, Q. Identification of domain structures in the propeptide of corin essential for the processing of proatrial natriuretic peptide. J. Biol. Chem. 2004, 279, 34464–34471. [Google Scholar] [CrossRef]
- Senöz, S.; Sahin, N.; Ozcan, T.; Direm, B.; Gökmen, O. The concentration of plasma atrial natriuretic peptide in normotensive and preeclamptic pregnancies. Eur. J. Obstet. Gynecol. Reprod. Biol. 1995, 62, 173–177. [Google Scholar] [CrossRef]
- Adam, B.; Malatyalioğlu, E.; Alvur, M.; Kökçü, A.; Bedir, A. Plasma Atrial Natriuretic Peptide Levels in Preeclampsia and Eclampsia. J. Matern.-Fetal Investig. 1998, 8, 85–88. [Google Scholar]
- Irons, D.W.; Baylis, P.H.; Butler, T.J.; Davison, J.M. Atrial natriuretic peptide in preeclampsia: Metabolic clearance, sodium excretion and renal hemodynamics. Am. J. Physiol. 1997, 273, F483–F487. [Google Scholar] [CrossRef]
- Hatjis, C.G.; Grogan, D.M. Changes in placental atrial natriuretic peptide receptors associated with severe toxemia of pregnancy. Placenta 1989, 10, 153–159. [Google Scholar] [CrossRef]
- Minegishi, T.; Nakamura, M.; Abe, K.; Tano, M.; Andoh, A.; Yoshida, M.; Takagi, T.; Nishikimi, T.; Kojima, M.; Kangawa, K. Adrenomedullin and atrial natriuretic peptide concentrations in normal pregnancy and pre-eclampsia. Mol. Hum. Reprod. 1999, 5, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Sala, C.; Campise, M.; Ambroso, G.; Motta, T.; Zanchetti, A.; Morganti, A. Atrial natriuretic peptide and hemodynamic changes during normal human pregnancy. Hypertension 1995, 25, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, J.K.; Fogh-Andersen, N.; Jaszczak, P.; Giese, J. Atrial natriuretic peptide (ANP) decrease during normal pregnancy as related to hemodynamic changes and volume regulation. Acta Obstet. Gynecol. Scand. 1993, 72, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Ellmers, L.J.; Knowles, J.W.; Kim, H.S.; Smithies, O.; Maeda, N.; Cameron, V.A. Ventricular expression of natriuretic peptides in Npr1(-/-) mice with cardiac hypertrophy and fibrosis. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H707–H714. [Google Scholar] [CrossRef]
- Khurana, M.L.; Mani, I.; Kumar, P.; Ramasamy, C.; Pandey, K.N. Ligand-Dependent Downregulation of Guanylyl Cyclase/Natriuretic Peptide Receptor-A: Role of miR-128 and miR-195. Int. J. Mol. Sci. 2022, 23, 13381. [Google Scholar] [CrossRef] [PubMed]
- Sunamura, M.; Duda, D.G.; Ghattas, M.H.; Lozonschi, L.; Motoi, F.; Yamauchi, J.; Matsuno, S.; Shibahara, S.; Abraham, N.G. Heme oxygenase-1 accelerates tumor angiogenesis of human pancreatic cancer. Angiogenesis 2003, 6, 15–24. [Google Scholar] [CrossRef]
- Was, H.; Cichon, T.; Smolarczyk, R.; Rudnicka, D.; Stopa, M.; Chevalier, C.; Leger, J.J.; Lackowska, B.; Grochot, A.; Bojkowska, K.; et al. Overexpression of heme oxygenase-1 in murine melanoma: Increased proliferation and viability of tumor cells, decreased survival of mice. Am. J. Pathol. 2006, 169, 2181–2198. [Google Scholar] [CrossRef]
- Evazi Bakhshi, S.; Mohammadi Roushandeh, A.; Habibi Roudkenar, M.; Shekarchi, S.; Bahadori, M.H. CRISPR/Cas9-mediated knockout of HO-1 decreased the proliferation and migration of T47D cells and increased cisplatin-induced apoptosis: An In Vitro study. Med. Oncol. 2022, 39, 175. [Google Scholar] [CrossRef]
- Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin, Number 222. Obstet. Gynecol. 2020, 135, e237–e260. [CrossRef]
- Burton, G.J.; Sebire, N.J.; Myatt, L.; Tannetta, D.; Wang, Y.L.; Sadovsky, Y.; Staff, A.C.; Redman, C.W. Optimising sample collection for placental research. Placenta 2014, 35, 9–22. [Google Scholar] [CrossRef]
- Brownfoot, F.C.; Hannan, N.; Onda, K.; Tong, S.; Kaitu’u-Lino, T. Soluble endoglin production is upregulated by oxysterols but not quenched by pravastatin in primary placental and endothelial cells. Placenta 2014, 35, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Kiemer, A.K.; Bildner, N.; Weber, N.C.; Vollmar, A.M. Characterization of heme oxygenase 1 (heat shock protein 32) induction by atrial natriuretic peptide in human endothelial cells. Endocrinology 2003, 144, 802–812. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 22) | Preeclampsia (n = 29) | |
|---|---|---|
| Maternal Age Median (IQR) | 31 (28–33) | 31 (28–35) |
| Gestation at Sampling Median (IQR) | 28.5 (27.4–30.3) | 29.6 (28.4–31.4) |
| Gestation at Delivery Median (IQR) | 39.6 (38.9–40.7) | 30.6 (28.3–31.7) **** |
| BMI (kg/m2) Median (IQR) | 23.5 (21.8–29.0) | 32.0 (25.0–37.3) *** |
| Parity no. (%) | ||
| 0 | 10 (45) | 22 (76) |
| 1 | 11 (50) | 4 (14) |
| ≥2 | 1 (5) | 3 (10) |
| Highest SBP prior to delivery (mmHg) Median (IQR) | 130 (125–131) | 174 (170–180) **** |
| Highest DBP prior to delivery (mmHg) Median (IQR) | 76 (70–81.3) | 103 (100–110) **** |
| Birth weight (g) Median (IQR) | 3550 (3158–3700) | 1305 (918–1497) **** |
| Control (n = 10) | Preeclampsia (n = 28) | |
|---|---|---|
| Maternal Age Median (IQR) | 29 (24–36) | 31 (27–34) |
| Gestation at Delivery Median (IQR) | 29.9 (27.4–32) | 30.4 (27.6–31.8) |
| BMI (kg/m2) Median (IQR) | 28.4 (24.0–40.9) | 27.0 (24.5–37.0) |
| Parity no. (%) | ||
| 0 | 2 (20) | 18 (64) |
| 1 | 6 (60) | 7 (25) |
| ≥2 | 2 (20) | 3 (11) |
| Highest SBP prior to delivery (mmHg) Median (IQR) | 125 (110–130) | 175 (160–180) **** |
| Highest DBP prior to delivery (mmHg) Median (IQR) | 70 (64–76) | 100 (90–110) **** |
| Birth weight (g) Median (IQR) | 1454 (941–2011) | 1314 (811–1424) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binder, N.K.; Beard, S.; de Alwis, N.; Fato, B.R.; Nguyen, T.-V.; Kaitu’u-Lino, T.J.; Hannan, N.J. Investigating the Effects of Atrial Natriuretic Peptide on the Maternal Endothelium to Determine Potential Implications for Preeclampsia. Int. J. Mol. Sci. 2023, 24, 6182. https://doi.org/10.3390/ijms24076182
Binder NK, Beard S, de Alwis N, Fato BR, Nguyen T-V, Kaitu’u-Lino TJ, Hannan NJ. Investigating the Effects of Atrial Natriuretic Peptide on the Maternal Endothelium to Determine Potential Implications for Preeclampsia. International Journal of Molecular Sciences. 2023; 24(7):6182. https://doi.org/10.3390/ijms24076182
Chicago/Turabian StyleBinder, Natalie K., Sally Beard, Natasha de Alwis, Bianca R. Fato, Tuong-Vi Nguyen, Tu’uhevaha J. Kaitu’u-Lino, and Natalie J. Hannan. 2023. "Investigating the Effects of Atrial Natriuretic Peptide on the Maternal Endothelium to Determine Potential Implications for Preeclampsia" International Journal of Molecular Sciences 24, no. 7: 6182. https://doi.org/10.3390/ijms24076182
APA StyleBinder, N. K., Beard, S., de Alwis, N., Fato, B. R., Nguyen, T.-V., Kaitu’u-Lino, T. J., & Hannan, N. J. (2023). Investigating the Effects of Atrial Natriuretic Peptide on the Maternal Endothelium to Determine Potential Implications for Preeclampsia. International Journal of Molecular Sciences, 24(7), 6182. https://doi.org/10.3390/ijms24076182