
Classification of Atretic Small Antral Follicles in the Human Ovary

 , , , ,
, , , ,  ,
,
Abstract
1. Introduction
2. Results
2.1. Characterization of Granulosa Cells in Type 1 Atretic Follicles in Adult Ovaries
2.2. Dynamics in the Theca Cell Layer and Oocyte in Type 1 Atretic Follicles in Adult Ovaries
2.3. ACTA2+ Fibroblast-like Cells Coat the Antrum in Type 2 Atretic Follicles in Adult Ovaries
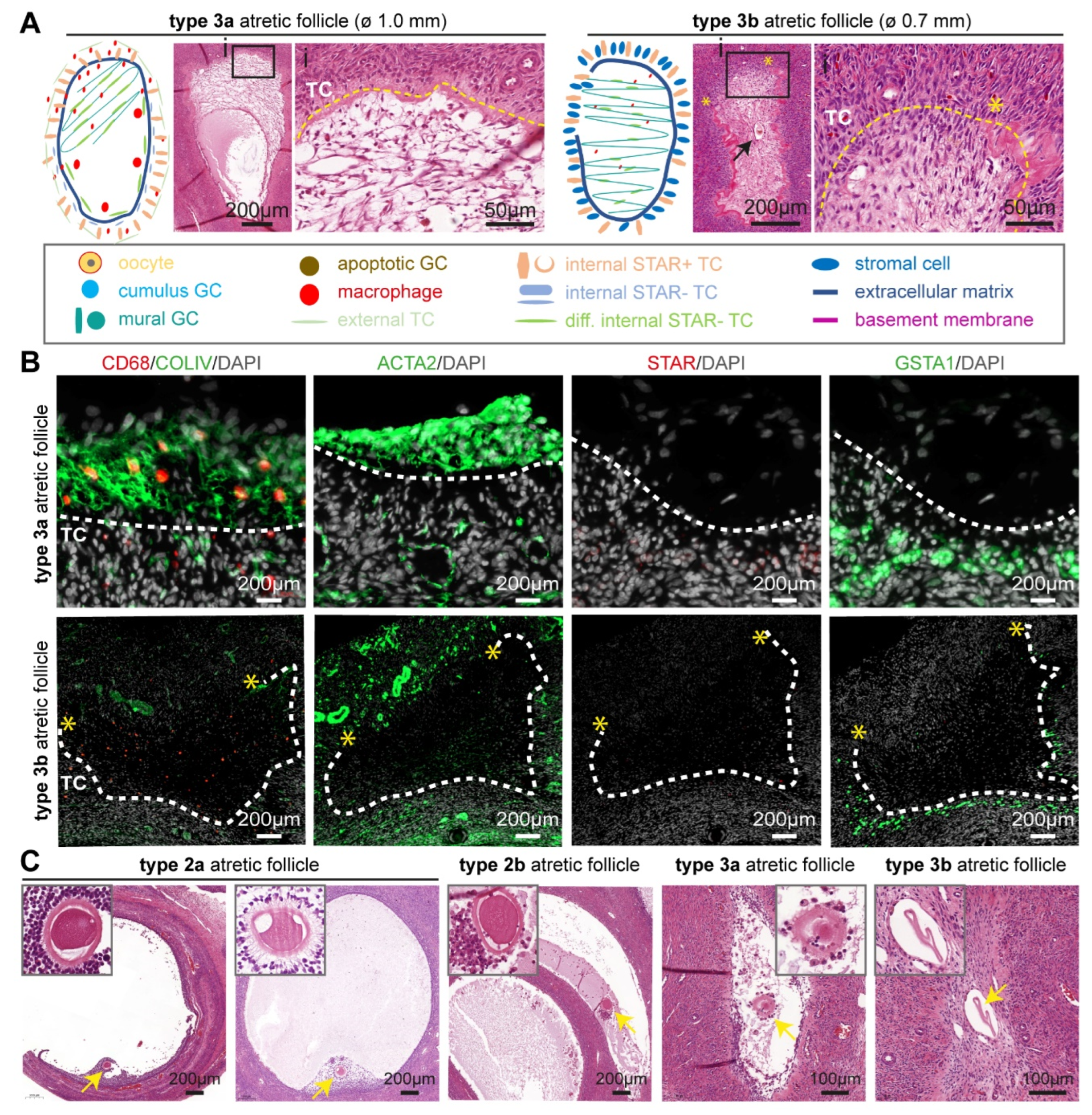
2.4. In Type 3 Atretic Follicles the Antral Cavity Filled and the Oocyte Degenerated
2.5. Quantification of Atretic Human Small Antral Follicles
2.6. Morphological and Molecular Characteristics of Atretic Small Antral Follicles
3. Discussion
4. Materials and Methods
4.1. Study Population and Tissue Collection
4.2. Histology
4.3. Immunofluorescence
4.4. Imaging
4.5. Follicular Measurements and Quantification
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gougeon, A. Human ovarian follicular development: From activation of resting follicles to preovulatory maturation. Ann. Endocrinol. 2010, 71, 132–143. [Google Scholar] [CrossRef]
- McNatty, K.P.; Hillier, S.G.; van den Boogaard, A.M.; Trimbos-Kemper, T.C.; Reichert, L.E., Jr.; van Hall, E.V. Follicular development during the luteal phase of the human menstrual cycle. J. Clin. Endocrinol. Metab. 1983, 56, 1022–1031. [Google Scholar] [CrossRef]
- Tilly, J.L.; Kowalski, K.I.; Johnson, A.L.; Hsueh, A.J. Involvement of apoptosis in ovarian follicular atresia and postovulatory regression. Endocrinology 1991, 129, 2799–2801. [Google Scholar] [CrossRef]
- Vaskivuo, T.E.; Tapanainen, J.S. Apoptosis in the human ovary. Reprod. Biomed. Online 2003, 6, 24–35. [Google Scholar] [CrossRef]
- Irving-Rodgers, H.F.; van Wezel, I.L.; Mussard, M.L.; Kinder, J.E.; Rodgers, R.J. Atresia revisited: Two basic patterns of atresia of bovine antral follicles. Reproduction 2001, 122, 761–775. [Google Scholar] [CrossRef]
- Osman, P. Rate and course of atresia during follicular development in the adult cyclic rat. J. Reprod. Fertil. 1985, 73, 261–270. [Google Scholar] [CrossRef]
- Sugimoto, M.; Manabe, N.; Kimura, Y.; Myoumoto, A.; Imai, Y.; Ohno, H.; Miyamoto, H. Ultrastructural Changes in Granulosa Cells in Porcine Antral Follicles Undergoing Atresia Indicate Apoptotic Cell Death. J. Reprod. Dev. 1998, 44, 7–14. [Google Scholar] [CrossRef]
- Wang, W.; Liu, H.L.; Tian, W.; Zhang, F.F.; Gong, Y.; Chen, J.W.; Mao, D.G.; Shi, F.X. Morphologic observation and classification criteria of atretic follicles in guinea pigs. J. Zhejiang Univ. Sci. B. 2010, 11, 307–314. [Google Scholar] [CrossRef]
- Baerwald, A.R. Human antral folliculogenesis: What we have learned from the bovine and equine models. Anim. Reprod. 2009, 6, 20–29. [Google Scholar]
- Baerwald, A.R.; Adams, G.P.; Pierson, R.A. Ovarian antral folliculogenesis during the human menstrual cycle: A review. Hum. Reprod. Update 2012, 18, 73–91. [Google Scholar] [CrossRef]
- Glamoclija, V.; Vilovic, K.; Saraga-Babic, M.; Baranovic, A.; Sapunar, D. Apoptosis and active caspase-3 expression in human granulosa cells. Fertil. Steril. 2005, 83, 426–431. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Irving-Rodgers, H.F. Morphological classification of bovine ovarian follicles. Reproduction 2010, 139, 309–318. [Google Scholar] [CrossRef]
- Hughes, F.M., Jr.; Gorospe, W.C. Biochemical identification of apoptosis (programmed cell death) in granulosa cells: Evidence for a potential mechanism underlying follicular atresia. Endocrinology 1991, 129, 2415–2422. [Google Scholar] [CrossRef]
- Hussein, M.R. Apoptosis in the ovary: Molecular mechanisms. Hum. Reprod. Update 2005, 11, 162–177. [Google Scholar] [CrossRef]
- Clark, L.J.; Irving-Rodgers, H.F.; Dharmarajan, A.M.; Rodgers, R.J. Theca interna: The other side of bovine follicular atresia. Biol. Reprod. 2004, 71, 1071–1078. [Google Scholar] [CrossRef]
- Maillet, G.; Benhaim, A.; Mittre, H.; Feral, C. Involvement of theca cells and steroids in the regulation of granulosa cell apoptosis in rabbit preovulatory follicles. Reproduction 2003, 125, 709–716. [Google Scholar] [CrossRef]
- Palumbo, A.; Yeh, J. In situ localization of apoptosis in the rat ovary during follicular atresia. Biol. Reprod. 1994, 51, 888–895. [Google Scholar] [CrossRef]
- Morita, Y.; Tilly, J.L. Oocyte apoptosis: Like sand through an hourglass. Dev. Biol. 1999, 213, 1–17. [Google Scholar] [CrossRef]
- Fan, X.; Chuva de Sousa Lopes, S.M. Molecular makeup of the human adult ovary. Opin. Endocrine. Metab. Res. 2021, 18, 187–193. [Google Scholar] [CrossRef]
- Young, J.M.; McNeilly, A.S. Theca: The forgotten cell of the ovarian follicle. Reproduction 2010, 140, 489–504. [Google Scholar] [CrossRef]
- Fan, X.; Bialecka, M.; Moustakas, I.; Lam, E.; Torrens-Juaneda, V.; Borggreven, N.V.; Trouw, L.; Louwe, L.A.; Pilgram, G.S.K.; Mei, H.; et al. Single-cell reconstruction of follicular remodeling in the human adult ovary. Nat. Commun. 2019, 10, 3164. [Google Scholar] [CrossRef] [PubMed]
- Man, L.; Lustgarten-Guahmich, N.; Kallinos, E.; Redhead-Laconte, Z.; Liu, S.; Schattman, B.; Redmond, D.; Hancock, K.; Zaninovic, N.; Schattman, G.; et al. Comparison of Human Antral Follicles of Xenograft versus Ovarian Origin Reveals Disparate Molecular Signatures. Cell. Rep. 2020, 32, 108027. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Huang, Y.; Lin, H. Estrogen disorders: Interpreting the abnormal regulation of aromatase in granulosa cells (Review). Int. J. Mol. Med. 2021, 47, 73. [Google Scholar] [CrossRef] [PubMed]
- Themmen, A.P. Anti-Mullerian hormone: Its role in follicular growth initiation and survival and as an ovarian reserve marker. J. Natl. Cancer Inst. Monogr. 2005, 2005, 18–21. [Google Scholar] [CrossRef]
- Walters, K.A.; Allan, C.M.; Handelsman, D.J. Androgen actions and the ovary. Biol. Reprod. 2008, 78, 380–389. [Google Scholar] [CrossRef]
- Richards, J.S.; Ren, Y.A.; Candelaria, N.; Adams, J.E.; Rajkovic, A. Ovarian Follicular Theca Cell Recruitment, Differentiation, and Impact on Fertility: 2017 Update. Endocr. Rev. 2018, 39, 1–20. [Google Scholar] [CrossRef]
- Rabahi, F.; Brule, S.; Sirois, J.; Beckers, J.F.; Silversides, D.W.; Lussier, J.G. High expression of bovine alpha glutathione S-transferase (GSTA1, GSTA2) subunits is mainly associated with steroidogenically active cells and regulated by gonadotropins in bovine ovarian follicles. Endocrinology 1999, 140, 3507–3517. [Google Scholar] [CrossRef]
- Gordon, S.; Pluddemann, A. Macrophage Clearance of Apoptotic Cells: A Critical Assessment. Front. Immunol. 2018, 9, 127. [Google Scholar] [CrossRef]
- Marion, G.B.; Gier, H.T.; Choudary, J.B. Micromorphology of the bovine ovarian follicular system. J. Anim. Sci. 1968, 27, 451–465. [Google Scholar] [CrossRef]
- Garcia, R.; Ballesteros, L.M.; Hernandez-Perez, O.; Rosales, A.M.; Espinosa, R.; Soto, H.; Diaz de Leon, L.; Rosado, A. Metalloproteinase activity during growth, maturation and atresia in the ovarian follicles of the goat. Anim. Reprod. Sci. 1997, 47, 211–228. [Google Scholar] [CrossRef]
- Byskov, A.G. Cell kinetic studies of follicular atresia in the mouse ovary. J. Reprod. Fertil. 1974, 37, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.G.; Ushizawa, K.; Hosoe, M.; Takahashi, T. Differential genome-wide gene expression profiling of bovine largest and second-largest follicles: Identification of genes associated with growth of dominant follicles. Reprod. Biol. Endocrinol. 2010, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Mlodawska, W.; Slomczynska, M. Immunohistochemical localization of aromatase during the development and atresia of ovarian follicles in prepubertal horses. Theriogenology 2010, 74, 1707–1712. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Watanabe, H.; Saito, H.; Hiroi, M.; Tonosaki, A. Elimination of atretic follicles from the mouse ovary: A TEM and immunohistochemical study in mice. J. Anat. 2000, 196, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.H.; Hales, D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef]
- Massett, M.P.; Bywaters, B.C.; Gibbs, H.C.; Trzeciakowski, J.P.; Padgham, S.; Chen, J.; Rivera, G.; Yeh, A.T.; Milewicz, D.M.; Trache, A. Loss of smooth muscle alpha-actin effects on mechanosensing and cell-matrix adhesions. Exp. Biol. Med. 2020, 245, 374–384. [Google Scholar] [CrossRef]
- Ruiz-Zapata, A.M.; Heinz, A.; Kerkhof, M.H.; van de Westerlo-van Rijt, C.; Schmelzer, C.E.H.; Stoop, R.; Kluivers, K.B.; Oosterwijk, E. Extracellular Matrix Stiffness and Composition Regulate the Myofibroblast Differentiation of Vaginal Fibroblasts. Int. J. Mol. Sci. 2020, 21, 4762. [Google Scholar] [CrossRef]
- Kinnear, H.M.; Tomaszewski, C.E.; Chang, F.L.; Moravek, M.B.; Xu, M.; Padmanabhan, V.; Shikanov, A. The ovarian stroma as a new frontier. Reproduction 2020, 160, R25–R39. [Google Scholar] [CrossRef]
- Kasuya, K. Elimination of apoptotic granulosa cells by intact granulosa cells and macrophages in atretic mature follicles of the guinea pig ovary. Arch. Histol. Cytol. 1997, 60, 175–184. [Google Scholar] [CrossRef]
- Lichtnekert, J.; Kawakami, T.; Parks, W.C.; Duffield, J.S. Changes in macrophage phenotype as the immune response evolves. Curr. Opin. Pharmacol. 2013, 13, 555–564. [Google Scholar] [CrossRef]
- Tarique, A.A.; Logan, J.; Thomas, E.; Holt, P.G.; Sly, P.D.; Fantino, E. Phenotypic, functional, and plasticity features of classical and alternatively activated human macrophages. Am. J. Respir. Cell. Mol. Biol. 2015, 53, 676–688. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue Features of Small Antral Follicles | Healthy | Type 1 | Type 2 | Type 3 | |||
|---|---|---|---|---|---|---|---|
| Type 2a | Type 2b | Type 3a | Type 3b | ||||
| Follicle | total number of follicles analysed | 47 | 59 | 49 | 72 | 51 | 225 |
| shape of follicle | round/ellipsoid | round/ellipsoid | ellipsoid/irregular | ellipsoid/irregular | irregular | irregular | |
| size of follicle ± SD (cm) (size range) | 2.2 ± 1.3 (0.8–5.0) | 2.0 ± 0.8 (0.9–4.3) | 1.6 ± 0.7 (0.5–3.6) | 1.2 ± 0.7 (0.15–2.7) | 0.5 ± 0.4 (0.08–2.1) | 0.2 ± 0.1 (0.02–0.25) | |
| Mural Granulosa Cells (mGC) | number of mGC layers ± SD (number range) | 4.2 ± 1.1 (2–10) | 3.4 ± 1.1 (1–9) | 2.2 ± 0.9 (0–8) | 0 | 0 | 0 |
| thickness of mGC ± SD (μm) (size range) | 32.5 ± 12.0 (10.0–91.1) | 25.5 ± 10.4 (4.0–89.0) | 16.5 ± 7.2 (0.0–75.0) | 0 | 0 | 0 | |
| shape of mGC in contact with BM | round | round/columnar | columnar/flat | n/a | n/a | n/a | |
| apoptotic index (% TUNEL+ mGCs) ± SD (index range) | 1.1 ± 1.5 (0.0–3.9) | 21.9 ± 10.1 (6.0–32.7) | 17.3 ± 10.6 (6.0–38.0) | 78.6 ± 5.9 (in cavity) (75.0–81.8) | 0 | 0 | |
| mitotic index (% MIK67+ mGCs) ± SD (index range) | 31.5 ± 3.9 (26.6–40) | 14.4 ± 6.8 (7.3–25) | 4.6 ± 2.0 (2.3–6.7) | 0 | 0 | 0 | |
| Basement Membrane (BM) | BM integrity | intact | intact | not intact | not intact | ECM deposition | ECM deposition |
| thickness of BM | + | + | ++ | ++ | +++ | +++ | |
| Internal Theca Cells (iTC) | number of (GSTA1+) iTC layers ± SD (number range) | 1.9 ± 0.6 (1–4) | 3.7 ± 1.5 (1–7) | 4.8 ± 1.4 (2–10) | 3.3 ± 1.1 (2–8) | 2.5 ± 0.6 (2–5) | 1.4 ± 1.3 (0–4) |
| thickness of (GSTA1+) iTC ± SD (μm) (size range) | 16.1 ± 6.0 (8.0–32.9) | 34.2 ± 13.6 (8.0–75.1) | 39.3 ± 16.8 (15.0–91.9) | 27.9 ± 8.7 (15.0–62.3) | 24.9 ± 5.0 (18.5–40.8) | 12.8 ± 11.6 (0–5) | |
| shape of iTC | ellipsoid | round (hyaline) | round (hyaline) | round (hyaline) | elongated | elongated | |
| orientation of (GSTA1+) iTC relative to BM | parallel | parallel | parallel | parallel | perpendicular | perpendicular | |
| Macrophages | accumulation | ovarian stroma | iTC layer | follicular cavity | follicular cavity | connective tissue in follicular cavity | connective tissue in follicular cavity |
| Oocyte | shape of zona pellucida | round | round | round/irregular | round/irregular | irregular | irregular |
| presence of oocyte | yes | yes | yes | yes | no | no | |
| Cell Type Markers Used | Healthy | Type 1 | Type 2 | Type 3a | |||
|---|---|---|---|---|---|---|---|
| Type 2a | Type 2b | Type 3a | Type 3b | ||||
| Mural Granulosa Cells (mGC) | MKI67 | +++ | + | +/− | − | − | − |
| cCASP3 | − | ++ | ++ | ++ (in cavity) | − | − | |
| TUNEL | − | ++ | ++ | ++ (in cavity) | − | − | |
| CYP19A1 | +++ | + | − | − | − | − | |
| AMH | +++ | + | +/− | − | − | − | |
| AR | ++ | ++ | +/− | − | − | − | |
| CDH1 | +++ | + | − | − | − | − | |
| GSTA1 | ++ | ++ | +/− | − | − | − | |
| Internal Theca Cells (iTC) | COLIV | + | ++ | ++ | ++ | ++/− | − |
| ACTA2 | − | − | ++/− | ++ | +++/+ | + | |
| STAR | +++ | ++ | + | +/− | +/− | +/− | |
| GSTA1 | +++ | +++ | +++ | +++ | ++ | ++ | |
| External Theca Cells (eTC) | ACTA2 | +++ | ++ | ++/+ | + | + | + |
| Macrophages | CD68 | +/− | ++ | +++ | +++ | ++ | ++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, F.; Fan, X.; del Valle, J.S.; Asseler, J.D.; van der Meeren, L.E.; Cheng, H.; Roelen, B.A.J.; Louwe, L.A.; Pilgram, G.S.K.; van der Westerlaken, L.A.J.; et al. Classification of Atretic Small Antral Follicles in the Human Ovary. Int. J. Mol. Sci. 2023, 24, 16846. https://doi.org/10.3390/ijms242316846
Wei F, Fan X, del Valle JS, Asseler JD, van der Meeren LE, Cheng H, Roelen BAJ, Louwe LA, Pilgram GSK, van der Westerlaken LAJ, et al. Classification of Atretic Small Antral Follicles in the Human Ovary. International Journal of Molecular Sciences. 2023; 24(23):16846. https://doi.org/10.3390/ijms242316846
Chicago/Turabian StyleWei, Fu, Xueying Fan, Julieta S. del Valle, Joyce D. Asseler, Lotte E. van der Meeren, Hui Cheng, Bernard A. J. Roelen, Leoni A. Louwe, Gonneke S. K. Pilgram, Lucette A. J. van der Westerlaken, and et al. 2023. "Classification of Atretic Small Antral Follicles in the Human Ovary" International Journal of Molecular Sciences 24, no. 23: 16846. https://doi.org/10.3390/ijms242316846
APA StyleWei, F., Fan, X., del Valle, J. S., Asseler, J. D., van der Meeren, L. E., Cheng, H., Roelen, B. A. J., Louwe, L. A., Pilgram, G. S. K., van der Westerlaken, L. A. J., van Mello, N. M., & Chuva de Sousa Lopes, S. M. (2023). Classification of Atretic Small Antral Follicles in the Human Ovary. International Journal of Molecular Sciences, 24(23), 16846. https://doi.org/10.3390/ijms242316846