
Antiviral Effect of Ginsenosides rk1 against Influenza a Virus Infection by Targeting the Hemagglutinin 1-Mediated Virus Attachment
 and
and
Abstract
1. Introduction
2. Results
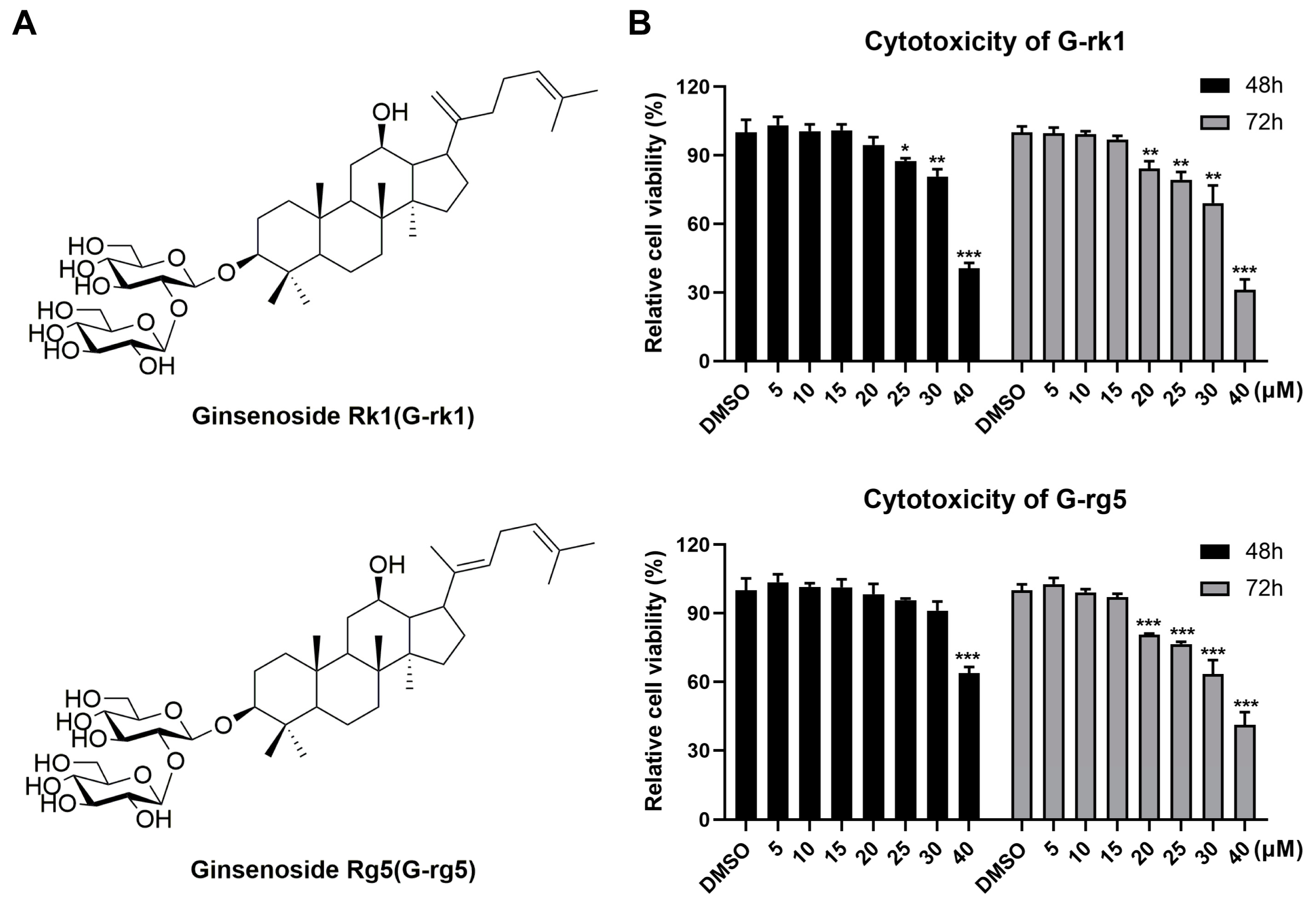
2.1. Cell Cytotoxicity and Inhibitory Effects of G-rk1 and G-rg5 against IAV Infection in A549 Cells
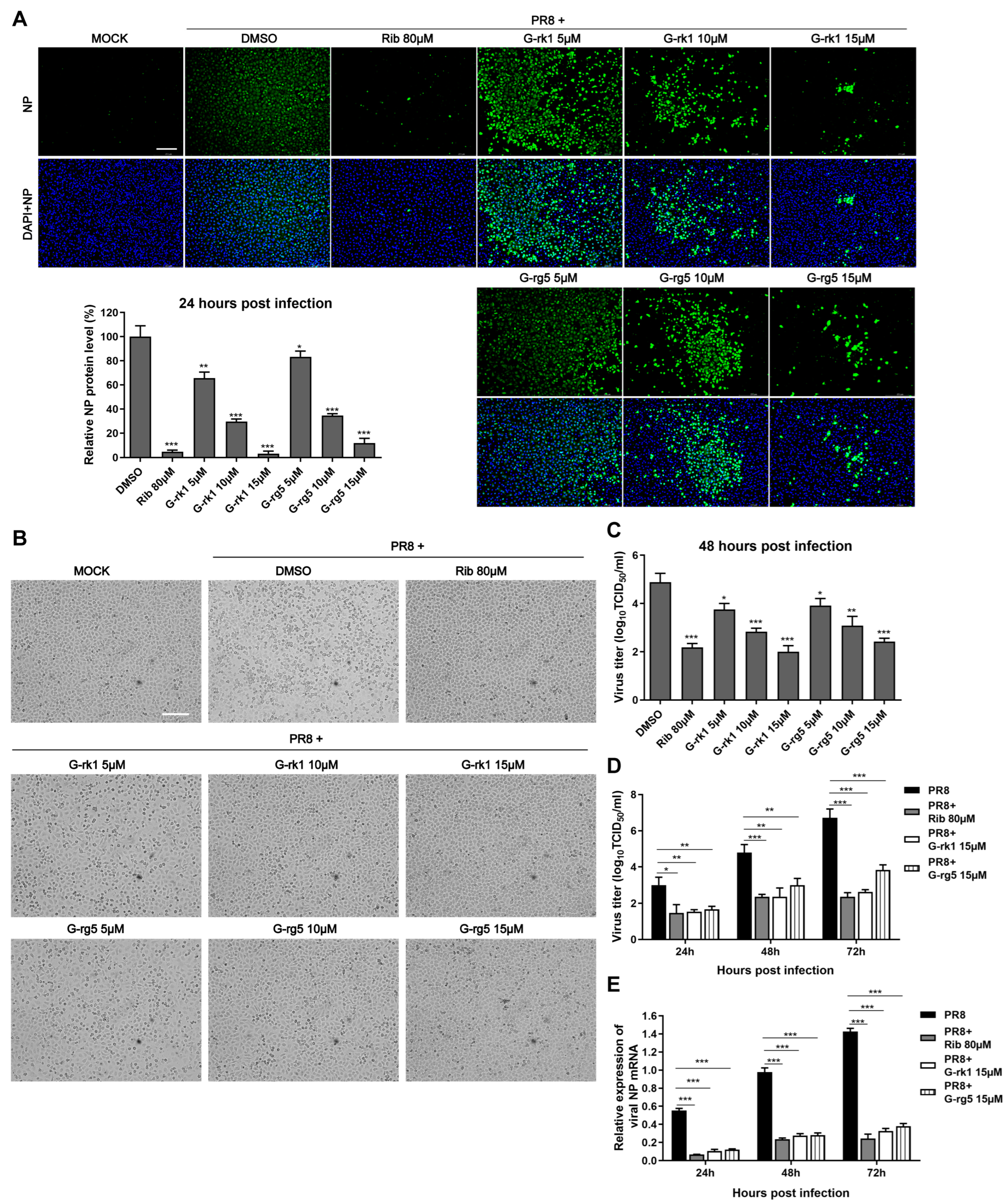
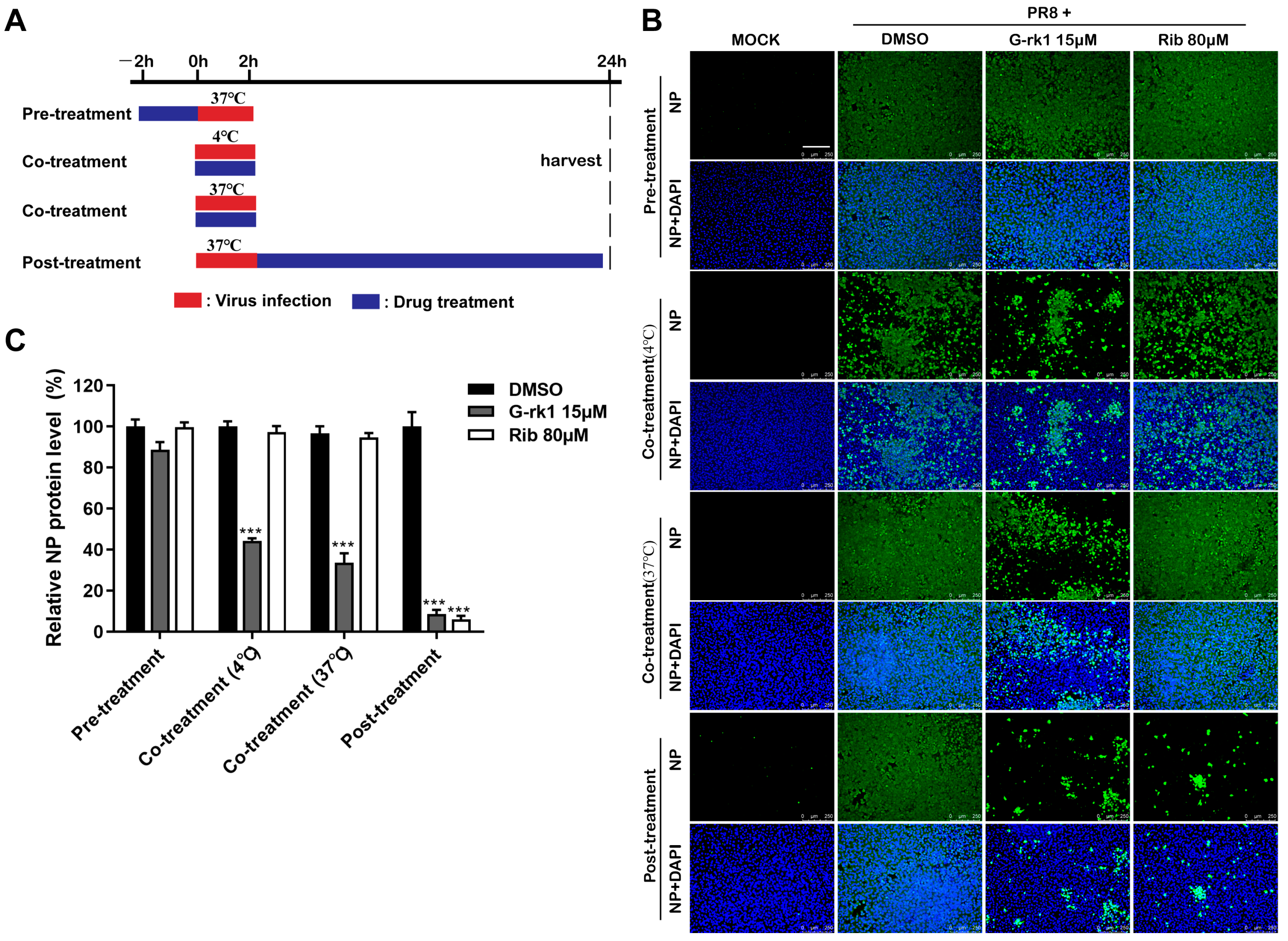
2.2. G-rk1 Interferes with IAV Entry
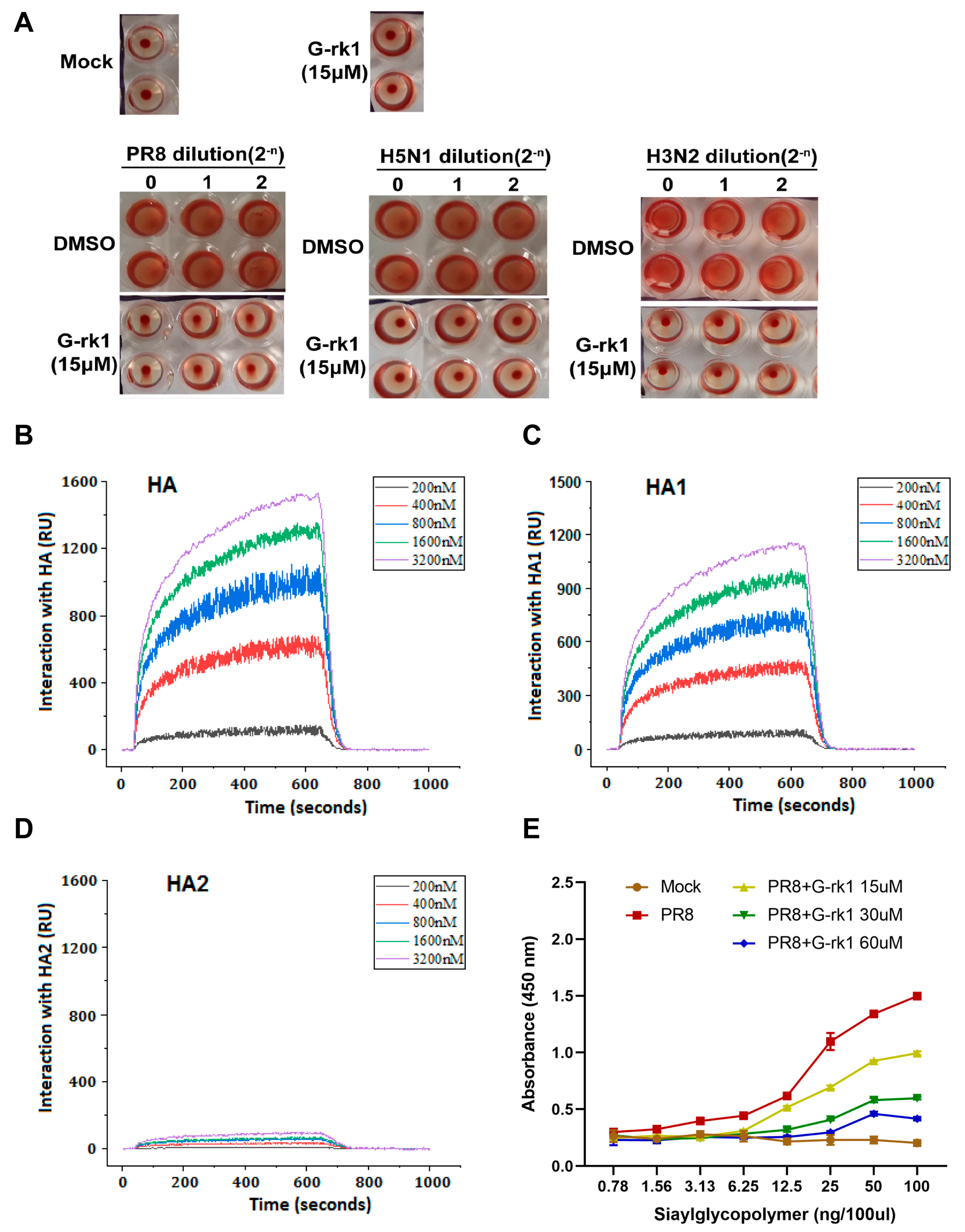
2.3. G-rk1 Interacts with HA via the HA1 Subunit
2.4. G-rk1 Reduces IAV-Induced Morbidity and Mortality following Lethal PR8 Infection in Mice
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Virus Strains
4.2. Compounds
4.3. Mice
4.4. Cytotoxicity Assay
4.5. Indirect Immunofluorescence Assay (IFA)
4.6. Viral Inhibition Assay
4.7. Real-Time Reverse-Transcription PCR (RT-PCR)
4.8. Time-Course Inhibition Assay
4.9. Hemagglutination Inhibition Assay (HAI)
4.10. Surface Plasmon Resonance (SPR) Analysis
4.11. The Influence of G-rk1 on Sialic Acid Receptor Binding Ability of IAV by an Indirect ELISA
4.12. In Vivo Viral Challenge and G-rk1 Treatment
4.13. Lung Tissue Viral Titers
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Basu, A.; Antanasijevic, A.; Wang, M.; Li, B.; Mills, D.; Ames, J.; Nash, P.; Williams, J.; Peet, N.; Moir, D.; et al. New small molecule entry inhibitors targeting hemagglutinin-mediated influenza a virus fusion. J. Virol. 2014, 88, 1447–1460. [Google Scholar] [CrossRef]
- Killingley, B.; Nguyen-Van-Tam, J. Routes of influenza transmission. Influenza Other Respir. Viruses 2013, 7 (Suppl. S2), 42–51. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Influenza (Seasonal). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 12 January 2023).
- Beigel, J.; Farrar, J.; Han, A.; Hayden, F.; Hyer, R.; de Jong, M.; Lochindarat, S.; Nguyen, T.; Nguyen, T.; Tran, T.; et al. Avian influenza A (H5N1) infection in humans. N. Engl. J. Med. 2005, 353, 1374–1385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Huang, Y.; Tian, D.; Lau, E.; Wan, Y.; Liu, X.; Dong, Y.; Song, Z.; Zhang, X.; Zhang, J.; et al. Kinetics of serological responses in influenza A(H7N9)-infected patients correlate with clinical outcome in China, 2013. Eurosurveillance 2013, 18, 20657. [Google Scholar] [CrossRef]
- Drexler, J.; Helmer, A.; Kirberg, H.; Reber, U.; Panning, M.; Müller, M.; Höfling, K.; Matz, B.; Drosten, C.; Eis-Hübinger, A. Poor clinical sensitivity of rapid antigen test for influenza A pandemic (H1N1) 2009 virus. Emerg. Infect. Dis. 2009, 15, 1662–1664. [Google Scholar] [CrossRef]
- De Clercq, E. Antiviral agents active against influenza A viruses. Nat. Rev. Drug Discov. 2006, 5, 1015–1025. [Google Scholar] [CrossRef]
- Heo, Y.A. Baloxavir: First Global Approval. Drugs 2018, 78, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Abdelwhab, E.; Veits, J.; Mettenleiter, T. Biological fitness and natural selection of amantadine resistant variants of avian influenza H5N1 viruses. Virus Res. 2017, 228, 109–113. [Google Scholar] [CrossRef]
- Ikematsu, H.; Hayden, F.; Kawaguchi, K.; Kinoshita, M.; de Jong, M.; Lee, N.; Takashima, S.; Noshi, T.; Tsuchiya, K.; Uehara, T. Baloxavir Marboxil for Prophylaxis against Influenza in Household Contacts. N. Engl. J. Med. 2020, 383, 309–320. [Google Scholar] [CrossRef]
- Lackenby, A.; Hungnes, O.; Dudman, S.; Meijer, A.; Paget, W.; Hay, A.; Zambon, M. Emergence of resistance to oseltamivir among influenza A(H1N1) viruses in Europe. Eurosurveillance 2008, 13, 8026. [Google Scholar] [CrossRef]
- de Jong, M.; Tran, T.; Truong, H.; Vo, M.; Smith, G.; Nguyen, V.; Bach, V.; Phan, T.; Do, Q.; Guan, Y.; et al. Oseltamivir resistance during treatment of influenza A (H5N1) infection. N. Engl. J. Med. 2005, 353, 2667–2672. [Google Scholar] [CrossRef] [PubMed]
- Ginex, T.; Luque, F. Searching for effective antiviral small molecules against influenza A virus: A patent review. Expert Opin. Ther. Pat. 2021, 31, 53–66. [Google Scholar] [CrossRef]
- Zeng, L.Y.; Yang, J.; Liu, S. Investigational hemagglutinin-targeted influenza virus inhibitors. Expert Opin. Investig. Drugs 2017, 26, 63–73. [Google Scholar] [CrossRef]
- Hussein, A.; Cheng, H.; Tundup, S.; Antanasijevic, A.; Varhegyi, E.; Perez, J.; AbdulRahman, E.; Elenany, M.; Helal, S.; Caffrey, M.; et al. Identification of entry inhibitors with 4-aminopiperidine scaffold targeting group 1 influenza A virus. Antivir. Res. 2020, 177, 104782. [Google Scholar] [CrossRef]
- Skehel, J.; Wiley, D. Receptor binding and membrane fusion in virus entry: The influenza hemagglutinin. Annu. Rev. Biochem. 2000, 69, 531–569. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.; Lee, G.; Park, S.; Bae, J.; Heo, J.; Kim, H.; Woo, S.; Lee, H.; Ahn, C.; et al. Novel Small Molecule Targeting the Hemagglutinin Stalk of Influenza Viruses. J. Virol. 2019, 93, e00878-19. [Google Scholar] [CrossRef] [PubMed]
- Nachbagauer, R.; Krammer, F. Universal influenza virus vaccines and therapeutic antibodies. Clin. Microbiol. Infect. 2017, 23, 222–228. [Google Scholar] [CrossRef]
- Gill, D.S.; Damle, N.K. Biopharmaceutical drug discovery using novel protein scaffolds. Curr. Opin. Biotechnol. 2006, 17, 653–658. [Google Scholar] [CrossRef]
- van Dongen, M.J.P.; Kadam, R.U.; Juraszek, J.; Lawson, E.; Brandenburg, B.; Schmitz, F.; Schepens, W.B.G.; Stoops, B.; van Diepen, H.A.; Jongeneelen, M.; et al. A small-molecule fusion inhibitor of influenza virus is orally active in mice. Science 2019, 363, eaar6221. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Lee, R.; Nam, S.; Kim, D.; Cho, I.; Kim, H.; Cho, Y.; Rhim, H.; Nah, S. Ginseng gintonin, aging societies, and geriatric brain diseases. Integr. Med. Res. 2021, 10, 100450. [Google Scholar] [CrossRef]
- Yang, W.; Hu, Y.; Wu, W.; Ye, M.; Guo, D. Saponins in the genus Panax L. (Araliaceae): A systematic review of their chemical diversity. Phytochemistry 2014, 106, 7–24. [Google Scholar] [CrossRef]
- Ratan, Z.; Haidere, M.; Hong, Y.; Park, S.; Lee, J.; Lee, J.; Cho, J. Pharmacological potential of ginseng and its major component ginsenosides. J. Ginseng Res. 2021, 45, 199–210. [Google Scholar] [CrossRef]
- Lee, J.; Ko, E.; Hwang, H.; Lee, Y.; Kwon, Y.; Kim, M.; Kang, S. Antiviral activity of ginseng extract against respiratory syncytial virus infection. Int. J. Mol. Med. 2014, 34, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Jung, Y.; Cho, Y. Beneficial effects of a combination of Korean red ginseng and highly active antiretroviral therapy in human immunodeficiency virus type 1-infected patients. Clin. Vaccine Immunol. 2009, 16, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Im, K.; Kim, J.; Min, H. Ginseng, the natural effectual antiviral: Protective effects of Korean Red Ginseng against viral infection. J. Ginseng Res. 2016, 40, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.; Altman, E. Inhibition of Herpes Simplex Viruses, Types 1 and 2, by Ginsenoside 20(S)-Rg3. J. Microbiol. Biotechnol. 2020, 30, 101–108. [Google Scholar] [CrossRef]
- Kang, L.; Choi, Y.; Lee, S. Stimulation of TRAF6/TAK1 degradation and inhibition of JNK/AP-1 signalling by ginsenoside Rg3 attenuates hepatitis B virus replication. Int. J. Biochem. Cell Biol. 2013, 45, 2612–2621. [Google Scholar] [CrossRef]
- Wang, Y.; Jung, Y.; Kim, K.; Kwon, Y.; Kim, Y.; Zhang, Z.; Kang, H.; Wang, B.; Quan, F.; Kang, S. Antiviral Activity of Fermented Ginseng Extracts against a Broad Range of Influenza Viruses. Viruses 2018, 10, 471. [Google Scholar] [CrossRef]
- Park, E.; Yum, J.; Ku, K.; Kim, H.; Kang, Y.; Kim, J.; Kim, J.; Kang, Y.; Seo, S. Red Ginseng-containing diet helps to protect mice and ferrets from the lethal infection by highly pathogenic H5N1 influenza virus. J. Ginseng Res. 2014, 38, 40–46. [Google Scholar] [CrossRef]
- Dong, W.; Farooqui, A.; Leon, A.J.; Kelvin, D.J. Inhibition of influenza A virus infection by ginsenosides. PLoS ONE 2017, 12, e0171936. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Kim, Y.J.; Jeon, J.N.; Wang, C.; Min, J.W.; Noh, H.Y.; Yang, D.C. Effect of white, red and black ginseng on physicochemical properties and ginsenosides. Plant Foods Hum. Nutr. 2015, 70, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Wong, G.; Liu, Y.; Liu, L.; Gao, G.; Shi, Y. Ribavirin is effective against drug-resistant H7N9 influenza virus infections. Protein Cell 2016, 7, 611–614. [Google Scholar] [CrossRef]
- Wu, N.C.; Wilson, I.A. Structural Biology of Influenza Hemagglutinin: An Amaranthine Adventure. Viruses 2020, 12, 1053. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, J.C. Hemagglutination-inhibition assay for influenza virus subtype identification and the detection and quantitation of serum antibodies to influenza virus. Methods Mol. Biol. 2014, 1161, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Edinger, T.; Pohl, M.; Stertz, S. Entry of influenza A virus: Host factors and antiviral targets. J. Gen. Virol. 2014, 95, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Luan, X.; Dou, D.; Huang, L. Comparative Analysis of Ginsenosides and Oligosaccharides in White Ginseng (WG), red Ginseng (RG) and Black Ginseng (BG). J. Chromatogr. Sci. 2019, 57, 403–410. [Google Scholar] [CrossRef]
- He, M.; Huang, X.; Liu, S.; Guo, C.; Xie, Y.; Meijer, A.H.; Wang, M. The Difference between White and Red Ginseng: Variations in Ginsenosides and Immunomodulation. Planta Med. 2018, 84, 845–854. [Google Scholar] [CrossRef]
- Han, M.J.; Kim, D.H. Effects of Red and Fermented Ginseng and Ginsenosides on Allergic Disorders. Biomolecules 2020, 10, 634. [Google Scholar] [CrossRef]
- Yoo, S.; Park, B.I.; Kim, D.H.; Lee, S.; Lee, S.H.; Shim, W.S.; Seo, Y.K.; Kang, K.; Lee, K.T.; Yim, S.V.; et al. Ginsenoside Absorption Rate and Extent Enhancement of Black Ginseng (CJ EnerG) over Red Ginseng in Healthy Adults. Pharmaceutics 2021, 13, 487. [Google Scholar] [CrossRef]
- Kim, E.; Kim, S.; Park, S.; Kim, S.; Yu, K.; Kim, S.; Lee, S.; Seo, Y.; Cho, N.; Kang, K.; et al. Greater Efficacy of Black Ginseng (CJ EnerG) over Red Ginseng against Lethal Influenza A Virus Infection. Nutrients 2019, 11, 1879. [Google Scholar] [CrossRef]
- De Clercq, E.; Li, G. Approved Antiviral Drugs over the Past 50 Years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Yang, G.; DeBeauchamp, J.; Crumpton, J.; Kim, H.; Li, L.; Wan, X.; Kercher, L.; Bowman, A.; Webster, R.; et al. HA stabilization promotes replication and transmission of swine H1N1 gamma influenza viruses in ferrets. eLife 2020, 9, e56236. [Google Scholar] [CrossRef] [PubMed]
- Wright, Z.; Wu, N.; Kadam, R.; Wilson, I.; Wolan, D. Structure-based optimization and synthesis of antiviral drug Arbidol analogues with significantly improved affinity to influenza hemagglutinin. Bioorganic Med. Chem. Lett. 2017, 27, 3744–3748. [Google Scholar] [CrossRef] [PubMed]
- Bereczki, I.; Kicsák, M.; Dobray, L.; Borbás, A.; Batta, G.; Kéki, S.; Nikodém, É.N.; Ostorházi, E.; Rozgonyi, F.; Vanderlinden, E.; et al. Semisynthetic teicoplanin derivatives as new influenza virus binding inhibitors: Synthesis and antiviral studies. Bioorganic Med. Chem. Lett. 2014, 24, 3251–3254. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Si, L.; Liu, D.; Proksch, P.; Zhang, L.; Zhou, D.; Lin, W. Neoechinulin B and its analogues as potential entry inhibitors of influenza viruses, targeting viral hemagglutinin. Eur. J. Med. Chem. 2015, 93, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Sacramento, C.Q.; Marttorelli, A.; Fintelman-Rodrigues, N.; de Freitas, C.S.; de Melo, G.R.; Rocha, M.E.; Kaiser, C.R.; Rodrigues, K.F.; da Costa, G.L.; Alves, C.M.; et al. Aureonitol, a Fungi-Derived Tetrahydrofuran, Inhibits Influenza Replication by Targeting Its Surface Glycoprotein Hemagglutinin. PLoS ONE 2015, 10, e0139236. [Google Scholar] [CrossRef]
- Ma, C.; Lin, Q.; Xue, Y.; Ju, Z.; Deng, G.; Liu, W.; Sun, Y.; Guan, H.; Cheng, X.; Wang, C. Pharmacokinetic studies of ginsenosides Rk1 and Rg5 in rats by UFLC-MS/MS. Biomed. Chromatogr. 2021, 35, e5108. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Fan, A.; Li, G.; Liu, Q. Pharmacokinetics and bioavailability study of ginsenoside Rk1 in rat by liquid chromatography/electrospray ionization tandem mass spectrometry. Biomed. Chromatogr. 2019, 33, e4580. [Google Scholar] [CrossRef]
- Douafer, H.; Andrieu, V.; Brunel, J.M. Scope and limitations on aerosol drug delivery for the treatment of infectious respiratory diseases. J. Control. Release 2020, 325, 276–292. [Google Scholar] [CrossRef]
- Dallas, R.H.; Rains, J.K.; Wilder, K.; Humphrey, W.; Cross, S.J.; Ghafoor, S.; Brazelton de Cardenas, J.N.; Hayden, R.T.; Hijano, D.R. The Aerogen(®) Solo Is an Alternative to the Small Particle Aerosol Generator (SPAG-2) for Administration of Inhaled Ribavirin. Pharmaceutics 2020, 12, 1163. [Google Scholar] [CrossRef]
- Gilbert, B.E.; Wyde, P.R.; Wilson, S.Z.; Robins, R.K. Aerosol and intraperitoneal administration of ribavirin and ribavirin triacetate: Pharmacokinetics and protection of mice against intracerebral infection with influenza A/WSN virus. Antimicrob. Agents Chemother. 1991, 35, 1448–1453. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Han, W.; Du, J.; Yang, X.; Duan, M.; Xu, C.; Zeng, Z.; Chen, W.; Chen, J. Chenodeoxycholic Acid from Bile Inhibits Influenza A Virus Replication via Blocking Nuclear Export of Viral Ribonucleoprotein Complexes. Molecules 2018, 23, 3315. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Liu, Y.; Li, S.; Sun, H.; Pan, L.; Cui, X.; Zhu, X.; Feng, Y.; Li, M.; Yu, Y.; Wu, M.; et al. Variation and Molecular Basis for Enhancement of Receptor Binding of H9N2 Avian Influenza Viruses in China Isolates. Front. Microbiol. 2020, 11, 602124. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | a CC50 (μM) | H1N1 (PR8) | H5N1 | H3N2 | |||
|---|---|---|---|---|---|---|---|
| b EC50 (μM) | c SI | b EC50 (μM) | c SI | EC50 (μM) | c SI | ||
| G-rk1 | 34.8 | 6.2 | 5.6 | 10.4 | 3.3 | 13.5 | 2.6 |
| G-rg5 | >40.0 | 8.2 | >5.0 | 14.8 | >2.8 | 11.5 | >3.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Sun, H.; Zhang, Z.; Ou, W.; Xu, F.; Luo, L.; Liu, Y.; Chen, W.; Chen, J. Antiviral Effect of Ginsenosides rk1 against Influenza a Virus Infection by Targeting the Hemagglutinin 1-Mediated Virus Attachment. Int. J. Mol. Sci. 2023, 24, 4967. https://doi.org/10.3390/ijms24054967
Yang X, Sun H, Zhang Z, Ou W, Xu F, Luo L, Liu Y, Chen W, Chen J. Antiviral Effect of Ginsenosides rk1 against Influenza a Virus Infection by Targeting the Hemagglutinin 1-Mediated Virus Attachment. International Journal of Molecular Sciences. 2023; 24(5):4967. https://doi.org/10.3390/ijms24054967
Chicago/Turabian StyleYang, Xia, Hailiang Sun, Zhening Zhang, Weixin Ou, Fengxiang Xu, Ling Luo, Yahong Liu, Weisan Chen, and Jianxin Chen. 2023. "Antiviral Effect of Ginsenosides rk1 against Influenza a Virus Infection by Targeting the Hemagglutinin 1-Mediated Virus Attachment" International Journal of Molecular Sciences 24, no. 5: 4967. https://doi.org/10.3390/ijms24054967
APA StyleYang, X., Sun, H., Zhang, Z., Ou, W., Xu, F., Luo, L., Liu, Y., Chen, W., & Chen, J. (2023). Antiviral Effect of Ginsenosides rk1 against Influenza a Virus Infection by Targeting the Hemagglutinin 1-Mediated Virus Attachment. International Journal of Molecular Sciences, 24(5), 4967. https://doi.org/10.3390/ijms24054967