
Intra-Airway Treatment with Synthetic Lipoxin A4 and Resolvin E2 Mitigates Neonatal Asthma Triggered by Maternal Exposure to Environmental Particles

{kind=link}
{kind=link}
Abstract
1. Introduction
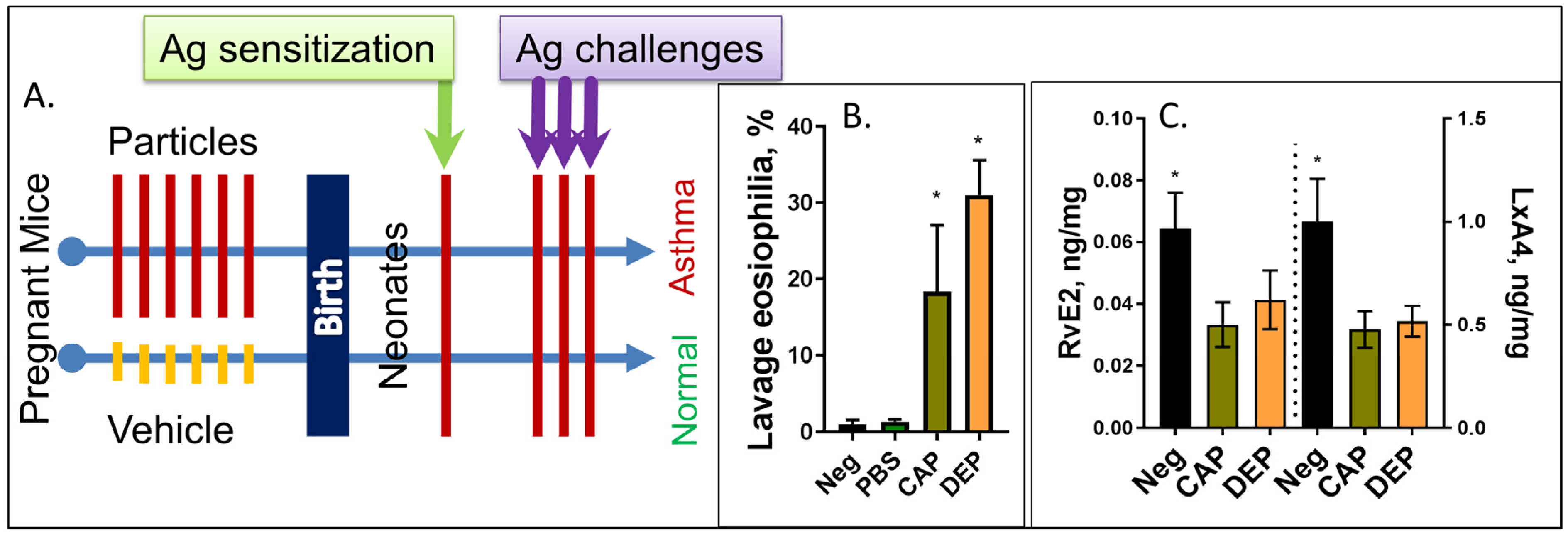
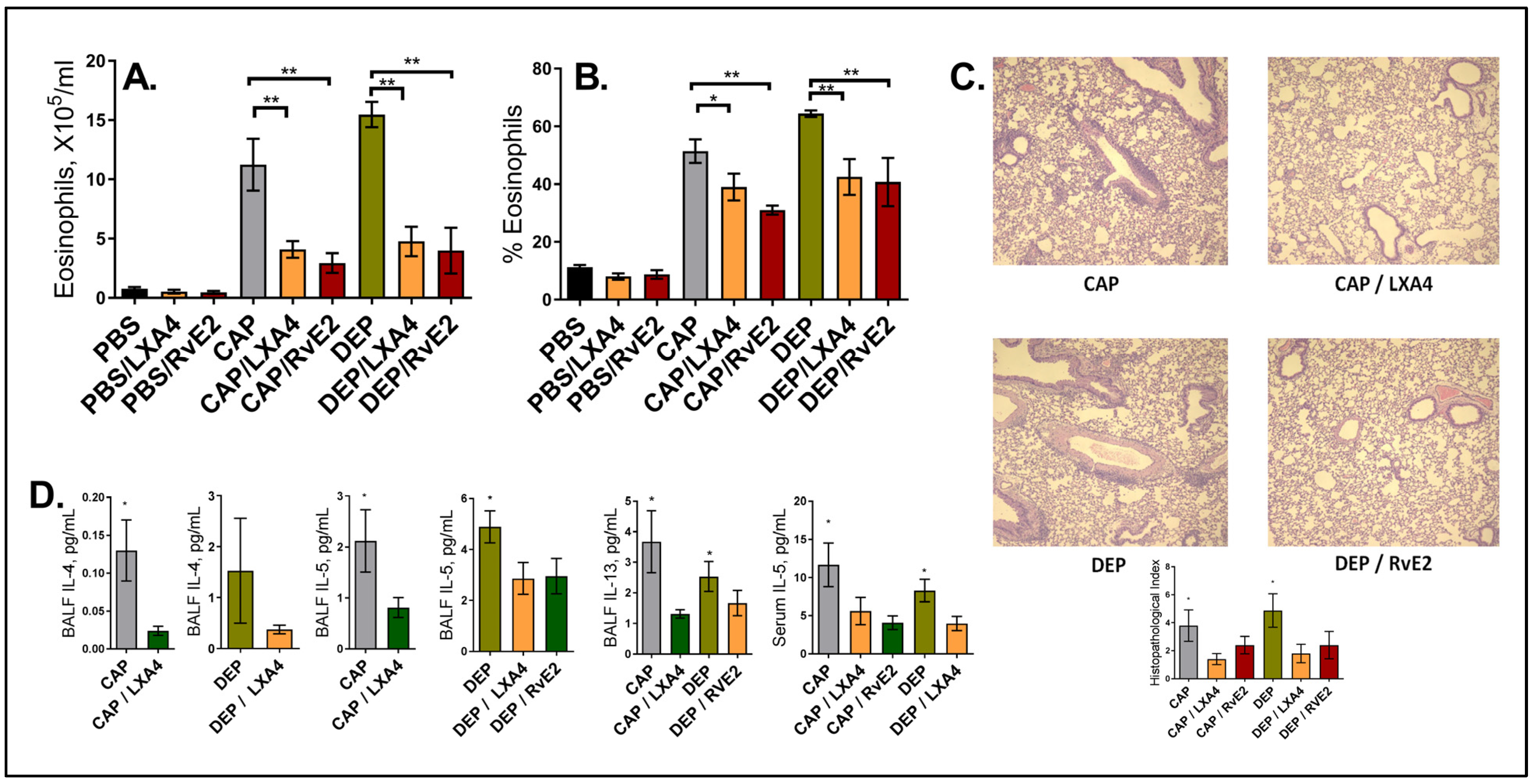
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Particles
4.3. Exposure
4.4. Pathologic Analysis
4.5. SPMs
4.6. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, D.; Chen, Z.; Zhou, L.F.; Huang, S.X. Air pollutants and early origins of respiratory diseases. Chronic Dis. Transl. Med. 2018, 4, 75–94. [Google Scholar] [CrossRef] [PubMed]
- Brunekreef, B.; Beelen, R.; Hoek, G.; Schouten, L.; Bausch-Goldbohm, S.; Fischer, P.; Armstrong, B.; Hughes, E.; Jerrett, M.; van den Brandt, P. Effects of long-term exposure to traffic-related air pollution on respiratory and cardiovascular mortality in The Netherlands: The NLCS-AIR study. Res. Rep. Health Eff. Inst. 2009, 139, 5–71; discussion 73–89. [Google Scholar]
- Bouazza, N.; Foissac, F.; Urien, S.; Guedj, R.; Carbajal, R.; Tréluyer, J.M.; Chappuy, H. Fine particulate pollution and asthma exacerbations. Arch. Dis. Child. 2018, 103, 828–831. [Google Scholar] [CrossRef]
- Host, S.; Chatignoux, E.; Leal, C.; Grémy, I. Exposition à la pollution atmosphérique de proximité liée au trafic: Quelles méthodes pour quels risques sanitaires? [Health risk assessment of traffic-related air pollution near busy roads]. Rev. Epidemiol. Sante Publique 2012, 60, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Sompornrattanaphan, M.; Thongngarm, T.; Ratanawatkul, P.; Wongsa, C.; Swigris, J.J. The contribution of particulate matter to respiratory allergy. Asian Pac. J. Allergy Immunol. 2020, 38, 19–28. [Google Scholar] [CrossRef]
- Hussain, S.; Parker, S.; Edwards, K.; Finch, J.; Jeanjean, A.; Leigh, R.; Gonem, S. Effects of indoor particulate matter exposure on daily asthma control. Ann. Allergy Asthma Immunol. 2019, 123, 375–380.e3. [Google Scholar] [CrossRef]
- Guarnieri, M.; Balmes, J.R. Outdoor air pollution and asthma. Lancet 2014, 383, 1581–1592. [Google Scholar] [CrossRef]
- Kiser, D.; Metcalf, W.J.; Elhanan, G.; Schnieder, B.; Schlauch, K.; Joros, A.; Petersen, C.; Grzymski, J. Particulate matter and emergency visits for asthma: A time-series study of their association in the presence and absence of wildfire smoke in Reno, Nevada, 2013–2018. Environ. Health 2020, 19, 92. [Google Scholar] [CrossRef]
- Goldsmith, C.A.; Kobzik, L. Particulate air pollution and asthma: A review of epidemiological and biological studies. Rev. Environ. Health 1999, 14, 121–134. [Google Scholar] [CrossRef]
- Venn, A.J.; Lewis, S.A.; Cooper, M.; Hubbard, R.; Britton, J. Living near a main road and the risk of wheezing illness in children. Am. J. Respir. Crit. Care Med. 2001, 164, 2177–2180. [Google Scholar] [CrossRef]
- Khreis, H.; Kelly, C.; Tate, J.; Parslow, R.; Lucas, K.; Nieuwenhuijsen, M. Exposure to traffic-related air pollution and risk of development of childhood asthma: A systematic review and meta-analysis. Environ. Int. 2017, 100, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Beamer, P.I. Air Pollution Contributes to Asthma Deaths. Am. J. Respir. Crit. Care Med. 2019, 200, 1–2. [Google Scholar] [CrossRef]
- Alexis, N.E.; Huang, Y.C.; Rappold, A.G.; Kehrl, H.; Devlin, R.; Peden, D.B. Patients with asthma demonstrate airway inflammation after exposure to concentrated ambient particulate matter. Am. J. Respir. Crit. Care Med. 2014, 190, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Oliver, B.G.; Pant, A.; Olivera, A.; Poronnik, P.; Pollock, C.A.; Saad, S. Effects of air pollution on human health—Mechanistic evidence suggested by in vitro and in vivo modelling. Environ. Res. 2022, 212 Pt C, 113378. [Google Scholar] [CrossRef]
- Lee, Y.G.; Lee, P.H.; Choi, S.M.; An, M.H.; Jang, A.S. Effects of Air Pollutants on Airway Diseases. Int. J. Environ. Res. Public Health 2021, 18, 9905. [Google Scholar] [CrossRef] [PubMed]
- Harkema, J.R.; Keeler, G.; Wagner, J.; Morishita, M.; Timm, E.; Hotchkiss, J.; Marsik, F.; Dvonch, T.; Kaminski, N.; Barr, E. Effects of concentrated ambient particles on normal and hypersecretory airways in rats. Res. Rep. Health Eff. Inst. 2004, 120, 1–68. [Google Scholar]
- Suzuki, T.; Kanoh, T.; Ishimori, M.; Ikeda, S.; Ohkuni, H. Adjuvant activity of diesel exhaust particulates (DEP) in production of anti-IgE and anti-IgG1 antibodies to mite allergen in mice. J. Clin. Lab. Immunol. 1996, 48, 187–199. [Google Scholar]
- Boehmer, T.K.; Foster, S.L.; Henry, J.R.; Woghiren-Akinnifesi, E.L.; Yip, F.Y.; Centers for Disease Control and Prevention. Residential proximity to major highways—United States, 2010. MMWR Suppl. 2013, 62, 46–50. [Google Scholar] [PubMed]
- Jung, C.R.; Chen, W.T.; Tang, Y.H.; Hwang, B.F. Fine particulate matter exposure during pregnancy and infancy and incident asthma. J. Allergy Clin. Immunol. 2019, 143, 2254–2262.e5. [Google Scholar] [CrossRef]
- Hehua, Z.; Qing, C.; Shanyan, G.; Qijun, W.; Yuhong, Z. The impact of prenatal exposure to air pollution on childhood wheezing and asthma: A systematic review. Environ. Res. 2017, 159, 519–530. [Google Scholar] [CrossRef]
- Gheissari, R.; Liao, J.; Garcia, E.; Pavlovic, N.; Gilliland, F.D.; Xiang, A.H.; Chen, Z. Health Outcomes in Children Associated with Prenatal and Early-Life Exposures to Air Pollution: A Narrative Review. Toxics 2022, 10, 458. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.W.; Hsieh, H.M.; Lee, C.H.; Lin, Y.C.; Tsao, Y.H.; Wu, H.W.; Kuo, F.C.; Hung, C.H. Prenatal and Postnatal Exposure to Ambient Air Pollution and Preschool Asthma in Neonatal Jaundice Infants. J. Inflamm. Res. 2022, 15, 3771–3781. [Google Scholar] [CrossRef] [PubMed]
- Ghazi, T.; Naidoo, P.; Naidoo, R.N.; Chuturgoon, A.A. Prenatal Air Pollution Exposure and Placental DNA Methylation Changes: Implications on Fetal Development and Future Disease Susceptibility. Cells 2021, 10, 3025. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Norbäck, D.; Li, Y.; Deng, Q. Early-life exposure to air pollution and childhood allergic diseases: An update on the link and its implications. Expert Rev. Clin. Immunol. 2020, 16, 813–827. [Google Scholar] [CrossRef]
- Ha, S.; Yeung, E.; Bell, E.; Insaf, T.; Ghassabian, A.; Bell, G.; Muscatiello, N.; Mendola, P. Prenatal and early life exposures to ambient air pollution and development. Environ. Res. 2019, 174, 170–175. [Google Scholar] [CrossRef]
- Fedulov, A.V.; Leme, A.; Yang, Z.; Dahl, M.; Lim, R.; Mariani, T.J.; Kobzik, L. Pulmonary exposure to particles during pregnancy causes increased neonatal asthma susceptibility. Am. J. Respir. Cell Mol. Biol. 2008, 38, 57–67. [Google Scholar] [CrossRef]
- Gregory, D.J.; Kobzik, L.; Yang, Z.; McGuire, C.C.; Fedulov, A.V. Transgenerational transmission of asthma risk after exposure to environmental particles during pregnancy. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 313, L395–L405. [Google Scholar] [CrossRef]
- Fedulov, A.V.; Kobzik, L. Allergy risk is mediated by dendritic cells with congenital epigenetic changes. Am. J. Respir. Cell Mol. Biol. 2011, 44, 285–292. [Google Scholar] [CrossRef]
- Fedulov, A.; Silverman, E.; Xiang, Y.; Leme, A.; Kobzik, L. Immunostimulatory CpG oligonucleotides abrogate allergic susceptibility in a murine model of maternal asthma transmission. J. Immunol. 2005, 175, 4292–4300. [Google Scholar] [CrossRef]
- Fedulov, A.V.; Leme, A.S.; Kobzik, L. Duration of allergic susceptibility in maternal transmission of asthma risk. Am. J. Reprod. Immunol. 2007, 58, 120–128. [Google Scholar] [CrossRef]
- Mikhaylova, L.; Zhang, Y.; Kobzik, L.; Fedulov, A.V. Link between epigenomic alterations and genome-wide aberrant transcriptional response to allergen in dendritic cells conveying maternal asthma risk. PLoS ONE 2013, 8, e70387. [Google Scholar] [CrossRef] [PubMed]
- Tiotiu, A.I.; Novakova, P.; Nedeva, D.; Chong-Neto, H.J.; Novakova, S.; Steiropoulos, P.; Kowal, K. Impact of Air Pollution on Asthma Outcomes. Int. J. Environ. Res. Public Health 2020, 17, 6212. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Dalli, J. The resolution code of acute inflammation: Novel pro-resolving lipid mediators in resolution. Semin. Immunol. 2015, 27, 200–215. [Google Scholar] [CrossRef]
- Barnig, C.; Levy, B.D. Innate immunity is a key factor for the resolution of inflammation in asthma. Eur. Respir. Rev. 2015, 24, 141–153. [Google Scholar] [CrossRef]
- Pirault, J.; Bäck, M. Lipoxin and Resolvin Receptors Transducing the Resolution of Inflammation in Cardiovascular Disease. Front. Pharmacol. 2018, 9, 1273. [Google Scholar] [CrossRef] [PubMed]
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef]
- Barnig, C.; Frossard, N.; Levy, B.D. Towards targeting resolution pathways of airway inflammation in asthma. Pharmacol. Ther. 2018, 186, 98–113. [Google Scholar] [CrossRef]
- Duvall, M.G.; Bruggemann, T.R.; Levy, B.D. Bronchoprotective mechanisms for specialized pro-resolving mediators in the resolution of lung inflammation. Mol Aspects Med. 2017, 58, 44–56. [Google Scholar] [CrossRef]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef]
- Planaguma, A.; Levy, B.D. Uncontrolled airway inflammation in lung disease represents a defect in counter-regulatory signaling. Future Lipidol. 2008, 3, 697–704. [Google Scholar] [CrossRef]
- Lu, X.; Fu, H.; Han, F.; Fang, Y.; Xu, J.; Zhang, L.; Du, Q. Lipoxin A4 regulates PM2.5-induced severe allergic asthma in mice via the Th1/Th2 balance of group 2 innate lymphoid cells. J. Thorac. Dis. 2018, 10, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Beck-Speier, I.; Karg, E.; Behrendt, H.; Stoeger, T.; Alessandrini, F. Ultrafine particles affect the balance of endogenous pro- and anti-inflammatory lipid mediators in the lung: In-vitro and in-vivo studies. Part. Fibre Toxicol. 2012, 9, 27. [Google Scholar] [CrossRef]
- Kumar, M.; Yano, N.; Fedulov, A.V. Gestational exposure to titanium dioxide, diesel exhaust, and concentrated urban air particles affects levels of specialized pro-resolving mediators in response to allergen in asthma-susceptible neonate lungs. J. Toxicol. Environ. Health A 2022, 85, 243–261. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.H.; Kobzik, L.; Dahl, M. Risk for asthma in offspring of asthmatic mothers versus fathers: A metaanalysis. PLoS ONE 2010, 5, e10134. [Google Scholar] [CrossRef]
- Barrett, E.G. Maternal influence in the transmission of asthma susceptibility. Pulm. Pharmacol. Ther. 2008, 21, 474–484. [Google Scholar] [CrossRef]
- Murdzoska, J.; Devadason, S.G.; Khoo, S.K.; Landau, L.I.; Young, S.; Goldblatt, J.; Zhang, G.; Le Souef, P.N.; Hayden, C.M. In utero smoke exposure and role of maternal and infant glutathione s-transferase genes on airway responsiveness and lung function in infancy. Am. J. Respir. Crit. Care Med. 2010, 181, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Breton, C.V.; Byun, H.M.; Wenten, M.; Pan, F.; Yang, A.; Gilliland, F.D. Prenatal tobacco smoke exposure affects global and gene-specific DNA methylation. Am. J. Respir. Crit. Care Med. 2009, 180, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Shankardass, K.; McConnell, R.; Jerrett, M.; Milam, J.; Richardson, J.; Berhane, K. Parental stress increases the effect of traffic-related air pollution on childhood asthma incidence. Proc. Natl. Acad. Sci. USA 2009, 106, 12406–12411. [Google Scholar] [CrossRef]
- Blacquiere, M.J.; Timens, W.; Melgert, B.N.; Geerlings, M.; Postma, D.S.; Hylkema, M.N. Maternal smoking during pregnancy induces airway remodelling in mice offspring. Eur. Respir. J. 2009, 33, 1133–1140. [Google Scholar] [CrossRef]
- Keil, T.; Lau, S.; Roll, S.; Gruber, C.; Nickel, R.; Niggemann, B.; Wahn, U.; Willich, S.N.; Kulig, M. Maternal smoking increases risk of allergic sensitization and wheezing only in children with allergic predisposition: Longitudinal analysis from birth to 10 years. Allergy 2009, 64, 445–451. [Google Scholar] [CrossRef]
- Wang, L.; Pinkerton, K.E. Air pollutant effects on fetal and early postnatal development. Birth Defects Res. C Embryo Today 2007, 81, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.L.; Clifton, V. Asthma and pregnancy: Emerging evidence of epigenetic interactions in utero. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 417–426. [Google Scholar] [CrossRef]
- Barker, D.J. The origins of the developmental origins theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Hamada, K.; Suzaki, Y.; Goldman, A.; Ning, Y.Y.; Goldsmith, C.; Palecanda, A.; Coull, B.; Hubeau, C.; Kobzik, L. Allergen-independent maternal transmission of asthma susceptibility. J. Immunol. 2003, 170, 1683–1689. [Google Scholar] [CrossRef]
- Ghio, A.J.; Kim, C.; Devlin, R.B. Concentrated ambient air particles induce mild pulmonary inflammation in healthy human volunteers. Am. J. Respir. Crit. Care Med. 2000, 162 Pt 1, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Kelly, F.J.; Fussell, J.C. Air pollution and public health: Emerging hazards and improved understanding of risk. Environ. Geochem. Health 2015, 37, 631–649. [Google Scholar] [CrossRef]
- Watson, A.Y.; Bates, R.R.; Kennedy, D. Air Pollution, the Automobile, and Public Health. In Biological Disposition of Airborne Particles: Basic Principles and Application to Vehicular Emissions; Schlesinger, R.B., Ed.; Academies Press: Washington, DC, USA, 1988. Available online: https://www.ncbi.nlm.nih.gov/books/NBK218161/ (accessed on 21 April 2020).
- Jakubzick, C.; Tacke, F.; Llodra, J.; van Rooijen, N.; Randolph, G.J. Modulation of dendritic cell trafficking to and from the airways. J. Immunol. 2006, 176, 3578–3584. [Google Scholar] [CrossRef] [PubMed]
- Imrich, A.; Ning, Y.; Lawrence, J.; Coull, B.; Gitin, E.; Knutson, M.; Kobzik, L. Alveolar macrophage cytokine response to air pollution particles: Oxidant mechanisms. Toxicol. Appl. Pharmacol. 2007, 218, 256–264. [Google Scholar] [CrossRef]
- Godleski, J.J.; Clarke, R.W.; Coull, B.A.; Saldiva, P.H.; Jiang, N.F.; Lawrence, J.; Koutrakis, P. Composition of inhaled urban air particles determines acute pulmonary responses. Ann. Occup. Hyg. 2002, 46, 419–424. [Google Scholar]
- Mauad, T.; Rivero, D.H.; de Oliveira, R.C.; Lichtenfels, A.J.; Guimaraes, E.T.; de Andre, P.A.; Kasahara, D.I.; Bueno, H.M.; Saldiva, P.H. Chronic exposure to ambient levels of urban particles affects mouse lung development. Am. J. Respir. Crit. Care Med. 2008, 178, 721–728. [Google Scholar] [CrossRef]
- Bommel, H.; Li-Weber, M.; Serfling, E.; Duschl, A. The environmental pollutant pyrene induces the production of IL-4. J. Allergy Clin. Immunol. 2000, 105, 796–802. [Google Scholar] [CrossRef] [PubMed]
- McConnell, R.; Berhane, K.; Yao, L.; Jerrett, M.; Lurmann, F.; Gilliland, F.; Kunzli, N.; Gauderman, J.; Avol, E.; Thomas, D.; et al. Traffic, susceptibility, and childhood asthma. Environ. Health Perspect. 2006, 114, 766–772. [Google Scholar] [CrossRef]
- New Scientist and Press Association. Diesel Fumes Lead to Thousands More Deaths Than Thought. 15 May 2017. Available online: https://www.newscientist.com/article/2131067-diesel-fumes-lead-to-thousands-more-deaths-than-thought/ (accessed on 14 February 2023).
- Grange, S.K.; Farren, N.J.; Vaughan, A.R.; Davison, J.; Carslaw, D.C. Post-Dieselgate: Evidence of NOx Emission Reductions Using On-Road Remote Sensing. Environ. Sci. Technol. Lett. 2020, 7, 382–387. [Google Scholar] [CrossRef]
- Long, E.; Carlsten, C. Controlled human exposure to diesel exhaust: Results illuminate health effects of traffic-related air pollution and inform future directions. Part. Fibre Toxicol. 2022, 19, 11. [Google Scholar] [CrossRef]
- Edwards, M.R.; Saglani, S.; Schwarze, J.; Skevaki, C.; Smith, J.A.; Ainsworth, B.; Almond, M.; Andreakos, E.; Belvisi, M.G.; Chung, K.F.; et al. Addressing unmet needs in understanding asthma mechanisms: From the European Asthma Research and Innovation Partnership (EARIP) Work Package (WP)2 collaborators. Eur. Respir. J. 2017, 49, 1602448. [Google Scholar] [CrossRef] [PubMed]
- Haworth, O.; Levy, B.D. Lipoxins, resolvins and protectins: New leads for the treatment of asthma. Expert Opin. Drug Discov. 2008, 3, 1209–1222. [Google Scholar] [CrossRef] [PubMed]
- Hisada, T.; Aoki-Saito, H.; Koga, Y. Are specialized pro-resolving mediators promising therapeutic agents for severe bronchial asthma? J. Thorac. Dis. 2017, 9, 4266–4269. [Google Scholar] [CrossRef]
- Hsiao, H.M.; Sapinoro, R.E.; Thatcher, T.H.; Croasdell, A.; Levy, E.P.; Fulton, R.A.; Olsen, K.C.; Pollock, S.J.; Serhan, C.N.; Phipps, R.P.; et al. A novel anti-inflammatory and pro-resolving role for resolvin D1 in acute cigarette smoke-induced lung inflammation. PLoS ONE 2013, 8, e58258. [Google Scholar] [CrossRef]
- Karra, L.; Haworth, O.; Priluck, R.; Levy, B.D.; Levi-Schaffer, F. Lipoxin B₄ promotes the resolution of allergic inflammation in the upper and lower airways of mice. Mucosal Immunol. 2015, 8, 852–862. [Google Scholar] [CrossRef]
- Dalli, J.; Winkler, J.W.; Colas, R.A.; Arnardottir, H.; Cheng, C.Y.; Chiang, N.; Petasis, N.A.; Serhan, C.N. Resolvin D3 and aspirin-triggered resolvin D3 are potent immunoresolvents. Chem. Biol. 2013, 20, 188–201. [Google Scholar] [CrossRef]
- Cezar, T.L.C.; Martinez, R.M.; Rocha, C.D.; Melo, C.P.B.; Vale, D.L.; Borghi, S.M.; Fattori, V.; Vignoli, J.A.; Camilios-Neto, D.; Baracat, M.M.; et al. Treatment with maresin 1, a docosahexaenoic acid-derived pro-resolution lipid, protects skin from inflammation and oxidative stress caused by UVB irradiation. Sci. Rep. 2019, 9, 3062. [Google Scholar] [CrossRef] [PubMed]
- Levy, B.D.; Kohli, P.; Gotlinger, K.; Haworth, O.; Hong, S.; Kazani, S.; Israel, E.; Haley, K.J.; Serhan, C.N. Protectin D1 is generated in asthma and dampens airway inflammation and hyperresponsiveness. J. Immunol. 2007, 178, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Van Hove, C.L.; Maes, T.; Joos, G.F.; Tournoy, K.G. Chronic inflammation in asthma: A contest of persistence vs resolution. Allergy 2008, 63, 1095–1109. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef]
- Serhan, C.N.; Krishnamoorthy, S.; Recchiuti, A.; Chiang, N. Novel anti-inflammatory--pro-resolving mediators and their receptors. Curr. Top. Med. Chem. 2011, 11, 629–647. [Google Scholar] [CrossRef]
- Chiang, N.; Serhan, C.N. Structural elucidation and physiologic functions of specialized pro-resolving mediators and their receptors. Mol. Aspects Med. 2017, 58, 114–129. [Google Scholar] [CrossRef]
- Oh, S.F.; Dona, M.; Fredman, G.; Krishnamoorthy, S.; Irimia, D.; Serhan, C.N. Resolvin E2 formation and impact in inflammation resolution. J. Immunol. 2012, 188, 4527–4534. [Google Scholar] [CrossRef]
- Liu, Y.; Wei, L.; He, C.; Chen, R.; Meng, L. Lipoxin A4 inhibits ovalbumin-induced airway inflammation and airway remodeling in a mouse model of asthma. Chem. Biol. Interact. 2021, 349, 109660. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Haworth, O.; Croze, R.; Oh, S.F.; Uddin, M.; Carlo, T.; Pfeffer, M.A.; Priluck, R.; Serhan, C.N.; Levy, B.D. Resolvin D1 and aspirin-triggered resolvin D1 promote resolution of allergic airways responses. J. Immunol. 2012, 189, 1983–1991. [Google Scholar] [CrossRef]
- Flesher, R.P.; Herbert, C.; Kumar, R.K. Resolvin E1 promotes resolution of inflammation in a mouse model of an acute exacerbation of allergic asthma. Clin. Sci. 2014, 126, 805–814. [Google Scholar] [CrossRef]
- Aoki, H.; Hisada, T.; Ishizuka, T.; Utsugi, M.; Kawata, T.; Shimizu, Y.; Okajima, F.; Dobashi, K.; Mori, M. Resolvin E1 dampens airway inflammation and hyperresponsiveness in a murine model of asthma. Biochem. Biophys. Res. Commun. 2008, 367, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Demokritou, P.; Gupta, T.; Ferguson, S.; Koutrakis, P. Development of a high-volume concentrated ambient particles system (CAPS) for human and animal inhalation toxicological studies. Inhal. Toxicol. 2003, 15, 111–129. [Google Scholar] [CrossRef]
- Lawrence, J.; Wolfson, J.M.; Ferguson, S.; Koutrakis, P.; Godleski, J. Performance Stability of the Harvard Ambient Particle Concentrator. Aerosol Sci. Technol. 2004, 38, 219–227. [Google Scholar] [CrossRef]
- Savage, S.T.; Lawrence, J.; Katz, T.; Stearns, R.C.; Coull, B.A.; Godleski, J.J. Does the Harvard/U.S. Environmental Protection Agency Ambient Particle Concentrator change the toxic potential of particles? J. Air Waste Manag. Assoc. 2003, 53, 1088–1097. [Google Scholar] [CrossRef] [PubMed]
- Koutrakis, P.; Godleski, J.J.; Coull, B.; Lawrence, J. Identifying the Physical and Chemical Properties of Particulate Matter Responsible for the Observed Adverse Health Effects. EPA Grant Number: R827353C014 Report Summary. Available online: https://cfpub.epa.gov/ncer_abstracts/index.cfm/fuseaction/display.abstractDetail/abstract/8578 (accessed on 28 February 2023).
- Mandarino, A.; Gregory, D.J.; McGuire, C.C.; Leblanc, B.W.; Witt, H.; Rivera, L.M.; Godleski, J.J.; Fedulov, A.V. The effect of talc particles on phagocytes in co-culture with ovarian cancer cells. Environ. Res. 2020, 180, 108676. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramar, M.; Yano, N.; Fedulov, A.V. Intra-Airway Treatment with Synthetic Lipoxin A4 and Resolvin E2 Mitigates Neonatal Asthma Triggered by Maternal Exposure to Environmental Particles. Int. J. Mol. Sci. 2023, 24, 6145. https://doi.org/10.3390/ijms24076145
Ramar M, Yano N, Fedulov AV. Intra-Airway Treatment with Synthetic Lipoxin A4 and Resolvin E2 Mitigates Neonatal Asthma Triggered by Maternal Exposure to Environmental Particles. International Journal of Molecular Sciences. 2023; 24(7):6145. https://doi.org/10.3390/ijms24076145
Chicago/Turabian StyleRamar, Mohankumar, Naohiro Yano, and Alexey V. Fedulov. 2023. "Intra-Airway Treatment with Synthetic Lipoxin A4 and Resolvin E2 Mitigates Neonatal Asthma Triggered by Maternal Exposure to Environmental Particles" International Journal of Molecular Sciences 24, no. 7: 6145. https://doi.org/10.3390/ijms24076145
APA StyleRamar, M., Yano, N., & Fedulov, A. V. (2023). Intra-Airway Treatment with Synthetic Lipoxin A4 and Resolvin E2 Mitigates Neonatal Asthma Triggered by Maternal Exposure to Environmental Particles. International Journal of Molecular Sciences, 24(7), 6145. https://doi.org/10.3390/ijms24076145