
Mitochondrial Dysfunction in Skeletal Muscle of Rotenone-Induced Rat Model of Parkinson’s Disease: SC-Nanophytosomes as Therapeutic Approach

 ,
,  , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. In Vivo Assays—Benefits of SC-Nanophytosomes Formulation on Associated Motor Symptoms of the Rotenone-Induced Rat Model of PD
2.2. Ex Vivo Assays in Skeletal Muscle Tissues-Effects on Mitochondrial Redox Chain Complex Functionality, Cell Redox State and Fatty Acid Profile
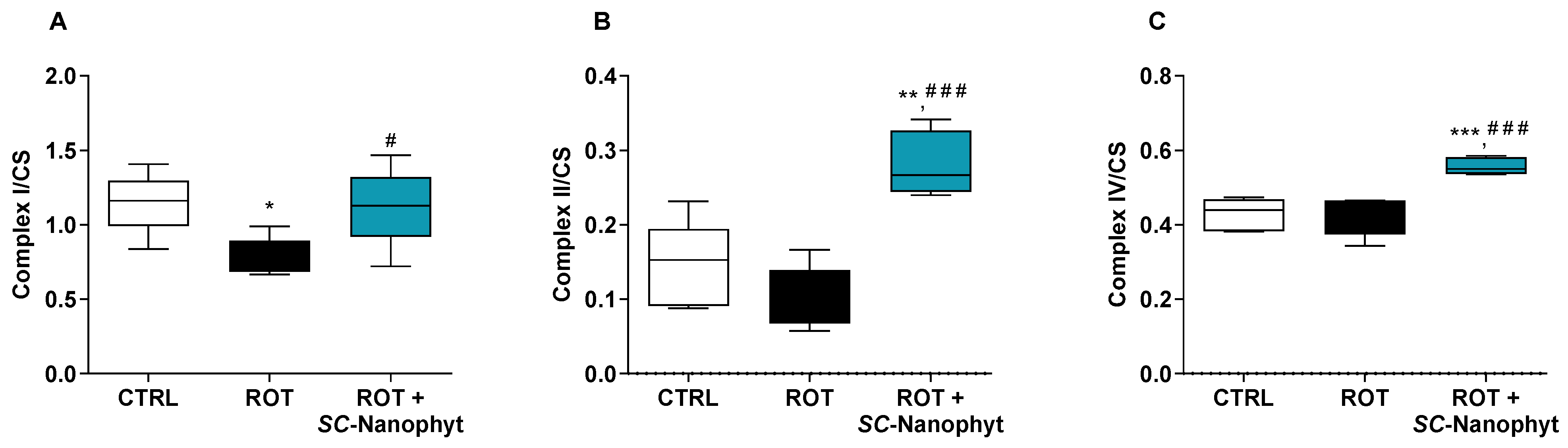
2.2.1. Effects on the Activity of Mitochondrial Redox Chain Complexes
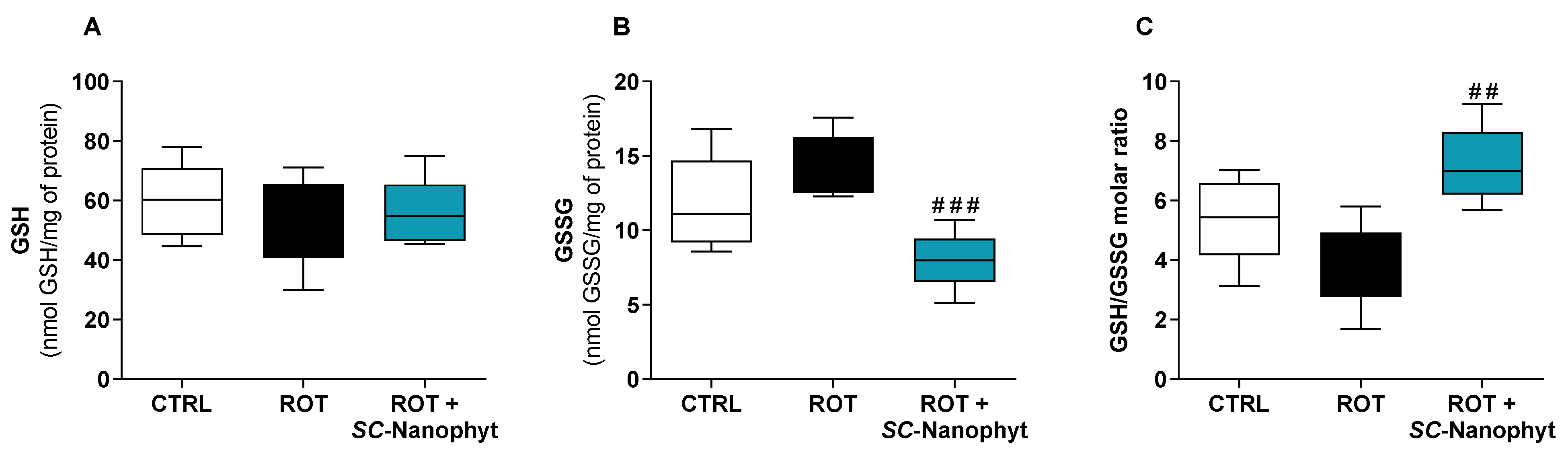
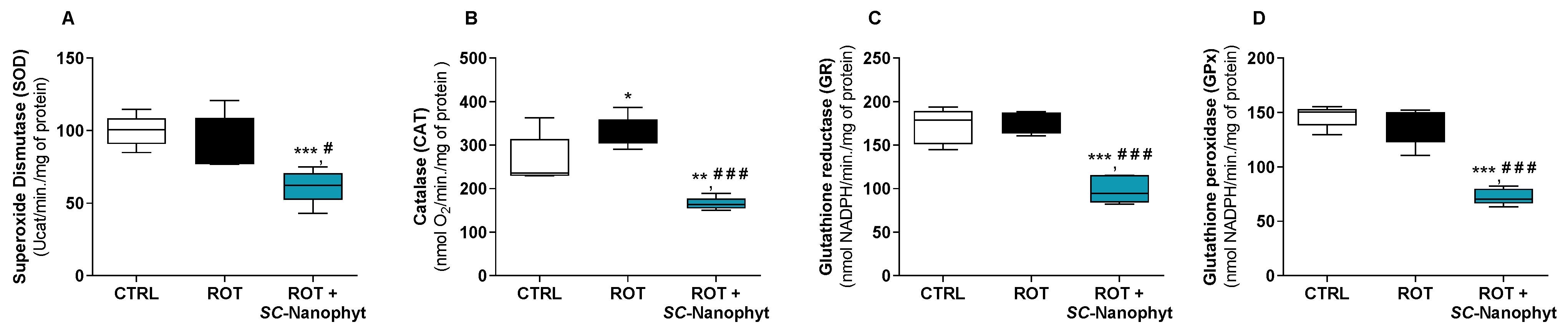
2.2.2. Effects on the Cell Redox State
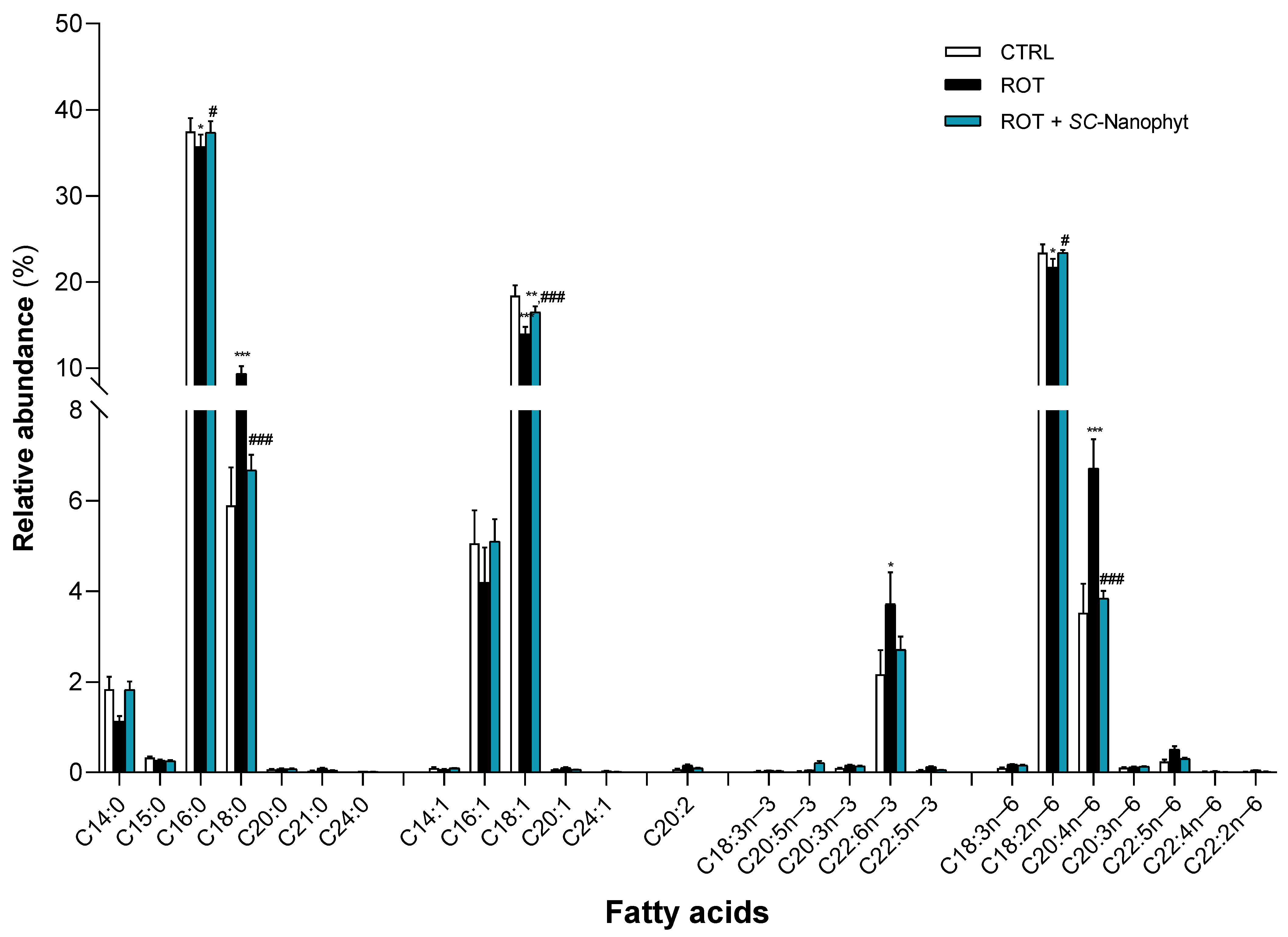
2.2.3. Effects on the Fatty Acid Profile of the Skeletal-Muscle Tissue
3. Materials and Methods
3.1. SC-Nanophytosome Preparation and Characterization
3.2. Rotenone-Induced Rat Model of PD, Treatment with SC-Nanophytosomes and the Beam Walking Test
3.3. Ex Vivo Studies: Skeletal-Muscle Tissues Processing for Bioenergetic, Cell Redox State and Lipidomic Assays
3.4. Assessment of the Mitochondrial Respiratory Chain Complex Activities
3.5. Determination of Enzymatic and Non-Enzymatic Antioxidant Defences in Skeletal Muscle Mitochondria-Free Cytosolic Fraction
3.6. Analysis of Fatty Acid Methyl Esters from Skeletal Muscle Tissues
3.7. Data and Statistical Analysis
4. Conclusions
SC-Nanophytosomes as Therapeutic Tool for PD—Limitations and Future Line of Research
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winklhofer, K.F.; Haass, C. Mitochondrial dysfunction in Parkinson’s disease. Biochim. et Biophys. Acta (BBA)-Mol. Basis Dis. 2010, 1802, 29–44. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A.H.V. Parkinson disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Poston, K.L.; Eidelberg, D. Functional brain networks and abnormal connectivity in the movement disorders. NeuroImage 2012, 62, 2261–2270. [Google Scholar] [CrossRef]
- Dezsi, L.; Vecsei, L. Monoamine Oxidase B Inhibitors in Parkinson’s Disease. CNS Neurol. Disord. - Drug Targets 2017, 16, 425–439. [Google Scholar] [CrossRef]
- Tambasco, N.; Romoli, M.; Calabresi, P. Levodopa in Parkinson’s Disease: Current Status and Future Developments. Curr. Neuropharmacol. 2018, 16, 1239–1252. [Google Scholar] [CrossRef] [PubMed]
- Schneider, L.S.; Mangialasche, F.; Andreasen, N.; Feldman, H.; Giacobini, E.; Jones, R.; Mantua, V.; Mecocci, P.; Pani, L.; Winblad, B.; et al. Clinical trials and late-stage drug development for Alzheimer’s disease: An appraisal from 1984 to 2014. J. Intern. Med. 2014, 275, 251–283. [Google Scholar] [CrossRef] [PubMed]
- Fields, C.R.; Bengoa-Vergniory, N.; Wade-Martins, R. Targeting Alpha-Synuclein as a Therapy for Parkinson’s Disease. Front. Mol. Neurosci. 2019, 12, 299. [Google Scholar] [CrossRef]
- Sherer, T.B.; Betarbet, R.; Testa, C.M.; Seo, B.B.; Richardson, J.R.; Kim, J.H.; Miller, G.W.; Yagi, T.; Matsuno-Yagi, A.; Greenamyre, J.T. Mechanism of Toxicity in Rotenone Models of Parkinson’s Disease. J. Neurosci. 2003, 23, 10756–10764. [Google Scholar] [CrossRef]
- Richardson, J.R.; Caudle, W.M.; Guillot, T.S.; Watson, J.L.; Nakamaru-Ogiso, E.; Seo, B.B.; Sherer, T.B.; Greenamyre, J.T.; Yagi, T.; Matsuno-Yagi, A.; et al. Obligatory Role for Complex I Inhibition in the Dopaminergic Neurotoxicity of 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). Toxicol. Sci. 2006, 95, 196–204. [Google Scholar] [CrossRef]
- Mendes, D.; Valentão, P.; Oliveira, M.M.; Andrade, P.; Videira, R.A. A nanophytosomes formulation based on elderberry anthocyanins and Codium lipids to mitigate mitochondrial dysfunctions. Biomed. Pharmacother. 2021, 143, 112157. [Google Scholar] [CrossRef]
- da Costa, E.; Melo, T.; Moreira, A.S.; Alves, E.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Decoding bioactive polar lipid profile of the macroalgae Codium tomentosum from a sustainable IMTA system using a lipidomic approach. Algal Res. 2015, 12, 388–397. [Google Scholar] [CrossRef]
- Haines, T.H.A.; Dencher, N. Cardiolipin: A proton trap for oxidative phosphorylation. FEBS Lett. 2002, 528, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Stanacev, N.Z.; Chang, Y.-Y.; Kennedy, E.P. Biosynthesis of Cardiolipin in Escherichia coli. J. Biol. Chem. 1967, 242, 3018–3019. [Google Scholar] [CrossRef]
- Bazinet, R.P.; Layé, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nat. Rev. Neurosci. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Neves, D.; Valentão, P.; Bernardo, J.; Oliveira, M.C.; Ferreira, J.M.; Pereira, D.M.; Andrade, P.B.; Videira, R.A. A new insight on elderberry anthocyanins bioactivity: Modulation of mitochondrial redox chain functionality and cell redox state. J. Funct. Foods 2019, 56, 145–155. [Google Scholar] [CrossRef]
- Mendes, D.; Peixoto, F.; Oliveira, M.M.; Andrade, P.B.; Videira, R.A. Brain Effects of SC-Nanophytosomes on a Rotenone-Induced Rat Model of Parkinson’s Disease—A Proof of Concept for a Mitochondria-Targeted Therapy. Int. J. Mol. Sci. 2022, 23, 12699. [Google Scholar] [CrossRef]
- Magaña, J.C.; Deus, C.M.; Giné-Garriga, M.; Montané, J.; Pereira, S.P. Exercise-Boosted Mitochondrial Remodeling in Parkinson’s Disease. Biomedicines 2022, 10, 3228. [Google Scholar] [CrossRef]
- Blin, O.; Desnuelle, C.; Rascol, O.; Borg, M.; Peyro Saint Paul, H.; Azulay, J.-P.; Billè, F.; Figarella, D.; Coulom, F.; Pellissier, J.F.; et al. Mitochondrial respiratory failure in skeletal muscle from patients with Parkinson’s disease and multiple system atrophy. J. Neurol. Sci. 1994, 125, 95–101. [Google Scholar] [CrossRef]
- Bindoff, L.; Birch-Machin, M.; Cartlidge, N.; Parker, W.; Turnbull, D. Respiratory chain abnormalities in skeletal muscle from patients with Parkinson’s disease. J. Neurol. Sci. 1991, 104, 203–208. [Google Scholar] [CrossRef]
- Burtscher, J.; Millet, G.P.; Place, N.; Kayser, B.; Zanou, N. The Muscle-Brain Axis and Neurodegenerative Diseases: The Key Role of Mitochondria in Exercise-Induced Neuroprotection. Int. J. Mol. Sci. 2021, 22, 6479. [Google Scholar] [CrossRef] [PubMed]
- Nay, K.; Smiles, W.J.; Kaiser, J.; McAloon, L.M.; Loh, K.; Galic, S.; Oakhill, J.S.; Gundlach, A.L.; Scott, J.W. Molecular Mechanisms Underlying the Beneficial Effects of Exercise on Brain Function and Neurological Disorders. Int. J. Mol. Sci. 2021, 22, 4052. [Google Scholar] [CrossRef] [PubMed]
- Hallett, M.; Hess, C.W. The Phenomenology of Parkinson’s Disease. Semin. Neurol. 2017, 37, 109–117. [Google Scholar] [CrossRef]
- Chen, M.; Wang, Y.; Deng, S.; Lian, Z.; Yu, K. Skeletal muscle oxidative stress and inflammation in aging: Focus on antioxidant and anti-inflammatory therapy. Front. Cell Dev. Biol. 2022, 10, 964130. [Google Scholar] [CrossRef]
- Ademowo, O.S.; Dias, H.K.I.; Burton, D.G.A.; Griffiths, H.R. Lipid (per) oxidation in mitochondria: An emerging target in the ageing process? Biogerontology 2017, 18, 859–879. [Google Scholar] [CrossRef] [PubMed]
- Rosen, K.M.; Veereshwarayya, V.; Moussa, C.E.-H.; Fu, Q.; Goldberg, M.S.; Schlossmacher, M.G.; Shen, J.; Querfurth, H.W. Parkin Protects against Mitochondrial Toxins and beta-Amyloid Accumulation in Skeletal Muscle Cells. J. Biol. Chem. 2006, 281, 12809–12816. [Google Scholar] [CrossRef]
- Büeler, H. Impaired mitochondrial dynamics and function in the pathogenesis of Parkinson’s disease. Exp. Neurol. 2009, 218, 235–246. [Google Scholar] [CrossRef]
- Li, H.; Sharma, L.K.; Li, Y.; Hu, P.; Idowu, A.; Liu, D.; Lu, J.; Bai, Y. Comparative bioenergetic study of neuronal and muscle mitochondria during aging. Free. Radic. Biol. Med. 2013, 63, 30–40. [Google Scholar] [CrossRef]
- Masella, R.; Di Benedetto, R.; Varì, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ J. 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef]
- Yap, L.-P.; Garcia, J.V.; Han, D.; Cadenas, E. The energy–redox axis in aging and age-related neurodegeneration. Adv. Drug Deliv. Rev. 2009, 61, 1283–1298. [Google Scholar] [CrossRef]
- Galper, J.; Dean, N.J.; Pickford, R.; Lewis, S.J.G.; Halliday, G.M.; Kim, W.S.; Dzamko, N. Lipid pathway dysfunction is prevalent in patients with Parkinson’s disease. Brain 2022, 145, 3472–3487. [Google Scholar] [CrossRef] [PubMed]
- Fernandis, A.Z.; Wenk, M.R. Membrane lipids as signaling molecules. Curr. Opin. Infect. Dis. 2007, 18, 121–128. [Google Scholar] [CrossRef]
- Xicoy, H.; Wieringa, B.; Martens, G.J.M. The Role of Lipids in Parkinson’s Disease. Cells 2019, 8, 27. [Google Scholar] [CrossRef]
- Mårtensson, C.U.; Doan, K.N.; Becker, T. Effects of lipids on mitochondrial functions. Biochim. et Biophys. Acta (BBA) - Mol. Cell Biol. Lipids 2017, 1862, 102–113. [Google Scholar] [CrossRef]
- Abbott, S.K.; Else, P.L.; Hulbert, A.J. Membrane fatty acid composition of rat skeletal muscle is most responsive to the balance of dietaryn-3 andn-6 PUFA. Br. J. Nutr. 2009, 103, 522–529. [Google Scholar] [CrossRef]
- Luong, T.N.; Carlisle, H.J.; Southwell, A.; Patterson, P.H. Assessment of Motor Balance and Coordination in Mice using the Balance Beam. J. Vis. Exp. 2011, 10, e2376. [Google Scholar] [CrossRef]
- Mendes, D.; Oliveira, M.M.; Moreira, P.I.; Coutinho, J.; Nunes, F.M.; Pereira, D.M.; Valentão, P.; Andrade, P.B.; Videira, R.A. Beneficial effects of white wine polyphenols-enriched diet on Alzheimer’s disease-like pathology. J. Nutr. Biochem. 2018, 55, 165–177. [Google Scholar] [CrossRef]
- Mendes, D.; Silva, A.M.; Oliveira, M.M.; Andrade, P.B.; Videira, R.A. An Experimental Approach to Address the Functional Relationship between Antioxidant Enzymes and Mitochondrial Respiratory Complexes. Methods Protoc. 2023, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, E.; Lewis, D. Spectrophotometric determination of phosphate esters in the presence and absence of orthophosphate. Anal. Biochem. 1970, 36, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, F.; Vicente, J.; Madeira, V.M. A comparative study of plant and animal mitochondria exposed to paraquat reveals that hydrogen peroxide is not related to the observed toxicity. Toxicol. Vitr. 2004, 18, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Severinsen, M.C.K.; Pedersen, B.K. Muscle–Organ Crosstalk: The Emerging Roles of Myokines. Endocr. Rev. 2020, 41, 594–609. [Google Scholar] [CrossRef]
- Mendes, D.; Peixoto, F.; Oliveira, M.M.; Andrade, P.B.; Videira, R.A. Mitochondria research and neurodegenerative diseases: On the track to understanding the biological world of high complexity. Mitochondrion 2022, 65, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Ekstrand, M.I.; Galter, D. The MitoPark Mouse—An animal model of Parkinson’s disease with impaired respiratory chain function in dopamine neurons. Park. Relat. Disord. 2009, 15, S185–S188. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendes, D.; Peixoto, F.; Oliveira, M.M.; Andrade, P.B.; Videira, R.A. Mitochondrial Dysfunction in Skeletal Muscle of Rotenone-Induced Rat Model of Parkinson’s Disease: SC-Nanophytosomes as Therapeutic Approach. Int. J. Mol. Sci. 2023, 24, 16787. https://doi.org/10.3390/ijms242316787
Mendes D, Peixoto F, Oliveira MM, Andrade PB, Videira RA. Mitochondrial Dysfunction in Skeletal Muscle of Rotenone-Induced Rat Model of Parkinson’s Disease: SC-Nanophytosomes as Therapeutic Approach. International Journal of Molecular Sciences. 2023; 24(23):16787. https://doi.org/10.3390/ijms242316787
Chicago/Turabian StyleMendes, Daniela, Francisco Peixoto, Maria Manuel Oliveira, Paula Branquinho Andrade, and Romeu António Videira. 2023. "Mitochondrial Dysfunction in Skeletal Muscle of Rotenone-Induced Rat Model of Parkinson’s Disease: SC-Nanophytosomes as Therapeutic Approach" International Journal of Molecular Sciences 24, no. 23: 16787. https://doi.org/10.3390/ijms242316787
APA StyleMendes, D., Peixoto, F., Oliveira, M. M., Andrade, P. B., & Videira, R. A. (2023). Mitochondrial Dysfunction in Skeletal Muscle of Rotenone-Induced Rat Model of Parkinson’s Disease: SC-Nanophytosomes as Therapeutic Approach. International Journal of Molecular Sciences, 24(23), 16787. https://doi.org/10.3390/ijms242316787