
RNA and Single-Stranded DNA Phages: Unveiling the Promise from the Underexplored World of Viruses

 , , and
, , and 
Abstract
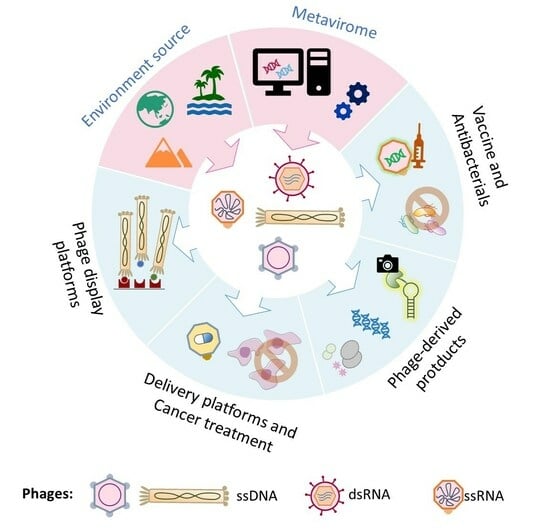
1. Introduction
2. A Brief History of ssDNA and RNA Phages
2.1. ssDNA Phages

{kind=link}
{kind=link}
{kind=link}
| Group | Family | Virion | Genome | Replication | Host Range | Life Cycle | Members 1 | Example [References] |
|---|---|---|---|---|---|---|---|---|
| ssDNA | Inoviridae | Non-enveloped flexible filaments | Circular +ssDNA | Rolling circle | LPS+ Gram-negative bacteria | Chronic infection | 25 genera, 43 species | Phage M13 [24] |
| Paulinoviridae | Non-enveloped flexible filaments | Circular +ssDNA | Rolling circle | LPS− Gram-negative and -positive bacteria | Chronic infection | 2 genera, 2 species | Phage B5; phage OH3 [27,32] | |
| Plectroviridae | Non-enveloped rigid rods | Circular +ssDNA | Rolling circle or transposition | Cell wall-less bacteria | Chronic infection | 4 genera, 6 species | Phage MV-L1 [33] | |
| Microviridae | Non-enveloped icosahedral virions, spikes −/+ | Circular +ssDNA | Rolling circle and other mechanism(s) | Enterobacteria, intracellular parasitic bacteria, cell wall-less bacteria | Lytic | 7 genera, 22 species | Phage φX174; phage 4 [34] | |
| Finnlakeviridae | Icosahedral virion with spikes, internal lipid membrane | Circular +ssDNA | Possibly rolling circle | Gram-negative Flavobacterium | Lytic | 1 genus, 1 species | Phage FLiP [30] | |
| dsRNA | Cystoviridae | Enveloped multi-layer icosahedral virions with spikes | Segmented, linear dsRNA | ssRNA → dsRNA | Gram-negative bacteria, mostly Pseudomonas | Lytic | 1 genus, 7 species | Phage phi6 [35] |
| ssRNA (before 2021) | Leviviridae | Non-enveloped icosahedral virions | Linear +ssRNA | −ssRNA → +ssRNA | Gram-negative bacteria | Lytic | 2 genera, 4 species | Phage MS2 [36], phage Qβ [34] |
| ssRNA (since 2021) 3 | Atkinsviridae | NA 2 | NA 2 | NA 2 | NA 2 | NA 2 | 56 genera, 91 species | Uncultured viral genomes [37] |
| Duinviridae | NA 2 | NA 2 | NA 2 | NA 2 | NA 2 | 6 genera, 6 species | Uncultured viral genomes [37] | |
| Fiersviridae (formerly Leviviridae) | Non-enveloped icosahedral virions | Linear +ssRNA | −ssRNA → +ssRNA | Gram-negative bacteria | Lytic | 185 genera, 298 species | Phage MS2 [36], phage Qβ [37] | |
| Solspiviridae | NA 2 | NA 2 | NA 2 | NA 2 | NA 2 | 24 genera, 31 species | Uncultured viral genomes [37] | |
| Blumeviridae | NA 2 | NA 2 | NA 2 | NA 2 | NA 2 | 31 genera, 35 species | Uncultured viral genomes [37] | |
| Steitzviridae | NA 2 | NA 2 | NA 2 | NA 2 | NA 2 | 117 genera, 412 species | Uncultured viral genomes [37] | |
| Unassigned | NA 2 | NA 2 | NA 2 | NA 2 | NA 2 | 9 genera, 9 species | Uncultured viral genomes [37] |
2.2. dsRNA Phages
2.3. ssRNA Phages
2.4. Current Opportunities and Challenges in Metaviromic Studies
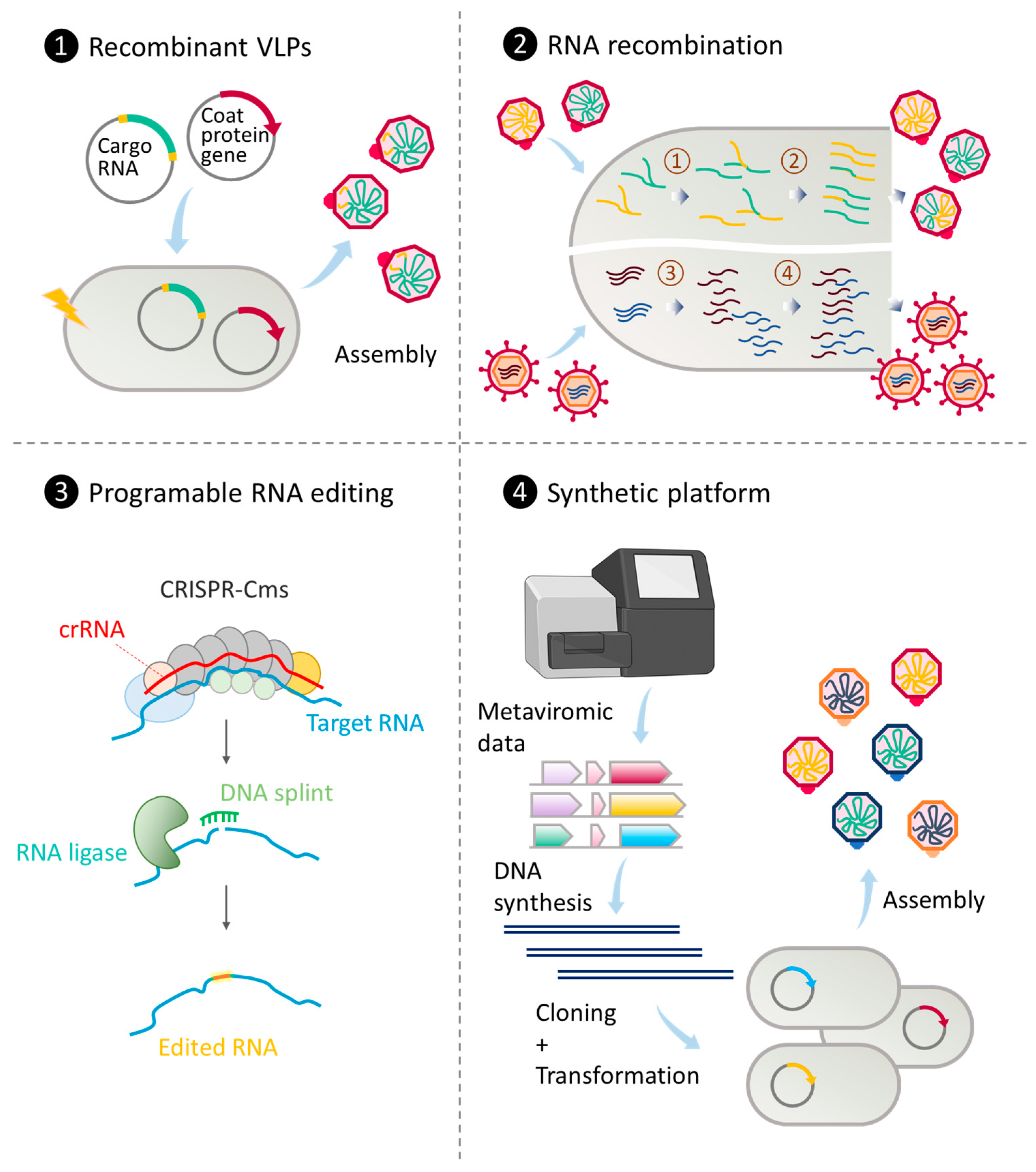
3. Genetic Engineering of RNA and ssDNA Phages
| Phage Components 1 | Tools and Technologies | Applications 1 | Reference(s) |
|---|---|---|---|
| ssDNA phages | Phage display | Library screening | [66,67,68,69] |
| Cancer treatment | [70,71,72] | ||
| Cell adhesive substrates using electrospinning | Drug delivery | [73,74,75] | |
| Carbon nanotubes | Biosensing and imaging | [76,77,78,79,80,81] | |
| dsRNA phages | dsRNA production | RNAi-based crop protection | [82] |
| Surrogate model | dsRNA virus research | [83,84,85] | |
| CP and TR of ssRNA phages | Protein–RNA tethering | ||
| Single tagging | In vivo RNA imaging | [58,86] | |
| Riboswitch screening | [64] | ||
| Dual tagging | In vivo two-color RNA imaging | [87,88,89] | |
| Affinity purification | |||
| RNA–protein | In vitro complex | [59] | |
| In vivo complex | [90] | ||
| RNA–RNA | In vivo complex | [91] | |
| CRISPR/Cas9-based gene regulation | In vivo transcription activation | [61,92] | |
| Recombinant VLPs of ssRNA phages | Therapeutic display | ||
| Peptide | Cell targeting and penetrating | [93,94] | |
| Glycan | Cell targeting | [95] | |
| DNA aptamer | Cell targeting | [96] | |
| Antibody | Cell targeting | [97] | |
| Therapeutic delivery | |||
| RNAs | Small interfering RNA delivery | [98] | |
| Toxins | Protein toxin delivery | [99] | |
| Small molecules | Chemotherapeutic delivery | [100] | |
| Antigen display | Vaccine development | [101,102] | |
| RNA cargo | RNA vaccine development | [103] | |
| Armored RNA | RNA virus detection | [104,105] | |
| Nanoreactor | Controlled enzymatic reaction | [106,107] |
3.1. Engineering ssDNA Phages
3.2. Engineering dsRNA Phages
3.3. Engineering ssRNA Phages
3.4. Techniques for Genetic Engineering of ssDNA and RNA Phages

4. Current Applications of RNA and ssDNA Phages as Therapeutic Agents against Multi-Drug Resistant Bacteria
4.1. Phage-Derived Lytic Enzymes as Antibacterial Agents

4.2. Natural ssDNA and RNA Phages as Antibacterial Agents
4.3. Engineering ssDNA and RNA Phages for Antimicrobial Therapy
5. Concluding Thoughts and Future Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Twort, F.W. An investigation on the nature of ultra-microscopic viruses. Lancet 1915, 186, 4814. [Google Scholar] [CrossRef]
- d’Herelle, F. An invisible microbe that is antagonistic to the dysentery bacillus. Les Comptes Rendus del’Académie Sci. 1917, 165, 373–375. [Google Scholar]
- Kristensen, D.M.; Waller, A.S.; Yamada, T.; Bork, P.; Mushegian, A.R.; Koonin, E.V. Orthologous gene clusters and taxon signature genes for viruses of prokaryotes. J. Bacteriol. 2013, 195, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Casey, A.; Coffey, A.; McAuliffe, O. Genetics and Genomics of Bacteriophages: The Evolution of Bacteriophage Genomes and Genomic Research. Bacteriophages Biol. Technol. Ther. 2021, 2, 193–218. [Google Scholar] [CrossRef]
- Chanishvili, N. Phage Therapy-History from Twort and d’Herelle Through Soviet Experience to Current Approaches. In Advances in Virus Research; Academic Press Inc.: Cambridge, MA, USA, 2012; Volume 83, pp. 3–40. [Google Scholar] [CrossRef]
- Ventola, C.L. The Antibiotic Resistance Crisis Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Watts, G. Phage therapy: Revival of the bygone antimicrobial. Lancet 2017, 390, 2539–2540. [Google Scholar] [CrossRef]
- Pires, D.P.; Costa, A.R.; Pinto, G.; Meneses, L.; Azeredo, J. Current challenges and future opportunities of phage therapy. FEMS Microbiol. Rev. 2020, 44, 684–700. [Google Scholar] [CrossRef]
- Veeranarayanan, S.; Azam, A.H.; Kiga, K.; Watanabe, S.; Cui, L. Bacteriophages as solid tumor theragnostic agents. Int. J. Mol. Sci. 2022, 23, 402. [Google Scholar] [CrossRef]
- Azam, A.H.; Tan, X.E.; Veeranarayanan, S.; Kiga, K.; Cui, L. Bacteriophage technology and modern medicine. Antibiotics 2021, 10, 999. [Google Scholar] [CrossRef]
- Ackermann, H.W. Phage classification and characterization. Methods Mol. Biol. 2009, 501, 127–140. [Google Scholar] [CrossRef]
- Turner, D.; Shkoporov, A.N.; Lood, C.; Millard, A.D.; Dutilh, B.E.; Alfenas-Zerbini, P.; van Zyl, L.J.; Aziz, R.K.; Oksanen, H.M.; Poranen, M.M.; et al. Abolishment of morphology-based taxa and change to binomial species names: 2022 taxonomy update of the ICTV bacterial viruses subcommittee. Arch. Virol. 2023, 168, 74. [Google Scholar] [CrossRef] [PubMed]
- Sertic, V.; Boulgakov, N. Classification et identification des typhiphage. CR Soc. Biol. Paris 1935, 119, 1270–1272. [Google Scholar]
- Vidaver, A.K.; Koski, R.K.; Van Etten, J.L. Bacteriophage φ6: A Lipid-Containing Virus of Pseudomonas phaseolicola. J. Virol. 1973, 11, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Loeb, T.; Zinder, N.D. A bacteriophage containing RNA. Proc. Natl. Acad. Sci. USA 1961, 47, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Callanan, J.; Stockdale, S.R.; Shkoporov, A.; Draper, L.A.; Ross, R.P.; Hill, C. Expansion of known ssRNA Phage Genomes: From Tens to over a Thousand. Sci. Adv. 2020, 6, eaay5981. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, J.; Santamaría, R.I.; Bustos, P.; González, V. Novel lineages of single-stranded DNA phages that coevolved with the symbiotic bacteria Rhizobium. Front. Microbiol. 2022, 13, 990394. [Google Scholar] [CrossRef]
- Pumpens, P. Single-Stranded RNA Phages; Taylor & Francis Ltd.: London, UK, 2020. [Google Scholar] [CrossRef]
- Callanan, J.; Stockdale, S.R.; Shkoporov, A.; Draper, L.A.; Ross, R.P.; Hill, C. RNA phage biology in a metagenomic Era. Viruses 2018, 10, 386. [Google Scholar] [CrossRef]
- Mata, C.P.; Rodríguez, J.M.; Suzuki, N.; Castón, J.R. Structure and assembly of double-stranded RNA mycoviruses. Adv. Virus. Res. 2020, 108, 213–247. [Google Scholar] [CrossRef]
- Tucker, K.P.; Parsons, R.; Symonds, E.M.; Breitbart, M. Diversity and distribution of single-stranded DNA phages in the North Atlantic Ocean. ISME J. 2010, 5, 822–830. [Google Scholar] [CrossRef]
- Székely, A.J.; Breitbart, M. Single-stranded DNA phages: From early molecular biology tools to recent revolutions in environmental microbiology. FEMS Microbiol. Lett. 2016, 363, fnw027. [Google Scholar] [CrossRef]
- Tisza, M.J.; Buck, C.B.; Chang, Y. A catalog of tens of thousands of viruses from human metagenomes reveals hidden associations with chronic diseases. Proc. Natl. Acad. Sci. USA 2021, 118, e2023202118. [Google Scholar] [CrossRef]
- Knezevic, P.; Adriaenssens, E.M. ICTV virus taxonomy profile: Inoviridae. J. Gen. Virol. 2021, 102, 001614. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.O.; Hutchison, C.A.; Pfannkoch, C.; Venter, J.C. Generating a Synthetic Genome by Whole Genome Assembly: X174 Bacteriophage from Synthetic Oligonucleotides. Proc. Natl. Acad. Sci. USA 2003, 100, 15440–15445. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.; Borrebaeck, C.A.K.; Nilsson, N.; Malmborg-Hager, A.C. The mechanism of bacterial infection by filamentous phages involves molecular interactions between TolA and phage protein 3 domains. J. Bacteriol. 2003, 185, 2628–2634. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Roux, S.; Krupovic, M.; Daly, R.A.; Borges, A.L.; Nayfach, S.; Schulz, F.; Sharrar, A.; Carnevali, P.B.M.; Cheng, J.-F.; Ivanova, N.N.; et al. Cryptic inoviruses revealed as pervasive in bacteria and archaea across Earth’s biomes. Nat. Microbiol. 2019, 4, 1895–1906. [Google Scholar] [CrossRef] [PubMed]
- Krupovic, M.; Forterre, P. Microviridae goes temperate: Microvirus-related proviruses reside in the genomes of bacteroidetes. PLoS ONE 2011, 6, e19893. [Google Scholar] [CrossRef] [PubMed]
- Kirchberger, P.C.; Martinez, Z.A.; Ochman, H. Organizing the Global Diversity of Microviruses. mBio 2022, 13, e0058822. [Google Scholar] [CrossRef]
- Mäntynen, S.; Laanto, E.; Sundberg, L.R.; Poranen, M.M.; Oksanen, H.M. ICTV virus taxonomy profile: Finnlakeviridae. J. Gen. Virol. 2020, 101, 894–895. [Google Scholar] [CrossRef]
- Laanto, E.; Mäntynen, S.; De Colibus, L.; Marjakangas, J.; Gillum, A.; Stuart, D.I.; Ravantti, J.J.; Huiskonen, J.T.; Sundberg, L.-R. Virus found in a boreal lake links ssDNA and dsDNA viruses. Proc. Natl. Acad. Sci. USA 2017, 114, 8378–8383. [Google Scholar] [CrossRef]
- Gupta, R.S. Origin of diderm (Gram-negative) bacteria: Antibiotic selection pressure rather than endosymbiosis likely led to the evolution of bacterial cells with two membranes. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2011, 100, 171–182. [Google Scholar] [CrossRef]
- Knezevic, P.; Adriaenssens, E.M. ICTV virus taxonomy profile: Plectroviridae. J. Gen. Virol. 2021, 102, 001597. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2011; Volume 9. [Google Scholar]
- Poranen, M.M.; Mäntynen, S. ICTV virus taxonomy profile: Cystoviridae. J. Gen. Virol. 2017, 98, 2423–2424. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.E.; Strauss, J.H.; Sinsheimer, R.L. Bacteriophage MS2–another RNA phage. Science 1961, 134, 1427. [Google Scholar]
- Callanan, J.; Stockdale, S.R.; Adriaenssens, E.M.; Kuhn, J.H.; Rumnieks, J.; Pallen, M.J.; Shkoporov, A.N.; Draper, L.A.; Ross, R.P.; Hill, C. Leviviricetes: Expanding and restructuring the taxonomy of bacteria-infecting single-stranded RNA viruses. Microb. Genom. 2021, 7, 000686. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, P.; Alimova, A. RNA Packaging in the Cystovirus Bacteriophages: Dynamic Interactions during Capsid Maturation. Int. J. Mol. Sci. 2022, 23, 2677. [Google Scholar] [CrossRef]
- Mindich, L.; Qiao, X.; Qiao, J.; Onodera, S.; Romantschuk, M.; Hoogstraten, D. Isolation of additional bacteriophages with genomes of segmented double- stranded RNA. J. Bacteriol. 1999, 181, 4505–4508. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Sun, Y.; Qiao, J.; Di Sanzo, F.; Mindich, L. Characterization of Φ2954, a newly isolated bacteriophage containing three dsRNA genomic segments. BMC Microbiol. 2010, 10, 55. [Google Scholar] [CrossRef]
- Mäntynen, S.; Laanto, E.; Kohvakka, A.; Poranen, M.M.; Bamford, J.K.H.; Ravantti, J.J. New enveloped dsRNA phage from freshwater habitat. J. Gen. Virol. 2015, 96, 1180–1189. [Google Scholar] [CrossRef]
- Yang, Y.; Lu, S.; Shen, W.; Zhao, X.; Shen, M.; Tan, Y.; Li, G.; Li, M.; Wang, J.; Hu, F.; et al. Characterization of the first double-stranded RNA bacteriophage infecting Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 38795. [Google Scholar] [CrossRef]
- Sanz-Gaitero, M.; Seoane-Blanco, M.; Van Raaij, M.J. Structure and Function of Bacteriophages. In Bacteriophages; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Crawford, E.M.; Gesteland, R.F. The adsorption of bacteriophage R-17. Virology 1964, 22, 165–167. [Google Scholar] [CrossRef]
- Harb, L.; Chamakura, K.; Khara, P.; Christie, P.J.; Young, R.; Zeng, L. SsRNA phage penetration triggers detachment of the F-pilus. Proc. Natl. Acad. Sci. USA 2020, 117, 25751–25758. [Google Scholar] [CrossRef] [PubMed]
- Bamford, D.H.; Palva, E.T.; Lounatmaa, K. Ultrastructure and Life Cycle of the Lipid-containing Bacteriophage phi6. J. Gen. Virol. 1976, 32, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Bollback, J.P.; Phylogeny, J.P. Phylogeny, Genome Evolution, and Host Specificity of Single-Stranded RNA Bacteriophage (Family Leviviridae). J. Mol. Evol. 2001, 52, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Li, Z.; Lai, M.; Shu, S.; Du, Y.; Zhou, Z.H.; Sun, R. In situ structures of the genome and genome-delivery apparatus in a single-stranded RNA virus. Nature 2017, 541, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Benler, S.; Koonin, E.V. Fishing for phages in metagenomes: What do we catch, what do we miss? Curr. Opin. Virol. 2021, 49, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Malathi, V.G.; Devi, P.R. ssDNA viruses: Key players in global virome. Virusdisease 2019, 30, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Dolja, V.V.; Krupovic, M.; Varsani, A.; Wolf, Y.I.; Yutin, N.; Kuhn, J.H. Global organization and proposed megataxonomy of the virus world. Microbiol. Mol. Biol. Rev. 2020, 84, e00061-19. [Google Scholar] [CrossRef]
- Neri, U.; Wolf, Y.I.; Roux, S.; Camargo, A.P.; Lee, B.; Kazlauskas, D.; Chen, I.M.; Ivanova, N.; Allen, L.Z.; Paez-Espino, D.; et al. Expansion of the global RNA virome reveals diverse clades of bacteriophages. Cell 2022, 185, 4023–4037.e18. [Google Scholar] [CrossRef]
- Thurber, R.V.; Haynes, M.; Breitbart, M.; Wegley, L.; Rohwer, F. Laboratory procedures to generate viral metagenomes. Nat. Protoc. 2009, 4, 470–483. [Google Scholar] [CrossRef]
- Nakagawa, S.; Sakaguchi, S.; Ogura, A.; Mineta, K.; Endo, T.; Suzuki, Y.; Gojobori, T. Current trends in RNA virus detection through metatranscriptome sequencing data. FEBS Open Bio 2023, 13, 992–1000. [Google Scholar] [CrossRef]
- Gann, E.R.; Kang, Y.; Dyhrman, S.T.; Gobler, C.J.; Wilhelm, S.W. Metatranscriptome Library Preparation Influences Analyses of Viral Community Activity During a Brown Tide Bloom. Front. Microbiol. 2021, 12, 4189. [Google Scholar] [CrossRef] [PubMed]
- Urayama, S.-I.; Takaki, Y.; Nishi, S.; Yoshida-Takashima, Y.; Deguchi, S.; Takai, K.; Nunoura, T. Unveiling the RNA virosphere associated with marine microorganisms. Mol. Ecol. Resour. 2018, 18, 1444–1455. [Google Scholar] [CrossRef]
- Pierrel, J. An RNA Phage Lab: MS2 in Walter Fiers’ Laboratory of Molecular Biology in Ghent, from Genetic Code to Gene and Genome, 1963–1976. J. Hist. Biol. 2010, 45, 109–138. [Google Scholar] [CrossRef] [PubMed]
- Lampasona, A.A.; Czaplinski, K. RNA voyeurism: A coming of age story. Methods 2016, 98, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Bardwell, V.J.; Wickens, M. Purification of RNA and RNA-protein complexes by an R17 coat protein affinity method. Nucleic Acids Res. 1990, 18, 6587–6594. [Google Scholar] [CrossRef] [PubMed]
- Aalto, A.P.; Sarin, L.P.; van Dijk, A.A.; Saarma, M.; Poranen, M.M.; Arumäe, U.; Bamford, D.H. Large-scale production of dsRNA and siRNA pools for RNA interference utilizing bacteriophage φ6 RNA-dependent RNA polymerase. RNA 2007, 13, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Dahlman, J.E.; Abudayyeh, O.O.; Joung, J.; Gootenberg, J.S.; Zhang, F.; Konermann, S. Orthogonal gene knockout and activation with a catalytically active Cas9 nuclease. Nat. Biotechnol. 2015, 33, 1159–1161. [Google Scholar] [CrossRef] [PubMed]
- Petrov, G.; Dymova, M.; Richter, V. Bacteriophage-Mediated Cancer Gene Therapy. Int. J. Mol. Sci. 2022, 23, 14245. [Google Scholar] [CrossRef]
- Lee, B.Y.; Lee, J.; Ahn, D.J.; Lee, S.; Oh, M.K. Optimizing protein V untranslated region sequence in M13 phage for increased production of single-stranded DNA for origami. Nucleic Acids Res. 2021, 49, 6596–6603. [Google Scholar] [CrossRef]
- Wu, M.J.; Andreasson, J.O.L.; Kladwang, W.; Greenleaf, W.; Das, R. Automated Design of Diverse Stand-Alone Riboswitches. ACS Synth. Biol. 2019, 8, 1838–1846. [Google Scholar] [CrossRef]
- Kiga, K.; Tan, X.-E.; Ibarra-Chávez, R.; Watanabe, S.; Aiba, Y.; Sato’o, Y.; Li, F.-Y.; Sasahara, T.; Cui, B.; Kawauchi, M.; et al. Development of CRISPR-Cas13a-based antimicrobials capable of sequence-specific killing of target bacteria. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.J.; Smith, P.G. Searching for peptide ligands with an epitope library. Science 1990, 249, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Parmley, S.F.; Smith, G.P. Antibody-selectable filamentous fd phage vectors: Affinity purification of target genes. Gene 1988, 73, 305–318. [Google Scholar] [CrossRef] [PubMed]
- John, M.; Andrew, G.D.; Greg, W.; David, C.J. Phage antibodies: Filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554. [Google Scholar]
- Smith, G.P.; Petrenko, V.A. Phage Display. Chem. Rev. 1997, 97, 391–410. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Chang, Y.-C.; Hu, C.-W.; Kao, C.-Y.; Yu, Y.-A.; Lim, S.-K.; Mou, K.Y. Development of a Novel Cytokine Vehicle Using Filamentous Phage Display for Colorectal Cancer Treatment. ACS Synth. Biol. 2021, 10, 2087–2095. [Google Scholar] [CrossRef]
- Dong, X.; Pan, P.; Zheng, D.-W.; Bao, P.; Zeng, X.; Zhang, X.-Z. Bioinorganic hybrid bacteriophage for modulation of intestinal microbiota to remodel tumor-immune microenvironment against colorectal cancer. Sci. Adv. 2020, 6, eaba1590. [Google Scholar] [CrossRef]
- Jin, H.E.; Farr, R.; Lee, S.W. Collagen mimetic peptide engineered M13 bacteriophage for collagen targeting and imaging in cancer. Biomaterials 2014, 35, 9236–9245. [Google Scholar] [CrossRef]
- Shin, Y.C.; Lee, J.H.; Kim, M.J.; Park, J.H.; Kim, S.E.; Kim, J.S.; Oh, J.-W.; Han, D.-W. Biomimetic Hybrid Nanofiber Sheets Composed of RGD Peptide-Decorated PLGA as Cell-Adhesive Substrates. J. Funct. Biomater. 2015, 6, 367–378. [Google Scholar] [CrossRef]
- Ling, H.; Lou, X.; Luo, Q.; He, Z.; Sun, M.; Sun, J. Recent advances in bacteriophage-based therapeutics: Insight into the post-antibiotic era. Acta Pharm. Sin. B 2022, 12, 4348–4364. [Google Scholar] [CrossRef]
- Chang, C.; Guo, W.; Yu, X.; Guo, C.; Zhou, N.; Guo, X.; Huang, R.-L.; Li, Q.; Zhu, Y. Engineered M13 phage as a novel therapeutic bionanomaterial for clinical applications: From tissue regeneration to cancer therapy. Mater. Today Bio 2023, 20, 100612. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Bagley, A.F.; Na, Y.J.; Birrer, M.J.; Bhatia, S.N.; Belcher, A.M. Deep, noninvasive imaging and surgical guidance of submillimeter tumors using targeted M13-stabilized single-walled carbon nanotubes. Proc. Natl. Acad. Sci. USA 2014, 111, 13948–13953. [Google Scholar] [CrossRef] [PubMed]
- Bardhan, N.M.; Ghosh, D.; Belcher, A.M. Carbon nanotubes as in vivo bacterial probes. Nat. Commun. 2014, 5, 4918. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Ghosh, D.; Ham, M.-H.; Qi, J.; Barone, P.W.; Strano, M.S.; Belcher, A.M. M13 phage-functionalized single-walled carbon nanotubes as nanoprobes for second near-infrared window fluorescence imaging of targeted tumors. Nano Lett. 2012, 12, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Passaretti, P.; Sun, Y.; Dafforn, T.R.; Oppenheimer, P.G. Determination and characterisation of the surface charge properties of the bacteriophage M13 to assist bio-nanoengineering. RSC Adv. 2020, 10, 25385–25392. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Qi, J.; Dequilettes, D.W.; Huang, M.; Lin, C.-W.; Bardhan, N.M.; Dang, X.; Bulović, V.; Belcher, A.M. M13 Virus-Based Framework for High Fluorescence Enhancement. Small 2019, 15, e1901233. [Google Scholar] [CrossRef]
- Lentini, G.; Fazio, E.; Calabrese, F.; De Plano, L.M.; Puliafico, M.; Franco, D.; Nicolò, M.S.; Carnazza, S.; Trusso, S.; Allegra, A.; et al. Phage-AgNPs complex as SERS probe for U937 cell identification. Biosens. Bioelectron. 2015, 74, 398–405. [Google Scholar] [CrossRef]
- Niehl, A.; Soininen, M.; Poranen, M.M.; Heinlein, M. Synthetic biology approach for plant protection using dsRNA. Plant Biotechnol. J. 2018, 16, 1679–1687. [Google Scholar] [CrossRef]
- Adcock, N.J.; Rice, E.W.; Sivaganesan, M.; Brown, J.D.; Stallknecht, D.E.; Swayne, D.E. The use of bacteriophages of the family Cystoviridae as surrogates for H5N1 highly pathogenic avian influenza viruses in persistence and inactivation studies. J. Environ. Sci. Health Part A 2009, 44, 1362–1366. [Google Scholar] [CrossRef]
- Fedorenko, A.; Grinberg, M.; Orevi, T.; Kashtan, N. Survival of the enveloped bacteriophage Phi6 (a surrogate for SARS-CoV-2) in evaporated saliva microdroplets deposited on glass surfaces. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Wood, J.P.; Richter, W.; Sunderman, M.; Calfee, M.W.; Serre, S.; Mickelsen, L. Evaluating the Environmental Persistence and Inactivation of MS2 Bacteriophage and the Presumed Ebola Virus Surrogate Phi6 Using Low Concentration Hydrogen Peroxide Vapor. Environ. Sci. Technol. 2020, 54, 3581–3590. [Google Scholar] [CrossRef]
- Tyagi, S. Imaging intracellular RNA distribution and dynamics in living cells. Nat. Methods 2009, 6, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Moon, H.C.; Park, H.Y. Live-cell imaging of single mRNA dynamics using split superfolder green fluorescent proteins with minimal background. RNA 2020, 26, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Chen, J.; Singer, R.H. Background free imaging of single mRNAs in live cells using split fluorescent proteins. Sci. Rep. 2014, 4, 3615. [Google Scholar] [CrossRef] [PubMed]
- Hocine, S.; Raymond, P.; Zenklusen, D.; Chao, J.A.; Singer, R.H. Single-molecule analysis of gene expression using two-color RNA labeling in live yeast. Nat. Methods 2012, 10, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Tsai, B.P.; Wang, X.; Huang, L.; Waterman, M.L. Quantitative profiling of in vivo-assembled RNA-protein complexes using a novel integrated proteomic approach. Mol. Cell. Proteom. 2011, 10, 1–15. [Google Scholar] [CrossRef]
- Je-Hyun, Y.; Myriam, G. Identification of mRNA-Interacting Factors by MS2-TRAP (MS2-Tagged RNA Affinity Purification). In RNA-Protein Complexes and Interactions; Methods in Molecular Biology; Lin, R.-J., Ed.; Springer Science+Business Media: New York, NY, USA, 2016; Volume 1421, pp. 15–22. [Google Scholar] [CrossRef]
- Konermann, S.; Brigham, M.D.; Trevino, A.E.; Joung, J.; Abudayyeh, O.O.; Barcena, C.; Hsu, P.D.; Habib, N.; Gootenberg, J.S.; Nishimasu, H.; et al. Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature 2014, 517, 583–588. [Google Scholar] [CrossRef]
- Carrico, Z.M.; Romanini, D.W.; Mehl, R.A.; Francis, M.B. Oxidative coupling of peptides to a virus capsid containing unnatural amino acids. Chem. Commun. 2008, 10, 1205–1207. [Google Scholar] [CrossRef]
- Wang, G.; Jia, T.; Xu, X.; Chang, L.; Zhang, R.; Fu, Y.; Li, Y.; Yang, X.; Zhang, K.; Lin, G.; et al. Novel miR-122 delivery system based on MS2 virus like particle surface displaying cell-penetrating peptide TAT for hepatocellular carcinoma. Oncotarget 2016, 7, 59402–59416. [Google Scholar] [CrossRef]
- Rhee, J.K.; Baksh, M.; Nycholat, C.; Paulson, J.C.; Kitagishi, H.; Finn, M.G. Glycan-targeted virus-like nanoparticles for photodynamic therapy. Biomacromolecules 2012, 13, 2333–2338. [Google Scholar] [CrossRef]
- Cohen, B.A.; Bergkvist, M. Targeted in vitro photodynamic therapy via aptamer-labeled, porphyrin-loaded virus capsids. J. Photochem. Photobiol. B Biol. 2013, 121, 67–74. [Google Scholar] [CrossRef] [PubMed]
- ElSohly, A.M.; Netirojjanakul, C.; Aanei, I.L.; Jager, A.; Bendall, S.C.; Farkas, M.E.; Nolan, G.P.; Francis, M.B. Synthetically Modified Viral Capsids as Versatile Carriers for Use in Antibody-Based Cell Targeting. Bioconjugate Chem. 2015, 26, 1590–1596. [Google Scholar] [CrossRef] [PubMed]
- Galaway, F.A.; Stockley, P.G. MS2 Viruslike Particles: A Robust, Semisynthetic Targeted Drug Delivery Platform. Mol. Pharm. 2012, 10, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Brown, W.L.; Stockley, P.G. Cell-Specific Delivery of Bacteriophage-Encapsidated Ricin a Chain. Bioconjugate Chem. 1995, 6, 587–595. [Google Scholar] [CrossRef]
- Finbloom, J.A.; Aanei, I.L.; Bernard, J.M.; Klass, S.H.; Elledge, S.K.; Han, K.; Ozawa, T.; Nicolaides, T.P.; Berger, M.S.; Francis, M.B. Evaluation of three morphologically distinct virus-like particles as nanocarriers for convection-enhanced drug delivery to glioblastoma. Nanomaterials 2018, 8, 1007. [Google Scholar] [CrossRef]
- Kozlovska, T.M.; Cielens, I.; Vasiljeva, I.; Strelnikova, A.; Kazaks, A.; Dislers, A.; Pumpens, P. RNA phage Qb Coat Protein as a Carrier for Foreign Epitopes. Intervirology 1996, 39, 9–15. [Google Scholar] [CrossRef]
- Peabody, D.S.; Peabody, J.; Bradfute, S.B.; Chackerian, B. RNA Phage VLP-Based Vaccine Platforms. Pharmaceuticals 2021, 14, 764. [Google Scholar] [CrossRef]
- Sun, S.; Li, W.; Sun, Y.; Pan, Y.; Li, J. A new RNA vaccine platform based on MS2 virus-like particles produced in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2011, 407, 124–128. [Google Scholar] [CrossRef]
- Pasloske, B.L.; Walkerpeach, C.R.; Obermoeller, R.D.; Winkler, M.; Dubois, D.B. Armored RNA technology for production of ribonuclease-resistant viral RNA controls and standards. J. Clin. Microbiol. 1998, 36, 3590–3594. [Google Scholar] [CrossRef]
- Yao, L.; Li, F.; Qu, M.; Guo, Y.; Jiang, Y.; Wang, L.; Zhai, Y. Development and Evaluation of a Novel Armored RNA Technology Using Bacteriophage Qβ. Food Environ. Virol. 2019, 11, 383–392. [Google Scholar] [CrossRef]
- Fiedler, J.D.; Brown, S.D.; Lau, J.L.; Finn, M.G. RNA-directed packaging of enzymes within virus-like particles. Angew. Chem. Int. Ed. 2010, 49, 9648–9651. [Google Scholar] [CrossRef]
- Giessen, T.W.; Silver, P.A. A Catalytic Nanoreactor Based on in Vivo Encapsulation of Multiple Enzymes in an Engineered Protein Nanocompartment. ChemBioChem 2016, 17, 1931–1935. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, S.W.; Ji, S.T.; Kim, Y.J.; Jang, W.B.; Oh, J.-W.; Kim, J.; Yoo, S.Y.; Beak, S.H.; Kwon, S.-M. Engineered M13 Nanofiber Accelerates Ischemic Neovascularization by Enhancing Endothelial Progenitor Cells. Tissue Eng. Regen. Med. 2017, 14, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.-W. RNA-based antiviral immunity. Nat. Rev. Immunol. 2010, 10, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Makeyev, E.V.; Bamford, D.H. Replicase activity of purified recombinant protein P2 of double-stranded RNA bacteriophage phi 6. EMBO J. 2000, 19, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Tars, K. SsRNA phages: Life cycle, structure and applications. In Biocommunication of Phages; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 261–292. [Google Scholar] [CrossRef]
- Karin, V.; James, M.B.; Peter, S.G.; Nicola, S.J.; Lars, L. Crystal structure of an RNA bacteriophage coat protein-operator complex. Nature 1994, 371, 623–626. [Google Scholar]
- Chao, J.A.; Patskovsky, Y.; Almo, S.C.; Singer, R.H. Structural basis for the coevolution of a viral RNA-protein complex. Nat. Struct. Mol. Biol. 2008, 15, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Persson, M.; Tars, K.; Liljas, L. PRR1 coat protein binding to its RNA translational operator. Acta Crystallogr. Sect. D Struct. Biol. 2013, 69, 367–372. [Google Scholar] [CrossRef]
- Kozlovska, T.M.; Cielēns, I.; Dreilinņa, D.; Dišlers, A.; Baumanis, V.; Ose, V.; Pumpēns, P. Recombinant rna phage qβ capsid particles synthesized and self-assembled in escherichia coli. Gene 1993, 137, 133–137. [Google Scholar] [CrossRef]
- Freivalds, J.; Dislers, A.; Ose, V.; Skrastina, D.; Cielens, I.; Pumpens, P.; Sasnauskas, K.; Kazaks, A. Assembly of bacteriophage Qβ virus-like particles in yeast Saccharomyces cerevisiae and Pichia pastoris. J. Biotechnol. 2006, 123, 297–303. [Google Scholar] [CrossRef]
- Pumpens, P.; Renhofa, R.; Dishlers, A.; Kozlovska, T.; Ose, V.; Pushko, P.; Tars, K.; Grens, E.; Bachmann, M.F. The true story and advantages of RNA phage capsids as nanotools. Intervirology 2016, 59, 74–110. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Ahn, S.J.; Park, C.; Bae, H.W.; Kim, E.S.; Cho, Y.H. Reverse genetic systems for pseudomonas aeruginosa leviphages. Methods Protoc. 2019, 2, 22. [Google Scholar] [CrossRef]
- Kim, E.S.; Lee, J.-Y.; Park, C.; Ahn, S.-J.; Bae, H.-W.; Cho, Y.-H. cDNA-Derived RNA Phage Assembly Reveals Critical Residues in the Maturation Protein of the Pseudomonas aeruginosa Leviphage PP7. J. Virol. 2021, 95, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.-Y.; Kim, B.-O.; Han, J.-H.; Park, J.; Kang, H.K.; Park, Y.; Cho, Y.-H. A phage protein-derived antipathogenic peptide that targets type IV pilus assembly. Virulence 2021, 12, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Qiao, X.; Mindich, L. Construction of carrier state viruses with partial genomes of the segmented dsRNA bacteriophages. Virology 2004, 319, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Lund, P.E.; Hunt, R.C.; Gottesman, M.M.; Kimchi-Sarfaty, C. Pseudovirions as vehicles for the delivery of siRNA. Pharm. Res. 2009, 27, 400–420. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.; Jimenez, S.; Slavcev, R.A. Construction and characterization of a novel miniaturized filamentous phagemid for targeted mammalian gene transfer. Microb. Cell Factories 2023, 22, 124. [Google Scholar] [CrossRef]
- Bikard, D.; Euler, C.W.; Jiang, W.; Nussenzweig, P.M.; Goldberg, G.W.; Duportet, X.; A Fischetti, V.; A Marraffini, L. Exploiting CRISPR-cas nucleases to produce sequence-specific antimicrobials. Nat. Biotechnol. 2014, 32, 1146–1150. [Google Scholar] [CrossRef]
- Fang, P.Y.; Bowman, J.C.; Ramos, L.M.G.; Hsiao, C.; Williams, L.D. RNA: Packaged and protected by VLPs. RSC Adv. 2018, 8, 21399–21406. [Google Scholar] [CrossRef]
- Wang, H.; Cui, X.; Cai, X.; An, T. Recombination in Positive-Strand RNA Viruses. Front. Microbiol. 2022, 13, 870759. [Google Scholar] [CrossRef]
- Gottlieb, P.; Alimova, A. Heterologous RNA Recombination in the Cystoviruses φ6 and φ8: A Mechanism of Viral Variation and Genome Repair. Viruses 2022, 14, 2589. [Google Scholar] [CrossRef] [PubMed]
- Nemudryi, A.; Nemudraia, A.; Nichols, J.E.; Scherffius, A.M.; Zahl, T.; Wiedenheft, B.A. CRISPR-based engineering of RNA viruses. Sci. Adv. 2023, 9, eadj8277. [Google Scholar] [CrossRef] [PubMed]
- Liekniņa, I.; Kalniņš, G.; Akopjana, I.; Bogans, J.; Šišovs, M.; Jansons, J.; Rūmnieks, J.; Tārs, K. Production and characterization of novel ssRNA bacteriophage virus-like particles from metagenomic sequencing data. J. Nanobiotechnology 2019, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- van Meerten, D.; Groeneveld, H.; Miller, D.M.J.; Marechal, G.B.; Tsareva, N.V.; Olsthoorn, R.C.L.; de la Peña, M.; van Duin, J. In vivo generation of hybrids between different species of RNA phages. J. Gen. Virol. 2002, 83, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Chetverin, A.B.; Chetverina, H.V. Nonhomologous RNA Recombination in a Cell-Free System: Evidence for a Transesterification Mechanism Guided by Secondary Structure. Cell 1997, 88, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Lowry, K.; Woodman, A.; Cook, J.; Evans, D.J. Recombination in Enteroviruses Is a Biphasic Replicative Process Involving the Generation of Greater-than Genome Length ‘Imprecise’ Intermediates. PLoS Pathog. 2014, 10, e1004191. [Google Scholar] [CrossRef]
- Hulseberg, C.E.; Fénéant, L.; Szyman’ska, K.M.; Szymanska-De Wijs, S.; Kessler, N.P.; Nelson, E.A.; Shoemaker, C.J.; Schmaljohn, C.S.; Polyak, S.J.; White, J.M.; et al. Senecavirus-Specific Recombination Assays Reveal the Intimate Link between Polymerase Fidelity and RNA Recombination. J. Virol. 2019, 93, e02185-18. [Google Scholar] [CrossRef]
- Woodman, A.; Lee, K.-M.; Janissen, R.; Gong, Y.-N.; Dekker, N.H.; Shih, S.-R.; Cameron, C.E. Predicting Intraserotypic Recombination in Enterovirus 71. J. Virol. 2019, 93, 10-1128. [Google Scholar] [CrossRef]
- Salomaa, M. Host Range of Cystoviruses. 2021. Available online: https://helda.helsinki.fi/items/05b55016-d12d-44b2-baeb-e99c5b1f6a27 (accessed on 26 November 2023).
- Micoli, F.; Bagnoli, F.; Rappuoli, R.; Serruto, D. The role of vaccines in combatting antimicrobial resistance. Nat. Rev. Microbiol. 2021, 19, 287–302. [Google Scholar] [CrossRef]
- Rodríguez-Rubio, L.; Martínez, B.; Donovan, D.M.; Rodríguez, A.; García, P. Bacteriophage virion-associated peptidoglycan hydrolases: Potential new enzybiotics. Crit. Rev. Microbiol. 2013, 39, 427–434. [Google Scholar] [CrossRef]
- Latka, A.; Maciejewska, B.; Majkowska-Skrobek, G.; Briers, Y.; Drulis-Kawa, Z. Bacteriophage-encoded virion-associated enzymes to overcome the carbohydrate barriers during the infection process. Appl. Microbiol. Biotechnol. 2017, 101, 3103–3119. [Google Scholar] [CrossRef] [PubMed]
- Gerstmans, H.; Criel, B.; Briers, Y. Synthetic biology of modular endolysins. Biotechnol. Adv. 2018, 36, 624–640. [Google Scholar] [CrossRef] [PubMed]
- Caldentey, J.; Bamford, D.H. The lytic enzyme of the Pseudomonas phage b6. Purification and biochemical characterization. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1992, 1159, 44–50. [Google Scholar] [CrossRef]
- Chamakura, K.; Young, R. Phage single-gene lysis: Finding the weak spot in the bacterial cell wall. J. Biol. Chem. 2019, 294, 3350–3358. [Google Scholar] [CrossRef]
- Adler, B.A.; Chamakura, K.; Carion, H.; Krog, J.; Deutschbauer, A.M.; Young, R.; Mutalik, V.K.; Arkin, A.P. Multicopy suppressor screens reveal convergent evolution of single-gene lysis proteins. Nat. Chem. Biol. 2023, 19, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Le, S.; Shen, W.; Chen, Q.; Huang, Y.; Lu, S.; Tan, Y.; Li, M.; Hu, F.; Li, Y. Antibacterial activity of a lytic enzyme encoded by Pseudomonas aeruginosa double stranded RNA Bacteriophage phiYY. Front. Microbiol. 2018, 9, 1778. [Google Scholar] [CrossRef] [PubMed]
- Orta, A.K.; Riera, N.; Li, Y.E.; Tanaka, S.; Yun, H.G.; Klaic, L.; Clemons, W.M. The mechanism of the phage-encoded protein antibiotic from ΦX174. Science 2023, 381, 180. [Google Scholar] [CrossRef]
- Yang, Y.; Shen, W.; Zhong, Q.; Chen, Q.; He, X.; Baker, J.L.; Xiong, K.; Jin, X.; Wang, J.; Hu, F.; et al. Development of a Bacteriophage Cocktail to Constrain the Emergence of Phage-Resistant Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 327. [Google Scholar] [CrossRef]
- Li, L.; Zhong, Q.; Zhao, Y.; Bao, J.; Liu, B.; Zhong, Z.; Wang, J.; Yang, L.; Zhang, T.; Cheng, M.; et al. First-in-human application of double-stranded RNA bacteriophage in the treatment of pulmonary Pseudomonas aeruginosa infection. Microb. Biotechnol. 2023, 16, 862–867. [Google Scholar] [CrossRef]
- Huo, C.-X.; Dhara, D.; Baliban, S.M.; Nick, S.T.; Tan, Z.; Simon, R.; Misra, A.K.; Huang, X. Synthetic and immunological studies of: Salmonella Enteritidis O-antigen tetrasaccharides as potential anti- Salmonella vaccines. Chem. Commun. 2019, 55, 4519–4522. [Google Scholar] [CrossRef]
- Rashidijahanabad, Z.; Kelly, M.; Kamruzzaman, M.; Qadri, F.; Bhuiyan, T.R.; McFall-Boegeman, H.; Wu, D.; Piszczek, G.; Xu, P.; Ryan, E.T.; et al. Virus-like Particle Display of Vibrio cholerae O-Specific Polysaccharide as a Potential Vaccine against Cholera. ACS Infect. Dis. 2022, 8, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.K.; Collins, J.J. Engineered Bacteriophage Targeting Gene Networks as Adjuvants for Antibiotic Therapy. Proc. Natl. Acad. Sci. USA 2009, 106, 4629–4634. [Google Scholar] [CrossRef] [PubMed]
- Gan, T.; Wang, D. Picobirnaviruses encode proteins that are functional bacterial lysins. Proc. Natl. Acad. Sci. USA 2023, 120, e2309647120. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, T.G.; Wang, I.-N.; Struck, D.K.; Young, R. A Protein Antibiotic in the Phage Q Virion: Diversity in Lysis Targets. Science 2001, 292, 2326–2329. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Y.; Li, P.; Han, Q.; Zhang, T.; Yang, B.; Wu, W.; Yang, H. Phage phiZ98: A novel tri-segmented dsRNA cystovirus for controlling Pseudomonas strains with defective lipopolysaccharides in foods. Food Res. Int. 2022, 162, 112197. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Zhang, H.; Shen, W.; Zou, Z.; Lu, S.; Li, G.; He, X.; Agnello, M.; Shi, W.; Hu, F.; et al. Pseudomonas aeruginosa MutL promotes large chromosomal deletions through non-homologous end joining to prevent bacteriophage predation. Nucleic Acids Res. 2018, 46, 4505–4514. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, L.A.M.; Pereira, C.; Frazão, C.; Balcão, V.M.; Almeida, A. Efficiency of phage φ6 for biocontrol of pseudomonas syringae pv. Syringae: An in vitro preliminary study. Microorganisms 2019, 7, 286. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, L.A.M.; Pereira, C.; Barreal, M.E.; Gallego, P.P.; Balcão, V.M.; Almeida, A. Use of phage ϕ6 to inactivate Pseudomonas syringae pv. actinidiae in kiwifruit plants: In vitro and ex vivo experiments. Appl. Microbiol. Biotechnol. 2019, 104, 1319–1330. [Google Scholar] [CrossRef]
- Villalpando-Aguilar, J.L.; Matos-Pech, G.; López-Rosas, I.; Castelán-Sánchez, H.G.; Alatorre-Cobos, F. Phage Therapy for Crops: Concepts, Experimental and Bioinformatics Approaches to Direct Its Application. Int. J. Mol. Sci. 2022, 24, 325. [Google Scholar] [CrossRef]
- Bhattarai, S.R.; Yoo, S.Y.; Lee, S.W.; Dean, D. Engineered phage-based therapeutic materials inhibit Chlamydia trachomatis intracellular infection. Biomaterials 2012, 33, 5166–5174. [Google Scholar] [CrossRef]
- Huang, J.X.; Bishop-Hurley, S.L.; Cooper, M.A. Development of anti-infectives using phage display: Biological agents against bacteria, viruses, and parasites. Antimicrob. Agents Chemother. 2012, 56, 4569–4582. [Google Scholar] [CrossRef] [PubMed]
- Prokopczuk, F.I.; Im, H.; Campos-Gomez, J.; Orihuela, C.J.; Martínez, E. Engineered Superinfective Pf Phage Prevents Dissemination of Pseudomonas aeruginosa in a Mouse Burn Model. mBio 2023, 14, e0047223. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.N.; Spanogiannopoulos, P.; Soto-Perez, P.; Alexander, M.; Nalley, M.J.; Bisanz, J.E.; Nayak, R.R.; Weakley, A.M.; Yu, F.B.; Turnbaugh, P.J. Phage-delivered CRISPR-Cas9 for strain-specific depletion and genomic deletions in the gut microbiome. Cell Rep. 2021, 37, 109930. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, H.M.; Watanabe, S.; Sharmin, S.; Kawaguchi, T.; Tan, X.-E.; Wannigama, D.L.; Cui, L. RNA and Single-Stranded DNA Phages: Unveiling the Promise from the Underexplored World of Viruses. Int. J. Mol. Sci. 2023, 24, 17029. https://doi.org/10.3390/ijms242317029
Nguyen HM, Watanabe S, Sharmin S, Kawaguchi T, Tan X-E, Wannigama DL, Cui L. RNA and Single-Stranded DNA Phages: Unveiling the Promise from the Underexplored World of Viruses. International Journal of Molecular Sciences. 2023; 24(23):17029. https://doi.org/10.3390/ijms242317029
Chicago/Turabian StyleNguyen, Huong Minh, Shinya Watanabe, Sultana Sharmin, Tomofumi Kawaguchi, Xin-Ee Tan, Dhammika Leshan Wannigama, and Longzhu Cui. 2023. "RNA and Single-Stranded DNA Phages: Unveiling the Promise from the Underexplored World of Viruses" International Journal of Molecular Sciences 24, no. 23: 17029. https://doi.org/10.3390/ijms242317029
APA StyleNguyen, H. M., Watanabe, S., Sharmin, S., Kawaguchi, T., Tan, X.-E., Wannigama, D. L., & Cui, L. (2023). RNA and Single-Stranded DNA Phages: Unveiling the Promise from the Underexplored World of Viruses. International Journal of Molecular Sciences, 24(23), 17029. https://doi.org/10.3390/ijms242317029