

Optimization of Xylooligosaccharides Production by Native and Recombinant Xylanase Hydrolysis of Chicken Feed Substrates


Abstract
:1. Introduction
2. Results and Discussion
2.1. Optimization of Feed Hydrolysis for Enhanced Reducing Sugars
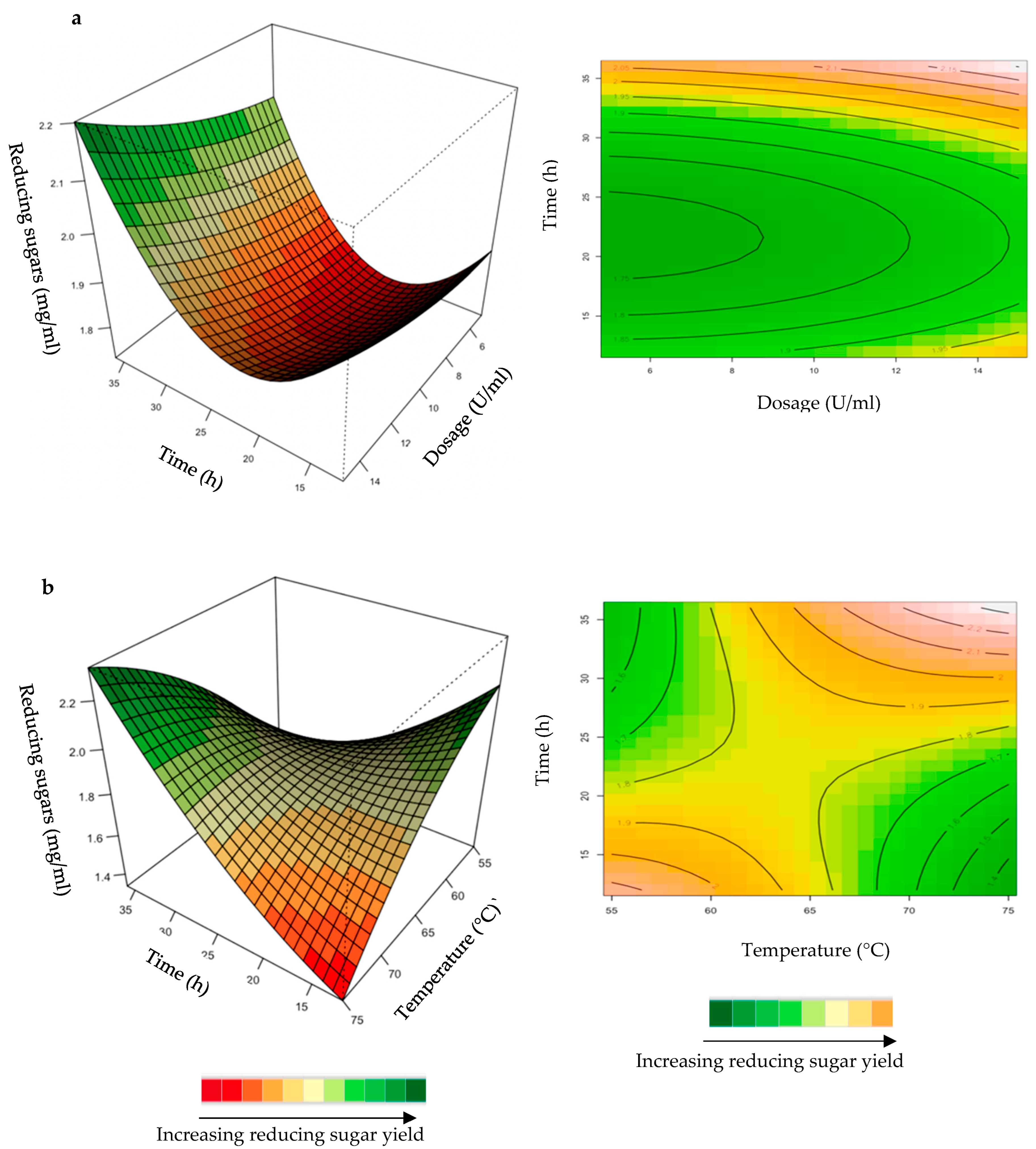
2.1.1. Hydrolysis of Starter and Grower Feeds with Crude T. harzianum Xylanase
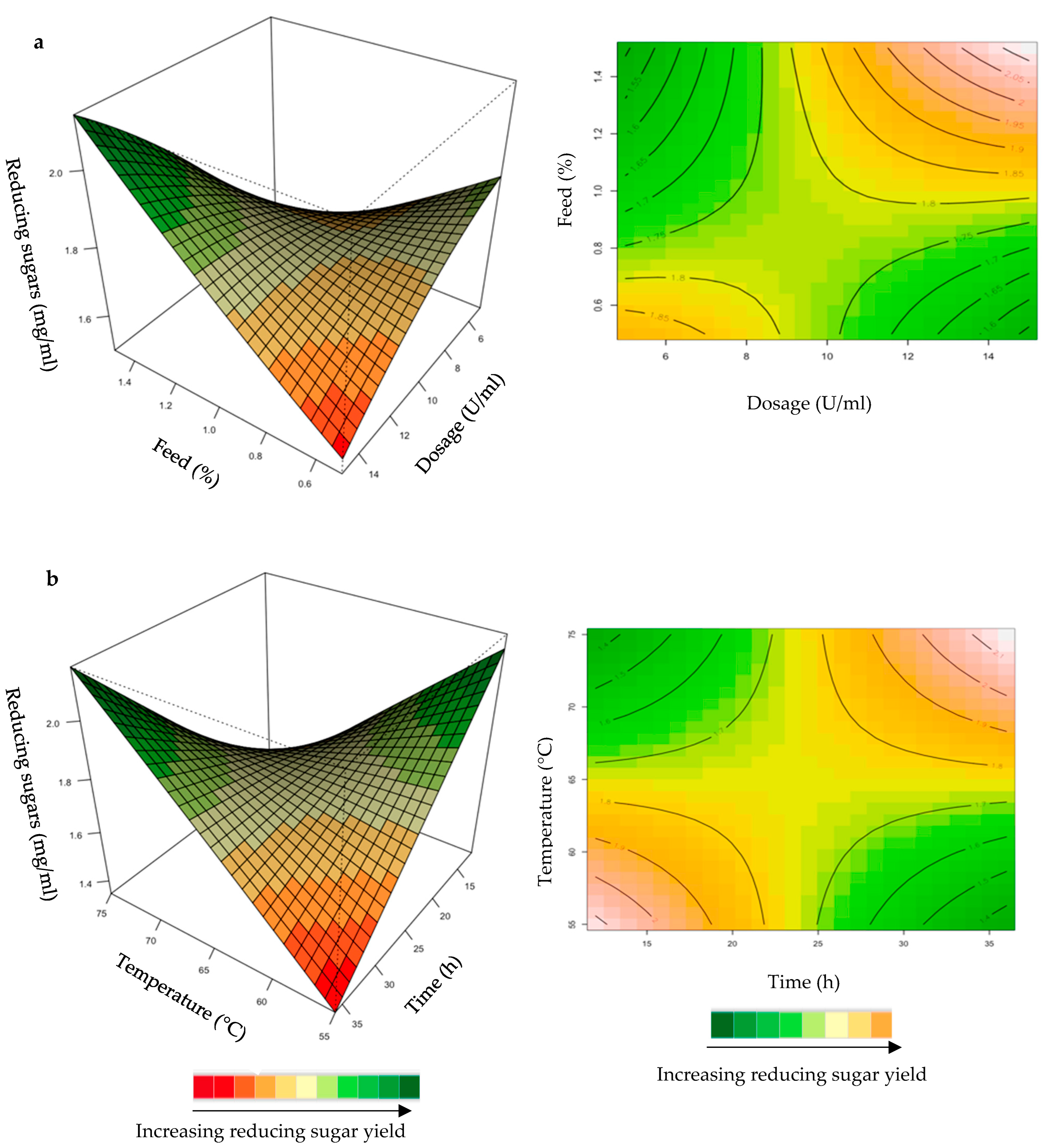
2.1.2. Hydrolysis of Starter and Grower Feeds with Purified T. harzianum Xylanase
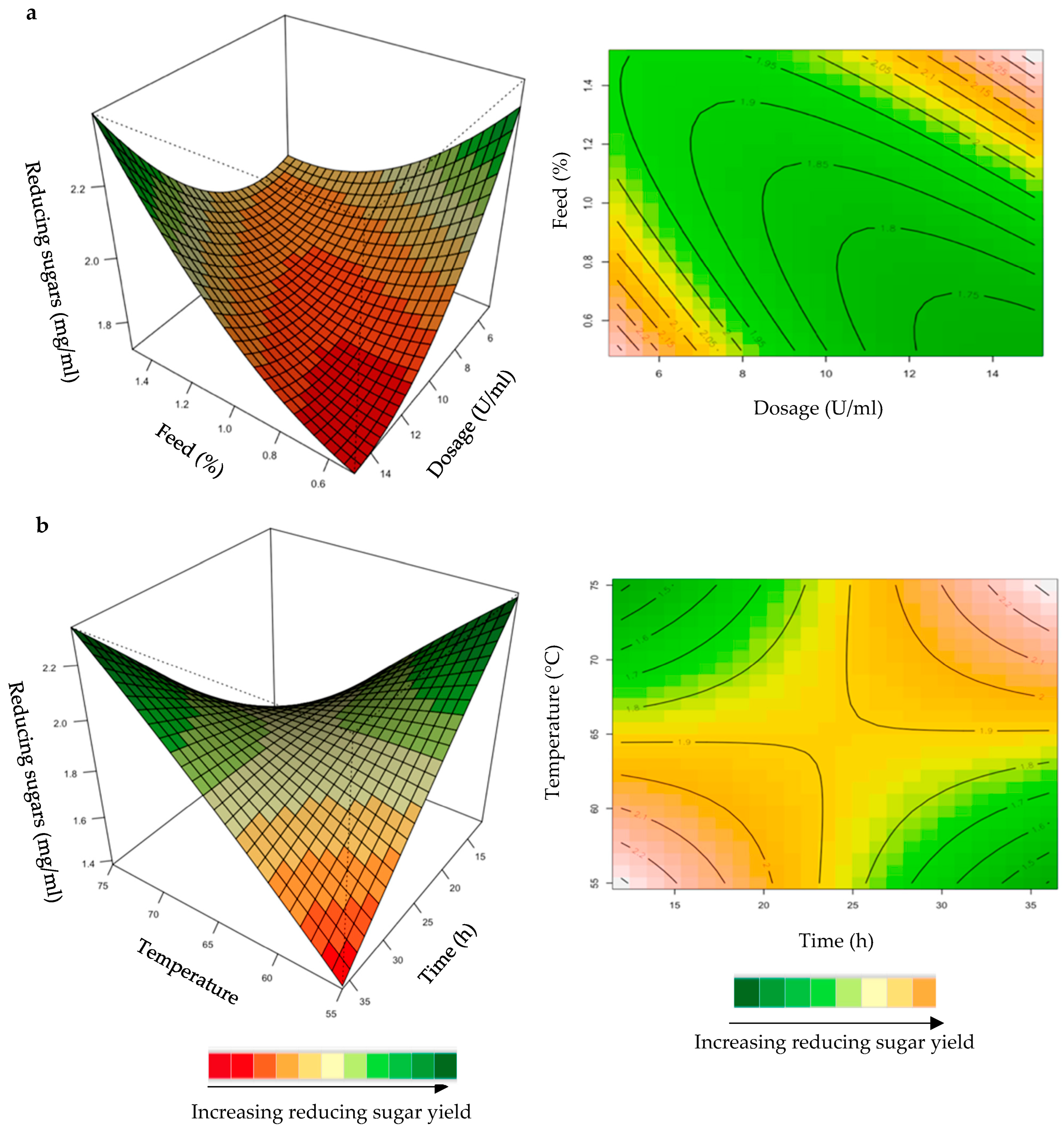
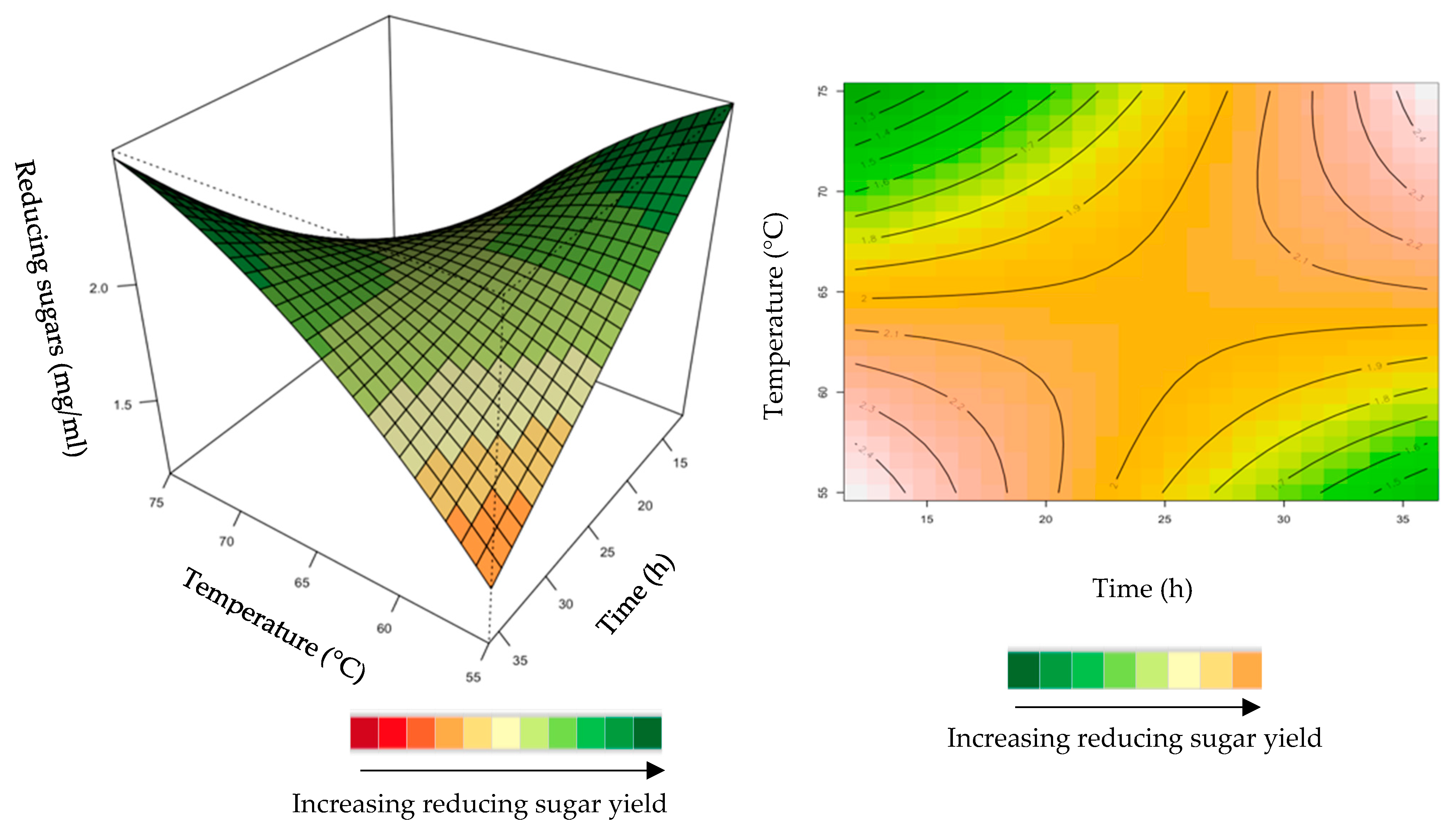
2.1.3. Hydrolysis of Starter and Grower Feeds with Recombinant XT6 Xylanase
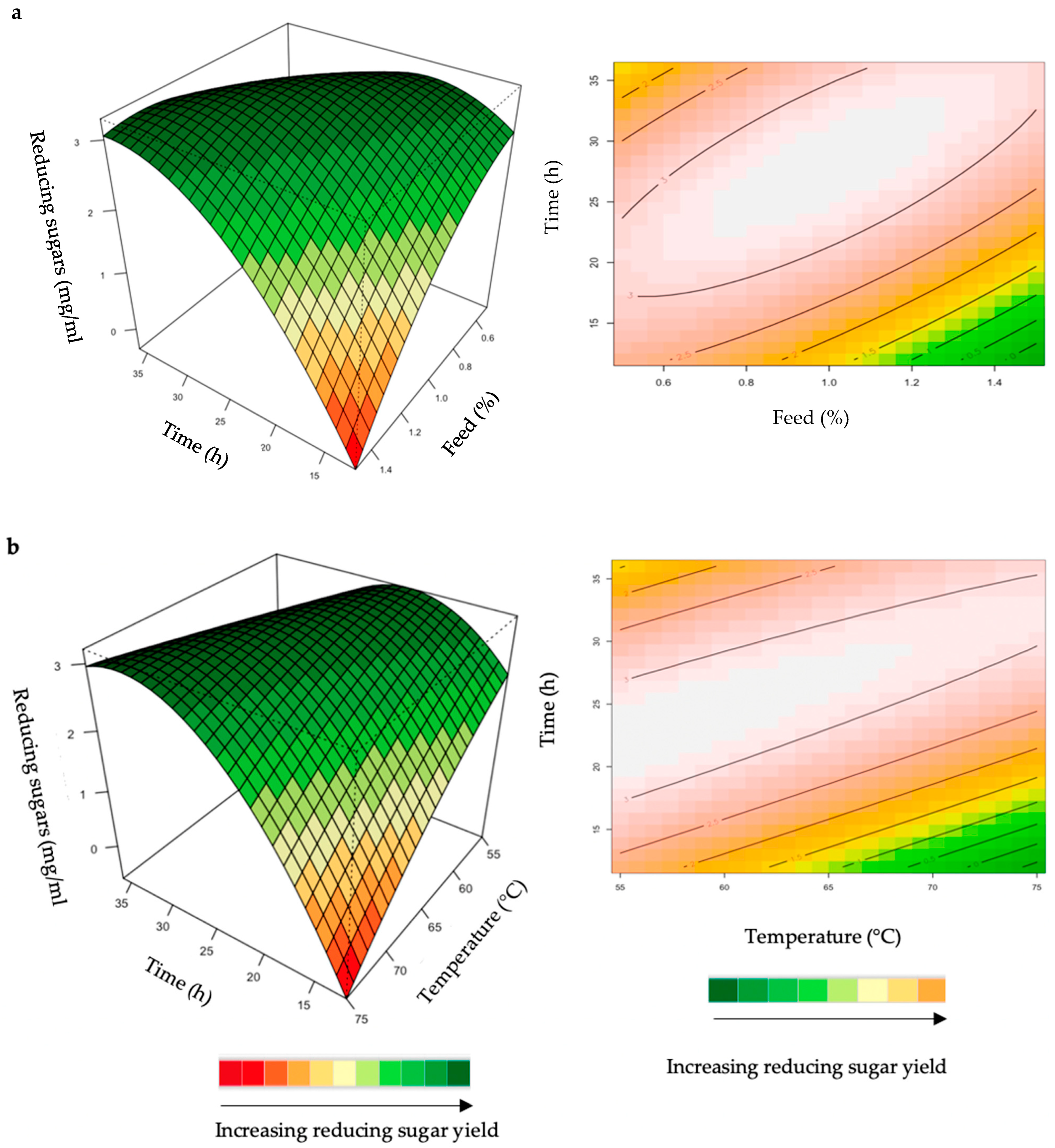
2.1.4. Interaction of Variables for Feed Hydrolysis
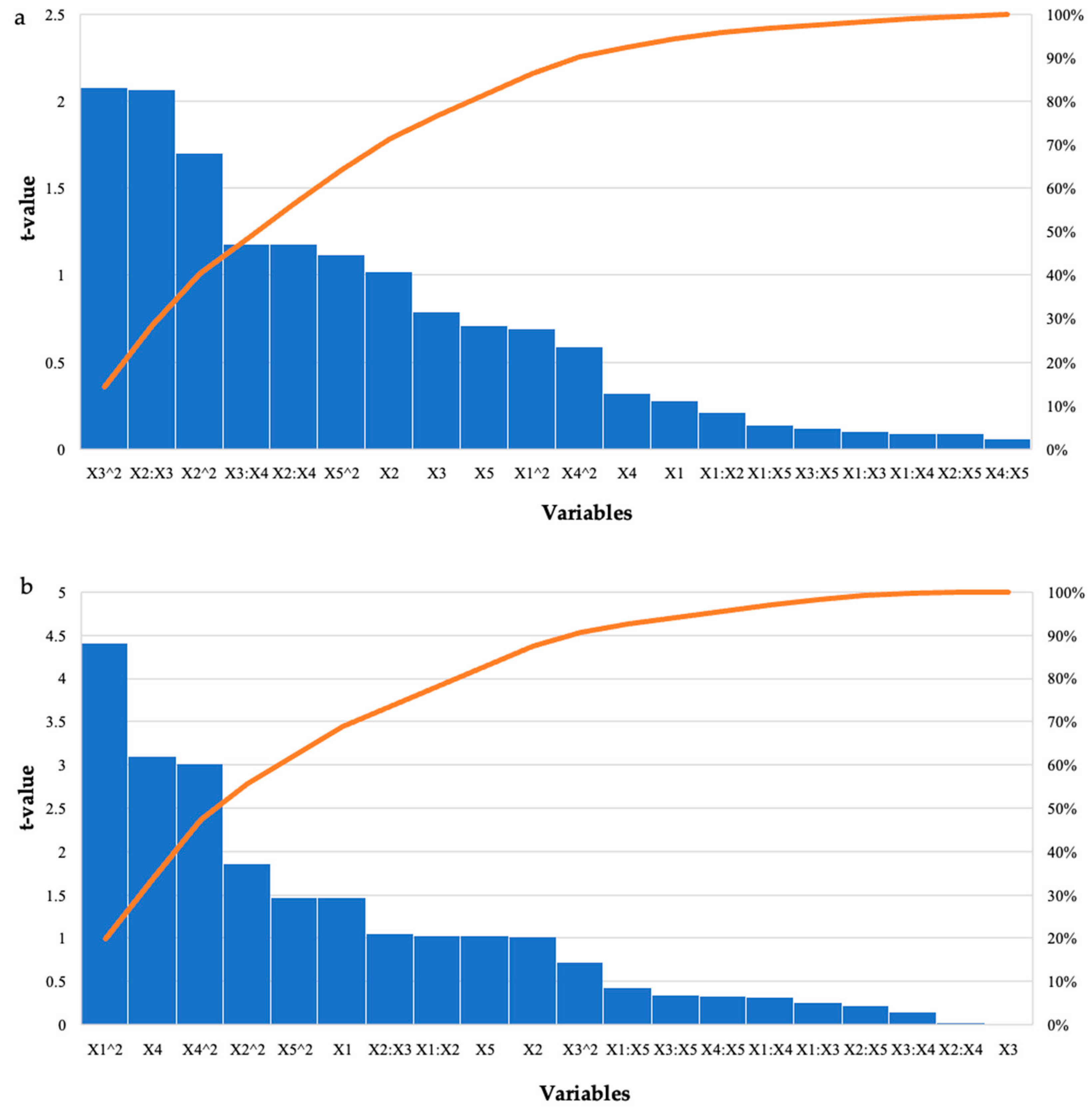
Effect of the Crude T. harzianum Xylanase on Reducing Sugar Yield Following Hydrolysis of Starter and Grower Chicken Feeds
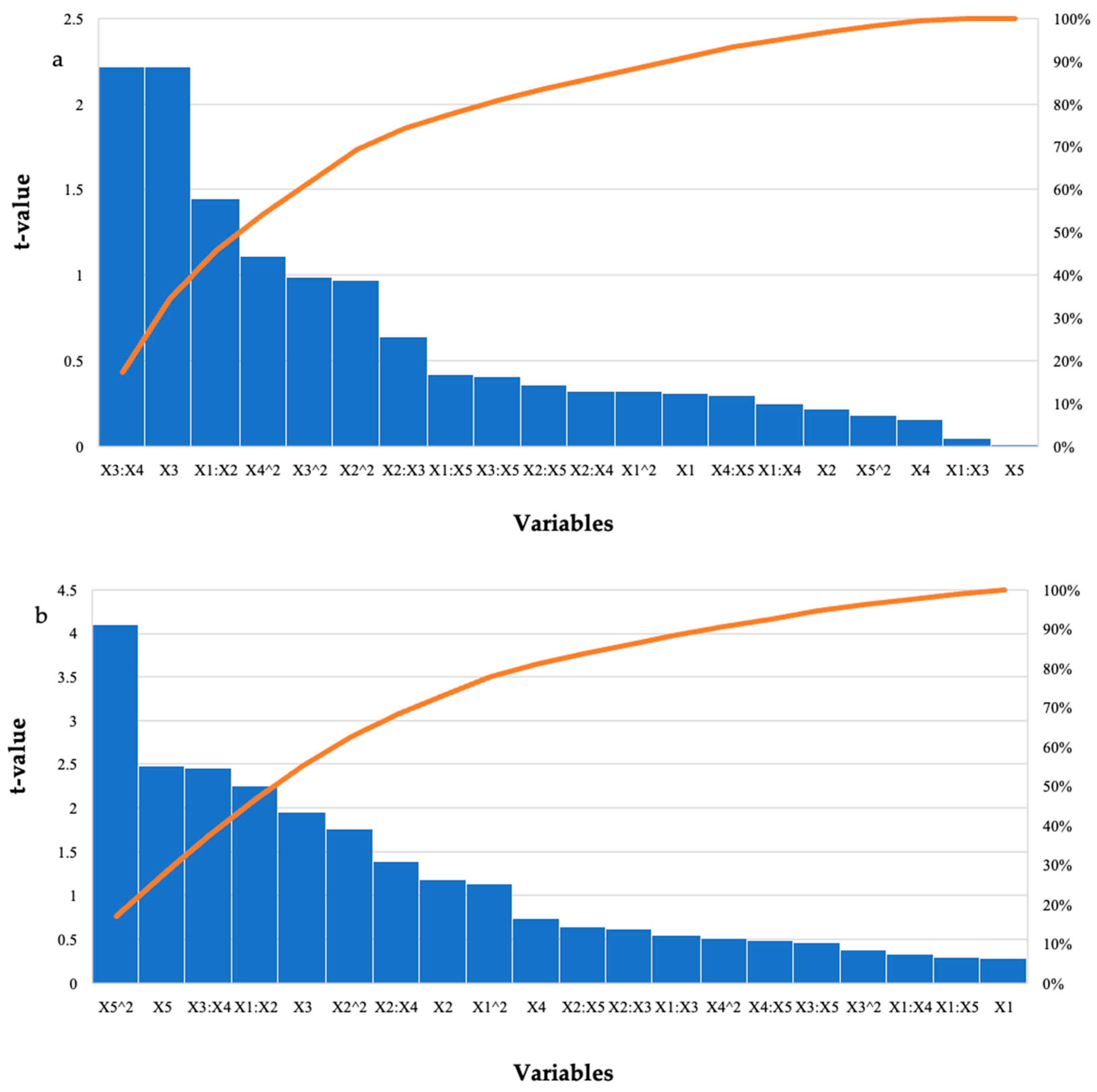
Effect of the Purified T. harzianum Xylanase on Reducing Sugar Yield Following Hydrolysis of Starter and Grower Chicken Feeds
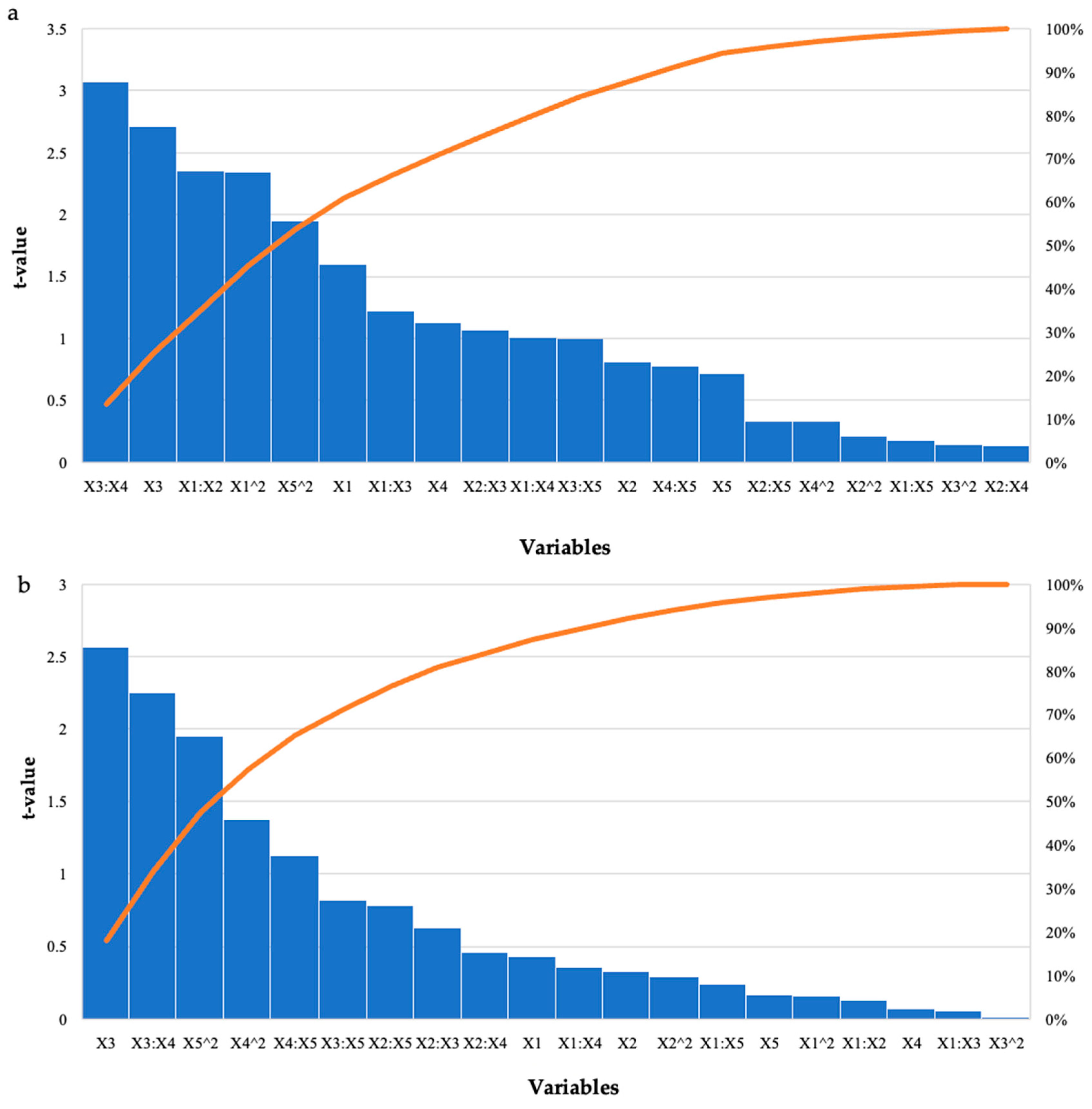
Effect of the Recombinant XT6 Xylanase on Reducing Sugar Yield Following Hydrolysis of Starter and Grower Chicken Feeds
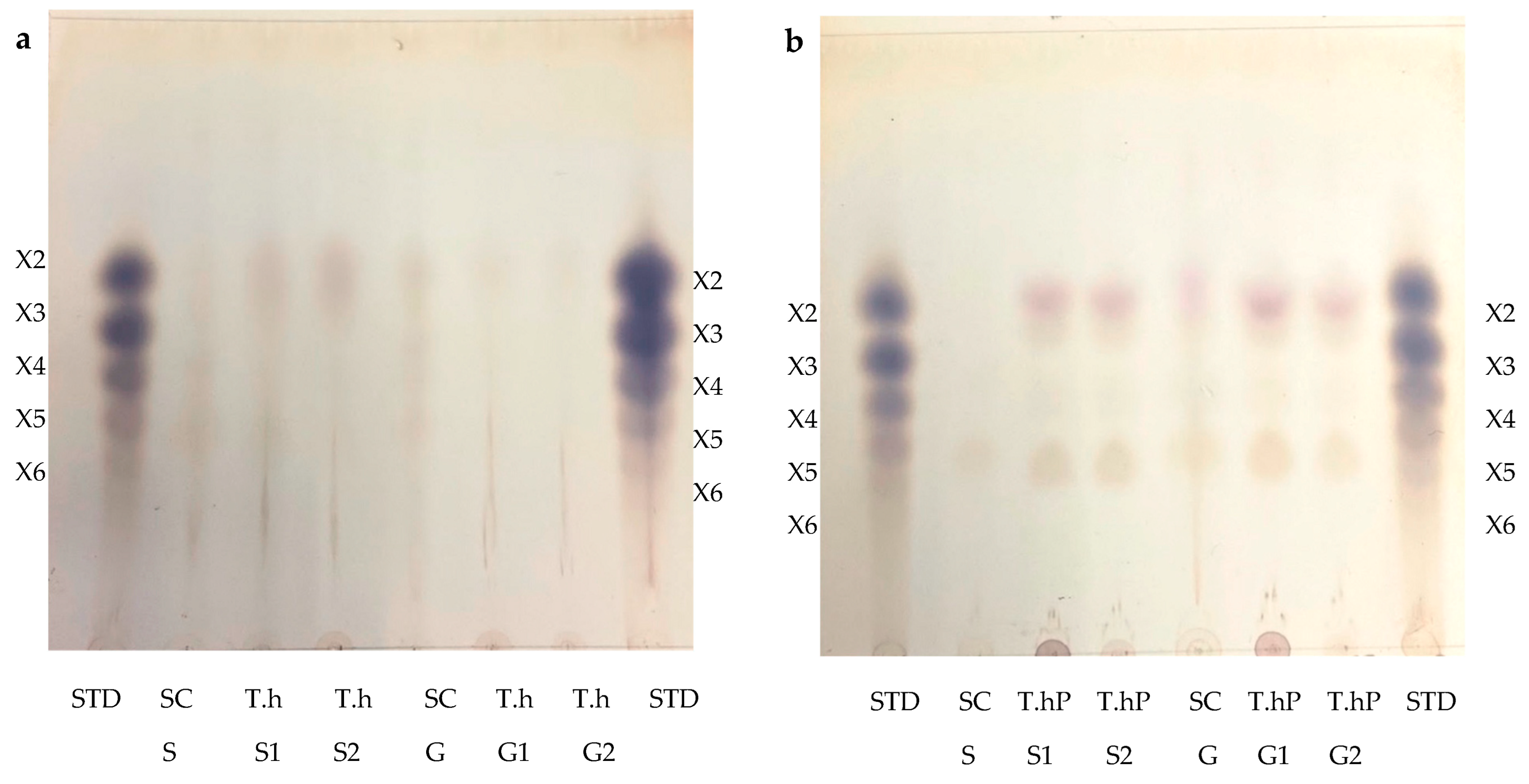
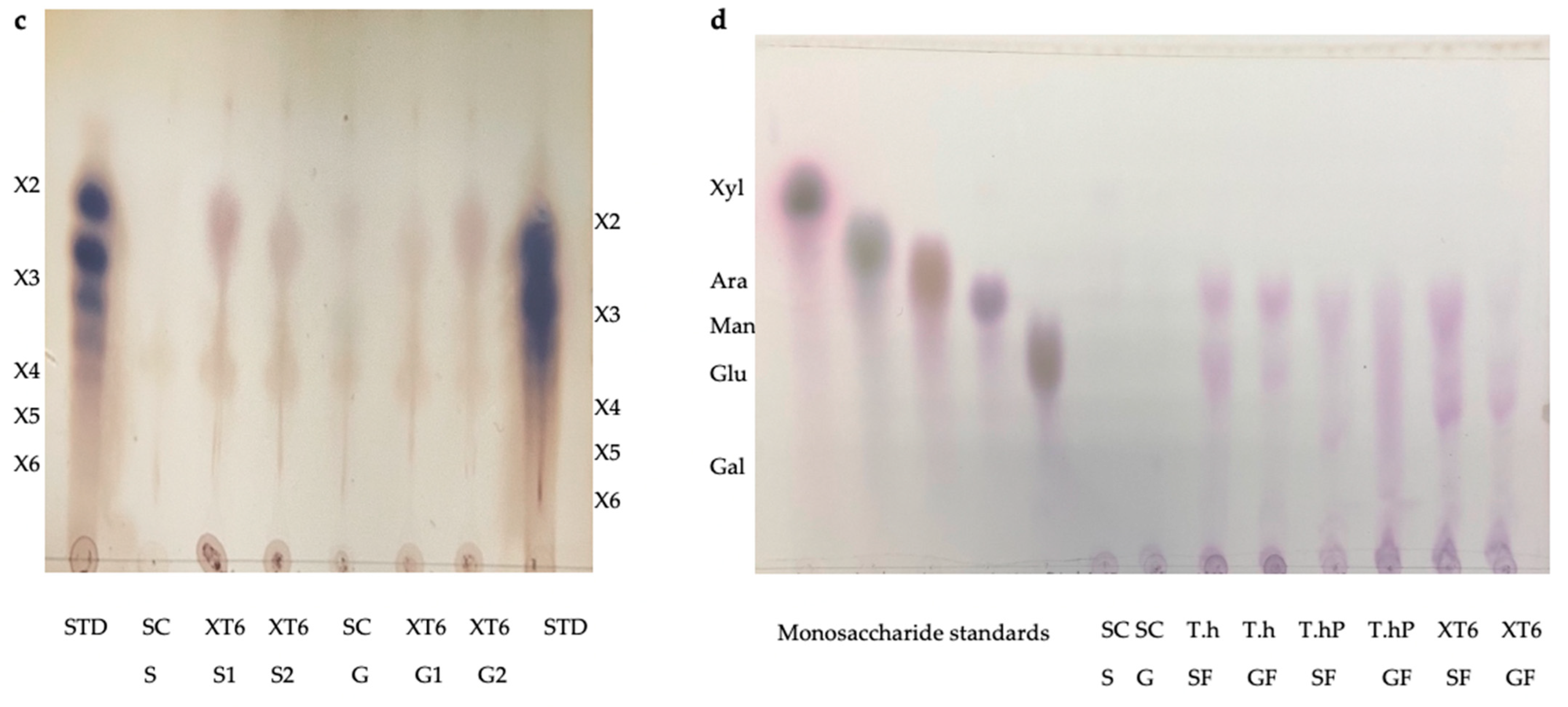
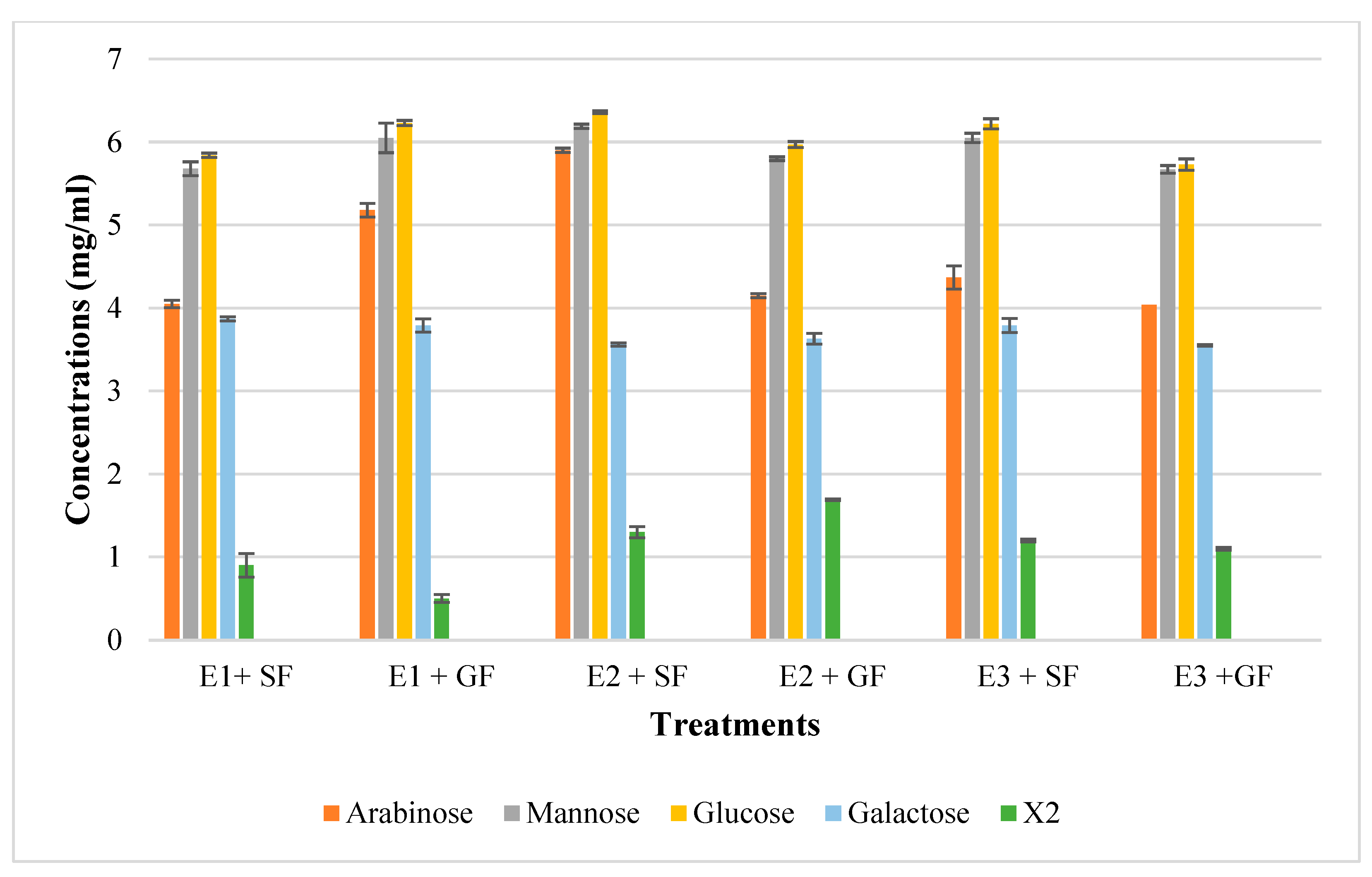
2.2. Thin Layer Chromatography (TLC) and High Performance Liquid Chromatography (HPLC) Analysis of Feed Hydrolysis Products
3. Materials and Methods
3.1. Feed Samples and Enzymes
3.2. Optimization of the Hydrolysis of Feed Using the Crude and Pure Fungal T. harzianum and Pure Recombinant XT6 Xylanases
3.3. Chromatographic Analysis of Hydrolysed Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- McLoughlin, R.F.; Berthon, B.S.; Jensen, M.E.; Baines, K.J.; Wood, L.G. Short-chain fatty acids, prebiotics, synbiotics, and systemic inflammation: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2017, 106, 930–945. [Google Scholar] [CrossRef] [PubMed]
- Passos, A.A.; Park, I.; Ferket, P.; von Heimendahl, E.; Kim, S.W. Effect of dietary supplementation of xylanase on apparent ileal digestibility of nutrients, viscosity of digesta, and intestinal morphology of growing pigs fed corn and soybean meal-based diet. Anim. Nutr. 2015, 1, 19–23. [Google Scholar] [CrossRef]
- Duarte, M.E.; Zhou, F.X.; Dutra, W.M.; Kim, S.W. Dietary supplementation of xylanase and protease on growth performance, digesta viscosity, nutrient digestibility, immune and oxidative stress status, and gut health of newly weaned pigs. Anim. Nutr. 2019, 5, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.E.; Tyus, J.; Kim, S.W. Synbiotic effects of enzyme and probiotics on intestinal health and growth of newly weaned pigs challenged with enterotoxigenic F18+Escherichia coli. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Aragon, C.C.; Ruiz-Matute, A.I.; Corzo, N.; Monti, R.; Guisán, J.M.; Mateo González, C. Production of Xylo-oligosaccharides (XOS) by controlled hydrolysis of xylan using immobilized xylanase from Aspergillus niger with improved properties. Integr. Food Nutr. Metab. 2020, 5, 1–9. [Google Scholar] [CrossRef]
- Donaldson, J.; Świątkiewicz, S.; Arczewka-Włosek, A.; Muszyński, S.; Szymańczyk, S.; Arciszewski, M.B.; Siembida, A.Z.; Kras, K.; Piedra, J.L.V.; Schwarz, T.; et al. Modern hybrid rye, as an alternative energy source for broiler chickens, improves the absorption surface of the small intestine depending on the intestinal part and xylanase supplementation. Animals 2021, 11, 1349. [Google Scholar] [CrossRef]
- Saini, R.; Patel, A.K.; Saini, J.K.; Chen, C.W.; Varjani, S.; Singhania, R.R.; Di Dong, C. Recent advancements in prebiotic oligomers synthesis via enzymatic hydrolysis of lignocellulosic biomass. Bioengineered 2022, 13, 2139–2172. [Google Scholar] [CrossRef]
- Dong, C.D.; Tsai, M.L.; Nargotra, P.; Kour, B.; Chen, C.W.; Sun, P.P.; Sharma, V. Bioprocess development for the production of xylooligosaccharides prebiotics from agro-industrial lignocellulosic waste. Heliyon 2023, 9, 18316. [Google Scholar] [CrossRef]
- Malgas, S.; Pletschke, B.I. The effect of an oligosaccharide reducing-end xylanase, BhRex8A, on the synergistic degradation of xylan backbones by an optimised xylanolytic enzyme cocktail. Enzyme Microb. Technol. 2019, 122, 74–81. [Google Scholar] [CrossRef]
- Carvalho, A.F.A.; Neto, P.D.O.; da Silva, D.F.; Pastore, G.M. Xylo-oligosaccharides from lignocellulosic materials: Chemical structure, health benefits and production by chemical and enzymatic hydrolysis. Food Res. Int. 2023, 51, 75–85. [Google Scholar] [CrossRef]
- Gurpilhares, D.D.B.; Cinelli, L.P.; Simas, N.K.; Pessoa, A., Jr.; Sette, L.D. Marine prebiotics: Polysaccharides and oligosaccharides obtained by using microbial enzymes. Food Chem. 2019, 280, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Y.H.; Wei, Q.Y.; Du, X.J.; Qu, Y.S. Investigating desorption during ethanol elution to improve the quality and antioxidant activity of xylo-oligosaccharides from corn stalk. Bioresour. Technol. 2018, 249, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wu, Y.; Liu, X.; Liu, B.; Cao, H.; Yu, H.; Sarker, S.D.; Nahar, L.; Xiao, J. Functional properties, structural studies and chemo-enzymatic synthesis of oligosaccharides. Trends Food Sci. Technol. 2017, 66, 135–145. [Google Scholar] [CrossRef]
- Bertacchi, S.; Jayaprakash, P.; Morrissey, J.P.; Branduardi, P. Interdependence between lignocellulosic biomasses, enzymatic hydrolysis and yeast cell factories in biorefineries. Microb. Biotechnol. 2022, 15, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.A.; Yalçın, S.; Latorre, J.D.; Basiouni, S.; Attia, Y.A.; Abd El-Wahab, A.; Visscher, C.; El-Seedi, H.R.; Huber, C.; Hafez, H.M.; et al. Probiotics, prebiotics, and phytogenic substances for optimizing gut health in poultry. Microorganisms 2022, 10, 395. [Google Scholar] [CrossRef] [PubMed]
- Morgan, N.K.; Wallace, A.; Bedford, M.R.; Choct, M. Efficiency of xylanases from families 10 and 11 in production of xylo-oligosaccharides from wheat arabinoxylans. Carbohydr. Polym. 2017, 167, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Zhou, J.; Chen, X.; Ji, W.; Zhang, L.; Hu, W.; Lu, Z. Evaluation of an efficient fed-batch enzymatic hydrolysis strategy to improve production of functional xylooligosaccharides from maize straws. Ind. Crops Prod. 2020, 157, 112920. [Google Scholar] [CrossRef]
- Wu, W.J.; Ahn, B.Y. Statistical optimization of medium components by response surface methodology to enhance menaquinone-7 (vitamin k2) production by Bacillus subtilis. J. Microbiol. Biotechnol. 2018, 28, 902–908. [Google Scholar] [CrossRef]
- Dhaver, P.; Pletschke, B.; Sithole, B.; Govinden, R. Optimization, purification, and characterization of xylanase production by a newly isolated Trichoderma harzianum strain by a two-step statistical experimental design strategy. Sci. Rep. 2022, 12, 17791. [Google Scholar] [CrossRef]
- Joshi, N.; Sharma, M.; Singh, S.P. Characterization of a novel xylanase from an extreme temperature hot spring metagenome for xylooligosaccharide production. Appl. Microbiol. Biotechnol. 2020, 104, 4889–4901. [Google Scholar] [CrossRef]
- Khangwal, I.; Nath, S.; Kango, N.; Shukla, P. Endo-xylanase induced xylooligosaccharides production from corn cobs, its structural features, and concentration-dependent antioxidant activities. Biomass Convers. Biorefinery 2022, 12, 5707–5717. [Google Scholar] [CrossRef]
- Stan, G.S.; Badea, I.A.; Aboul-Enein, H.Y. HPLC method for quantification of five compounds in a parenteral form used in treatment of companion animals. J. Chromatogr. Sci. 2016, 54, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Zhao, X.; Wang, Q.; Wang, Y.; Zhou, H. Development and validation of a HPLC method for determination of degree of polymerization of xylo-oligosaccharides. Food Chem. 2016, 213, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Debebe, A.; Temesgen, S.; Redi-Abshiro, M.; Chandravanshi, B.S.; Ele, E. Improvement in analytical methods for determination of sugars in fermented alcoholic beverages. J. Anal. Methods Chem. 2018, 8, 4010298. [Google Scholar] [CrossRef]
- Dhaver, P.; Pletschke, B.; Sithole, B.; Govinden, R. Isolation, screening and partial optimization of thermostable xylanase production under submerged fermentation by fungi in Durban, South Africa. Mycology 2022, 13, 271–292. [Google Scholar] [CrossRef]
- Dhaver, P.; Sithole, T.; Pletschke, B.; Sithole, B.; Govinden, R. Enhanced production of a recombinant xylanase (XT6): Optimization of production and purification, and scaled-up batch fermentation in a stirred tank bioreactor. Sci. Rep. 2023, 13, 20895. [Google Scholar] [CrossRef]
- Chen, Y.; Xie, Y.; Ajuwon, K.M.; Zhong, R.; Li, T.; Chen, L.; Zhang, H.; Beckers, Y.; Everaert, N. Xylo-oligosaccharides, preparation and application to human and animal health: A review. Front. Nutr. 2021, 8, 731930. [Google Scholar] [CrossRef]
- Ataei, D.; Hamidi-Esfahani, Z.; Ahmadi-Gavlighi, H. Enzymatic production of xylooligosaccharides from date (Phoenix dactylifera L.) seed. Food Sci. Nutr. 2020, 8, 6699–6707. [Google Scholar] [CrossRef] [PubMed]
- Chapa, D.; Pandit, P.; Shah, A. Production of xylooligosaccharides from corncob xylan by fungal xylanase and their utilization by probiotics. Bioresour. Technol. 2012, 115, 215–221. [Google Scholar] [CrossRef]
- Ai, Z.; Jiang, Z.; Li, L.; Deng, W.; Kusakabe, I.; Li, H. Immobilization of Streptomyces olivaceoviridis E-86 xylanase on Eudragit S-100 for xylo-oligosaccharide production. Process Biochem. 2005, 40, 2707–2714. [Google Scholar] [CrossRef]
- Yoon, K.Y.; Woodams, E.E.; Hang, Y.D. Enzymatic production of pentoses from the hemicellulose fraction of corn residues. LWT 2006, 39, 387–391. [Google Scholar] [CrossRef]
- Sun, Q.; Patil, P.J.; Singh, A.K.; Teng, C.; Zhou, M.; Zhou, Y.; Fan, G. Optimization of pre-treatment and enzymatic hydrolysis coupled with ultrasonication for the production of xylooligosaccharides from corn cob. Biomass Convers. Biorefinery 2023, 2023. [Google Scholar] [CrossRef]
- Gowdhaman, D.; Ponnusami, V. Production and optimization of xylooligosaccharides from corncob by Bacillus aerophilus KGJ2 xylanase and its antioxidant potential. Int. J. Biol. Macromol. 2015, 79, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Xu, S.; Wang, Z.; Yang, W. Aqueous extraction of corncob xylan and production of xylooligosaccharides. LWT 2005, 38, 677–682. [Google Scholar] [CrossRef]
- Yan, F.; Tian, S.; Du, K.; Xue, X.A.; Gao, P.; Chen, Z. Preparation and nutritional properties of xylooligosaccharides from agricultural and forestry by-products: A comprehensive review. Front. Nutr. 2022, 9, 977548. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, X.; Lu, C.; Lu, P.; Yin, C.; Ye, Z.; Huang, Z. Identification and characterization of a novel endo-β-1,4-xylanase from Streptomyces sp. T7 and its application in xylo-oligosaccharide production. Molecules 2022, 27, 2516. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, N.; Kahar, P.; Lisdiyanti, P.; Lee, J.; Prasetya, B.; Ogino, C.; Kondo, A. GH-10 and GH-11 Endo-1, 4-β-xylanase enzymes from Kitasatospora sp. produce xylose and xylooligosaccharides from sugarcane bagasse with no xylose inhibition. Bioresour. Technol. 2019, 272, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, U.M.; El-Khonezy, M.I.; Shokeer, A.; Abdel-Ghany, S.S.; Bassuny, R.I.; Barakat, A.Z.; Salama, W.H.; Azouz, R.A.M.; Fahmy, A.S. Revealing of a novel xylose-binding site of Geobacillus stearothermophilus xylanase by directed evolution. J. Biochem. 2019, 165, 177–184. [Google Scholar] [CrossRef]
- Rashin, R.; Sohail, M. Xylanolytic Bacillus species for xylooligosaccharides production: A critical review. Bioresour. Bioprocess. 2021, 8, 16. [Google Scholar] [CrossRef]
- Su, Y.; Fang, L.; Wang, P.; Lai, C.; Huang, C.; Ling, Z.; Sun, S.; Yong, Q. Efficient production of xylooligosaccharides rich in xylobiose and xylotriose from poplar by hydrothermal pre-treatment coupled with post-enzymatic hydrolysis. Bioresour. Technol. 2021, 342, 125955. [Google Scholar] [CrossRef]
- Valladares-Diestra, K.K.; de Souza Vandenberghe, L.P.; Soccol, C.R. Integrated xylooligosaccharides production from imidazole-treated sugarcane bagasse with application of in house produced enzymes. Bioresour. Technol. 2022, 362, 127800. [Google Scholar] [CrossRef]
- Finegold, S.M.; Li, Z.; Summanen, P.H.; Downes, J.; Thames, G.; Corbett, K.; Dowd, S.; Krak, M.; Heber, D. Xylooligosaccharides increases bifidobacteria but not lactobacilli in human gut microbiota. Food Funct. 2014, 5, 436–445. [Google Scholar] [CrossRef] [PubMed]
- De Maesschalck, C.; Eeckhaut, V.; Maertens, L.; De Lange, L.; Marchal, L.; Nezer, C.; De Baere, S.; Croubels, S.; Daube, G.; Dewulf, J.; et al. Effects of xylo-oligosaccharides on broiler chicken performance and microbiota. Appl. Environ. Microbiol. 2015, 81, 5880–5888. [Google Scholar] [CrossRef] [PubMed]
- Sithole, T. Cloning, Expression, Partial Characterization and Application of a Recombinant GH10 Xylanase, XT6, from Geobacillus stearothermophilus T6 as an Additive to Chicken Feeds. Master’s Thesis, Rhodes University, Grahamstown, South Africa, 2022. [Google Scholar]
- Biasato, I.; Ferrocino, I.; Biasibetti, E.; Grego, E.; Dabbou, S.; Sereno, A.; Gai, F.; Gasco, L.; Schiavone, A.; Cocolin, L.; et al. Modulation of intestinal microbiota, morphology and mucin composition by dietary insect meal inclusion in free-range chickens. BMC Vet. Res. 2018, 14, 383. [Google Scholar] [CrossRef] [PubMed]
- El-Deek, A.A.; Abdel-Wareth, A.A.; Osman, M.; El-Shafey, M.; Khalifah, A.M.; Elkomy, A.E.; Lohakare, J. Alternative feed ingredients in the finisher diets for sustainable broiler production. Sci. Rep. 2020, 10, 17743. [Google Scholar] [CrossRef]
- Saleh, E.A.; Watkins, S.E.; Waldroup, P.W. Changing time of feeding starter, grower, and finisher diets for broilers 2. birds grown to 2.2 kg. J. App. Poul. Res. 1997, 6, 64–73. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org/ (accessed on 15 March 2023).
- Singh, N.; Sithole, B.; Kumar, A.; Govinden, R. A glucose tolerant β-glucosidase from a newly isolated Neofusicoccum parvum strain F7: Production, purification, and characterization. Sci. Rep. 2023, 13, 5134. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Level | Reducing Sugars (mg/mL) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Run Order | X1 (U/mL) | X2 (%) | X3 (h) | X4 (°C) | X5 (pH) | Starter Chicken Feed | Grower Chicken Feed | ||||
| Crude T. harzianum Xylanase | Purified T. harzianum Xylanase | Recombinant XT6 Xylanase | Crude T. harzianum Xylanase | Purified T. harzianum Xylanase | Recombinant XT6 Xylanase | ||||||
| 1 | 5 (−) | 0.5 (−) | 24 (0) | 65 (0) | 5 (0) | 3.16 | 0.82 | 2.36 | 3.21 | 2.09 | 2.37 |
| 2 | 15 (+) | 0.5 (−) | 24 (0) | 65 (0) | 5 (0) | 3.10 | 1.90 | 1.68 | 2.92 | 1.59 | 2.17 |
| 3 | 5 (−) | 1.5 (+) | 24 (0) | 65 (0) | 5 (0) | 2.78 | 1.99 | 2.12 | 2.56 | 1.73 | 2.62 |
| 4 | 15 (+) | 1.5 (+) | 24 (0) | 65 (0) | 5 (0) | 1.95 | 2.22 | 2.46 | 2.96 | 2.21 | 2.34 |
| 5 | 5 (−) | 1 (0) | 12 (−) | 65 (0) | 5 (0) | 2.02 | 2.23 | 2.10 | 2.85 | 1.68 | 2.20 |
| 6 | 15 (+) | 1 (0) | 12 (−) | 65 (0) | 5 (0) | 2.74 | 2.25 | 2.57 | 2.84 | 1.93 | 2.14 |
| 7 | 5 (−) | 1 (0) | 36 (+) | 65 (0) | 5 (0) | 2.86 | 2.15 | 2.03 | 2.69 | 1.49 | 2.22 |
| 8 | 15 (+) | 1 (0) | 36(+) | 65 (0) | 5 (0) | 3.23 | 2.21 | 1.97 | 2.50 | 1.97 | 2.12 |
| 9 | 10 (0) | 0.5 (−) | 12 (−) | 55 (−) | 5 (0) | 2.60 | 1.87 | 2.18 | 2.46 | 2.00 | 2.16 |
| 10 | 10 (0) | 1.5 (+) | 12 (−) | 55 (−) | 5 (0) | 2.82 | 2.13 | 2.05 | 2.51 | 2.10 | 2.45 |
| 11 | 10 (0) | 0.5 (−) | 36 (+) | 75 (+) | 5 (0) | 2.21 | 2.11 | 2.27 | 2.40 | 2.17 | 2.53 |
| 12 | 10 (0) | 1.5 (+) | 36 (+) | 75 (+) | 5 (0) | 2.93 | 2.81 | 2.98 | 2.83 | 2.31 | 2.79 |
| 13 | 10 (0) | 1 (0) | 24 (0) | 55 (−) | 4 (−) | 2.74 | 2.05 | 2.17 | 2.79 | 1.85 | 2.62 |
| 14 | 10 (0) | 1 (0) | 24 (0) | 75 (+) | 4 (−) | 2.68 | 2.00 | 1.97 | 2.59 | 1.73 | 1.99 |
| 15 | 10 (0) | 1 (0) | 24 (0) | 55 (−) | 6 (+) | 2.38 | 1.68 | 1.67 | 2.57 | 1.55 | 1.81 |
| 16 | 10 (0) | 1 (0) | 24 (0) | 75 (+) | 6 (+) | 2.56 | 1.80 | 1.81 | 2.60 | 1.65 | 1.92 |
| 17 | 10 (0) | 0.5 (−) | 24 (0) | 65 (0) | 4 (−) | 2.60 | 1.88 | 1.71 | 2.28 | 1.29 | 1.94 |
| 18 | 10 (0) | 1.5 (+) | 24 (0) | 65 (0) | 4 (−) | 2.52 | 1.78 | 1.71 | 2.46 | 1.03 | 1.69 |
| 19 | 10 (0) | 0.5 (−) | 24 (0) | 65 (0) | 6 (+) | 2.50 | 1.80 | 1.57 | 2.28 | 1.65 | 1.73 |
| 20 | 10 (0) | 1.5 (+) | 24 (0) | 65 (0) | 6 (+) | 2.08 | 1.91 | 1.72 | 2.31 | 1.68 | 1.99 |
| 21 | 10 (0) | 1 (0) | 24 (0) | 65 (0) | 5 (0) | 1.92 | 2.01 | 1.86 | 1.95 | 1.81 | 1.41 |
| 22 | 10 (0) | 1 (0) | 24 (0) | 65 (0) | 5 (0) | 2.32 | 1.88 | 1.92 | 1.82 | 1.92 | 2.44 |
| 23 | 10 (0) | 1 (0) | 24 (0) | 65 (0) | 5 (0) | 2.21 | 2.13 | 1.94 | 1.77 | 1.91 | 1.95 |
| 24 | 10 (0) | 1 (0) | 24 (0) | 65 (0) | 5 (0) | 2.18 | 2.27 | 2.10 | 2.46 | 2.41 | 2.11 |
| 25 | 5 (−) | 1 (0) | 24 (0) | 55 (−) | 5 (0) | 3.03 | 1.78 | 2.20 | 3.07 | 1.77 | 1.92 |
| 26 | 15 (+) | 1 (0) | 24 (0) | 55 (−) | 5 (0) | 2.66 | 1.59 | 1.64 | 2.63 | 1.60 | 1.73 |
| 27 | 5 (−) | 1 (0) | 24 (0) | 75 (+) | 5 (0) | 2.56 | 1.63 | 1.85 | 2.45 | 1.89 | 1.75 |
| 28 | 15 (+) | 1 (0) | 24 (0) | 75 (+) | 5 (0) | 2.51 | 1.60 | 1.73 | 2.22 | 1.87 | 1.80 |
| 29 | 5 (−) | 1 (0) | 24 (0) | 65 (0) | 4 (−) | 2.33 | 1.86 | 1.87 | 2.16 | 1.25 | 1.65 |
| 30 | 15 (+) | 1 (0) | 24 (0) | 65 (0) | 4 (−) | 2.47 | 1.91 | 1.86 | 2.36 | 1.63 | 1.51 |
| 31 | 5 (−) | 1 (0) | 24 (0) | 65 (0) | 6 (+) | 2.41 | 1.90 | 1.89 | 2.22 | 1.51 | 1.89 |
| 32 | 15 (+) | 1 (0) | 24 (0) | 65 (0) | 6 (+) | 2.05 | 1.71 | 1.80 | 2.13 | 1.76 | 1.90 |
| 33 | 10 (0) | 0.5 (−) | 12 (−) | 65 (0) | 5 (0) | 2.36 | 1.78 | 1.74 | 2.03 | 1.70 | 2.13 |
| 34 | 10 (0) | 1.5 (+) | 12 (-) | 65 (0) | 5 (0) | −5.86 | 1.71 | 1.89 | 1.76 | 1.85 | 2.24 |
| 35 | 10 (0) | 0.5 (−) | 36 (+) | 65 (0) | 5 (0) | 2.28 | 1.90 | 1.69 | 1.58 | 1.72 | 1.91 |
| 36 | 10 (0) | 1.5 (+) | 36(+) | 65 (0) | 5 (0) | 2.48 | 2.16 | 1.92 | 2.10 | 1.89 | 2.48 |
| 37 | 10 (0) | 0.5 (−) | 24 (0) | 55 (−) | 5 (0) | 2.97 | 1.80 | 1.84 | 2.89 | 1.96 | 2.03 |
| 38 | 10 (0) | 1.5 (+) | 24 (0) | 55 (−) | 5 (0) | 4.45 | 1.60 | 2.07 | 2.56 | 1.79 | 1.92 |
| 39 | 10 (0) | 0.5 (−) | 24 (0) | 75 (+) | 5 (0) | 2.51 | 1.52 | 1.79 | 2.31 | 1.35 | 1.59 |
| 40 | 10 (0) | 1.5 (+) | 24 (0) | 75 (+) | 5 (0) | 2.61 | 1.49 | 1.66 | 2.22 | 1.91 | 1.36 |
| 41 | 10 (0) | 1 (0) | 12 (−) | 65 (0) | 4 (−) | 2.49 | 1.60 | 1.67 | 2.24 | 1.71 | 1.93 |
| 42 | 10 (0) | 1 (0) | 36 (+) | 65 (0) | 4 (−) | 2.40 | 1.80 | 1.69 | 2.48 | 1.77 | 1.60 |
| 43 | 10 (0) | 1 (0) | 12 (−) | 65 (0) | 6 (+) | 2.07 | 1.77 | 1.67 | 2.21 | 1.74 | 1.61 |
| 44 | 10 (0) | 1 (0) | 36 (+) | 65 (0) | 6 (+) | 2.41 | 1.73 | 1.64 | 2.22 | 1.60 | 1.83 |
| 45 | 10 (0) | 1 (0) | 24 (0) | 65 (0) | 5 (0) | 8.05 | 1.60 | 1.65 | 1.98 | 1.75 | 2.03 |
| 46 | 10 (0) | 1 (0) | 24 (0) | 65 (0) | 5 (0) | 4.91 | 1.51 | 1.80 | 1.81 | 1.75 | 2.16 |
| 47 | 10 (0) | 1 (0) | 24 (0) | 65 (0) | 5 (0) | 1.90 | 1.52 | 1.74 | 1.62 | 1.89 | 2.29 |
| 48 | 10 (0) | 1 (0) | 24 (0) | 65 (0) | 5 (0) | 7.84 | 1.95 | 2.11 | 1.96 | 2.18 | 2.33 |
| Variable | Estimate | Std. Error | t Value | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Starter Feed | Grower Feed | Starter Feed | Grower Feed | Starter Feed | Grower Feed | Starter Feed | Grower Feed | |
| Model | −23.24 | 27.40 | 49.43 | 8.95 | −0.47 | 3.06 | 0.05 * | 0.005 * |
| Enzyme Dosage (U/mL) | 0.48 | −0.45 | 1.68 | 0.30 | 0.28 | −1.47 | 0.78 | 0.15 |
| Feed Loading (%) | 14.41 | −2.59 | 14.11 | 2.55 | 1.02 | −1.02 | 0.32 | 0.32 |
| Incubation Time (h) | −0.61 | −0.01 | 0.77 | 0.14 | −0.79 | −0.01 | 0.43 | 0.99 |
| Incubation Temperature (°C) | 0.30 | −0.53 | 0.94 | 0.17 | 0.32 | −3.10 | 0.75 | 0.004 * |
| pH | 6.51 | −1.70 | 9.13 | 1.65 | 0.71 | −1.03 | 0.48 | 0.31 |
| Enzyme Dosage (U/mL): Feed Loading (%) | −0.08 | 0.07 | 0.37 | 0.07 | −0.21 | 1.03 | 0.84 | 0.31 |
| Enzyme Dosage (U/mL): Time (h) | −0.01 | −0.01 | 0.02 | 0.02 | −0.10 | −0.26 | 0.92 | 0.80 |
| Enzyme Dosage (U/mL): Temperature (°C) | 0.01 | −0.01 | 0.02 | 0.01 | 0.09 | 0.32 | 0.93 | 0.75 |
| Enzyme Dosage (U/mL): pH | −0.02 | −0.01 | 0.18 | 0.03 | −0.14 | −0.43 | 0.89 | 0.67 |
| Feed Loading (%): Incubation Time (h) | 0.26 | 0.02 | 0.13 | 0.02 | 2.07 | 1.06 | 0.03 * | 0.30 |
| Feed Loading (%): Incubation Temperature (°C) | −0.18 | 0.00 | 0.15 | 0.02 | −1.18 | 0.03 | 0.25 | 0.98 |
| Feed Loading (%): pH | −0.17 | −0.07 | 1.84 | 0.33 | −0.09 | −0.22 | 0.93 | 0.83 |
| Incubation Time (h): Incubation Temperature (°C) | 0.01 | −0.01 | 0.01 | 0.01 | 1.18 | −0.15 | 0.05 * | 0.88 |
| Incubation Time (h): pH | 0.01 | −0.01 | 0.08 | 0.01 | 0.12 | −0.34 | 0.91 | 0.73 |
| Temperature (°C): pH | 0.01 | 0.01 | 0.09 | 0.02 | 0.06 | 0.33 | 0.95 | 0.74 |
| Enzyme Dosage (U/mL)^2 | −0.02 | 0.02 | 0.02 | 0.01 | −0.69 | 4.41 | 0.50 | 0.0001 * |
| Feed Loading (%)^2 | −4.12 | 0.81 | 2.43 | 0.44 | −1.70 | 1.86 | 0.04 * | 0.07 |
| Incubation Time (h)^2 | −0.01 | 0.01 | 0.01 | 0.01 | −2.08 | 0.72 | 0.03 * | 0.48 |
| Incubation Temperature (°C)^2 | −0.01 | 0.03 | 0.01 | 0.01 | −0.59 | 3.02 | 0.56 | 0.005 * |
| pH^2 | −0.68 | 0.02 | 0.61 | 0.11 | −1.12 | 1.47 | 0.27 | 0.15 |
| Variable | Estimate | Std. Error | t Value | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Starter Feed | Grower Feed | Starter Feed | Grower Feed | Starter Feed | Grower Feed | Starter Feed | Grower Feed | |
| Model | 3.52 | 1.06 | 7.90 | 5.87 | 0.44 | 0.18 | 0.03 * | 0.03 * |
| Enzyme Dosage (U/mL) | 0.08 | −0.06 | 0.27 | 0.20 | 0.31 | −0.29 | 0.76 | 0.77 |
| Feed Loading (%) | 0.50 | −1.99 | 2.26 | 1.68 | 0.22 | −1.19 | 0.83 | 0.24 |
| Incubation Time (h) | −0.27 | −0.18 | 0.12 | 0.09 | −2.22 | −1.95 | 0.04 * | 0.06 |
| Incubation Temperature (°C) | 0.02 | −0.08 | 0.15 | 0.11 | 0.16 | −0.74 | 0.87 | 0.47 |
| pH | −0.015 | 2.69 | 1.46 | 1.08 | −0.01 | 2.48 | 0.99 | 0.02 * |
| Enzyme Dosage (U/mL): Feed Loading (%) | −0.09 | 0.10 | 0.06 | 0.04 | −1.45 | 2.25 | 0.16 | 0.03 * |
| Enzyme Dosage (U/mL): Time (h) | 0.00 | 0.01 | 0.00 | 0.01 | 0.05 | 0.55 | 0.05 * | 0.59 |
| Enzyme Dosage (U/mL): Temperature (°C) | 0.00 | 0.01 | 0.00 | 0.02 | 0.25 | 0.33 | 0.80 | 0.74 |
| Enzyme Dosage (U/mL): pH | −0.01 | −0.01 | 0.03 | 0.02 | −0.42 | −0.30 | 0.68 | 0.77 |
| Feed Loading (%): Incubation Time (h) | 0.01 | −0.01 | 0.02 | 0.01 | 0.64 | −0.62 | 0.53 | 0.54 |
| Feed Loading (%): Incubation Temperature (°C) | 0.01 | 0.02 | 0.02 | 0.01 | 0.32 | 1.39 | 0.75 | 0.18 |
| Feed Loading (%): pH | 0.11 | 0.14 | 0.2 | 0.22 | 0.36 | 0.65 | 0.72 | 0.52 |
| Incubation Time (h): Incubation Temperature (°C) | 0.00 | 0.03 | 0.00 | 0.01 | 2.22 | 2.46 | 0.04 * | 0.02 * |
| Incubation Time (h): pH | −0.00 | −0.04 | 0.01 | 0.02 | −0.41 | −0.47 | 0.69 | 0.64 |
| Temperature (°C): pH | 0.00 | 0.05 | 0.01 | 0.01 | −0.30 | 0.49 | 0.77 | 0.63 |
| Enzyme Dosage (U/mL)^2 | 0.00 | −0.03 | 0.00 | 0.02 | 0.32 | −1.14 | 0.75 | 0.27 |
| Feed Loading (%)^2 | −0.38 | −0.50 | 0.39 | 0.29 | −0.97 | −1.76 | 0.34 | 0.09 |
| Incubation Time (h)^2 | 0.00 | −0.01 | 0.00 | 0.01 | 0.99 | −0.38 | 0.33 | 0.71 |
| Incubation Temperature (°C)^2 | −0.00 | −0.01 | 0.00 | 0.01 | −1.11 | −0.51 | 0.28 | 0.61 |
| pH^2 | −0.01 | −0.30 | 0.10 | 0.07 | −0.18 | −4.10 | 0.86 | 0.0003 * |
| Variable | Estimate | Std. Error | t Value | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Starter Feed | Grower Feed | Starter Feed | Grower Feed | Starter Feed | Grower Feed | Starter Feed | Grower Feed | |
| Model | 9.32 | 6.51 | 5.83 | 8.73 | 1.59 | 0.75 | 0.04 * | 0.04 * |
| Enzyme Dosage (U/mL) | −3.17 | −1.28 | 1.98 | 2.96 | −1.60 | −0.43 | 0.12 | 0.67 |
| Feed Loading (%) | −1.34 | −8.30 | 1.67 | 2.49 | −0.81 | −0.33 | 0.43 | 0.74 |
| Incubation Time (h) | −2.48 | −3.51 | 9.14 | 1.37 | −2.71 | −2.57 | 0.01 * | 0.02 * |
| Incubation Temperature (°C) | −1.25 | 1.10 | 1.11 | 1.66 | −1.13 | 0.07 | 0.27 | 0.95 |
| pH | 7.80 | 2.75 | 1.08 | 1.61 | 0.72 | 0.17 | 0.48 | 0.87 |
| Enzyme Dosage (U/mL): Feed Loading (%) | 1.03 | −8.60 | 4.36 | 6.53 | 2.35 | −0.13 | 0.03 * | 0.90 |
| Enzyme Dosage (U/mL): Time (h) | −2.21 | −1.57 | 1.81 | 2.72 | −1.22 | −0.06 | 0.23 | 0.95 |
| Enzyme Dosage (U/mL): Temperature (°C) | 2.21 | 1.17 | 2.18 | 3.27 | 1.01 | 0.36 | 0.32 | 0.72 |
| Enzyme Dosage (U/mL): pH | −3.85 | 7.88 | 2.18 | 3.27 | −0.18 | 0.24 | 0.86 | 0.81 |
| Feed Loading (%): Incubation Time (h) | 1.59 | 1.40 | 1.48 | 2.22 | 1.07 | 0.63 | 0.29 | 0.53 |
| Feed Loading (%): Incubation Temperature (°C) | −2.30 | −1.24 | 1.78 | 2.67 | −0.13 | −0.46 | 0.90 | 0.65 |
| Feed Loading (%): pH | 7.25 | 2.54 | 2.18 | 3.27 | 0.33 | 0.78 | 0.74 | 0.44 |
| Incubation Time (h): Incubation Temperature (°C) | 4.04 | 4.43 | 1.32 | 1.97 | 3.07 | 2.25 | 0.004 * | 0.03 * |
| Incubation Time (h): pH | −8.96 | 1.12 | 9.09 | 1.36 | −1.00 | 0.82 | 0.92 | 0.42 |
| Temperature (°C): pH | 8.51 | 1.85 | 1.09 | 1.63 | 0.78 | 1.13 | 0.44 | 0.27 |
| Enzyme Dosage (U/mL)^2 | 6.70 | 7.01 | 2.86 | 4.29 | 2.34 | 0.16 | 0.03 * | 0.87 |
| Feed Loading (%)^2 | −6.10 | 1.24 | 2.86 | 4.29 | −0.21 | 0.29 | 0.83 | 0.77 |
| Incubation Time (h)^2 | −7.93 | −9.33 | 5.58 | 8.34 | −0.14 | −0.01 | 0.89 | 0.99 |
| Incubation Temperature (°C)^2 | −2.64 | −1.66 | 8.03 | 1.20 | −0.33 | −1.38 | 0.05 * | 0.18 |
| pH^2 | −1.40 | −2.09 | 7.16 | 1.07 | −1.95 | −1.95 | 0.06 | 0.05 * |
| Composition (%) | Starter Feed | Grower Feed |
|---|---|---|
| Methionine | 0.23 | 0.10 |
| Lysine | 0.10 | 0.16 |
| Kynofos 21 (Mono dicalcium phosphate (MDCP)) | 1.15 | 0.80 |
| Salt | 0.36 | 0.30 |
| Premix | 0.30 | 0.30 |
| Feed lime | 1.46 | 2.34 |
| Maize bran | 4.00 | 6.00 |
| Soybean | 35.00 | 20.00 |
| Maize | 57.40 | 70.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhaver, P.; Pletschke, B.; Sithole, B.; Govinden, R. Optimization of Xylooligosaccharides Production by Native and Recombinant Xylanase Hydrolysis of Chicken Feed Substrates. Int. J. Mol. Sci. 2023, 24, 17110. https://doi.org/10.3390/ijms242317110
Dhaver P, Pletschke B, Sithole B, Govinden R. Optimization of Xylooligosaccharides Production by Native and Recombinant Xylanase Hydrolysis of Chicken Feed Substrates. International Journal of Molecular Sciences. 2023; 24(23):17110. https://doi.org/10.3390/ijms242317110
Chicago/Turabian StyleDhaver, Priyashini, Brett Pletschke, Bruce Sithole, and Roshini Govinden. 2023. "Optimization of Xylooligosaccharides Production by Native and Recombinant Xylanase Hydrolysis of Chicken Feed Substrates" International Journal of Molecular Sciences 24, no. 23: 17110. https://doi.org/10.3390/ijms242317110
APA StyleDhaver, P., Pletschke, B., Sithole, B., & Govinden, R. (2023). Optimization of Xylooligosaccharides Production by Native and Recombinant Xylanase Hydrolysis of Chicken Feed Substrates. International Journal of Molecular Sciences, 24(23), 17110. https://doi.org/10.3390/ijms242317110