
Discovery of Anti-Coronavirus Cinnamoyl Triterpenoids Isolated from Hippophae rhamnoides during a Screening of Halophytes from the North Sea and Channel Coasts in Northern France

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
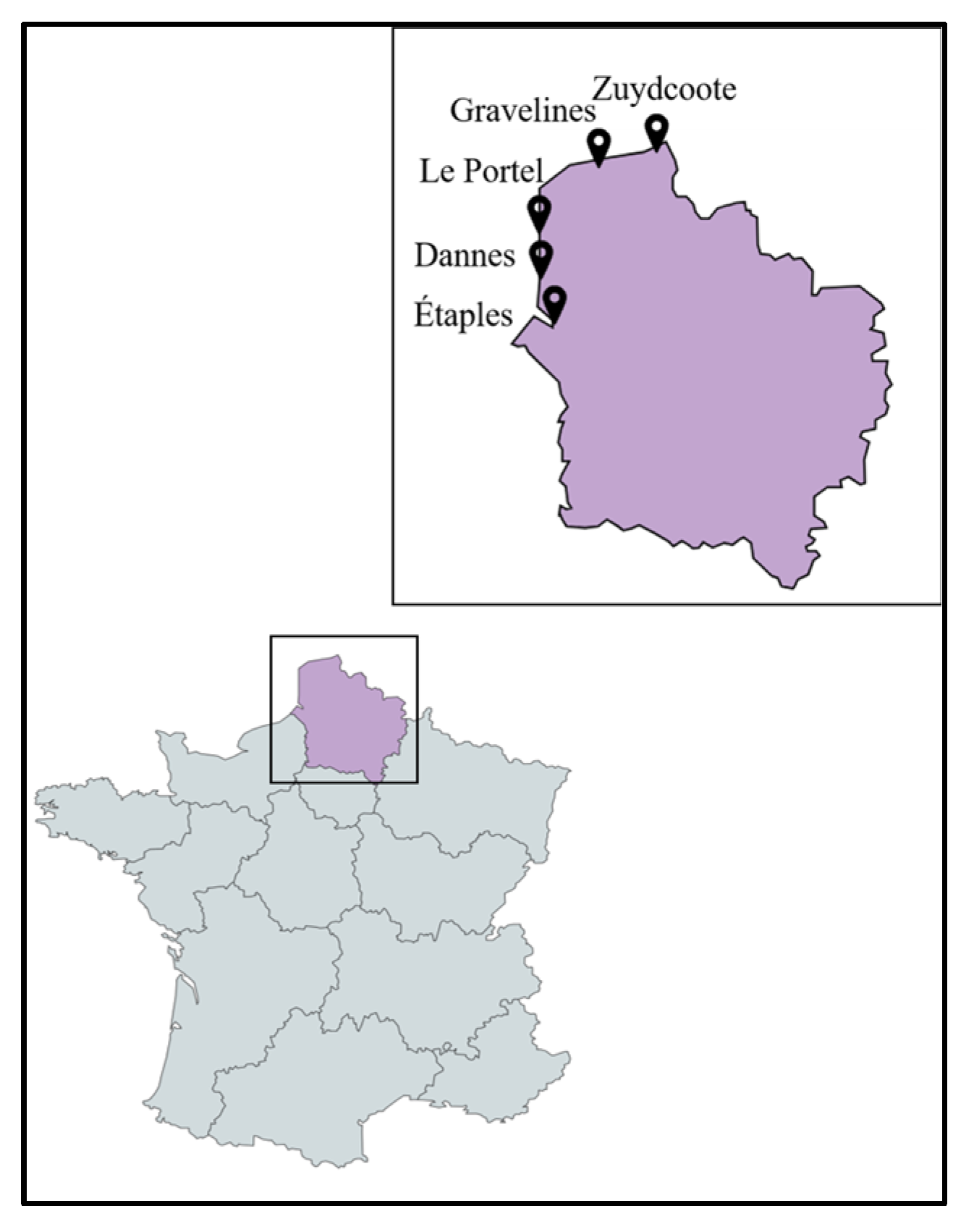
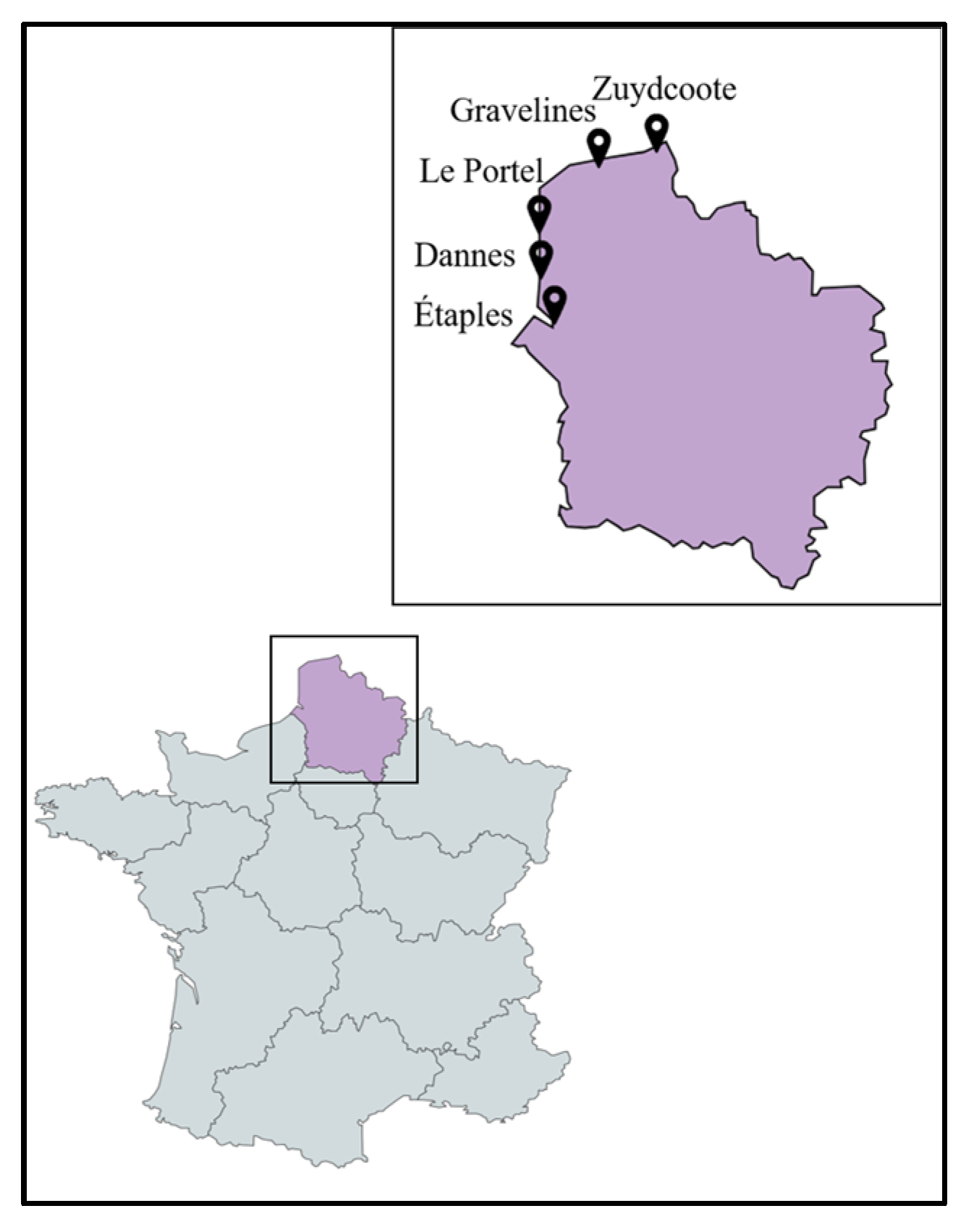
2.1. Sampling and Classification of the Collected Plant Species
2.2. Cytotoxicity and Antiviral Activity of Plant Crude Methanolic Extracts
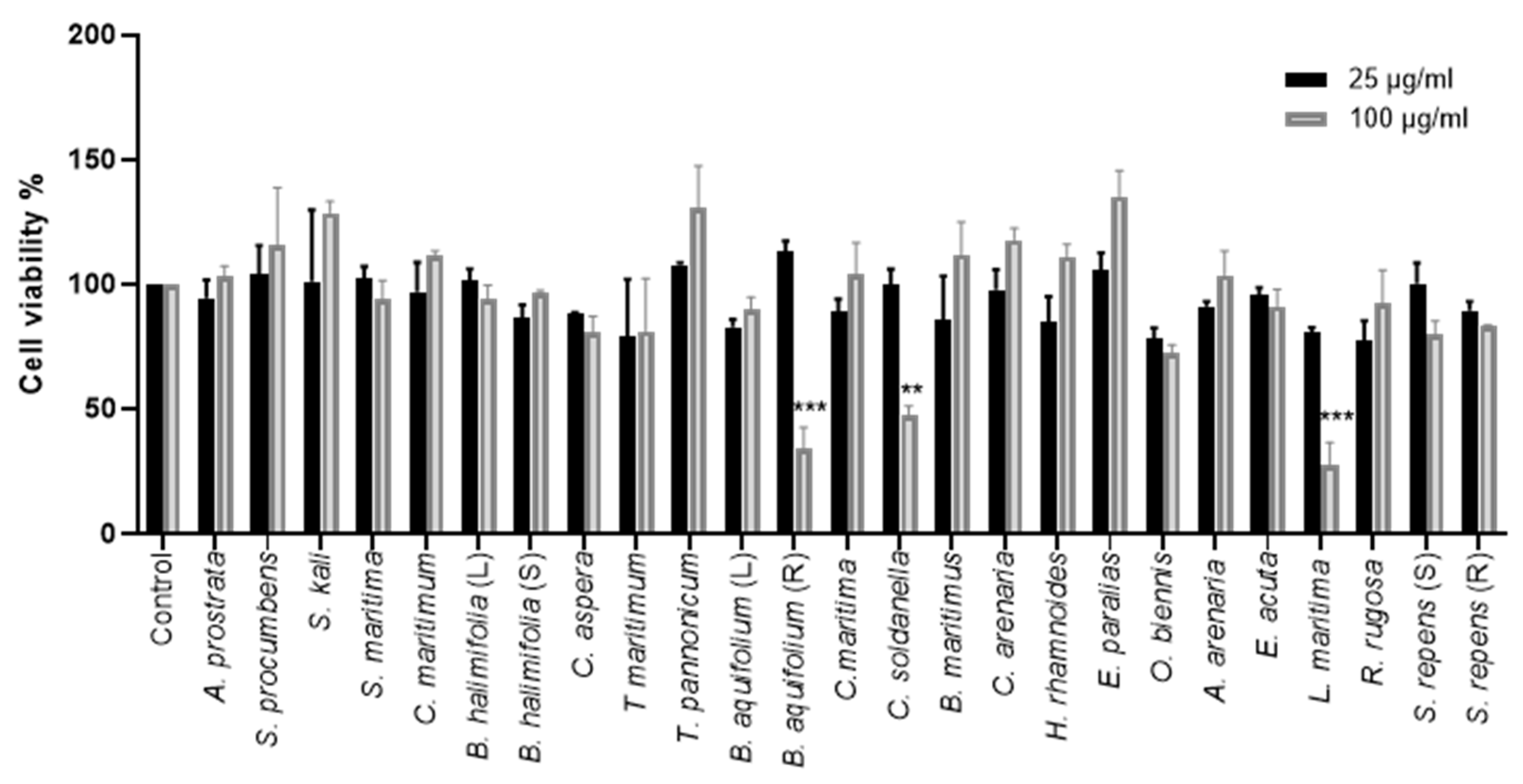
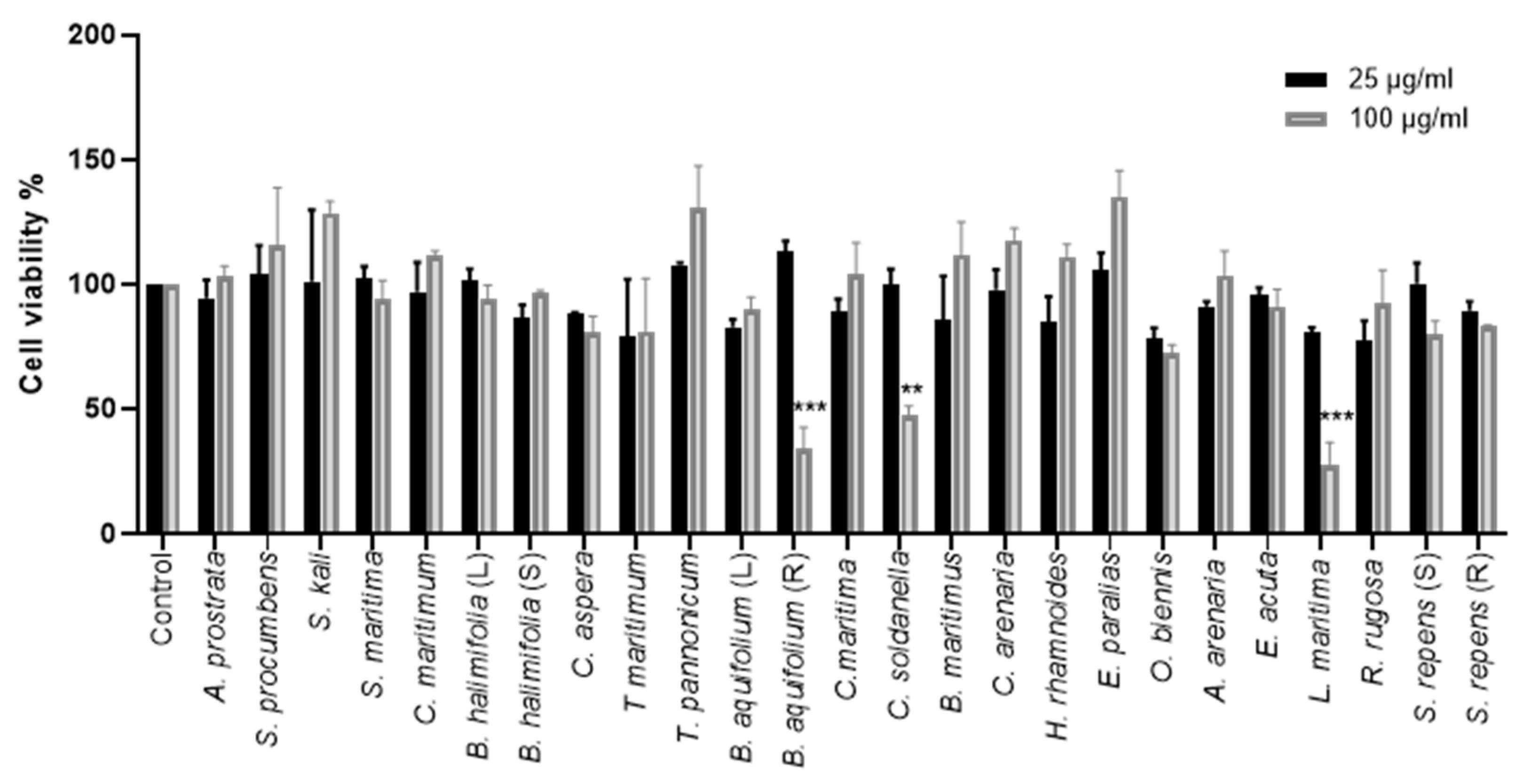
2.2.1. Effect of Crude Methanolic Extracts on Cell Viability
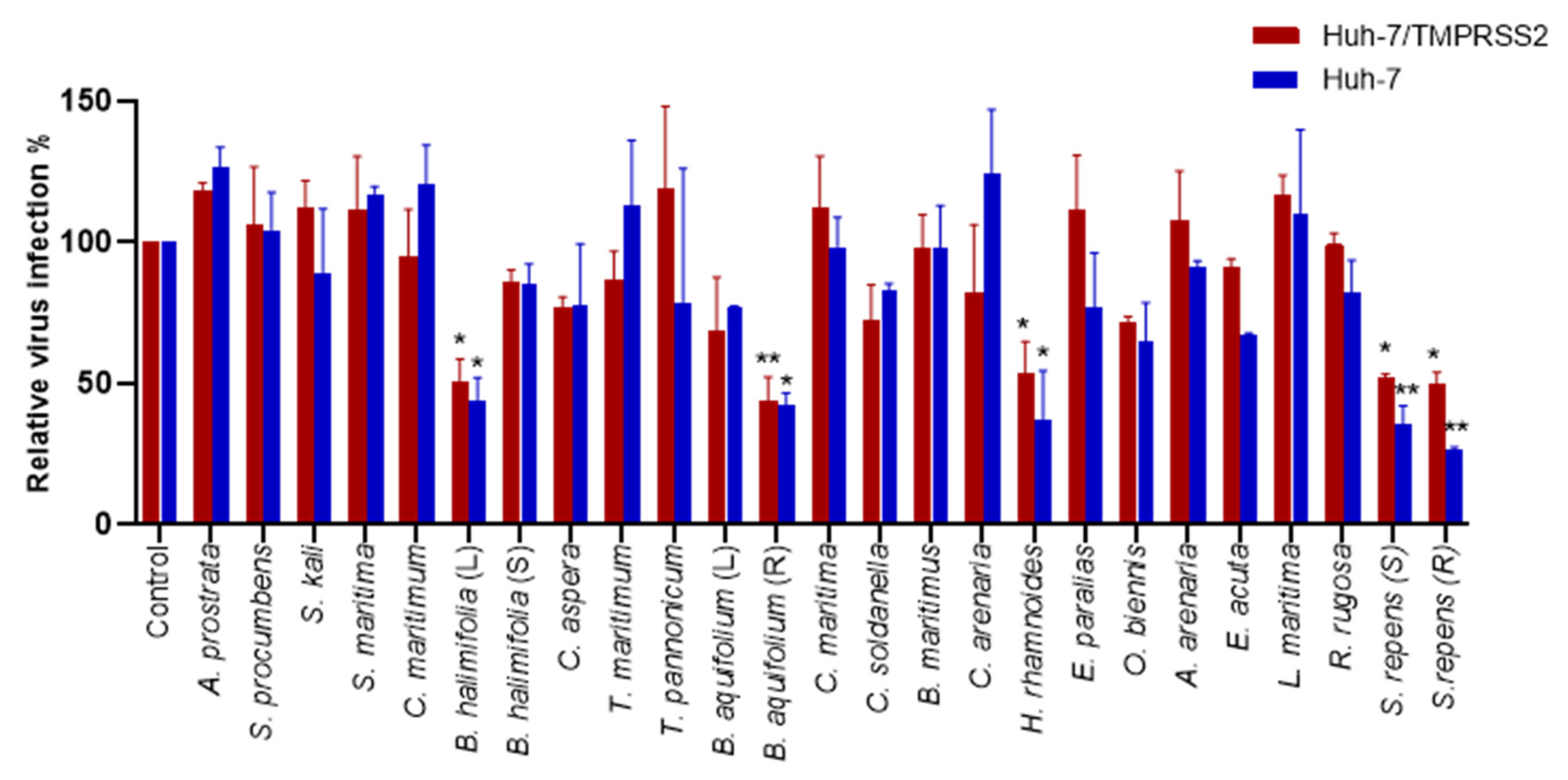
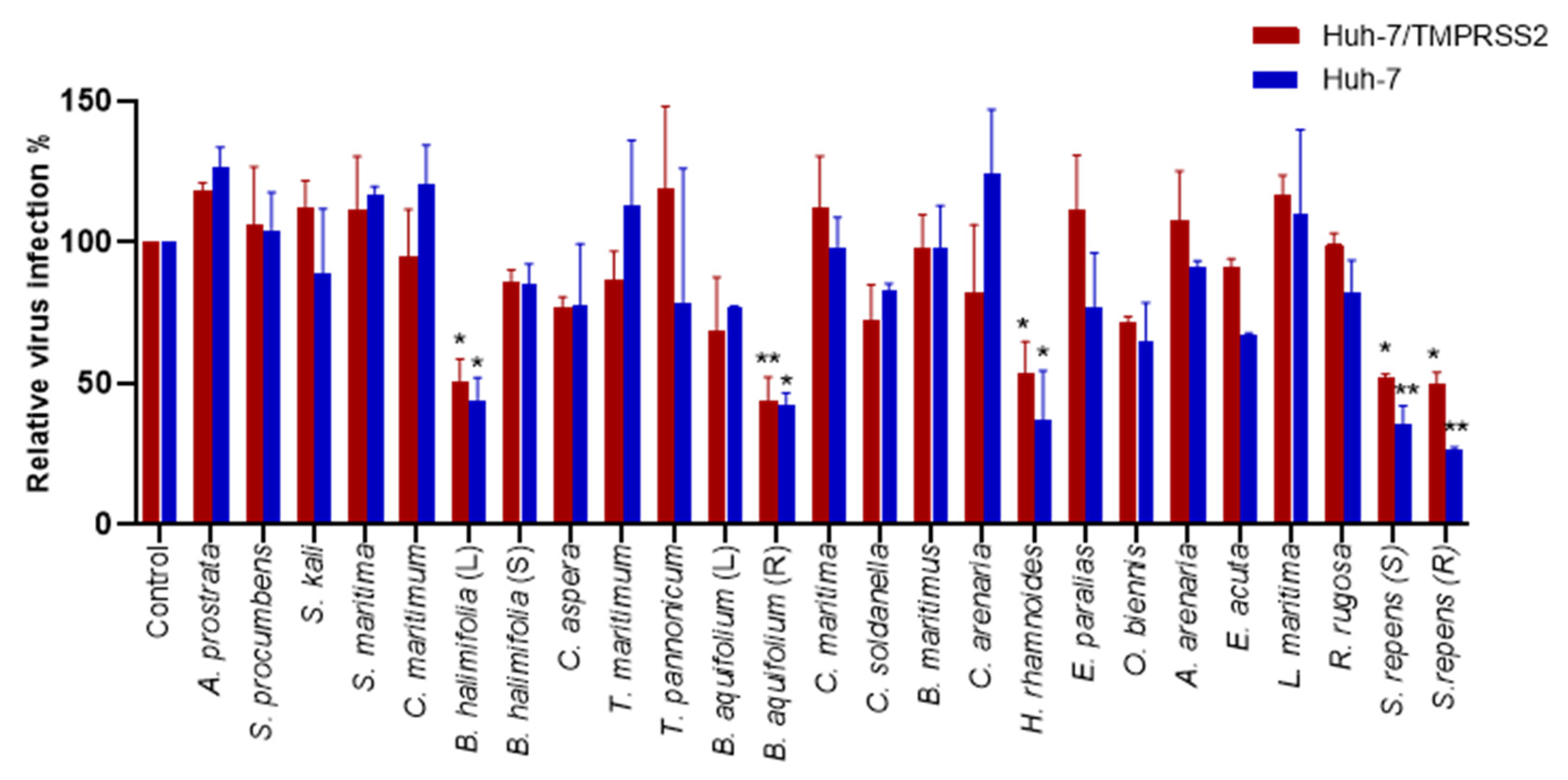
2.2.2. Antiviral Screening of the Plant Crude Methanolic Extracts on HCoV-229E
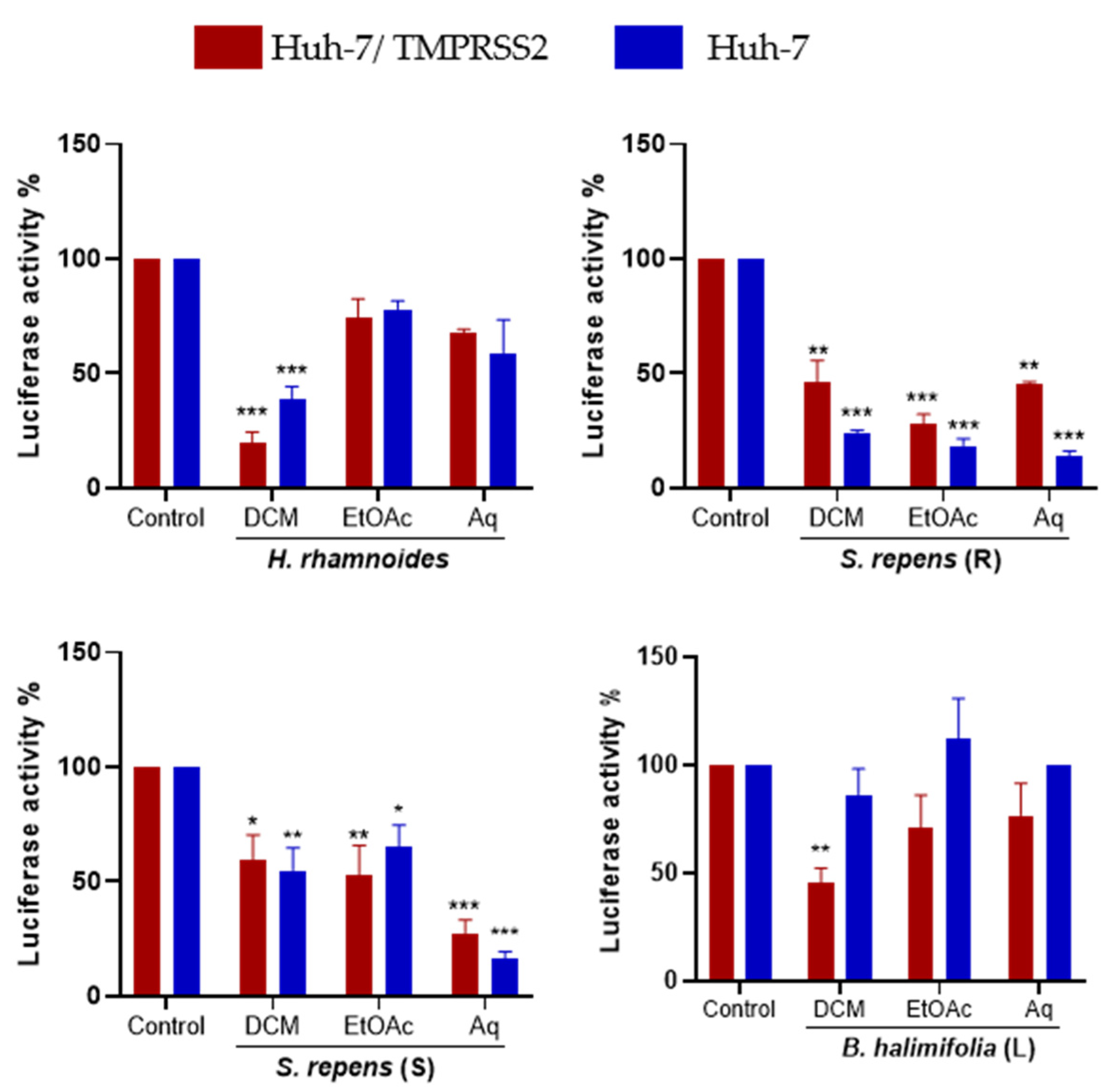
2.3. Dose-Response Antiviral Activity of Plant Extracts
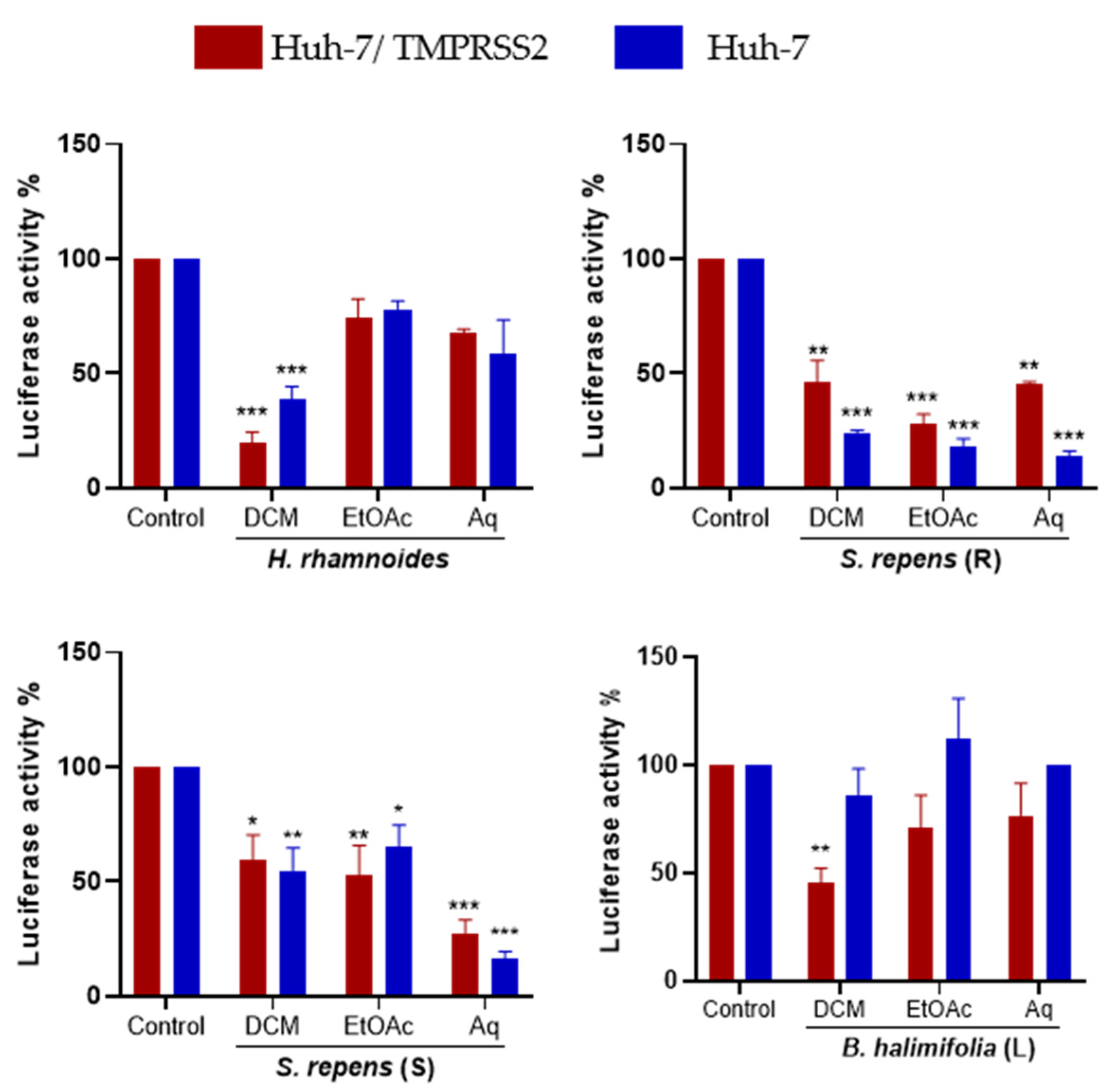
2.4. Bioguided Fractionation Assay to Determine the Active Sub-Extracts
2.5. Characterization of HR DCM SE by UHPLC UV-MS
2.6. Cytotoxicity of Ten Fractions Prepared from HR DCM SE
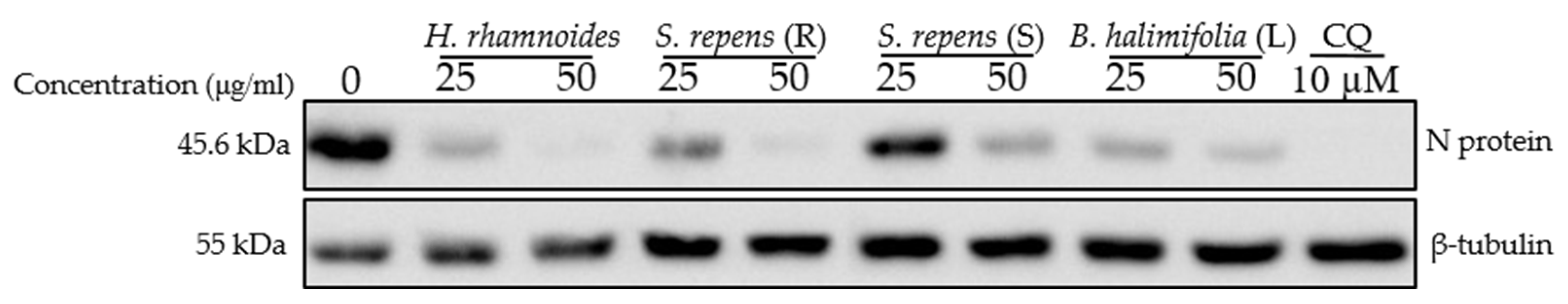
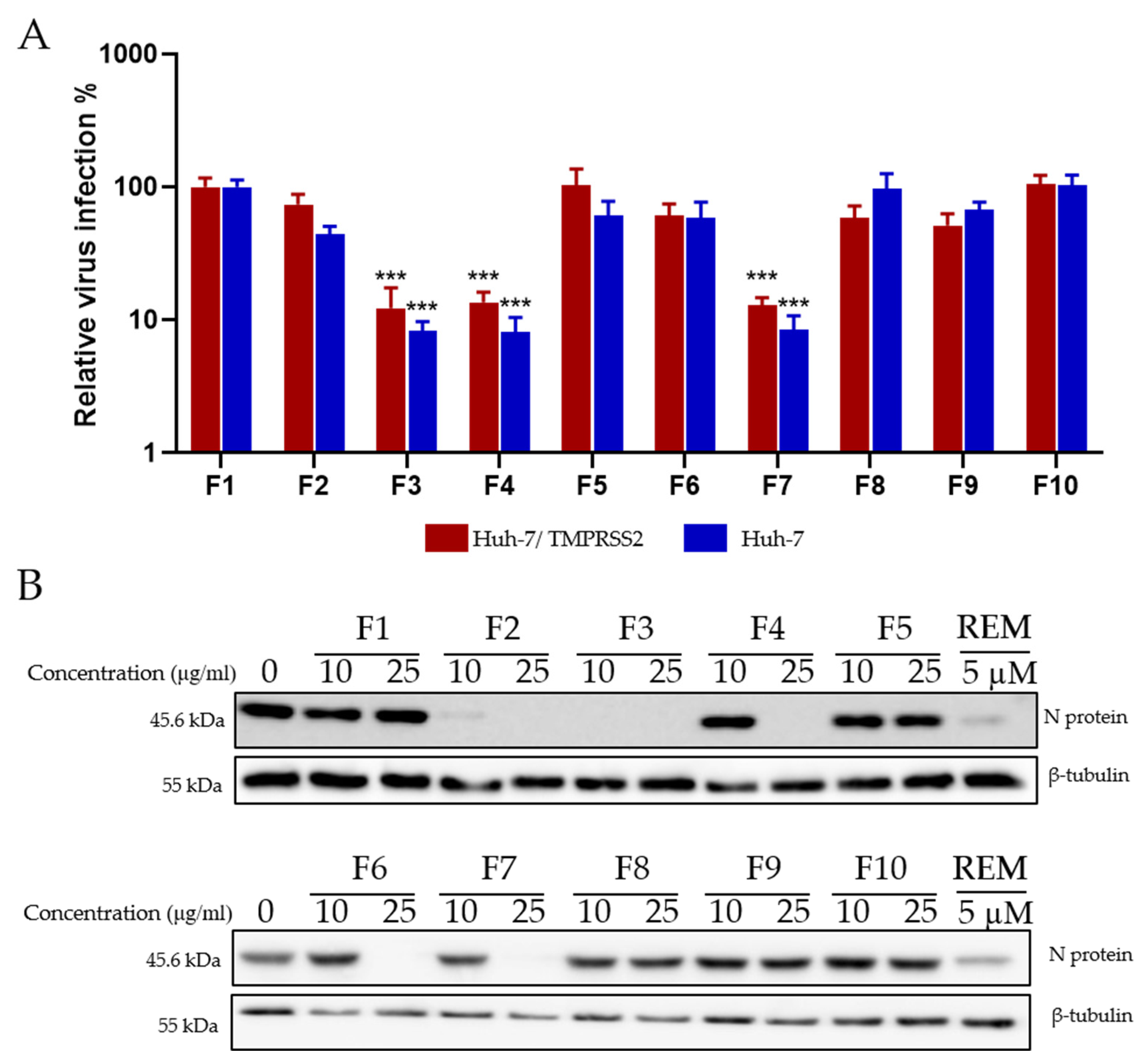
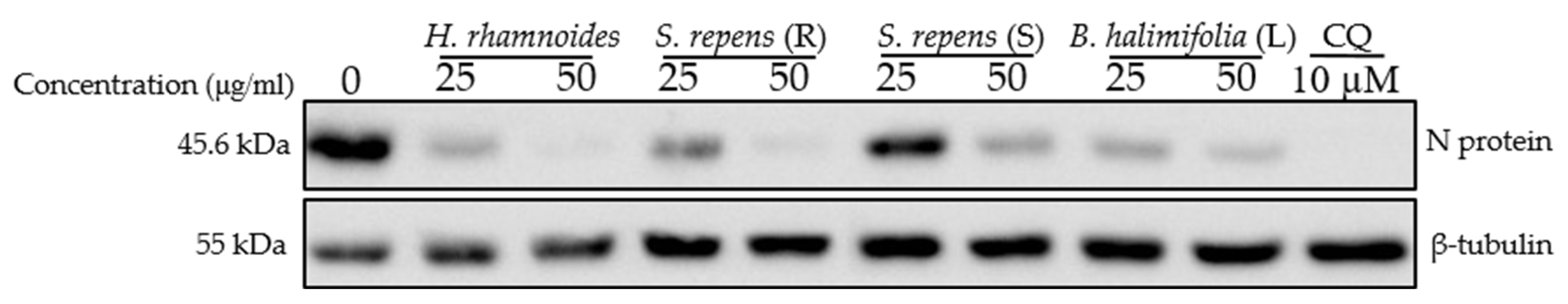
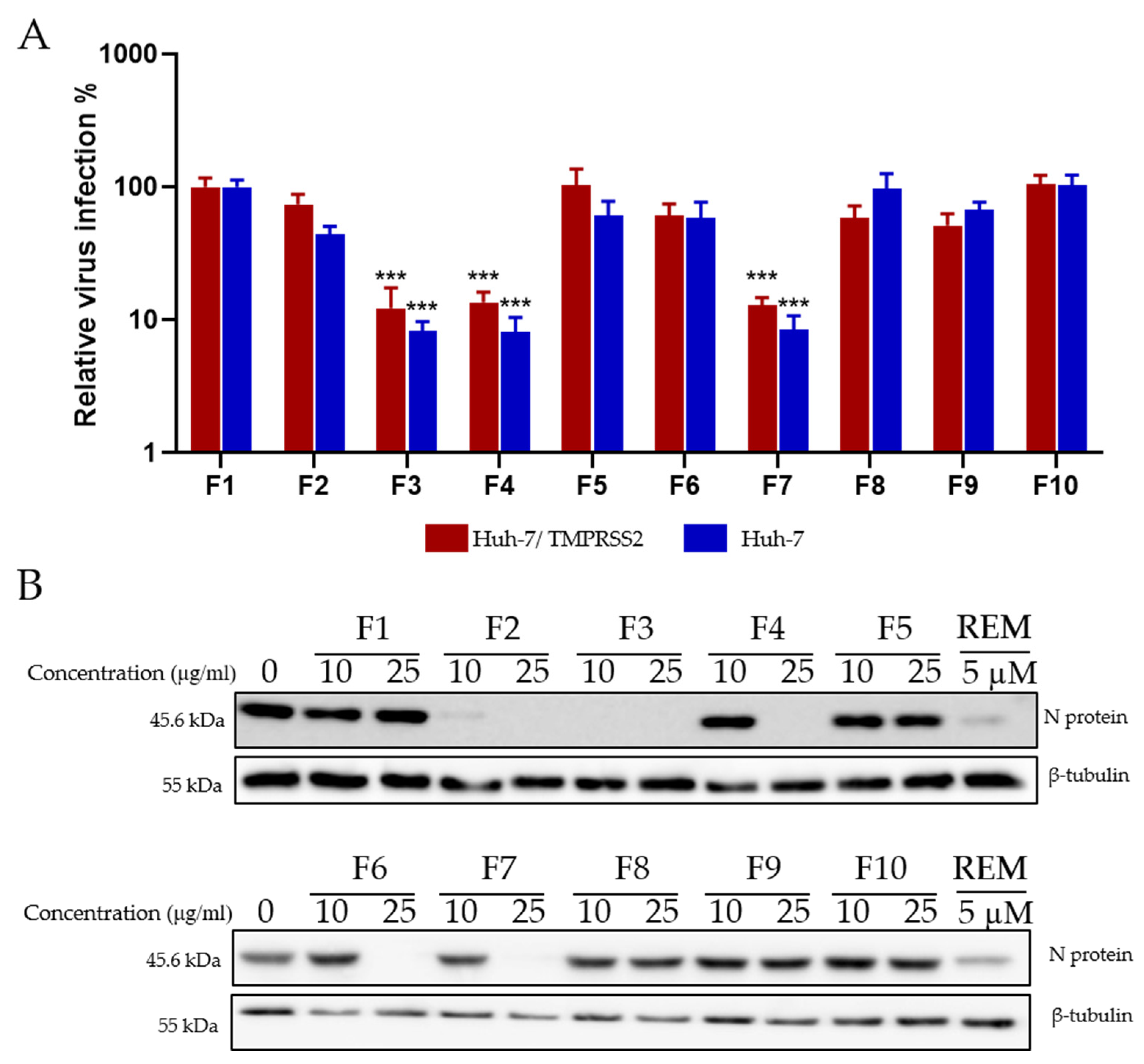
2.7. Antiviral Screening of Ten Fractions Obtained from HR DCM SE against HCoV-229E and SARS-CoV-2
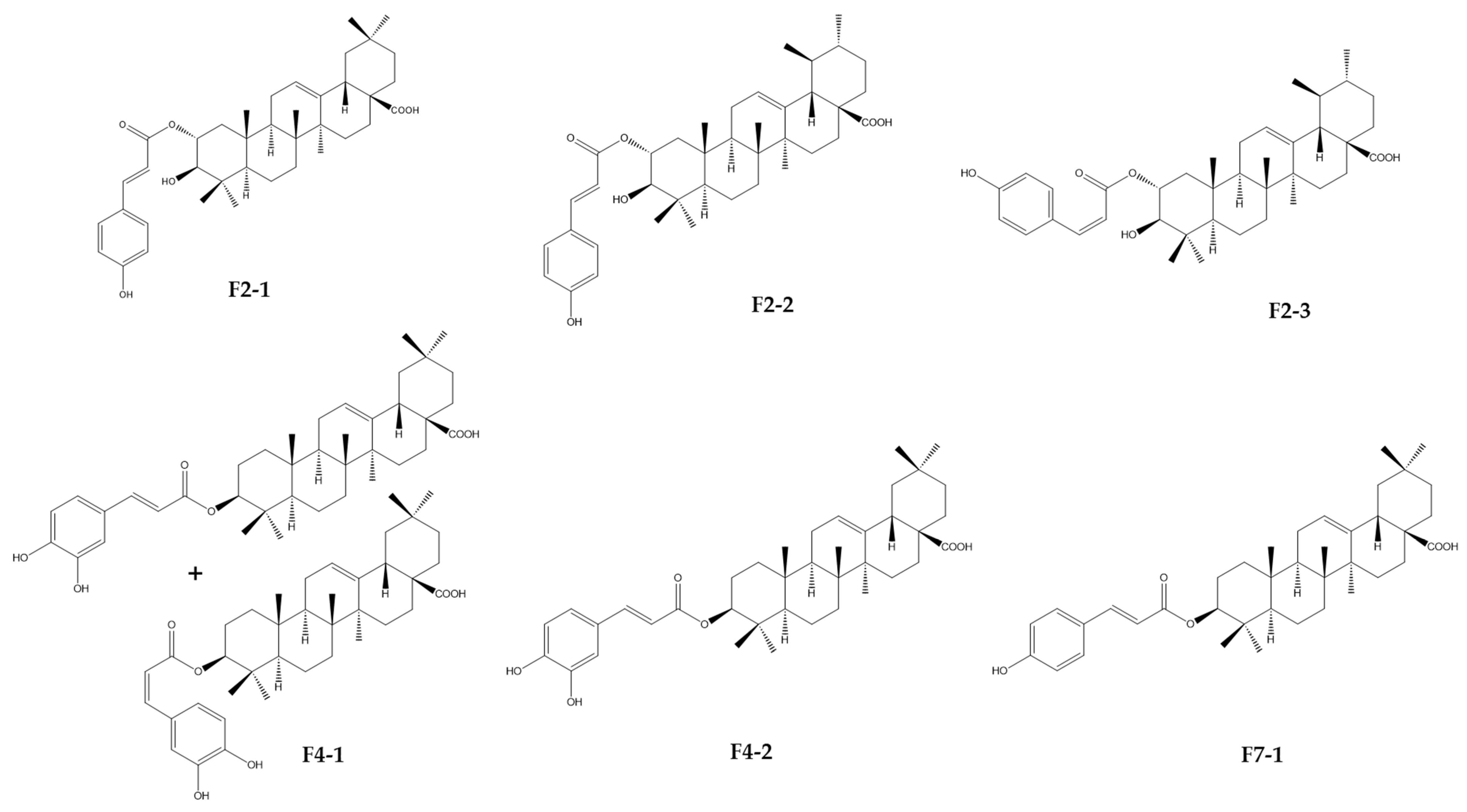
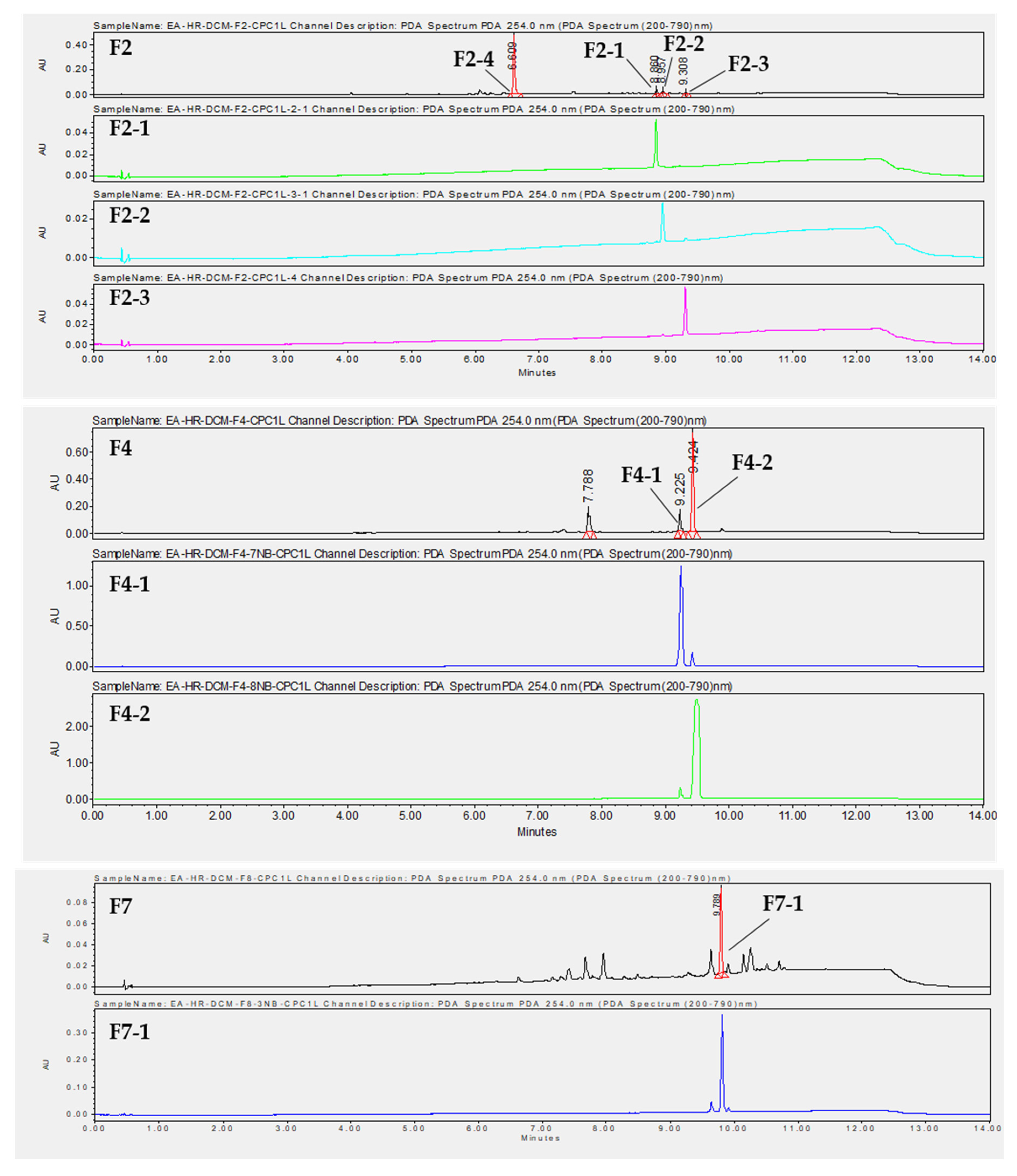
2.8. Isolation of Compounds by Preparative HPLC and Identification by HRMS and NMR
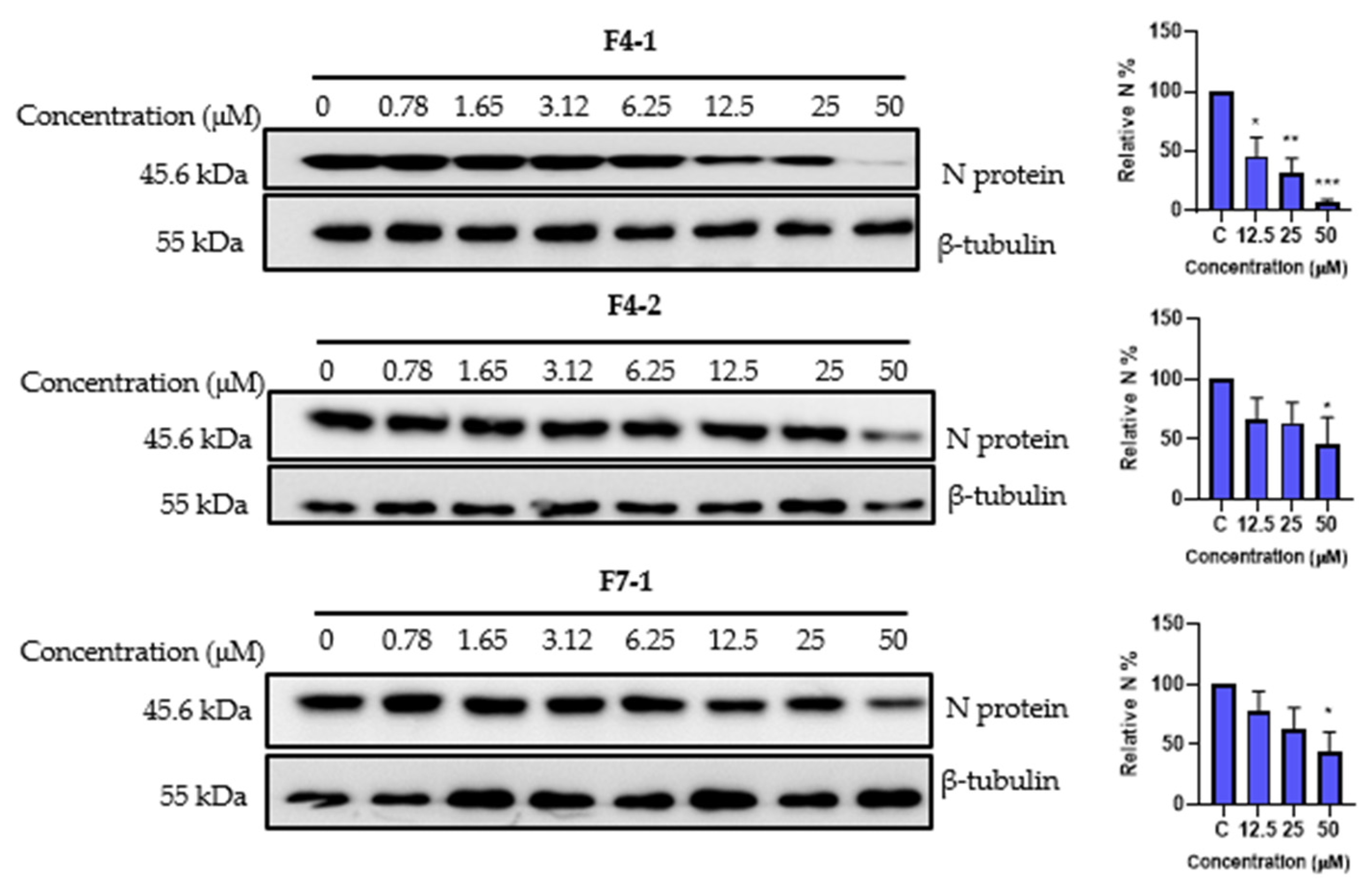
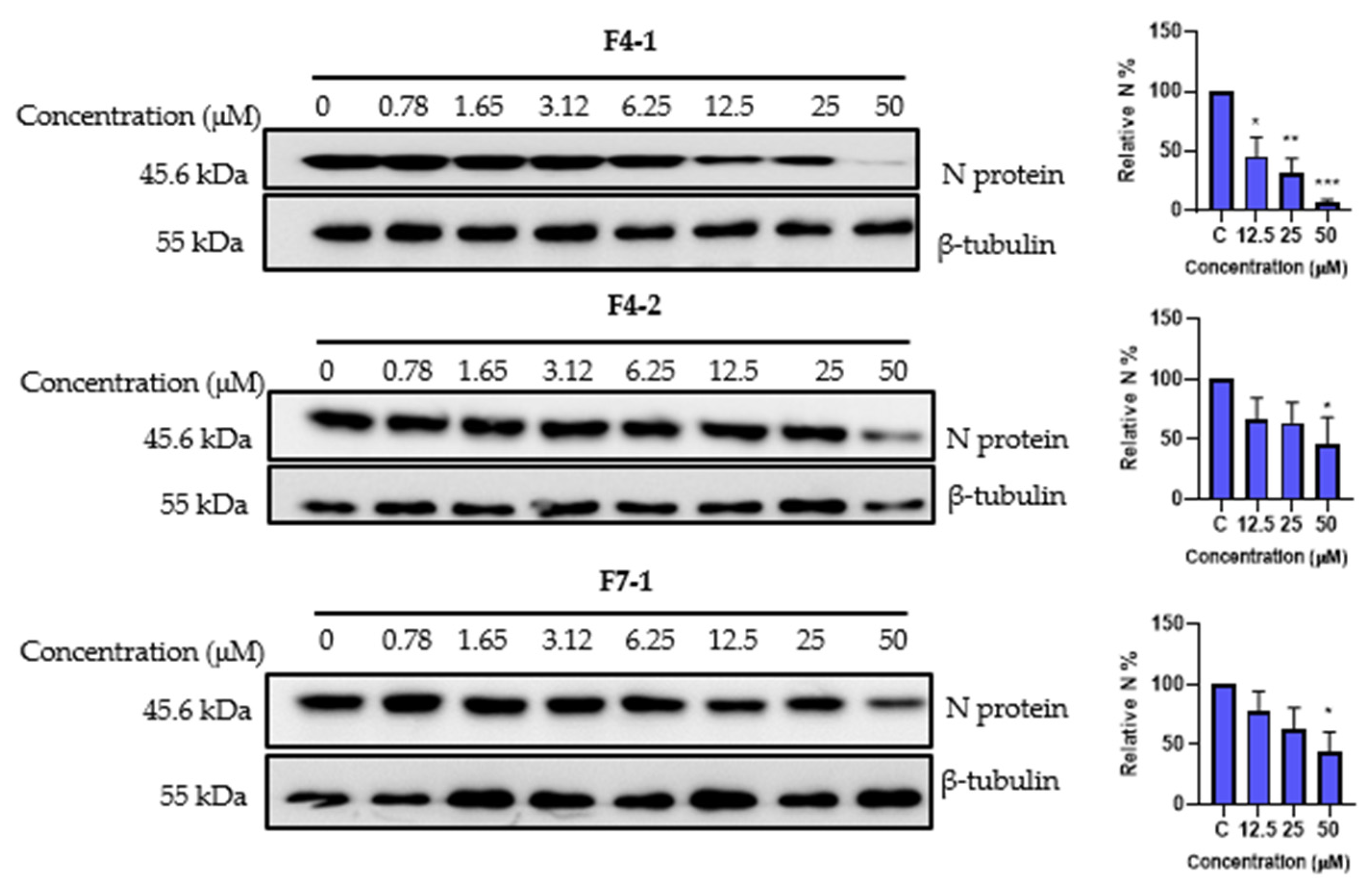
2.9. Cytotoxicity and Antiviral Activity of the Purified Compounds on HCoV-229E and SARS-CoV-2
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Solid/Liquid Extraction
4.3. Liquid/Liquid Extraction
4.4. Fractionation of the DCM Sub-Extract of Hippophae Rhamnoides
4.5. UHPLC-UV-MS Analysis
4.6. Preparative HPLC
4.7. NMR and HRMS
4.8. Virus and Cell Lines
4.9. Cell Viability Assay
4.10. Virus Infection Assay
4.10.1. HCoV-229E
4.10.2. SARS-CoV-2
4.10.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kesheh, M.M.; Hosseini, P.; Soltani, S.; Zandi, M. An overview on the seven pathogenic human coronaviruses. Rev. Med. Virol. 2022, 32, e2282. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.X.; Liang, J.Q.; Fung, T.S. Human Coronavirus-229E, -OC43, -NL63, and -HKU1 (Coronaviridae). Encycl. Virol. 2021, 2, 428–440. [Google Scholar] [CrossRef]
- Zhu, Z.; Lian, X.; Su, X.; Wu, W.; Marraro, G.A.; Zeng, Y. From SARS and MERS to COVID-19: A brief summary and comparison of severe acute respiratory infections caused by three highly pathogenic human coronaviruses. Respir. Res. 2020, 21, 224. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Hunt, B.J.; Stegermann, M.; Rochwerg, B.; Lamontagne, F.; Siemieniuk, R.A.; Agoritsas, T.; Askie, L.; Lytvyn, L.; Leo, Y.-S.; et al. A living WHO guideline on drugs for COVID-19. BMJ 2020, 370, m3379. [Google Scholar] [CrossRef] [PubMed]
- EMA (European Medicines Agency). Lagevrio: Withdrawn Application. Available online: https://www.ema.europa.eu/en/medicines/human/withdrawn-applications/lagevrio (accessed on 15 October 2023).
- Duroseau, B.; Kipshidze, N.; Limaye, R.J. The impact of delayed access to COVID-19 vaccines in low- and lower-middle-income countries. Front. Public Health 2023, 10, 1087138. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Li, Q.Y.; Liu, J.; Efferth, T. Traditional Chinese herbal medicine at the forefront battle against COVID-19: Clinical experience and scientific basis. Phytomedicine 2021, 80, 153337. [Google Scholar] [CrossRef] [PubMed]
- Villena-Tejada, M.; Vera-Ferchau, I.; Cardona-Rivero, A.; Zamalloa-Cornejo, R.; Quispe-Florez, M.; Frisancho-Triveño, Z.; Abarca-Meléndez, R.C.; Alvarez-Sucari, S.G.; Mejia, C.R.; Yañez, J.A. Use of medicinal plants for COVID-19 prevention and respiratory symptom treatment during the pandemic in Cusco, Peru: A cross-sectional survey. PLoS ONE 2021, 16, e0257165. [Google Scholar] [CrossRef]
- Khadka, D.; Dhamala, M.K.; Li, F.; Aryal, P.C.; Magar, P.R.; Bhatta, S.; Thakur, M.S.; Basnet, A.; Cui, D.; Shi, S. The use of medicinal plants to prevent COVID-19 in Nepal. J. Ethnobiol. Ethnomed. 2021, 17, 26. [Google Scholar] [CrossRef]
- Duan, Q.; Zhu, Z.; Wang, B.; Chen, M. Recent Progress on the Salt Tolerance Mechanisms and Application of Tamarisk. Int. J. Mol. Sci. 2022, 23, 3325. [Google Scholar] [CrossRef]
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. eHALOPH a Database of Salt-Tolerant Plants: Helping put Halophytes to Work. Plant Cell Physiol. 2016, 57, e10. [Google Scholar] [CrossRef]
- Cheeseman, J.M. The evolution of halophytes, glycophytes and crops, and its implications for food security under saline conditions. New Phytol. 2015, 206, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, G.; Rivière, C. Amaranthaceae halophytes from the French Flanders coast of the North Sea: A review of their phytochemistry and biological activities. Phytochem. Rev. 2020, 19, 1263–1302. [Google Scholar] [CrossRef]
- Kuo, Y.-C.; Lin, L.-C.; Tsai, W.-J.; Chou, C.-J.; Kung, S.-H.; Ho, Y.-H. Samarangenin B from Limonium sinense Suppresses Herpes Simplex Virus Type 1 Replication in Vero Cells by Regulation of Viral Macromolecular Synthesis. Antimicrob. Agents Chemother. 2002, 46, 2854–2864. [Google Scholar] [CrossRef] [PubMed]
- Medini, F.; Legault, J.; Pichette, A.; Abdelly, C.; Ksouri, R. Antiviral efficacy of Limonium densiflorum against HSV-1 and influenza viruses. S. Afr. J. Bot. 2014, 92, 65–72. Available online: https://cyberleninka.org/article/n/390317 (accessed on 7 September 2021). [CrossRef]
- Chiang, L.; Chiang, W.; Chang, M.; Ng, L.; Lin, C. Antiviral activity of Plantago major extracts and related compounds in vitro. Antivir. Res. 2002, 55, 53–62. [Google Scholar] [CrossRef]
- Sahuc, M.-E.; Sahli, R.; Rivière, C.; Pène, V.; Lavie, M.; Vandeputte, A.; Brodin, P.; Rosenberg, A.R.; Dubuisson, J.; Ksouri, R.; et al. Dehydrojuncusol, a Natural Phenanthrene Compound Extracted from Juncus maritimus, Is a New Inhibitor of Hepatitis C Virus RNA Replication. J. Virol. 2019, 93, e02009-18. [Google Scholar] [CrossRef] [PubMed]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of Coronavirus Cell Entry Mediated by the Viral Spike Protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Binder, J.; Yurgelonis, I.; Rai, D.K.; Lazarro, S.; Costales, C.; Kobylarz, K.; McMonagle, P.; Steppan, C.M.; Aschenbrenner, L.; et al. Generation of a VeroE6 Pgp gene knock out cell line and its use in SARS-CoV-2 antiviral study. Antivir. Res. 2022, 208, 105429. [Google Scholar] [CrossRef]
- Yang, Z.-G.; Li, H.-R.; Wang, L.-Y.; Li, Y.-H.; Lu, S.-G.; Wen, X.-F.; Wang, J.; Daikonya, A.; Kitanaka, S. Triterpenoids from Hippophae rhamnoides L. and Their Nitric Oxide Production-Inhibitory and DPPH Radical-Scavenging Activities. Chem. Pharm. Bull. 2007, 55, 15–18. [Google Scholar] [CrossRef]
- Siddiqui, S.; Siddiqui, B.S.; Hafeez, F.; Begum, S. Isolation and Structure of Neriucoumaric and Isoneriucoumaric Acids from the Leaves of Nerium oleander. Planta Medica 1987, 53, 424–427. [Google Scholar] [CrossRef]
- Fuchino, H.; Satoh, T.; Tanaka, N. Chemical Evaluation of Betula Species in Japan. I.Constituents of Betula ermanii. Chem. Pharm. Bull. 1995, 43, 1937–1942. [Google Scholar] [CrossRef]
- Takahashi, H.; Iuchi, M.; Fujita, Y.; Minami, H.; Fukuyama, Y. Coumaroyl triterpenes from Casuarina equisetifolia. Phytochemistry 1999, 51, 543–550. [Google Scholar] [CrossRef]
- Reshamwala, D.; Shroff, S.; Liimatainen, J.; Tienaho, J.; Laajala, M.; Kilpeläinen, P.; Viherä-Aarnio, A.; Karonen, M.; Jyske, T.; Marjomäki, V. Willow (Salix spp.) bark hot extracts inhibit both enveloped and nonenveloped viruses: Study on its anti-coronavirus and anti-enterovirus activities. Front. Microbiol. 2023, 14, 1249794. [Google Scholar] [CrossRef]
- Zhan, Y.; Ta, W.; Tang, W.; Hua, R.; Wang, J.; Wang, C.; Lu, W. Potential antiviral activity of isorhamnetin against SARS-CoV-2 spike pseudotyped virus in vitro. Drug Dev. Res. 2021, 82, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Rousi, A. The genus Hippophaë L. A taxonomic study. Ann. Bot. Fenn. 1971, 8, 177–227. [Google Scholar]
- Zakynthinos, G.; Varzakas, T. Hippophae rhamnoides: Safety and nutrition. Curr. Res. Nutr. Food Sci. J. 2015, 3, 89–97. [Google Scholar] [CrossRef]
- Żuchowski, J. Phytochemistry and pharmacology of sea buckthorn (Elaeagnus rhamnoides; syn. Hippophae rhamnoides): Progress from 2010 to 2021. Phytochem. Rev. 2023, 22, 3–33. [Google Scholar] [CrossRef]
- Kim, S.-J.; Hwang, E.; Yi, S.S.; Song, K.D.; Lee, H.-K.; Heo, T.-H.; Park, S.-K.; Jung, Y.J.; Jun, H.S. Sea Buckthorn Leaf Extract Inhibits Glioma Cell Growth by Reducing Reactive Oxygen Species and Promoting Apoptosis. Appl. Biochem. Biotechnol. 2017, 182, 1663–1674. [Google Scholar] [CrossRef]
- Upadhyay, N.K.; Kumar, M.Y.; Gupta, A. Antioxidant, cytoprotective and antibacterial effects of Sea buckthorn (Hippophae rhamnoides L.) leaves. Food Chem. Toxicol. 2010, 48, 3443–3448. [Google Scholar] [CrossRef]
- Basu, M.; Prasad, R.; Jayamurthy, P.; Pal, K.; Arumughan, C.; Sawhney, R. Anti-atherogenic effects of seabuckthorn (Hippophaea rhamnoides) seed oil. Phytomed. Int. J. Phytother. Phytopharm. 2007, 14, 770–777. [Google Scholar] [CrossRef]
- Solcan, C.; Gogu, M.; Floristean, V.; Oprisan, B.; Solcan, G. The hepatoprotective effect of sea buckthorn (Hippophae rhamnoides) berries on induced aflatoxin B1 poisoning in chickens. Poult. Sci. 2013, 92, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Goel, H.; Kumar, I.P.; Samanta, N.; Rana, S. Induction of DNA-protein cross-links by Hippophae rhamnoides: Implications in radioprotection and cytotoxicity. Mol. Cell. Biochem. 2003, 245, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, N.K.; Kumar, R.; Siddiqui, M.S.; Gupta, A. Mechanism of Wound-Healing Activity of Hippophae rhamnoides L. Leaf Extract in Experimental Burns. Evid.-Based Complement. Altern. Med. 2011, 2011, 659705. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, B.; Kontek, R.; Żuchowski, J.; Stochmal, A. Novel bioactive properties of low-polarity fractions from sea-buckthorn extracts (Elaeagnus rhamnoides (L.) A. Nelson)—(in vitro). Biomed. Pharmacother. 2021, 135, 111141. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Si, L.; Wang, Y.; Wu, Y.; Yu, F.; Jiao, P.; Shi, Y.; Wang, H.; Xiao, S.; Fu, G.; et al. Discovery of Pentacyclic Triterpenoids as Potential Entry Inhibitors of Influenza Viruses. J. Med. Chem. 2014, 57, 10058–10071. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Wang, Q.; Zhang, Z.; Peng, Y.; Qiu, Y.; Shi, Y.; Zheng, Y.; Xiao, S.; Wang, H.; Huang, X.; et al. Development of Oleanane-Type Triterpenes as a New Class of HCV Entry Inhibitors. J. Med. Chem. 2013, 56, 4300–4319. [Google Scholar] [CrossRef] [PubMed]
- Si, L.; Meng, K.; Tian, Z.; Sun, J.; Li, H.; Zhang, Z.; Soloveva, V.; Li, H.; Fu, G.; Xia, Q.; et al. Triterpenoids manipulate a broad range of virus-host fusion via wrapping the HR2 domain prevalent in viral envelopes. Sci. Adv. 2018, 4, eaau8408. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.B.; Park, S.-J.; Kim, Y.M.; Lee, J.-Y.; Seo, W.D.; Chang, J.S.; Park, K.H.; Rho, M.-C.; Lee, W.S. SARS-CoV 3CLpro inhibitory effects of quinone-methide triterpenes from Tripterygium regelii. Bioorganic Med. Chem. Lett. 2010, 20, 1873–1876. [Google Scholar] [CrossRef]
- Darshani, P.; Sarma, S.S.; Srivastava, A.K.; Baishya, R.; Kumar, D. Anti-viral triterpenes: A review. Phytochem. Rev. 2022, 21, 1761–1842. [Google Scholar] [CrossRef]
- Tkacz, K.; Wojdyło, A.; Turkiewicz, I.P.; Nowicka, P. Triterpenoids, phenolic compounds, macro- and microelements in anatomical parts of sea buckthorn (Hippophaë rhamnoides L.) berries, branches and leaves. J. Food Compos. Anal. 2021, 103, 104107. [Google Scholar] [CrossRef]
- Braca, A.; Morelli, I.; Mendez, J.; Battinelli, L.; Braghiroli, L.; Mazzanti, G. Antimicrobial Triterpenoids from Licania heteromorpha. Planta Medica 2000, 66, 768–769. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Alam, M.; Khatoon, F.; Fatima, U.; Elasbali, A.M.; Adnan, M.; Islam, A.; Hassan, I.; Snoussi, M.; De Feo, V. Natural products can be used in therapeutic management of COVID-19: Probable mechanistic insights. Biomed. Pharmacother. 2022, 147, 112658. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Choudhir, G.; Shukla, S.K.; Sharma, M.; Tyagi, P.; Bhushan, A.; Rathore, M. Identification of phytochemical inhibitors against main protease of COVID-19 using molecular modeling approaches. J. Biomol. Struct. Dyn. 2021, 39, 3760–3770. [Google Scholar] [CrossRef]
- Mitra, D.; Verma, D.; Mahakur, B.; Kamboj, A.; Srivastava, R.; Gupta, S.; Pandey, A.; Arora, B.; Pant, K.; Panneerselvam, P.; et al. Molecular docking and simulation studies of natural compounds of Vitex negundo L. against papain-like protease (PLpro) of SARS-CoV-2 (coronavirus) to conquer the pandemic situation in the world. J. Biomol. Struct. Dyn. 2022, 40, 5665–5686. [Google Scholar] [CrossRef] [PubMed]
- Anbazhagan, S.; Muthu, S.; Singh, R.K.; Pallab, C. In silico molecular docking analysis targeting SARS-CoV-2 spike protein and selected herbal constituents. J. Pure Appl. Microbiol. 2020, 14, 989–998. [Google Scholar]
- Duhamel, F.; Farvacques, C.; Blondel, C.; Delplanque, S.; Catteau, E.; Gelez, W.; Francois, R.; Prey, T.; Cholet, J.; Buchet, J.; et al. Guide des Végétations Littorales du Nord-Ouest de la France. CBN de Bailleul. Available online: https://www.cbnbl.org/guide-vegetations-littorales-du-nord-ouest-france (accessed on 15 October 2023).
- Hesp, P. Foredunes and blowouts: Initiation, geomorphology and dynamics. Geomorphology 2002, 48, 245–268. [Google Scholar] [CrossRef]
- Belouzard, S.; Machelart, A.; Sencio, V.; Vausselin, T.; Hoffmann, E.; Deboosere, N.; Rouillé, Y.; Desmarets, L.; Séron, K.; Danneels, A.; et al. Clofoctol inhibits SARS-CoV-2 replication and reduces lung pathology in mice. PLoS Pathog. 2022, 18, e1010498. [Google Scholar] [CrossRef]
- Meunier, T.; Desmarets, L.; Bordage, S.; Bamba, M.; Hervouet, K.; Rouillé, Y.; François, N.; Decossas, M.; Sencio, V.; Trottein, F.; et al. A Photoactivable Natural Product with Broad Antiviral Activity against Enveloped Viruses, Including Highly Pathogenic Coronaviruses. Antimicrob. Agents Chemother. 2022, 66, e0158121. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Botanical Family | Plant Species | Parts | Place of Collection | Voucher Code |
|---|---|---|---|---|
| Amaranthaceae | Atriplex prostrata Boucher ex DC. | Whole plant | Dannes (at the base of an incipient dune) | LIP007584 |
| Salicornia procumbens Sm. | Whole plant | Etaples (in the schorre) | LIP007585 | |
| Salsola kali L. | Whole plant | Dannes (at the base of an incipient dune) | LIP007586 | |
| Suaeda maritima (L.) Dumort. | Whole plant | Etaples (in the schorre) | LIP007587 | |
| Apiaceae | Crithmum maritimum L. | Aerial parts | Le Portel (on a coastal cliff) | LIP007588 |
| Baccharis halimifolia L. | Leaves (L), stems (S) | Gravelines (planted in a roadside hedge) | LIP007589 | |
| Asteraceae | Centaurea aspera L. | Whole plant | Zuydcoote (on a relict foredune) | LIP007590 |
| Tripleurospermum maritimum (L.) W. D. J. Koch | Whole plant | Le Portel (on a coastal cliff) | LIP007591 | |
| Tripolium pannonicum (Jacq.) Dobroc. | Whole plant | Dannes (in the schorre) | LIP007592 | |
| Berberidaceae | Berberis aquifolium Pursh | Leaves (L), roots (R) | Zuydcoote (on a relict foredune) | LIP007593 |
| Brassicaceae | Cakile maritima Scop. subsp. integrifolia (Hornem.) Greuter and Burdet | Whole plant | Dannes (at the base of an incipient dune) | LIP007594 |
| Convolvulaceae | Convolvulus soldanella L. | Whole plant | Dannes (on an incipient dune) | LIP007595 |
| Cyperaceae | Bolboschoenus maritimus (L.) Palla | Whole plant | Dannes (in the schorre) | LIP007596 |
| Carex arenaria L. | Whole plant | Dannes (in the schorre) | LIP007597 | |
| Elaeagnaceae | Hippophae rhamnoides L. | Whole plant | Dannes (on an established foredune) | LIP007598 |
| Euphorbiaceae | Euphorbia paralias L. | Whole plant | Dannes (on an incipient foredune) | LIP007599 |
| Onagraceae | Oenothera biennis L. | Whole plant | Zuydcoote (on a relict foredune) | LIP007600 |
| Poaceae | Ammophila arenaria subsp. arenaria (L.) Link | Whole plant | Dannes (on an established foredune) | LIP007601 |
| Elytrigia acuta (DC.) Tzvelev | Whole plant | Zuydcoote (on an incipient foredune) | LIP007602 | |
| Primulaceae | Lysimachia maritima (L.) Galasso, Banfi, and Soldano | Whole plant | Dannes (in the schorre) | LIP007603 |
| Rosaceae | Rosa rugosa Thumb. | Aerial parts | Dannes (on an established foredune) | LIP007604 |
| Salicaceae | Salix repens subsp. dunensis Rouy | Stems (S), roots (R) | Zuydcoote (on a relict foredune) | LIP007605 |
| Crude Extract | CC50 (μg/mL) Vero-81 | CC50 (μg/mL) Huh-7 | Huh-7 | Huh-7/TMPRSS2 | ||
|---|---|---|---|---|---|---|
| IC50 (μg/mL) | SI | IC50 (μg/mL) | SI | |||
| H. rhamnoides | 499 | 621 | 19.7 | 31.5 | 27.2 | 22 |
| S. repens (R) | 441 | 149 | 29.1 | 5.1 | 15.5 | 9 |
| S. repens (S) | 438 | 285 | 15.1 | 28.9 | 14.7 | 19 |
| B halimifolia (L) | 820 | >1000 | 65.3 | >15 | 11.2 | >89 |
| Sub-Extract | Vero-81 CC50 (μg/mL) | Huh-7 CC50 (μg/mL) | Huh-7 | Huh-7/TMPRSS2 | ||
|---|---|---|---|---|---|---|
| IC50 (μg/mL) | SI | IC50 (μg/mL) | SI | |||
| H. rhamnoides DCM | 264 | 410 | 18.7 | 21 | 36. | 11 |
| S. repens (R) EtOAc | 344 | 500 | 15.8 | 31 | 28.87 | 17 |
| S. repens (S) Aq | 262 | 550 | 7.6 | 71 | 30.5 | 18 |
| B. halimifolia (L) DCM | 347 | 368 | 31.1 | 11 | 38.3 | 9 |
| Sub-Extract | Huh-7 CC50 (µM) | Huh-7 | Huh-7/TMPRSS2 | ||
|---|---|---|---|---|---|
| IC50 (μM) | SI | IC50 (μM) | SI | ||
| F2-1 (2-O-trans-p-coumaroyl-maslinic acid) | 39.9 | 8.6 | 4 | 9.1 | 4 |
| F2-2 (3β-hydroxy-2α-trans-p-coumaryloxy-urs-12-en-28-oic acid) | 44.4 | 12.0 | 3 | 11.4 | 3 |
| F2-3 (3β-hydroxy-2α-cis-p-coumaryloxy-urs-12-en-28-oic acid) | 52.1 | 14.5 | 3 | 14.1 | 3 |
| F4-1 Mixture 3-O-trans-caffeoyl oleanolic acid / 3-O-cis-caffeoyl oleanolic acid (70/30) | 21.0 | 7.6 | 2 | 12.0 | 1 |
| F4-2 (3-O-trans-caffeoyl-oleanolic acid) | 81.4 | 11.6 | 6 | 11.5 | 7 |
| F7-1 (3-O-trans-p-coumaroyl-oleanolic acid) | 39.3 | 11.4 | 3 | 10.7 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Ibrahim, M.; Akissi, Z.L.E.; Desmarets, L.; Lefèvre, G.; Samaillie, J.; Raczkiewicz, I.; Sahpaz, S.; Dubuisson, J.; Belouzard, S.; Rivière, C.; et al. Discovery of Anti-Coronavirus Cinnamoyl Triterpenoids Isolated from Hippophae rhamnoides during a Screening of Halophytes from the North Sea and Channel Coasts in Northern France. Int. J. Mol. Sci. 2023, 24, 16617. https://doi.org/10.3390/ijms242316617
Al Ibrahim M, Akissi ZLE, Desmarets L, Lefèvre G, Samaillie J, Raczkiewicz I, Sahpaz S, Dubuisson J, Belouzard S, Rivière C, et al. Discovery of Anti-Coronavirus Cinnamoyl Triterpenoids Isolated from Hippophae rhamnoides during a Screening of Halophytes from the North Sea and Channel Coasts in Northern France. International Journal of Molecular Sciences. 2023; 24(23):16617. https://doi.org/10.3390/ijms242316617
Chicago/Turabian StyleAl Ibrahim, Malak, Zachee Louis Evariste Akissi, Lowiese Desmarets, Gabriel Lefèvre, Jennifer Samaillie, Imelda Raczkiewicz, Sevser Sahpaz, Jean Dubuisson, Sandrine Belouzard, Céline Rivière, and et al. 2023. "Discovery of Anti-Coronavirus Cinnamoyl Triterpenoids Isolated from Hippophae rhamnoides during a Screening of Halophytes from the North Sea and Channel Coasts in Northern France" International Journal of Molecular Sciences 24, no. 23: 16617. https://doi.org/10.3390/ijms242316617
APA StyleAl Ibrahim, M., Akissi, Z. L. E., Desmarets, L., Lefèvre, G., Samaillie, J., Raczkiewicz, I., Sahpaz, S., Dubuisson, J., Belouzard, S., Rivière, C., & Séron, K. (2023). Discovery of Anti-Coronavirus Cinnamoyl Triterpenoids Isolated from Hippophae rhamnoides during a Screening of Halophytes from the North Sea and Channel Coasts in Northern France. International Journal of Molecular Sciences, 24(23), 16617. https://doi.org/10.3390/ijms242316617