
TIMAP, a Regulatory Subunit of Protein Phosphatase 1, Inhibits In Vitro Neuronal Differentiation



Abstract
1. Introduction
2. Results
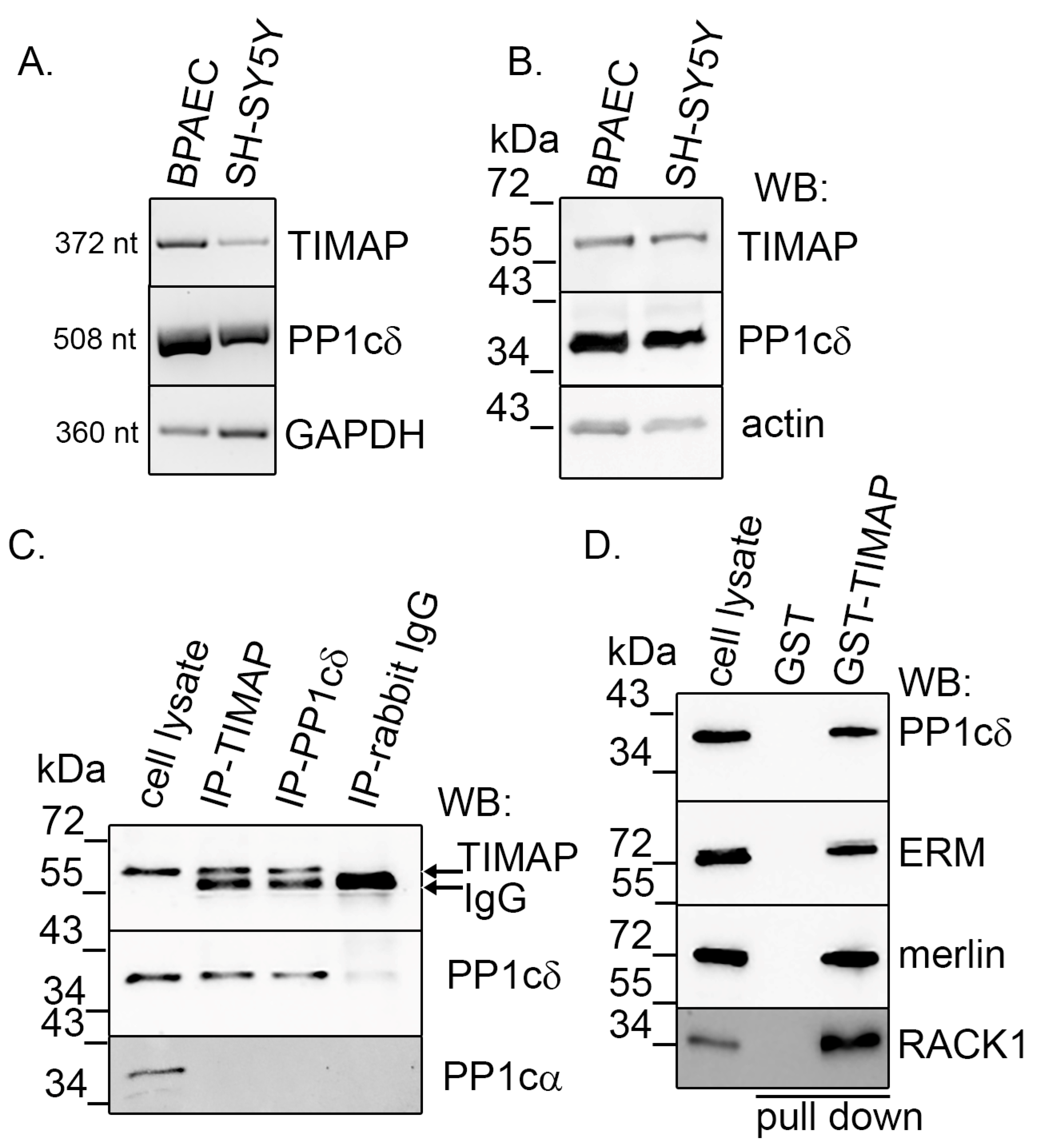
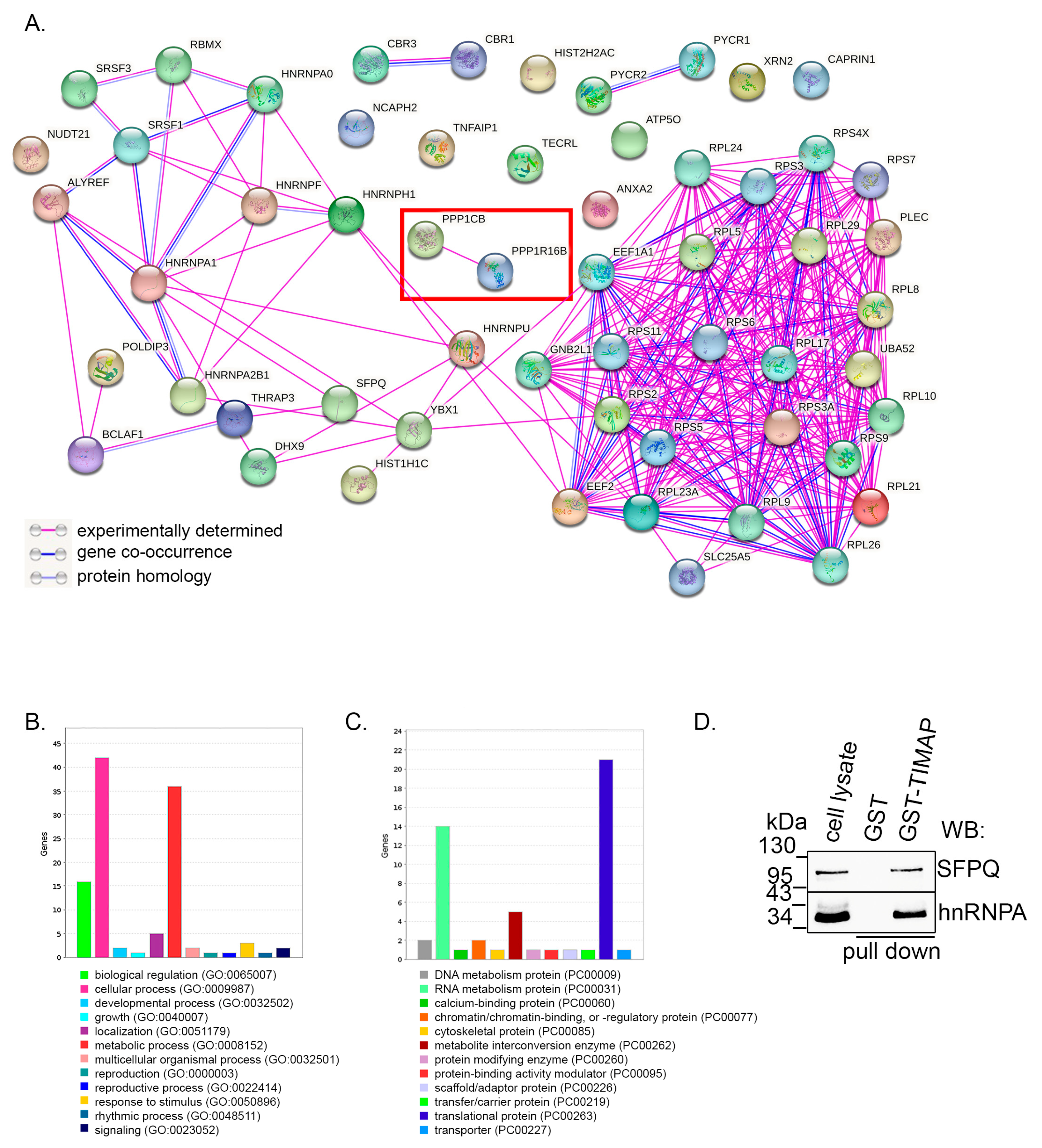
2.1. TIMAP–PP1c Complex Is Present in SH-SY5Y Cells
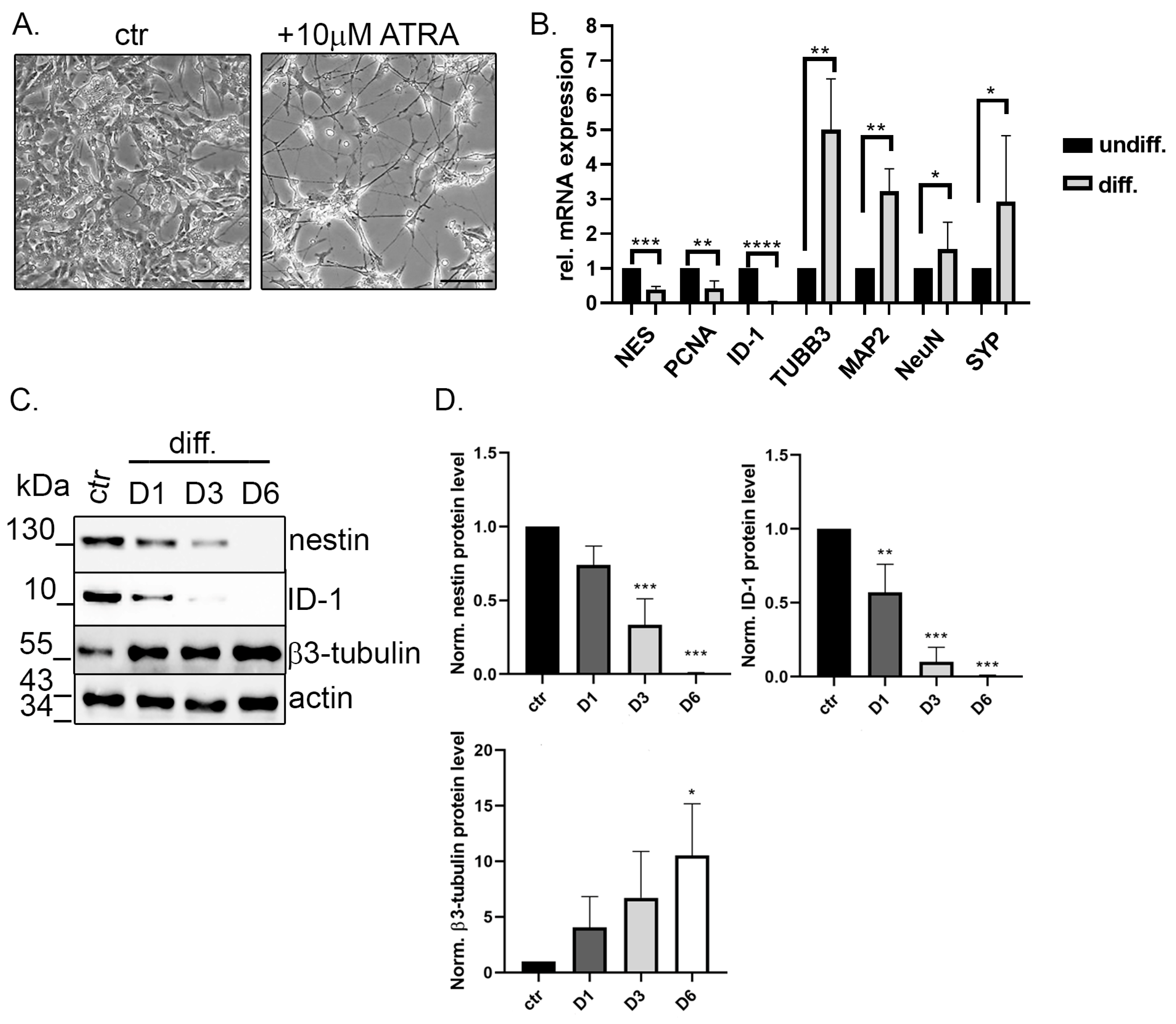
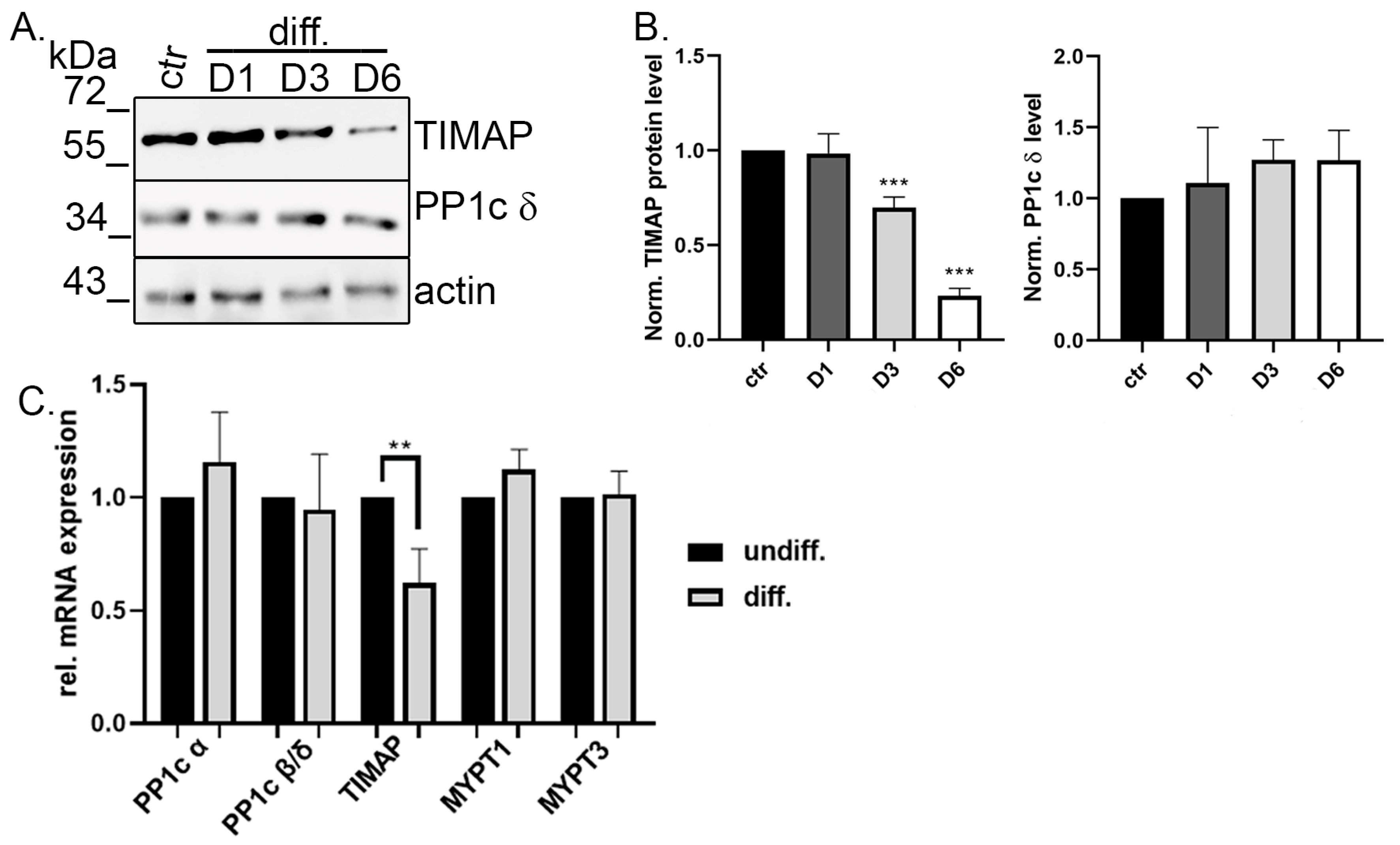
2.2. TIMAP Is Downregulated in Differentiated SH-SY5Y Cells
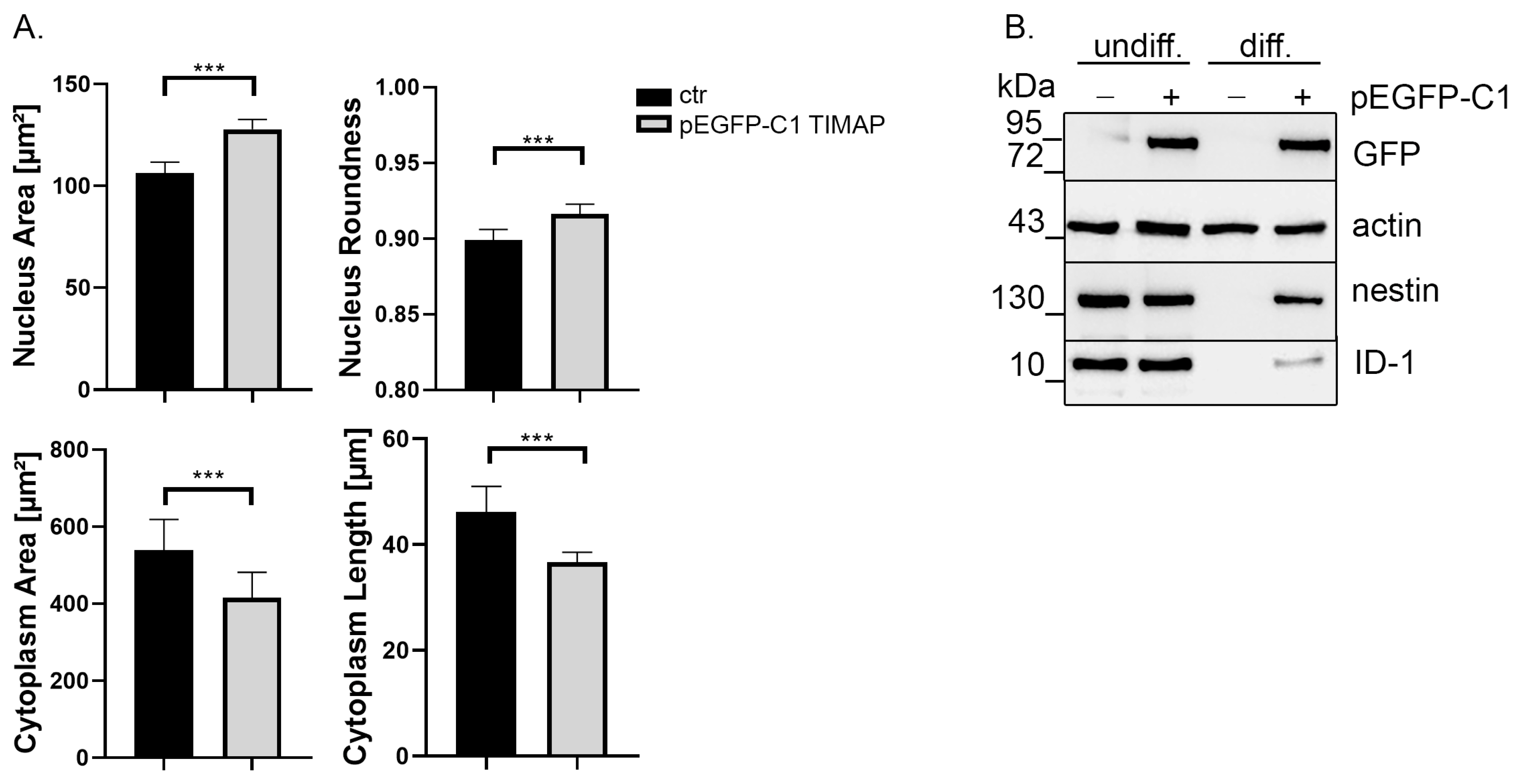
2.3. TIMAP Overexpression Delays Neuronal Differentiation
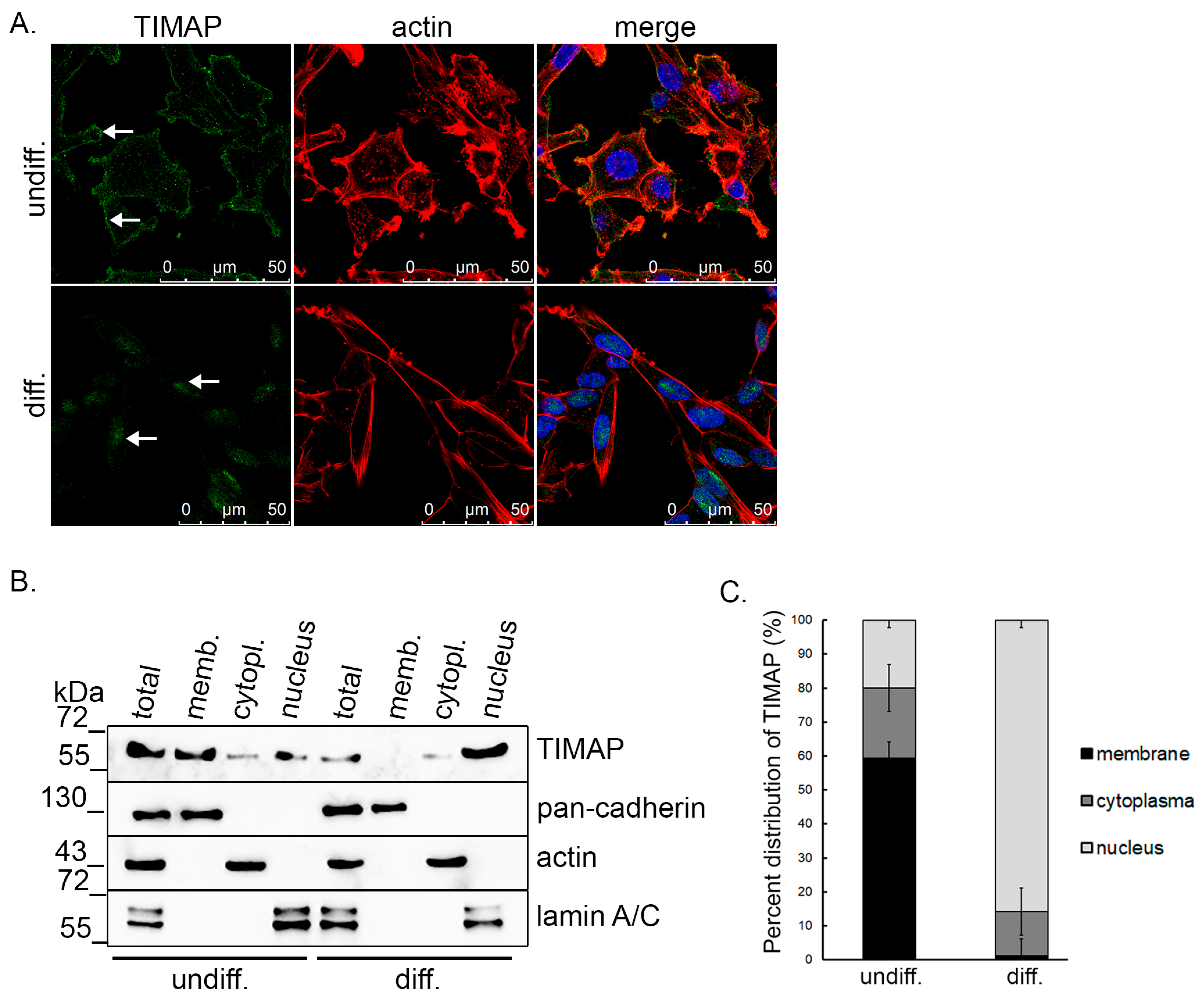
2.4. TIMAP Translocate into the Nucleus of SH-SY5Y Cells upon Differentiation
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Cell Differentiation
4.2. Bacterial Protein Expression and GST Pull-Down Assay, LC-MS/MS Analysis
4.3. SDS-PAGE and Western Blotting
4.4. RNA Isolation, cDNA Synthesis and qPCR
4.5. Immunoprecipitation, Immunofluorescence and Microscopy
4.6. Transfection
4.7. Subcellular Fractionation
4.8. High-Content Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barford, D. Protein phosphatases. Curr. Opin. Struct. Biol. 1995, 5, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Ariño, J.; Velázquez, D.; Casamayor, A. Ser/Thr protein phosphatases in fungi: Structure, regulation and function. Microb. Cell 2019, 6, 217–256. [Google Scholar] [CrossRef]
- Cohen, P.T.; Brewis, N.D.; Hughes, V.; Mann, D.J. Protein serine/threonine phosphatases; an expanding family. FEBS Lett. 1990, 268, 355–359. [Google Scholar] [CrossRef]
- Shi, Y. Serine/Threonine Phosphatases: Mechanism through Structure. Cell 2009, 139, 468–484. [Google Scholar] [CrossRef] [PubMed]
- Bollen, M.; Peti, W.; Ragusa, M.J.; Beullens, M. The extended PP1 toolkit: Designed to create specificity. Trends Biochem. Sci. 2010, 35, 450–458. [Google Scholar] [CrossRef]
- Cannon, J.F. Function of protein phosphatase-1, Glc7, in Saccharomyces cerevisiae. Adv. Appl. Microbiol. 2010, 73, 27–59. [Google Scholar]
- Peti, W.; Nairn, A.C.; Page, R. Structural basis for protein phosphatase 1 regulation and specificity. FEBS J. 2013, 280, 596–611. [Google Scholar] [CrossRef] [PubMed]
- Korrodi-Gregorio, L.; Esteves, S.L.; Fardilha, M. Protein phosphatase 1 catalytic isoforms: Specificity toward interacting proteins. Transl. Res. 2014, 164, 366–391. [Google Scholar] [CrossRef] [PubMed]
- Ceulemans, H.; Bollen, M. Functional diversity of protein phosphatase-1, a cellular economizer and reset button. Physiol. Rev. 2004, 84, 1–39. [Google Scholar] [CrossRef]
- Gibbons, J.A.; Kozubowski, L.; Tatchell, K.; Shenolikar, S. Expression of human protein phosphatase-1 in Saccharomyces cerevisiae highlights the role of phosphatase isoforms in regulating eukaryotic functions. J. Biol. Chem. 2007, 282, 21838–21847. [Google Scholar] [CrossRef]
- Verbinnen, I.; Ferreira, M.; Bollen, M. Biogenesis and activity regulation of protein phosphatase 1. Biochem. Soc. Trans. 2017, 45, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Csortos, C.; Czikora, I.; Bogatcheva, N.V.; Adyshev, D.M.; Poirier, C.; Olah, G.; Verin, A.D. TIMAP is a positive regulator of pulmonary endothelial barrier function. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, L440–L450. [Google Scholar] [CrossRef] [PubMed]
- Boratko, A.; Csortos, C. TIMAP, the versatile protein phosphatase 1 regulator in endothelial cells. IUBMB Life 2017, 69, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Mattagajasingh, S.N.; Xu, H.; Kim, K.; Fierlbeck, W.; Deng, J.; Lowenstein, C.J.; Ballermann, B.J. TIMAP, a novel CAAX box protein regulated by TGF-beta1 and expressed in endothelial cells. Am. J. Physiol. Cell Physiol. 2002, 283, C327–C337. [Google Scholar] [CrossRef] [PubMed]
- Czikora, I.; Kim, K.M.; Kasa, A.; Becsi, B.; Verin, A.D.; Gergely, P.; Erdodi, F.; Csortos, C. Characterization of the effect of TIMAP phosphorylation on its interaction with protein phosphatase 1. Biochimie 2011, 93, 1139–1145. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Boratko, A.; Csortos, C. PKC mediated phosphorylation of TIMAP regulates PP1c activity and endothelial barrier function. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 431–439. [Google Scholar] [CrossRef]
- Schulz, A.; Zoch, A.; Morrison, H. A neuronal function of the tumor suppressor protein merlin. Acta Neuropathol. Commun. 2014, 2, 82. [Google Scholar] [CrossRef][Green Version]
- Magdaleno, S.; Northcutt, G.M.; Curran, T.; Kurschner, C. mPPP1R16B is a novel mouse protein phosphatase 1 targeting subunit whose mRNA is located in cell bodies and dendrites of neurons in four distinct regions of the brain. Brain Res. Gene Expr. Patterns 2002, 1, 143–149. [Google Scholar] [CrossRef]
- Alivand, M.R.; Najafi, S.; Esmaeili, S.; Rahmanpour, D.; Zhaleh, H.; Rahmati, Y. Integrative analysis of DNA methylation and gene expression profiles to identify biomarkers of glioblastoma. Cancer Genet. 2021, 258, 135–150. [Google Scholar] [CrossRef]
- Bell, M.; Zempel, H. SH-SY5Y-derived neurons: A human neuronal model system for investigating TAU sorting and neuronal subtype-specific TAU vulnerability. Rev. Neurosci. 2021, 33, 1–15. [Google Scholar] [CrossRef]
- Xicoy, H.; Wieringa, B.; Martens, G.J. The SH-SY5Y cell line in Parkinson’s disease research: A systematic review. Mol. Neurodegener. 2017, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Strother, L.; Miles, G.B.; Holiday, A.R.; Cheng, Y.; Doherty, G.H. Long-term culture of SH-SY5Y neuroblastoma cells in the absence of neurotrophins: A novel model of neuronal ageing. J. Neurosci. Methods 2021, 362, 109301. [Google Scholar] [CrossRef]
- De Conto, V.; Cheung, V.; Maubon, G.; Souguir, Z.; Maubon, N.; Vandenhaute, E.; Berezowski, V. In vitro differentiation modifies the neurotoxic response of SH-SY5Y cells. Toxicol. In Vitro 2021, 77, 105235. [Google Scholar] [CrossRef] [PubMed]
- Christensen, J.; Steain, M.; Slobedman, B.; Abendroth, A. Differentiated neuroblastoma cells provide a highly efficient model for studies of productive varicella-zoster virus infection of neuronal cells. J. Virol. 2011, 85, 8436–8442. [Google Scholar] [CrossRef]
- Shipley, M.M.; Mangold, C.A.; Szpara, M.L. Differentiation of the SH-SY5Y Human Neuroblastoma Cell Line. J. Vis. Exp. 2016, 108, 53193. [Google Scholar]
- Lopes, F.M.; Schroder, R.; da Frota, M.L., Jr.; Zanotto-Filho, A.; Muller, C.B.; Pires, A.S.; Meurer, R.T.; Colpo, G.D.; Gelain, D.P.; Kapczinski, F.; et al. Comparison between proliferative and neuron-like SH-SY5Y cells as an in vitro model for Parkinson disease studies. Brain Res. 2010, 1337, 85–94. [Google Scholar] [CrossRef]
- Campos Cogo, S.; Gradowski Farias da Costa do Nascimento, T.; de Almeida Brehm Pinhatti, F.; de Franca Junior, N.; Santos Rodrigues, B.; Cavalli, L.R.; Elifio-Esposito, S. An overview of neuroblastoma cell lineage phenotypes and in vitro models. Exp. Biol. Med. 2020, 245, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
- Boratko, A.; Gergely, P.; Csortos, C. RACK1 is involved in endothelial barrier regulation via its two novel interacting partners. Cell Commun. Signal. 2013, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Kiraly, N.; Thalwieser, Z.; Fonodi, M.; Csortos, C.; Boratko, A. Dephosphorylation of annexin A2 by protein phosphatase 1 regulates endothelial cell barrier. IUBMB Life 2021, 73, 1257–1268. [Google Scholar] [CrossRef]
- Boratko, A.; Peter, M.; Thalwieser, Z.; Kovacs, E.; Csortos, C. Elongation factor-1A1 is a novel substrate of the protein phosphatase 1-TIMAP complex. Int. J. Biochem. Cell Biol. 2015, 69, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Khakha, N.; Khan, H.; Kaur, A.; Singh, T.G. Therapeutic implications of phosphorylation- and dephosphorylation-dependent factors of cAMP-response element-binding protein (CREB) in neurodegeneration. Pharmacol. Rep. 2023, 75, 1152–1165. [Google Scholar] [CrossRef]
- Brembati, V.; Faustini, G.; Longhena, F.; Outeiro, T.F.; Bellucci, A. Changes in alpha-Synuclein Posttranslational Modifications in an AAV-Based Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2023, 24, 13435. [Google Scholar] [CrossRef]
- Li, Z.; Yin, B.; Zhang, S.; Lan, Z.; Zhang, L. Targeting protein kinases for the treatment of Alzheimer’s disease: Recent progress and future perspectives. Eur. J. Med. Chem. 2023, 261, 115817. [Google Scholar] [CrossRef]
- Da Cruz e Silva, E.F.; Fox, C.A.; Ouimet, C.C.; Gustafson, E.; Watson, S.J.; Greengard, P. Differential expression of protein phosphatase 1 isoforms in mammalian brain. J. Neurosci. 1995, 15, 3375–3389. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Otey, C.A.; Boukhelifa, M.; Maness, P. B35 neuroblastoma cells: An easily transfected, cultured cell model of central nervous system neurons. Methods Cell Biol. 2003, 71, 287–304. [Google Scholar] [PubMed]
- Shafer, T.J.; Atchison, W.D. Transmitter, ion channel and receptor properties of pheochromocytoma (PC12) cells: A model for neurotoxicological studies. Neurotoxicology 1991, 12, 473–492. [Google Scholar] [PubMed]
- LePage, K.T.; Dickey, R.W.; Gerwick, W.H.; Jester, E.L.; Murray, T.F. On the use of neuro-2a neuroblastoma cells versus intact neurons in primary culture for neurotoxicity studies. Crit. Rev. Neurobiol. 2005, 17, 27–50. [Google Scholar] [CrossRef] [PubMed]
- Biedler, J.L.; Helson, L.; Spengler, B.A. Morphology and growth, tumorigenicity, and cytogenetics of human neuroblastoma cells in continuous culture. Cancer Res. 1973, 33, 2643–2652. [Google Scholar]
- Shopik, M.J.; Li, L.; Luu, H.A.; Obeidat, M.; Holmes, C.F.; Ballermann, B.J. Multi-directional function of the protein phosphatase 1 regulatory subunit TIMAP. Biochem. Biophys. Res. Commun. 2013, 435, 567–573. [Google Scholar] [CrossRef]
- Singh, J.; Kaur, G. Transcriptional regulation of polysialylated neural cell adhesion molecule expression by NMDA receptor activation in retinoic acid-differentiated SH-SY5Y neuroblastoma cultures. Brain Res. 2007, 1154, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Kume, T.; Kawato, Y.; Osakada, F.; Izumi, Y.; Katsuki, H.; Nakagawa, T.; Kaneko, S.; Niidome, T.; Takada-Takatori, Y.; Akaike, A. Dibutyryl cyclic AMP induces differentiation of human neuroblastoma SH-SY5Y cells into a noradrenergic phenotype. Neurosci. Lett. 2008, 443, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Cernaianu, G.; Brandmaier, P.; Scholz, G.; Ackermann, O.P.; Alt, R.; Rothe, K.; Cross, M.; Witzigmann, H.; Trobs, R.B. All-trans retinoic acid arrests neuroblastoma cells in a dormant state. Subsequent nerve growth factor/brain-derived neurotrophic factor treatment adds modest benefit. J. Pediatr. Surg. 2008, 43, 1284–1294. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.V.; Buensuceso, C.; Montague, F.; Mahadevan, L. Vanadate stimulates differentiation and neurite outgrowth in rat pheochromocytoma PC12 cells and neurite extension in human neuroblastoma SH-SY5Y cells. Neuroscience 1994, 60, 479–494. [Google Scholar] [CrossRef] [PubMed]
- Oe, T.; Sasayama, T.; Nagashima, T.; Muramoto, M.; Yamazaki, T.; Morikawa, N.; Okitsu, O.; Nishimura, S.; Aoki, T.; Katayama, Y.; et al. Differences in gene expression profile among SH-SY5Y neuroblastoma subclones with different neurite outgrowth responses to nerve growth factor. J. Neurochem. 2005, 94, 1264–1276. [Google Scholar] [CrossRef]
- Pahlman, S.; Odelstad, L.; Larsson, E.; Grotte, G.; Nilsson, K. Phenotypic changes of human neuroblastoma cells in culture induced by 12-O-tetradecanoyl-phorbol-13-acetate. Int. J. Cancer 1981, 28, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Sarkanen, J.R.; Nykky, J.; Siikanen, J.; Selinummi, J.; Ylikomi, T.; Jalonen, T.O. Cholesterol supports the retinoic acid-induced synaptic vesicle formation in differentiating human SH-SY5Y neuroblastoma cells. J. Neurochem. 2007, 102, 1941–1952. [Google Scholar] [CrossRef] [PubMed]
- Agholme, L.; Lindstrom, T.; Kagedal, K.; Marcusson, J.; Hallbeck, M. An in vitro model for neuroscience: Differentiation of SH-SY5Y cells into cells with morphological and biochemical characteristics of mature neurons. J. Alzheimers Dis. 2010, 20, 1069–1082. [Google Scholar] [CrossRef]
- Clagett-Dame, M.; McNeill, E.M.; Muley, P.D. Role of all-trans retinoic acid in neurite outgrowth and axonal elongation. J. Neurobiol. 2006, 66, 739–756. [Google Scholar] [CrossRef]
- Wang, X.; Obeidat, M.; Li, L.; Pasarj, P.; Aburahess, S.; Holmes, C.F.B.; Ballermann, B.J. TIMAP inhibits endothelial myosin light chain phosphatase by competing with MYPT1 for the catalytic protein phosphatase 1 subunit PP1cbeta. J. Biol. Chem. 2019, 294, 13280–13291. [Google Scholar] [CrossRef]
- Lontay, B.; Serfozo, Z.; Gergely, P.; Ito, M.; Hartshorne, D.J.; Erdodi, F. Localization of myosin phosphatase target subunit 1 in rat brain and in primary cultures of neuronal cells. J. Comp. Neurol. 2004, 478, 72–87. [Google Scholar] [CrossRef]
- Beckelman, B.C.; Day, S.; Zhou, X.; Donohue, M.; Gouras, G.K.; Klann, E.; Keene, C.D.; Ma, T. Dysregulation of Elongation Factor 1A Expression is Correlated with Synaptic Plasticity Impairments in Alzheimer’s Disease. J. Alzheimers Dis. 2016, 54, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Beckelman, B.C.; Zhou, X.; Keene, C.D.; Ma, T. Impaired Eukaryotic Elongation Factor 1A Expression in Alzheimer’s Disease. Neurodegener. Dis. 2016, 16, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Esparcia, P.; Hernandez-Ortega, K.; Koneti, A.; Gil, L.; Delgado-Morales, R.; Castano, E.; Carmona, M.; Ferrer, I. Altered machinery of protein synthesis is region- and stage-dependent and is associated with alpha-synuclein oligomers in Parkinson’s disease. Acta Neuropathol. Commun. 2015, 3, 76. [Google Scholar] [CrossRef] [PubMed]
- Kapur, M.; Monaghan, C.E.; Ackerman, S.L. Regulation of mRNA Translation in Neurons-A Matter of Life and Death. Neuron 2017, 96, 616–637. [Google Scholar] [CrossRef]
- Clarke, J.P.; Thibault, P.A.; Salapa, H.E.; Levin, M.C. A Comprehensive Analysis of the Role of hnRNP A1 Function and Dysfunction in the Pathogenesis of Neurodegenerative Disease. Front. Mol. Biosci. 2021, 8, 659610. [Google Scholar] [CrossRef]
- Roy, R.; Durie, D.; Li, H.; Liu, B.Q.; Skehel, J.M.; Mauri, F.; Cuorvo, L.V.; Barbareschi, M.; Guo, L.; Holcik, M.; et al. hnRNPA1 couples nuclear export and translation of specific mRNAs downstream of FGF-2/S6K2 signalling. Nucleic Acids Res. 2014, 42, 12483–12497. [Google Scholar] [CrossRef]
- Choi, Y.H.; Lim, J.K.; Jeong, M.W.; Kim, K.T. HnRNP A1 phosphorylated by VRK1 stimulates telomerase and its binding to telomeric DNA sequence. Nucleic Acids Res. 2012, 40, 8499–8518. [Google Scholar] [CrossRef]
- Koo, J.H.; Lee, H.J.; Kim, W.; Kim, S.G. Endoplasmic Reticulum Stress in Hepatic Stellate Cells Promotes Liver Fibrosis via PERK-Mediated Degradation of HNRNPA1 and Up-regulation of SMAD2. Gastroenterology 2016, 150, 181–193.e8. [Google Scholar] [CrossRef]
- Martin, J.; Masri, J.; Cloninger, C.; Holmes, B.; Artinian, N.; Funk, A.; Ruegg, T.; Anderson, L.; Bashir, T.; Bernath, A.; et al. Phosphomimetic substitution of heterogeneous nuclear ribonucleoprotein A1 at serine 199 abolishes AKT-dependent internal ribosome entry site-transacting factor (ITAF) function via effects on strand annealing and results in mammalian target of rapamycin complex 1 (mTORC1) inhibitor sensitivity. J. Biol. Chem. 2011, 286, 16402–16413. [Google Scholar]
- Lim, Y.W.; James, D.; Huang, J.; Lee, M. The Emerging Role of the RNA-Binding Protein SFPQ in Neuronal Function and Neurodegeneration. Int. J. Mol. Sci. 2020, 21, 7151. [Google Scholar] [CrossRef]
- Sasaki, Y.T.; Ideue, T.; Sano, M.; Mituyama, T.; Hirose, T. MENepsilon/beta noncoding RNAs are essential for structural integrity of nuclear paraspeckles. Proc. Natl. Acad. Sci. USA 2009, 106, 2525–2530. [Google Scholar] [CrossRef]
- Ha, K.; Takeda, Y.; Dynan, W.S. Sequences in PSF/SFPQ mediate radioresistance and recruitment of PSF/SFPQ-containing complexes to DNA damage sites in human cells. DNA Repair 2011, 10, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Kim, Y.C.; Adelstein, R.S.; Kawamoto, S. Fox-3 and PSF interact to activate neural cell-specific alternative splicing. Nucleic Acids Res. 2011, 39, 3064–3078. [Google Scholar] [CrossRef]
- Heyd, F.; Lynch, K.W. Phosphorylation-dependent regulation of PSF by GSK3 controls CD45 alternative splicing. Mol. Cell 2010, 40, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, R.; Subramanian, V. PA6 Stromal Cell Co-Culture Enhances SH-SY5Y and VSC4.1 Neuroblastoma Differentiation to Mature Phenotypes. PLoS ONE 2016, 11, e0159051. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Regdon, Z.; Demeny, M.A.; Kovacs, K.; Hajnady, Z.; Nagy-Penzes, M.; Bakondi, E.; Kiss, A.; Hegedus, C.; Virag, L. High-content screening identifies inhibitors of oxidative stress-induced parthanatos: Cytoprotective and anti-inflammatory effects of ciclopirox. Br. J. Pharmacol. 2021, 178, 1095–1113. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID (UniProt) | Protein Name | Nr. of Peptide | Gene Name |
|---|---|---|---|
| Q15149 | Plectin | 290 | PLEC |
| Q08211 | ATP-dependent RNA helicase A | 61 | DHX9 |
| Q9H0D6 | 5-3 exoribonuclease 2 | 47 | XRN2 |
| P13639 | Elongation factor 2 | 45 | EEF2 |
| Q9Y2W1 | Thyroid-hormone-receptor-associated protein 3 | 42 | THRAP3 |
| Q00839 | Heterogeneous nuclear ribonucleoprotein U | 37 | HNRNPU |
| Q9NYF8 | Bcl-2-associated transcription factor 1 | 35 | BCLAF1 |
| P23246 | Splicing factor, proline- and glutamine-rich | 30 | SFPQ |
| Q6IBW4 | Condensin-2 complex subunit H2 | 26 | NCAPH2 |
| P38159 | RNA-binding motif protein | 25 | RBMX |
| Q96T49 | Protein phosphatase 1 regulatory inhibitor subunit 16B | 24 | PPP1R16B |
| Q9BY77 | Polymerase delta-interacting protein 3 | 24 | POLDIP3 |
| P07355 | Annexin A2 | 23 | ANXA2 |
| Q5VTE0 | Elongation factor 1-alpha 1 | 21 | EEF1A1 |
| Q13829 | BTB/POZ domain-containing adapter for CUL3-mediated RhoA degradation protein 2 | 21 | TNFAIP1 |
| Q14444 | Caprin-1 | 21 | CAPRIN1 |
| P22626 | Heterogeneous nuclear ribonucleoproteins A2/B1 | 20 | HNRNPA2B1 |
| P63244 | Guanine nucleotide-binding protein subunit beta-2-like 1 | 20 | GNB2L1 |
| P05141 | ADP/ATP translocase 2 | 19 | SLC25A5 |
| P23396 | 40S ribosomal protein S3 | 19 | RPS3 |
| P09651 | Heterogeneous nuclear ribonucleoprotein A1 | 18 | HNRNPA1 |
| P15880 | 40S ribosomal protein S2 | 18 | RPS2 |
| P31943 | Heterogeneous nuclear ribonucleoprotein H | 18 | HNRNPH1 |
| P16152 | Carbonyl reductase [NADPH] 1 | 17 | CBR1 |
| P61247 | 40S ribosomal protein S3a | 17 | RPS3A |
| P62140 | Serine/threonine-protein phosphatase PP1-beta catalytic subunit | 17 | PPP1CB |
| O75828 | Carbonyl reductase [NADPH] 3 | 16 | CBR3 |
| P62701 | 40S ribosomal protein S4, X isoform | 16 | RPS4X |
| Q96C36 | Pyrroline-5-carboxylate reductase 2 | 16 | PYCR2 |
| P32322 | Pyrroline-5-carboxylate reductase 1, mitochondrial | 15 | PYCR1 |
| P52597 | Heterogeneous nuclear ribonucleoprotein F | 15 | HNRNPF |
| O43809 | Cleavage and polyadenylation specificity factor subunit 5 | 14 | NUDT21 |
| P67809 | Nuclease-sensitive element-binding protein 1 | 13 | YBX1 |
| Q07955 | Serine/arginine-rich splicing factor 1 | 13 | SRSF1 |
| Q13151 | Heterogeneous nuclear ribonucleoprotein A0 | 13 | HNRNPA0 |
| Q86V81 | THO complex subunit 4 | 13 | ALYREF |
| P48047 | ATP synthase subunit O, mitochondrial | 12 | ATP5O |
| Q5HYJ1 | Trans-2,3-enoyl-CoA reductase-like | 12 | TECRL |
| P46777 | 60S ribosomal protein L5 | 11 | RPL5 |
| P46781 | 40S ribosomal protein S9 | 11 | RPS9 |
| P46782 | 40S ribosomal protein S5 | 10 | RPS5 |
| P62081 | 40S ribosomal protein S7 | 10 | RPS7 |
| P62280 | 40S ribosomal protein S11 | 10 | RPS11 |
| P62917 | 60S ribosomal protein L8 | 10 | RPL8 |
| P16403 | Histone H1.2 | 9 | HIST1H1C |
| P18621 | 60S ribosomal protein L17 | 9 | RPL17 |
| P32969 | 60S ribosomal protein L9 | 9 | RPL9 |
| P27635 | 60S ribosomal protein L10 | 8 | RPL10 |
| P62753 | 40S ribosomal protein S6 | 8 | RPS6 |
| P83731 | 60S ribosomal protein L24 | 8 | RPL24 |
| P84103 | Serine/arginine-rich splicing factor 3 | 7 | SRSF3 |
| P62987 | Ubiquitin-60S ribosomal protein L40 | 6 | UBA52 |
| P46778 | 60S ribosomal protein L21 | 6 | RPL21 |
| P61254 | 60S ribosomal protein L26;60S ribosomal protein L26-like 1 | 6 | RPL26 |
| P62750 | 60S ribosomal protein L23a | 6 | RPL23A |
| Q16777 | Histone H2A type 2-C | 5 | HIST2H2AC |
| P47914 | 60S ribosomal protein L29 | 4 | RPL29 |
| Antibody | Dilution | Vendor (cat. #) |
|---|---|---|
| TIMAP | 1:1000 | ABGENT (San Diego, CA, USA)(AP17590c) |
| PP1cδ | 1:1000 | Millipore (Burlington, MA, USA) (#07-270) |
| PP1cα | 1:1000 | R&D Systems (Minneapolis, MN, USA) (MAB6105) |
| actin | 1:1000 | Sigma (St. Louis, MO, USA) (A5060) |
| ERM | 1:1000 | Cell Signaling Technologies (Danvers, MA, USA) (#3142) |
| merlin | 1:1000 | Cell Signaling Technologies (#6995) |
| RACK1 | 1:1000 | BD Transduction Laboratories (Franklin Lakes, NJ, USA) (#610177) |
| GFP | 1:1000 | Millipore (#AB10145; #MAB1083) |
| Nestin | 1:2000 | Cell Signaling Technologies (#4760) |
| ID-1 | 1:500 | Novus Biologicals (Littleton, CO, USA) (#H00003397-M02) |
| lamin A/C | 1:2000 | Santa Cruz (Dallas, TX, USA) (sc-20681) |
| pan-cadherin | 1:1000 | Cell Signaling Technologies (#4068) |
| HRP-linked anti-rabbit IgG | 1:5000 | Cell Signaling Technologies (#7074) |
| HRP-linked anti-mouse IgG | 1:5000 | Cell Signaling Technologies (#7076) |
| Gene Name | Primer 1 | Primer 2 |
|---|---|---|
| GAPDH (glyceraldehyde-3-phosphate dehydrogenase) | 5′-GTC TCC TCT GAC TTC AAC AGC G-3′ | 5′-ACC ACC CTG TTG CTG TAG CCA A-3′ |
| NES (nestin) | 5′-TCA AGA TGT CCC TCA GCC TGG A-3′ | 5′-AAG CTG AGG GAA GTC TTG GAG C-3′ |
| PCNA (proliferating cell nuclear antigen) | 5′-CAA GTA ATG TCG ATA AAG AGG AGG-3′ | 5′-GTG TCA CCG TTG AAG AGA GTG G-3′ |
| ID1 (Inhibitor of differentiation 1) | 5′-GTT GGA GCT GAA CTC GGA ATC C-3′ | 5′-ACA CAA GAT GCG ATC GTC CGC A-3′ |
| TUBB3 (β3-tubulin) | 5′-TCA GCG TCT ACT ACA ACG AGG C-3′ | 5′-GCC TGA AGA GAT GTC CAA AGG C-3′ |
| MAP2 (microtubule-associated protein 2) | 5′-AGG CTG TAG CAG TCC TGA AAG G-3′ | 5′-CTT CCT CCA CTG TGA CAG TCT G-3′ |
| NeuN (RBFOX3) | 5′-TAC GCA GCC TAC AGA TAC GCT C-3′ | 5′-TGG TTC CAA TGC TGT AGG TCG C-3′ |
| SYP (synaptophysin) | 5′-TCG GCT TTG TGA AGG TGC TGC A-3′ | 5′-TCA CTC TCG GTC TTG TTG GCA C-3′ |
| ACTB (b-actin) | 5′-CAC CAT TGG CAA TGA GCG GTT C-3′ | 5′-AGG TCT TTG CGG ATG TCC ACG T-3′ |
| PPP1CA (PP1c α) | 5′-GCT GCT GGC CTA TAA GAT CAA-3′ | 5′-GTC TCT TGC ACT CAT CGT AGA A-3′ |
| PPP1CB (PP1c δ) | 5′-CAG CCA TTG TGG ATG AGA AGA-3′ | 5′-AGG GAC ATC AGT AGG TCT CAT AA-3′ |
| PPP1R16B (TIMAP) | 5′-TGG AGC TAG TCT CAG TGC AAG G-3′ | 5′-CTC AAG GAT GAC TTG TGC CTC AG-3′ |
| PPP1R12A (MYPT1) | 5′-CAG GTC AAG CAA CAC CTA CA-3′ | 5′-GCC CGT CTT TCT AAG TCC TAA C-3′ |
| PPP1R16A (MYPT3) | 5′-TCA ATG CCT GTG ACA GTG AG-3′ | 5′-ATG TTC CCG TCG GTG TTG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonódi, M.; Thalwieser, Z.; Csortos, C.; Boratkó, A. TIMAP, a Regulatory Subunit of Protein Phosphatase 1, Inhibits In Vitro Neuronal Differentiation. Int. J. Mol. Sci. 2023, 24, 17360. https://doi.org/10.3390/ijms242417360
Fonódi M, Thalwieser Z, Csortos C, Boratkó A. TIMAP, a Regulatory Subunit of Protein Phosphatase 1, Inhibits In Vitro Neuronal Differentiation. International Journal of Molecular Sciences. 2023; 24(24):17360. https://doi.org/10.3390/ijms242417360
Chicago/Turabian StyleFonódi, Márton, Zsófia Thalwieser, Csilla Csortos, and Anita Boratkó. 2023. "TIMAP, a Regulatory Subunit of Protein Phosphatase 1, Inhibits In Vitro Neuronal Differentiation" International Journal of Molecular Sciences 24, no. 24: 17360. https://doi.org/10.3390/ijms242417360
APA StyleFonódi, M., Thalwieser, Z., Csortos, C., & Boratkó, A. (2023). TIMAP, a Regulatory Subunit of Protein Phosphatase 1, Inhibits In Vitro Neuronal Differentiation. International Journal of Molecular Sciences, 24(24), 17360. https://doi.org/10.3390/ijms242417360