
Red LED Light Improves Pepper (Capsicum annuum L.) Seed Radicle Emergence and Growth through the Modulation of Aquaporins, Hormone Homeostasis, and Metabolite Remobilization
 , ,
, ,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Results
2.1. R LED Light Accelerates Seed Hydration and Suppresses the Inhibitory Effect of Azide during Germination
2.2. R LED Light Increases the Radicle Emergence Rate, Germination Percentage, Germination Speed Index (GSI), and Humidity Percentage
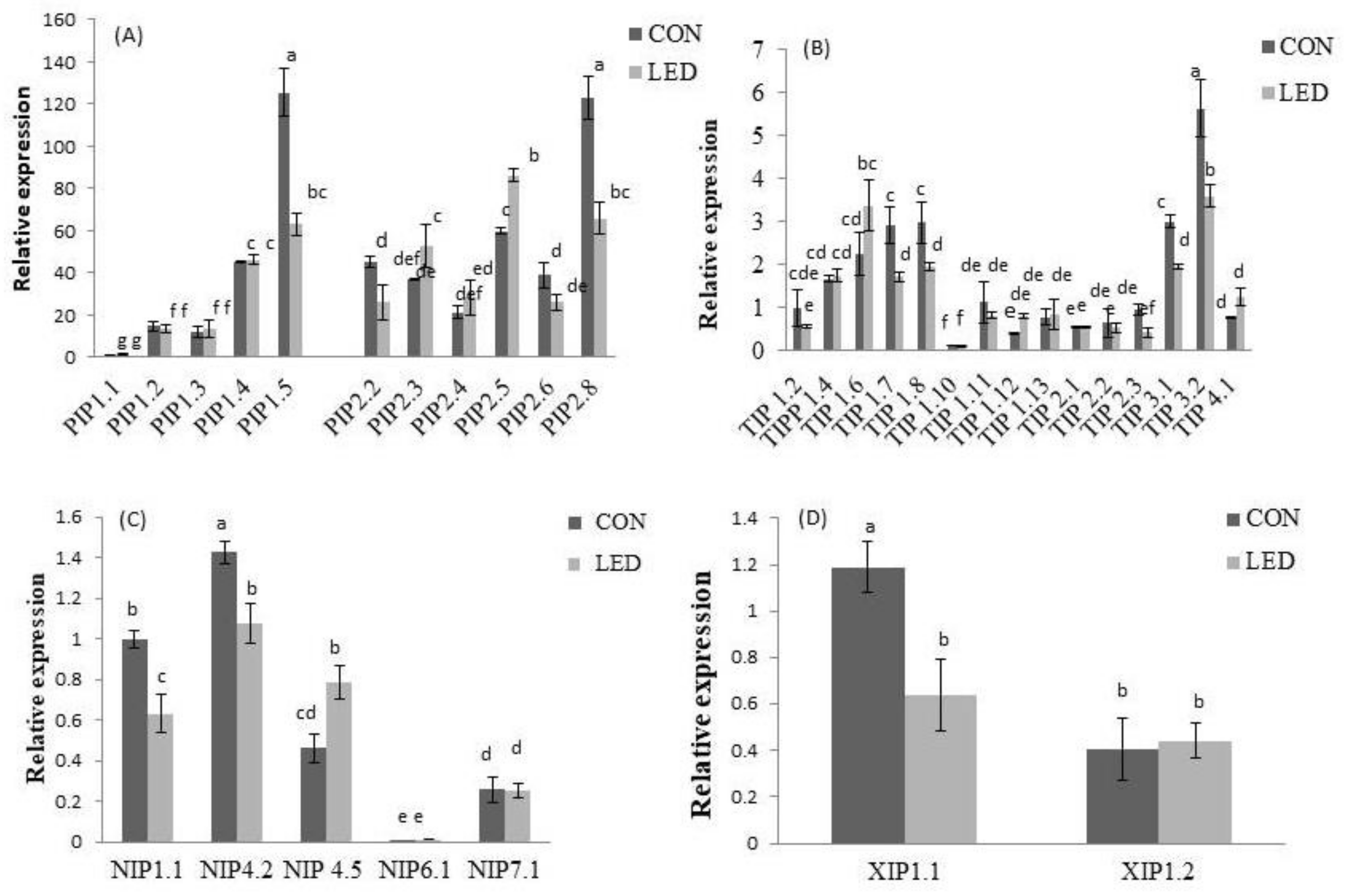
2.3. Seeds AQP Isoforms Were Differentially Expressed under Control and R LED-Irradiation during Phase III
2.4. R LED Irradiance Modulates the Levels of Phytohormones during Phase III
2.5. R LED Light Induces Changes in the Metabolite Content during Phase III
2.5.1. Sugars
2.5.2. Amino Acids
2.5.3. Organic Acids
2.5.4. Other Metabolites
2.6. Principal Component Analysis (PCA)
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
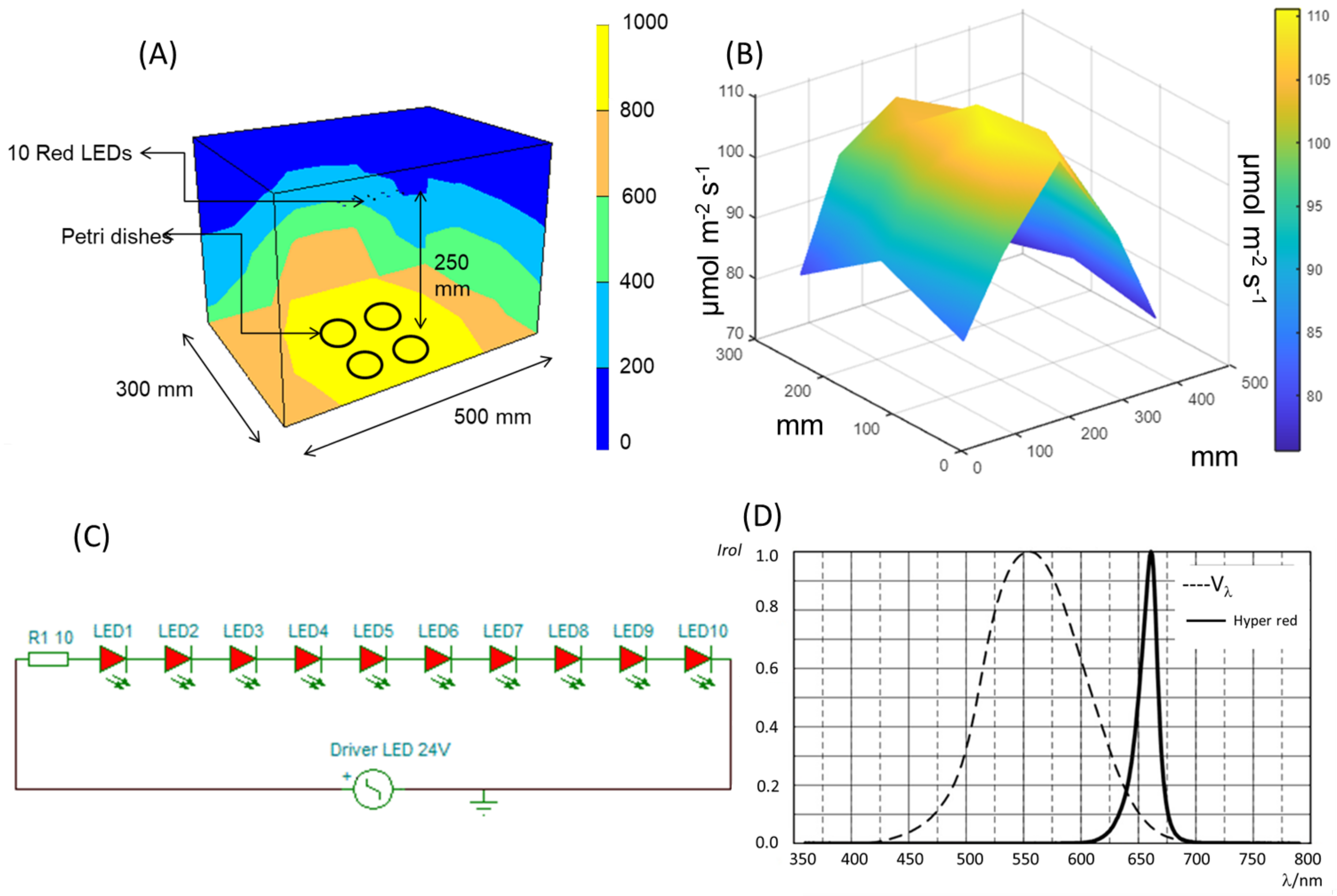
4.2. Red LED Light Design
4.3. Sodium Azide Treatment
4.4. Germination Assays
4.5. Viability of the Seeds
4.6. Imbibition Kinetics
4.7. RNA Extraction and Reverse Transcription
4.8. Quantitative Real-Time PCR (qRT-PCR) Analyses
4.9. Metabolite Analysis by 1H-NMR Spectroscopy
4.10. Hormone Analysis by UPLC-QToF-MS/MS
4.10.1. Sample Preparation
4.10.2. UPLC-QToF-MS/MS Analysis
4.11. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuan, P.A.; Sun, M.; Nguyen, T.N.; Park, S.; Ayele, B.T. Molecular mechanisms of seed germination. In Sprouted Grains: Nutritional Value, Production and Applications; Feng, H., Nemzer, B., DeVries, J.W., Eds.; AACC International Press: Cambridge, MA, USA, 2019; pp. 1–24. [Google Scholar]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination-Still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Kumari, A.; Kaur, R. Evaluation of benzyl-butyl phthalate induced germination and early growth vulnerability to barley seedlings (Hordeum vulgare L.). Indian J. Ecol. 2018, 45, 174–177. [Google Scholar]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Alves Junior, C.; de Oliveira Vitoriano, J.; da Silva, D.L.S.; de Lima Farias, M.; de Lima Dantas, N.B. Water uptake mechanism and germination of Erythrina velutina seeds treated with atmospheric plasma. Sci. Rep. 2016, 6, 33722. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, W. Molecular Biology of Aquaporins. Adv. Exp. Med. Biol. 2017, 969, 1–34. [Google Scholar] [PubMed]
- Bezerra-Neto, J.P.; de Araújo, F.C.; Ferreira-Neto, J.R.C.; da Silva, M.D.; Pandolfi, V.; Aburjaile, F.F.; Sakamoto, T.; de Oliveira Silva, R.L.; Kido, E.A.; Amorima, L.L.B.; et al. Plant aquaporins: Diversity, evolution and biotechnological applications. Curr. Protein Pept. Sci. 2019, 20, 368–395. [Google Scholar] [CrossRef]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef]
- Forrest, K.L.; Bhave, M. Major intrinsic proteins (MIPs) in plants: A complex gene family with major impacts on plant phenotype. Funct. Integr. Genom. 2007, 7, 263–289. [Google Scholar] [CrossRef]
- Cardoso, A.Á.; Souza, G.A.; Silva, C.J.; Pires, R.M.; Dias, D.C. Seed imbibition and germination of Plathymenia reticulata Benth. (Fabaceae) affected by mercury: Possible role of aquaporins. Acta Bot. Bras. 2015, 29, 285–291. [Google Scholar] [CrossRef]
- Obroucheva, N.V.; Sinkevich, I.A.; Lityagina, S.V.; Novikova, G.V. Water relations in germinating seeds. Rus. J. Plant Physiol. 2017, 64, 625–633. [Google Scholar] [CrossRef]
- Su, Y.; Liu, Z.; Sun, J.; Wu, C.; Li, Y.; Zhang, C.; Zhao, L. Genome-wide identification of maize aquaporin and functional analysis during seed germination and seedling establishment. Front. Plant Sci. 2022, 13, 831916. [Google Scholar] [CrossRef]
- Hoai, P.T.T.; Tyerman, S.D.; Schnell, N.; Tucker, M.; McGaughey, S.A.; Qiu, J.; Groszmann, M.; Byrt, C.S. Deciphering aquaporin regulation and roles in seed biology. J. Exp. Bot. 2020, 71, 1763–1773. [Google Scholar] [CrossRef]
- Veselova, T.V.; Veselovskii, V.A.; Usmanov, P.D.; Usmanova, O.V.; Kozar, V.I. Hypoxia and imbibition injuries to aging seeds. Russ. J. Plant Physiol. 2003, 50, 835–842. [Google Scholar] [CrossRef]
- Willigen, C.V.; Postaire, O.; Tournaire-Roux, C.; Boursiac, Y.; Maurel, C. Expression and inhibition of aquaporins in germinating Arabidopsis seeds. Plant Cell Physiol. 2006, 47, 1241–1250. [Google Scholar] [CrossRef]
- Obroucheva, N.V.; Lityagina, S.V.; Novikova, G.V.; Sinkevich, I.A. Vacuolar status and water relations in embryonic axes of recalcitrant Aesculus hippocastanum seeds during stratification and early germination. AoB Plants 2012, 2012, pls008. [Google Scholar] [CrossRef]
- Novikova, G.V.; Tournaire-Roux, C.; Sinkevich, I.A.; Lityagina, S.V.; Maurel, C.; Obroucheva, N. Vacuolar biogenesis and aquaporin expression at early germination of broad bean seeds. Plant Physiol. Biochem. 2014, 82, 123–132. [Google Scholar] [CrossRef]
- Balarynová, J.; Fellner, M. Expression of genes encoding PIP aquaporins in tomato seeds exposed to blue radiation and mercury. Biol. Plant. 2019, 63, 1–10. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Z.; Liu, F.; Sun, L.; Hao, F. Versatile roles of aquaporins in plant growth and development. Int. J. Mol. Sci. 2020, 21, 9485. [Google Scholar] [CrossRef]
- Béré, E.; Lahbib, K.; Merceron, B.; Fleurat-Lessard, P.; Boughanmi, N.G. α-TIP aquaporin distribution and size tonoplast variation in storage cells of Vicia faba cotyledons at seed maturation and germination stages. J. Plant Physiol. 2017, 216, 145–151. [Google Scholar] [CrossRef]
- Maurel, C.; Verdoucq, L.; Luu, D.T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef]
- Reinhardt, H.; Hachez, C.; Bienert, M.D.; Beebo, A.; Swarup, K.; Voß, U.; Bouhidel, K.; Frigerio, L.; Schjoerring, J.K.; Bennett, M.J.; et al. Tonoplast aquaporins facilitate lateral root emergence. Plant Physiol. 2016, 170, 1640–1654. [Google Scholar] [CrossRef]
- Footitt, S.; Clewes, R.; Feeney, M.; Finch-Savage, W.E.; Frigerio, L. Aquaporins influence seed dormancy and germination in response to stress. Plant Cell Environ. 2019, 8, 2325–2339. [Google Scholar] [CrossRef] [PubMed]
- Amir, R.; Galili, G.; Cohen, H. The metabolic roles of free amino acids during seed development. Plant Sci. 2018, 275, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Marsolais, F.; Bernards, M.A.; Sumarah, M.W.; Bykova, N.V.; Igamberdiev, A.U. Glyoxylate cycle and metabolism of organic acids in the scutellum of barley seeds during germination. Plant Sci. 2016, 248, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Lynch, T.J. Abscisic acid inhibition of radicle emergence but not seedling growth is suppressed by sugars. Plant Physiol. 2000, 122, 1179–1186. [Google Scholar] [CrossRef]
- Manz, B.; Muller, K.; Kucera, B.; Volke, F.; Leubner-Metzger, G. Water uptake and distribution in germinating tobacco seeds investigated in vivo by nuclear magnetic resonance imaging. Plant Physiol. 2005, 138, 1538–1551. [Google Scholar] [CrossRef]
- Ogawa, M.; Hanada, A.; Yamauchi, Y.; Kuwahara, A.; Kamiya, Y.; Yamaguchi, S. Gibberellin biosynthesis and response during Arabidopsis seed germination. Plant Cell 2003, 15, 1591–1604. [Google Scholar] [CrossRef]
- Stamm, P.; Topham, A.T.; Mukhtar, N.K.; Jackson, M.D.; Tomé, D.F.; Beynon, J.L.; Bassel, G.W. The transcription factor ATHB5 affects GA-mediated plasticity in hypocotyl cell growth during seed germination. Plant Physiol. 2017, 173, 907–917. [Google Scholar] [CrossRef]
- Leubner-Metzger, G. Brassinosteroids promote seed germination. In Brassinosteroids; Hayat, S., Ahmad, A., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 119–128. [Google Scholar]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant. 2016, 9, 34–45. [Google Scholar] [CrossRef]
- Gomes, M.P.; Smedbol, É.; Carneiro, M.M.L.C.; Garcia, Q.S.; Juneau, P. Reactive oxygen species and plant hormones. In Oxidative Damage to Plants. Antioxidant Networks and Signalling; Ahmad, P., Ed.; Academic Press: Amsterdam, The Netherlands, 2014; pp. 65–88. [Google Scholar]
- Sánchez-Linares, L.; Gavilanes-Ruíz, M.; Díaz-Pontones, D.; Guzmán-Chávez, F.; Calzada-Alejo, V.; Zurita-Villegas, V.; Luna-Loaiza, V.; Moreno-Sánchez, R.; Bernal-Lugo, I.; Sánchez-Nieto, S. Early carbon mobilization and radicle protrusion in maize germination. J. Exp. Bot. 2012, 63, 4513–4526. [Google Scholar] [CrossRef]
- Neff, M.M. Light-mediated seed germination: Connecting phytochrome B to gibberellic acid. Dev. Cell. 2012, 22, 687–688. [Google Scholar] [CrossRef]
- Yang, L.; Liu, S.; Lin, R. The role of light in regulating seed dormancy and germination. J. Integr. Plant Biol. 2020, 62, 1310–1326. [Google Scholar] [CrossRef]
- Diel, M.I.; Valera, O.V.S.; Pinheiro, M.V.M.; Thiesen, L.A.; Meira, D.; de Melo, P.J.; Junges, D.L.; Caron, B.O.; Schmidt, D. Temperature and light quality influence seed germination of two biquinho pepper cultivars. Bulg. J. Agric. Sci. 2019, 25, 1007–1014. [Google Scholar]
- Sanoubar, R.; Calone, R.; Noli, E.; Barbanti, L. Data on seed germination using LED versus fluorescent light under growth chamber conditions. Data Brief 2018, 19, 594–600. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.D. Impact of light-emitting diodes (LEDs) and its potential on plant growth and development in controlled-environment plant production system. Curr. Biotechnol. 2016, 5, 28–43. [Google Scholar] [CrossRef]
- McCree, K.J. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Meteorol. 1971, 9, 191–216. [Google Scholar] [CrossRef]
- Kusuma, P.; Pattison, P.M.; Bugbee, B. From physics to fixtures to food: Current and potential LED efficacy. Hort. Res. 2020, 7, 56. [Google Scholar] [CrossRef]
- Tang, D.; Hamayun, M.; Latif, A. Germination of some important weeds influenced by red light and nitrogenous compounds. Pak. J. Bot. 2010, 42, 3739–3745. [Google Scholar]
- Pardo, G.P.; Velázquez, S.T.; Orea, A.C.; Aguilar, C.H.; Pacheco, F.A.D.; Martínez, F.R. Pulsed LED light in germination and growth of lettuce seeds. Bothalia J. 2016, 46, 13–26. [Google Scholar]
- Hernández-Adasme, C.; Silva, H.; Escalona, V. In-door germination and seedling growth of green and red lettuce under LED-light spectrum and subsequent effect on baby leaf lettuce. Ital. J. Agron. 2022, 17, 1982. [Google Scholar] [CrossRef]
- Tehrani, F.P.; Majd, A.; Mahmoodzadeh, H.; Satari, T.N. Effect of red and blue light-emitting diodes on germination, morphological and anatomical features of Brassica napus . Adv. Stud. Biol. 2016, 8, 173–180. [Google Scholar] [CrossRef]
- Hawley, D.W. Development of a Variable-Spectra LED Array for Optimized Plant Development. Master’s Thesis, University of Guelph, Guelph, ON, Canada, 2013. Available online: https://atrium.lib.uoguelph.ca/xmlui/bitstream/handle/10214/7718/Hawley_David_2013_MSc.pdf?sequence=1&isAllowed=y (accessed on 12 October 2022).
- Cho, J.N.; Ryu, J.Y.; Jeong, Y.M.; Park, J.; Song, J.J.; Amasino, R.M.; Noh, B.; Noh, Y.S. Control of seed germination by light-induced histone arginine demethylation activity. Dev. Cell 2012, 22, 736–748. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, K.L.; Reid, R.J. The involvement of aquaglyceroporins in transport of boron in barley roots. Plant Cell Environ. 2009, 32, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Sutka, M.; Li, G.; Boudet, J.; Boursiac, Y.; Doumas, P.; Maurel, C. Natural variation of root hydraulics in Arabidopsis grown in normal and salt-stressed conditions. Plant Physiol. 2011, 155, 1264–1276. [Google Scholar] [CrossRef] [PubMed]
- Grondin, A.; Mauleon, R.; Vadez, V.; Henry, A. Root aquaporins contribute to whole plant water fluxes under drought stress in rice (Oryza sativa L.). Plant Cell Environ. 2016, 39, 347–365. [Google Scholar] [CrossRef]
- Schuurmans, J.A.; van Dongen, J.T.; Rutjens, B.P.; Boonman, A.; Pieterse, C.M.; Borstlap, A.C. Members of the aquaporin family in the developing pea seed coat include representatives of the PIP, TIP, and NIP subfamilies. Plant Mol. Biol. 2003, 53, 655–667. [Google Scholar] [CrossRef]
- Liu, H.-Y.; Yu, X.; Cui, D.-Y.; Sun, M.-H.; Sun, W.-N.; Tang, Z.-C.; Kwak, S.-S.; Su, W.-A. The role of water channel proteins and nitric oxide signaling in rice seed germination. Cell Res. 2007, 17, 638–649. [Google Scholar] [CrossRef]
- Liu, C.; Fukumoto, T.; Matsumoto, T.; Gena, P.; Frascaria, D.; Kaneko, T.; Katsuhara, M.; Zhong, S.; Sun, X.; Zhu, Y.; et al. Aquaporin OsPIP1;1 promotes rice salt resistance and seed germination. Plant Physiol. Biochem. 2013, 63, 151–158. [Google Scholar] [CrossRef]
- Houreld, N. Visible red and near infra-red light is absorbed by cytochrome c oxidase and stimulates the production of ATP. In Adenosine Triphosphate: Chemical Properties, Biosynthesis and Function in Cells; Kuester, E., Traugott, G., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2013; pp. 161–176. [Google Scholar]
- Karu, T.I. Cellular and molecular mechanisms of photobiomodulation (low-power laser therapy). IEEE J. Sel. Top. Quantum Electron. 2013, 20, 143–148. [Google Scholar] [CrossRef]
- Catalán, J.; Papas, M.; Trujillo-Rojas, L.; Blanco-Prieto, O.; Bonilla-Correal, S.; Rodríguez-Gil, J.E.; Miró, J.; Yeste, M. Red LED light acts on the mitochondrial electron chain of donkey sperm and its effects depend on the time of exposure to light. Front. Cell Dev. Biol. 2020, 8, 588621. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Aroca, R.; Chaumont, F.; Ruiz-Lozano, J.M. Contribution of the arbuscular mycorrhizal symbiosis to the regulation of radial root water transport in maize plants under water deficit. Environ. Exp. Bot. 2019, 167, 103821. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R. Exogenous catalase and ascorbate modify the effects of abscisic acid (ABA) on root hydraulic properties in Phaseolus vulgaris L. plants. J. Plant Growth Regul. 2006, 25, 10–17. [Google Scholar] [CrossRef]
- Benabdellah, K.; Ruiz-Lozano, J.M.; Aroca, R. Hydrogen peroxide effects on root hydraulic properties and plasma membrane aquaporin regulation in Phaseolus vulgaris . Plant Mol. Biol. 2009, 70, 647–661. [Google Scholar] [CrossRef]
- Lechowska, K.; Kubala, S.; Wojtyla, Ł.; Nowaczyk, G.; Quinet, M.; Lutts, S.; Garnczarska, M. New insight on water status in germinating Brassica napus seeds in relation to priming-improved germination. Int. J. Mol. Sci. 2019, 20, 540. [Google Scholar] [CrossRef]
- Steinbrecher, T.; Leubner-Metzger, G. The biomechanics of seed germination. J. Exp. Bot. 2017, 68, 765–783. [Google Scholar] [CrossRef]
- Nonogaki, H. Seed germination and dormancy: The classic story, new puzzles, and evolution. J. Integr. Plant Biol. 2019, 61, 541–563. [Google Scholar] [CrossRef]
- Obroucheva, N.V. Aquaporins in seeds. Seed Sci. Res. 2013, 23, 213–216. [Google Scholar] [CrossRef]
- Shijneva, I.A.; Novikova, G.V.; Obroucheva, N.V. Aquaporins of tonoplast and plasmalemma in axial organs of germinating broad bean seeds. Dokl. Biochem. Biophys. 2007, 413, 43–46. [Google Scholar] [CrossRef]
- Gattolin, S.; Sorieul, M.; Frigerio, L. Mapping of tonoplast intrinsic proteins in maturing and germinating Arabidopsis seeds reveals dual localization of embryonic tips to the tonoplast and plasma membrane. Mol. Plant 2011, 4, 180–189. [Google Scholar] [CrossRef]
- Maurel, C.; Kado, R.T.; Guern, J.; Chrispeels, M.J. Phosphorylation regulates the water channel activity of the seed-specific aquaporin alpha-TIP. EMBO J. 1995, 14, 3028–3035. [Google Scholar] [CrossRef]
- Li, G.W.; Peng, Y.H.; Yu, X.; Zhang, M.H.; Cai, W.M.; Sun, W.N.; Su, W.A. Transport functions and expression analysis of vacuolar membrane aquaporins in response to various stresses in rice. J. Plant Physiol. 2008, 165, 1879–1888. [Google Scholar] [CrossRef]
- Shiota, H.; Sudoh, T.; Tanaka, I. Expression analysis of genes encoding plasma membrane aquaporins during seed and fruit development in tomato. Plant Sci. 2006, 171, 277–285. [Google Scholar] [CrossRef]
- Ludevid, D.; Höfte, H.; Himelblau, E.; Chrispeels, M.J. The expression pattern of the tonoplast intrinsic protein γ-TIP in Arabidopsis thaliana is correlated with cell enlargement. Plant Physiol. 1992, 100, 1633–1639. [Google Scholar] [CrossRef]
- Kamiya, T.; Tanaka, M.; Mitani, N.; Ma, J.F.; Maeshima, M.; Fujiwara, T. NIP1;1, an aquaporin homolog, determines the arsenite sensitivity of Arabidopsis thaliana . J. Biol. Chem. 2009, 284, 2114–2120. [Google Scholar] [CrossRef]
- Schmid, M.; Davison, T.S.; Henz, S.R.; Pape, U.J.; Demar, M.; Vingron, M.; Schölkopf, B.; Weigel, D.; Lohmann, J.U. A gene expression map of Arabidopsis thaliana development. Nat. Genet. 2005, 37, 501–506. [Google Scholar] [CrossRef]
- Le, B.H.; Cheng, C.; Bui, A.Q.; Wagmaister, J.A.; Henry, K.F.; Pelletier, J.; Kwong, L.; Belmonte, M.; Kirkbride, R.; Horvath, S.; et al. Global analysis of gene activity during Arabidopsis seed development and identification of seed-specific transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 8063–8070. [Google Scholar] [CrossRef]
- Barozzi, F.; Papadia, P.; Stefano, G.; Renna, L.; Brandizzi, F.; Migoni, D.; Fanizzi, F.P.; Piro, G.; Di Sansebastiano, G.-P. Variation in membrane trafficking linked to SNARE AtSYP51 interaction with aquaporin NIP1;1. Front. Plant Sci. 2019, 9, 1949. [Google Scholar] [CrossRef]
- De Caroli, M.; Barozzi, F.; Renna, L.; Piro, G.; Di Sansebastiano, G.-P. Actin and microtubules differently contribute to vacuolar targeting specificity during the export from the ER. Membranes 2021, 11, 299. [Google Scholar] [CrossRef]
- Kuo, T.M.; Doehlert, D.C.; Crawford, C.G. Sugar metabolism in germinating soybean seeds: Evidence for the sorbitol pathway in soybean axes. Plant Physiol. 1990, 93, 1514–1520. [Google Scholar] [CrossRef]
- Tesfay, S.Z.; Modi, A.T.; Mohammed, F. The effect of temperature in moringa seed phytochemical compounds and carbohydrate mobilization. S. Afr. J. Bot. 2016, 102, 190–196. [Google Scholar] [CrossRef]
- Romero-Rodríguez, M.C.; Archidona-Yuste, A.; Abril, N.; Gil-Serrano, A.M.; Meijón, M.; Jorrín-Novo, J.V. Germination and early seedling development in Quercus ilex recalcitrant and non-dormant seeds: Targeted transcriptional, hormonal, and sugar analysis. Front. Plant Sci. 2018, 9, 1508. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.; Schlappal, A.; Geller, E.; Castonguay, T.W. Fructose-induced hypertriglyceridemia: A review. In Nutrition in the Prevention and Treatment of Abdominal Obesity; Watson, R.R., Ed.; Academic Press: Amsterdam, The Netherlands, 2014; pp. 197–205. [Google Scholar]
- Koops, A.J.; Groeneveld, H.W. Mobilization of endosperm reserves and uptake of sucrose and valine by the cotyledons of Euphorbia lathyris L. J. Exp. Bot. 1990, 41, 1279–1285. [Google Scholar] [CrossRef]
- Buckeridge, M.S.; Santos, H.P.; Tine, M.A.; Aidar, M.P.M. Mobilização de reservas. In Germinação: Do Básico ao Aplicado; Ferreira, A.G., Borguetti, F., Eds.; Artmed Editora: Porto Alegre, Brazil, 2004; pp. 163–185. [Google Scholar]
- Boter, M.; Calleja-Cabrera, J.; Carrera-Castaño, G.; Wagner, G.; Hatzig, S.V.; Snowdon, R.J.; Legoahec, L.; Bianchetti, G.; Bouchereau, A.; Nesi, N.; et al. An integrative approach to analyze seed germination in Brassica napus . Front. Plant Sci. 2019, 10, 1342. [Google Scholar] [CrossRef]
- Lea, P.J.; Joy, K.W. Amino acid interconversion in germinating seeds. In Mobilization of Reserves in Germination; Nozzolillo, C., Lea, P.J., Loewus, F.A., Eds.; Springer: New York, NY, USA, 1983; Volume 17, pp. 77–109. [Google Scholar]
- Hildebrandt, T.M.; Nesi, A.N.; Araújo, W.L.; Braun, H.-P. Amino acid catabolism in plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef]
- Newsholme, P.; Stenson, L.; Sulvucci, M.; Sumayao, R.; Krause, M. Amino acid metabolism. In Comprehensive Biotechnology; Young, M.M., Ed.; Academic Press: Dublin, Ireland, 2011; pp. 3–14. [Google Scholar]
- Glevarec, G.; Bouton, S.; Jaspard, E.; Riou, M.-T.; Cliquet, J.-B.; Suzuki, A.; Limami, A.M. Respective roles of the glutamine synthetase/glutamate synthase cycle and glutamate dehydrogenase in ammonium and amino acid metabolism during germination and post-germinative growth in the model legume Med . Truncatula. Planta 2004, 219, 286–297. [Google Scholar] [CrossRef]
- Lee, N.; Kim, D. Toxic metabolites and inborn errors of amino acid metabolism: What one informs about the other. Metabolites 2022, 12, 527. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G.; Hodges, M. Respiration and nitrogen assimilation: Targeting mitochondria-associated metabolism as a means to enhance nitrogen use efficiency. J. Exp. Bot. 2011, 62, 1467–1482. [Google Scholar] [CrossRef]
- Hooks, M.A.; Allwood, J.W.; Harrison, J.K.D.; Kopka, J.; Erban, A.; Goodacre, R.; Balk, J. Selective induction and subcellular distribution of ACONITASE 3 reveal the importance of cytosolic citrate metabolism during lipid mobilization in Arabidopsis . Biochem. J. 2014, 463, 309–317. [Google Scholar] [CrossRef]
- Silva, A.T.; Ligterink, W.; Hilhorst, H.W.M. Metabolite profiling and associated gene expression reveal two metabolic shifts during the seed-to-seedling transition in Arabidopsis thaliana . Plant Mol. Biol. 2017, 95, 481–496. [Google Scholar] [CrossRef]
- Tahjib-Ul-Arif, M.; Zahan, M.I.; Karim, M.M.; Imran, S.; Hunter, C.T.; Islam, M.S.; Mia, M.A.; Hannan, M.A.; Rhaman, M.S.; Hossain, M.A.; et al. Citric acid-mediated abiotic stress tolerance in plants. Int. J. Mol. Sci. 2021, 22, 7235. [Google Scholar] [CrossRef]
- Monroy-Vázquez, M.E.; Peña-Valdivia, C.B.; García-Nava, J.R.; Solano-Camacho, E.; Campos, H.; García-Villanueva, E. Imbibition, viability and vigor of seeds of four Opuntia species with different degree of domestication. Agrociencia 2017, 51, 27–42. [Google Scholar]
- Victoria, T.; Jorge, A.; Bonilla, C.; Carmen, R.; Sánchez, O.; Manuel, S. Viabilidad en tetrazolio de semillas de caléndula y eneldo. Acta Agron. 2006, 55, 31–41. [Google Scholar]
- Ferraresi, D.A.; Yamashita, O.M.; de Carvalho, M.A.C. Superação da dormência e qualidade de luz na germinação de sementes de Murdannia nudiflora (L.) Brenans. Rev. Bras. Sementes 2009, 31, 126–132. [Google Scholar] [CrossRef]
- Wang, G.; Chen, Y.; Fan, H.; Huang, P. Effects of light-emitting diode (LED) red and blue light on the growth and photosynthetic characteristics of Momordica charantia L. J. Agric. Chem. Environ. 2021, 10, 105798. [Google Scholar]
- Bae, G.; Choi, G. Decoding of light signals by plant phytochromes and their interacting proteins. Annu. Rev. Plant Biol. 2008, 59, 281–311. [Google Scholar] [CrossRef]
- Legris, M.; Ince, Y.Ç.; Fankhauser, C. Molecular mechanisms underlying phytochrome-controlled morphogenesis in plants. Nat. Commun. 2019, 10, 5219. [Google Scholar] [CrossRef]
- Seo, M.; Nambara, E.; Choi, G.; Yamaguchi, S. Interaction of light and hormone signals in germinating seeds. Plant Mol. Biol. 2009, 69, 463–472. [Google Scholar] [CrossRef]
- Mérai, Z.; Graeber, K.; Wilhelmsson, P.; Ullrich, K.K.; Arshad, W.; Grosche, C.; Tarkowská, D.; Turečková, V.; Strnad, M.; Rensing, S.A.; et al. Aethionema arabicum: A novel model plant to study the light control of seed germination. J. Exp. Bot. 2019, 70, 3313–3328. [Google Scholar] [CrossRef]
- Millar, A.A.; Jacobson, J.V.; Ross, J.J.; Helliwell, C.A.; Poole, A.T.; Scofield, G.; Reid, J.B.; Gubler, F. Seed dormancy and ABA metabolism in Arabidopsis and barley: The role of ABA 8’-hydroxylase. Plant J. 2006, 45, 942–954. [Google Scholar] [CrossRef]
- Okamoto, M.; Kuwahara, A.; Seo, M.; Kushiro, T.; Asami, T.; Hirai, N.; Kamiya, Y.; Koshiba, T.; Nambara, E. CYP707A1 and CYP707A2, which encode abscisic acid 8’-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant Physiol. 2006, 141, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Pawela, A.; Banasiak, J.; Biała, W.; Martinoia, E.; Jasiński, M. MtABCG20 is an ABA exporter influencing root morphology and seed germination of Medicago truncatula . Plant J. 2019, 98, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, S.; Sun, N.; Liu, H.; Zhao, Y.; Liang, Y.; Zhang, L.; Han, Y. Functional diversity of jasmonates in rice. Rice 2015, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Alallaq, S.; Ranjan, A.; Brunoni, F.; Novák, O.; Lakehal, A.; Bellini, C. Red light controls adventitious root regeneration by modulating hormone homeostasis in Picea abies seedlings. Front. Plant Sci. 2020, 11, 586140. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Hu, Y.; Wang, H.; Guo, Q.; Chen, Y.; Howe, G.A.; Yu, D. Molecular mechanism underlying the synergetic effect of jasmonate on abscisic acid signaling during seed germination in Arabidopsis. Plant Cell 2020, 32, 3846–3865. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ma, W.; Shen, S.; Gu, A. Underlying biochemical and molecular mechanisms for seed germination. Int. J. Mol. Sci. 2022, 23, 8502. [Google Scholar] [CrossRef]
- Chung, I.M.; Paudel, N.; Kim, S.-H.; Yu, C.Y.; Ghimire, B.K. Correction to: The influence of light wavelength on growth and antioxidant capacity in Pachyrhizus erosus (L.) urban. J. Plant Growth Regul. 2020, 39, 954. [Google Scholar] [CrossRef]
- Dhakal, R.; Park, E.; Lee, S.-W.; Baek, K.-H. Soybean (Glycine max L. Merr.) sprouts germinated under red light irradiation induce disease resistance against bacterial rotting disease. PLoS ONE 2015, 10, e0117712. [Google Scholar] [CrossRef]
- Gallé, Á.; Czékus, Z.; Tóth, L.; Galgóczy, L.; Poór, P. Pest and disease management by red light. Plant Cell Environ. 2021, 44, 3197–3210. [Google Scholar] [CrossRef]
- Barba-Espín, G.; Diaz-Vivancos, P.; Job, D.; Belghazi, M.; Job, C.; Hernández, J.A. Understanding the role of H2O2 during pea seed germination: A combined proteomic and hormone profiling approach. Plant Cell Environ. 2011, 34, 1907–1919. [Google Scholar] [CrossRef]
- Koornneef, A.; Leon-Reyes, A.; Ritsema, T.; Verhage, A.; Den Otter, F.C.; Van Loon, L.C.; Pieterse, C.M.J. Kinetics of salicylate-mediated suppression of jasmonate signalling reveal a role for redox modulation. Plant Physiol. 2008, 147, 1358–1368. [Google Scholar] [CrossRef]
- Van der Does, D.; Leon-Reyes, A.; Koornneef, A.; Van Verk, M.C.; Rodenburg, N.; Pauwels, L.; Goossens, A.; Körbes, A.P.; Memelink, J.; Ritsema, T.; et al. Salicylic acid suppresses jasmonic acid signaling downstream of SCFCOI1-JAZ by targeting GCC promoter motifs via transcription factor ORA59. Plant Cell 2013, 25, 744–761. [Google Scholar] [CrossRef]
- Kumar, V.; Shriram, V.; Hoque, T.S.; Hasan, M.M.; Burritt, D.J.; Hossain, M.A. Glycinebetaine-mediated abiotic oxidative-stress tolerance in plants: Physiological and biochemical mechanisms. In Stress Signaling in Plants: Genomics and Proteomics Perspective; Sarwat, M., Ahmad, A., Abdin, M.Z., Ibrahim, M.M., Eds.; Springer: Cham, Switzerland, 2017; Volume 2, pp. 111–133. [Google Scholar]
- Khan, N.; Syeed, S.; Masood, A.; Nazar, R.; Iqbal, N. Application of salicylic acid increases contents of nutrients and antioxidative metabolism in mungbean and alleviates adverse effects of salinity stress. Int. J. Plant Biol. 2010, 1, e1. [Google Scholar] [CrossRef]
- Yang, N.; Ding, R.; Liu, J. Synthesizing glycine betaine via choline oxidation pathway as an osmoprotectant strategy in Haloferacales. Gene 2022, 847, 146886. [Google Scholar] [CrossRef]
- Strader, L.C.; Wheeler, D.L.; Christensen, S.E.; Berens, J.C.; Cohen, J.D.; Rampey, R.A.; Bartel, B. Multiple facets of Arabidopsis seedling development require indole-3-butyric acid-derived auxin. Plant Cell 2011, 23, 984–999. [Google Scholar] [CrossRef]
- Damodaran, S.; Strader, L.C. Indole 3-butyric acid metabolism and transport in Arabidopsis thaliana . Front. Plant Sci. 2019, 10, 851. [Google Scholar] [CrossRef]
- Rinaldi, M.A.; Patel, A.B.; Park, J.; Lee, K.; Strader, L.C.; Bartel, B. The roles of β-oxidation and cofactor homeostasis in peroxisome distribution and function in Arabidopsis thaliana . Genetics 2016, 204, 1089–1115. [Google Scholar] [CrossRef]
- Ross, J.J.; O’Neill, D.P.; Smith, J.J.; Kerckhoffs, L.H.; Elliott, R.C. Evidence that auxin promotes gibberellin A1 biosynthesis in pea. Plant J. 2000, 21, 547–552. [Google Scholar] [CrossRef]
- ISTA (2014). International Seed Testing Association web site. Available online: www.seedtest.org (accessed on 12 October 2022).
- Martínez-Solís, J.; Romero, A.S.; Pena, G.; Virgen, J. Índice de velocidad de emergencia en líneas de maíz. Rev. Mex. Cienc. Agríc. 2010, 1, 288–303. [Google Scholar]
- Domínguez-Domínguez, S.; Domínguez-López, A.; González-Huerta, A.; Navarro-Galindo, S. Cinética de imbibición e isotermas de adsorción de humedad de la semilla de jamaica (Hibiscus sabdariffa L.). Rev. Mex. Ing. Quím. 2007, 6, 309–316. [Google Scholar]
- Muries, B.; Faize, M.; Carvajal, M.; Martínez Ballesta, M.C. Identification and differential induction of the expression of aquaporins by salinity in broccoli plants. Mol. Biosyst. 2011, 7, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Uppuluri, L.S.; Motukuri, S.R.K.; Kumar, D. Genome-wide identification, characterization of aquaporin gene family and understanding aquaporin transport system in hot pepper (Capsicum annuum L.). Sci. Hortic. 2021, 286, 110206. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Pang, X.; Wan, H.; Ahammed, G.J.; Yu, J.; Yao, Z.; Ruan, M.; Ye, Q.; Li, Z.; Wang, R.; et al. Identification of optimal reference genes for normalization of qPCR analysis during pepper fruit development. Front. Plant Sci. 2017, 8, 1128. [Google Scholar] [CrossRef]
- Van de Weijer, T.; Schrauwen-Hinderling, V.B. Application of magnetic resonance spectroscopy in metabolic research. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 741–748. [Google Scholar] [CrossRef]
- Müller, M.; Munné-Bosch, S. Rapid and sensitive hormonal profiling of complex plant samples by liquid chromatography coupled to electrospray ionization tandem mass spectrometry. Plant Methods 2011, 7, 37. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GP (%) | GSI | Viability (%) | Humidity (%) | |
|---|---|---|---|---|
| CON | 88 ± 1.63 b | 0.35 ± 0.03 b | 86 ± 4 a | 2.64 ± 0.18 b |
| LED | 93 ± 1.91 a | 0.37 ± 0.03 a | 90 ± 5 a | 3.27 ± 0.20 a |
| SA | IBA | IAA | GA3 | 6-BA | ABA | JA | |
|---|---|---|---|---|---|---|---|
| CON | 88.04 ± 1.59 b | n.d | n.d | n.d | n.d | 13.44 ± 1.13 | 648.67 ± 6.35 a |
| LED | 116.24 ± 2.58 a | 7.26 ± 0.23 | n.d | n.d | n.d | n.d | 573.11 ± 10.95 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaghdoud, C.; Ollio, I.; Solano, C.J.; Ochoa, J.; Suardiaz, J.; Fernández, J.A.; Martínez Ballesta, M.d.C. Red LED Light Improves Pepper (Capsicum annuum L.) Seed Radicle Emergence and Growth through the Modulation of Aquaporins, Hormone Homeostasis, and Metabolite Remobilization. Int. J. Mol. Sci. 2023, 24, 4779. https://doi.org/10.3390/ijms24054779
Zaghdoud C, Ollio I, Solano CJ, Ochoa J, Suardiaz J, Fernández JA, Martínez Ballesta MdC. Red LED Light Improves Pepper (Capsicum annuum L.) Seed Radicle Emergence and Growth through the Modulation of Aquaporins, Hormone Homeostasis, and Metabolite Remobilization. International Journal of Molecular Sciences. 2023; 24(5):4779. https://doi.org/10.3390/ijms24054779
Chicago/Turabian StyleZaghdoud, Chokri, Irene Ollio, Cristóbal J. Solano, Jesús Ochoa, Juan Suardiaz, Juan A. Fernández, and María del Carmen Martínez Ballesta. 2023. "Red LED Light Improves Pepper (Capsicum annuum L.) Seed Radicle Emergence and Growth through the Modulation of Aquaporins, Hormone Homeostasis, and Metabolite Remobilization" International Journal of Molecular Sciences 24, no. 5: 4779. https://doi.org/10.3390/ijms24054779
APA StyleZaghdoud, C., Ollio, I., Solano, C. J., Ochoa, J., Suardiaz, J., Fernández, J. A., & Martínez Ballesta, M. d. C. (2023). Red LED Light Improves Pepper (Capsicum annuum L.) Seed Radicle Emergence and Growth through the Modulation of Aquaporins, Hormone Homeostasis, and Metabolite Remobilization. International Journal of Molecular Sciences, 24(5), 4779. https://doi.org/10.3390/ijms24054779