
Expression Patterns and Functional Analysis of Three SmTAT Genes Encoding Tyrosine Aminotransferases in Salvia miltiorrhiza

Abstract
:1. Introduction
2. Results
2.1. Bioinformatics Analysis of SmTATs in S. miltiorrhiza
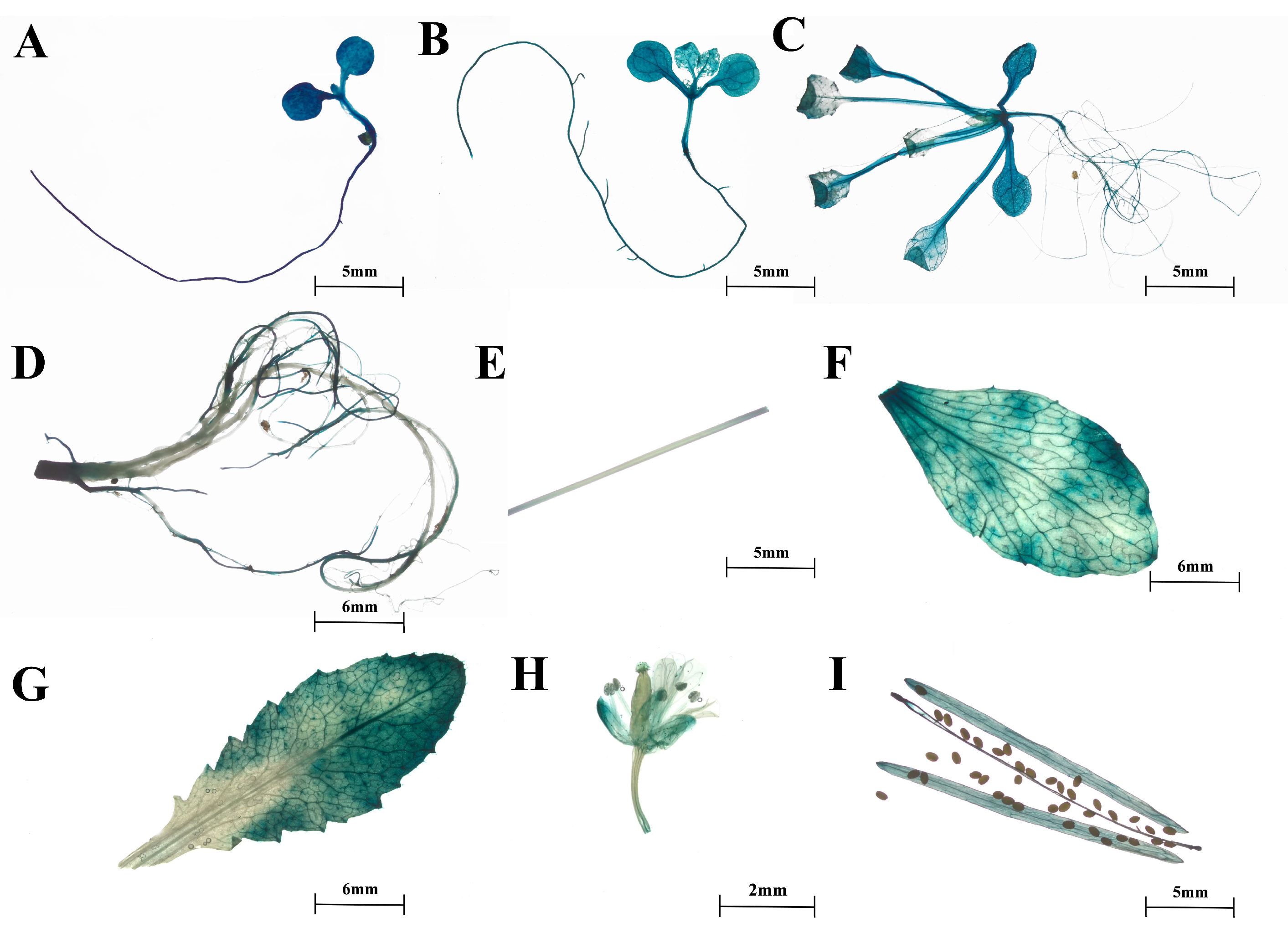
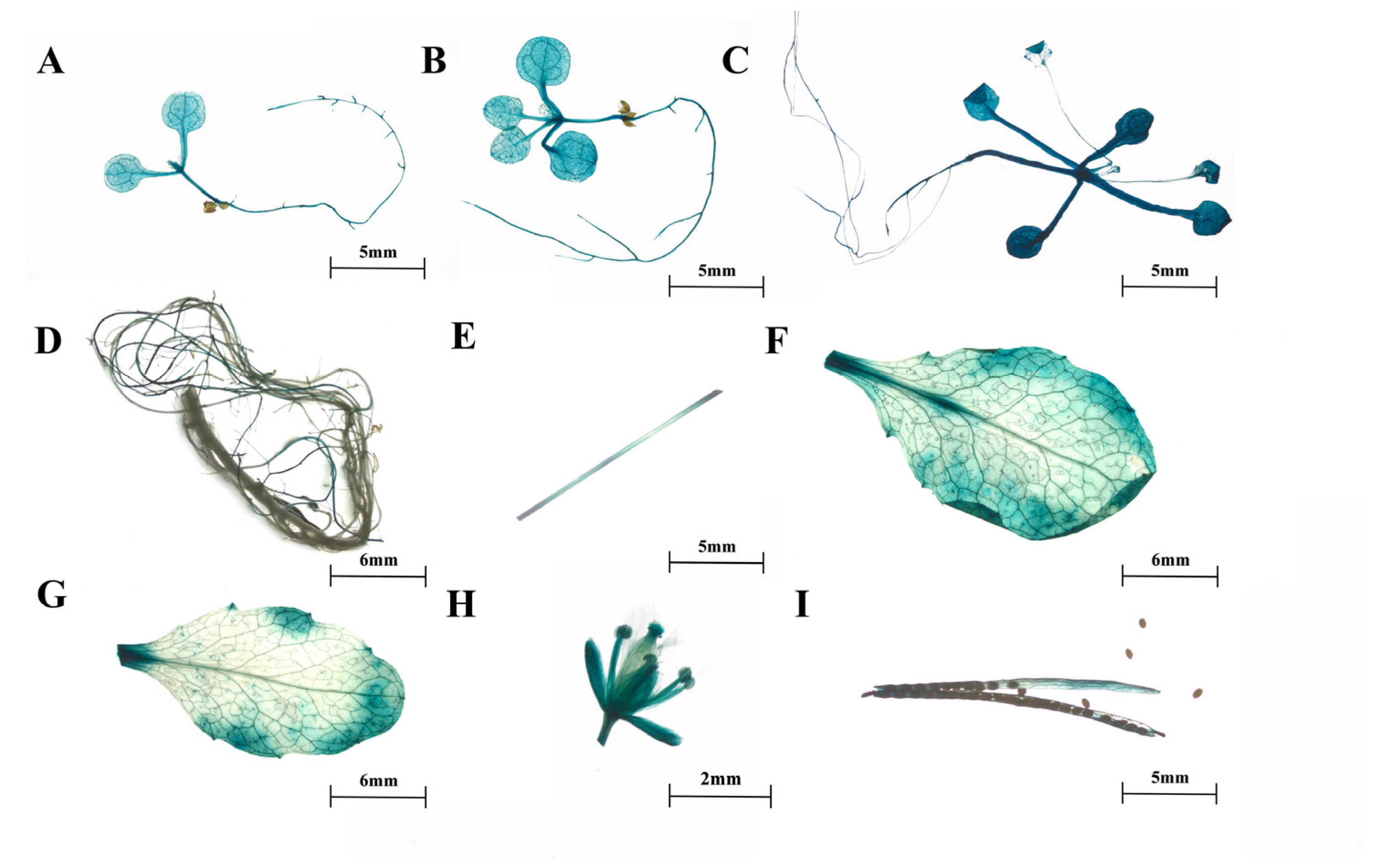
2.2. Expression Patterns of SmTATs
2.3. SmTATs Are Located in the Cytoplasm
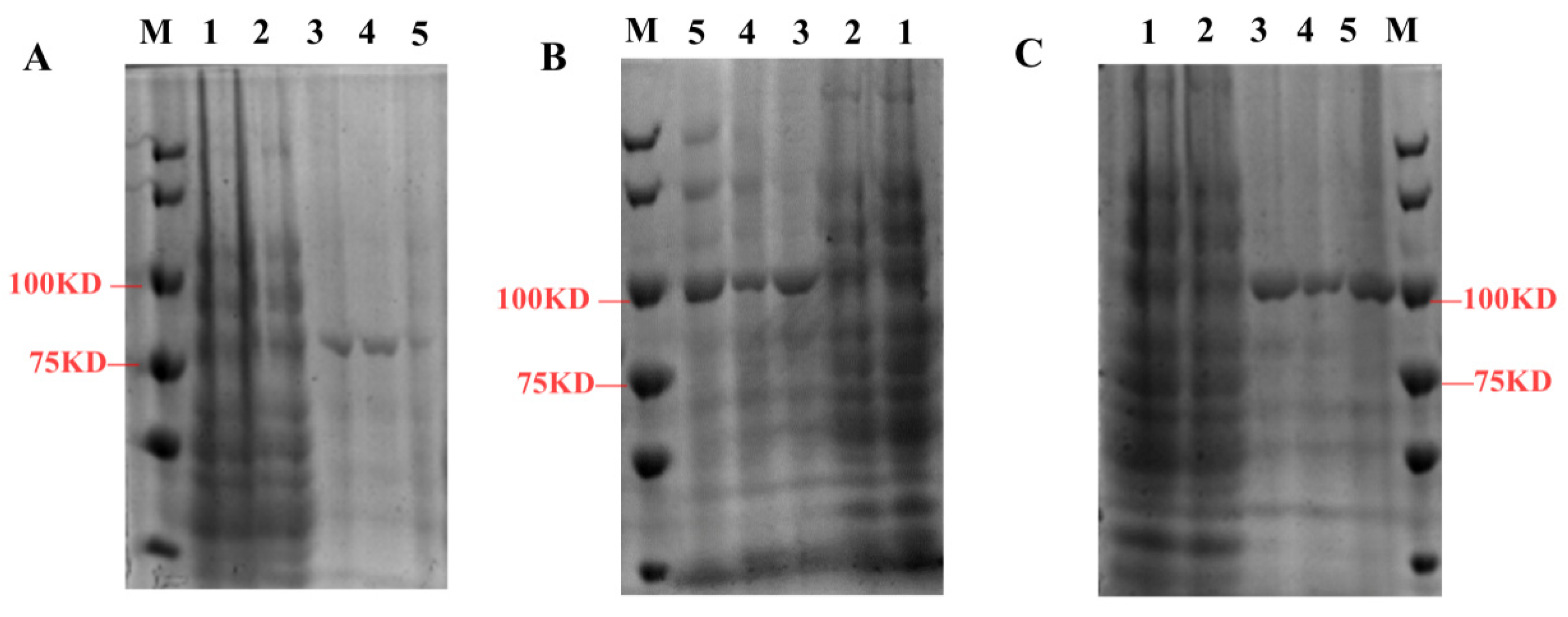
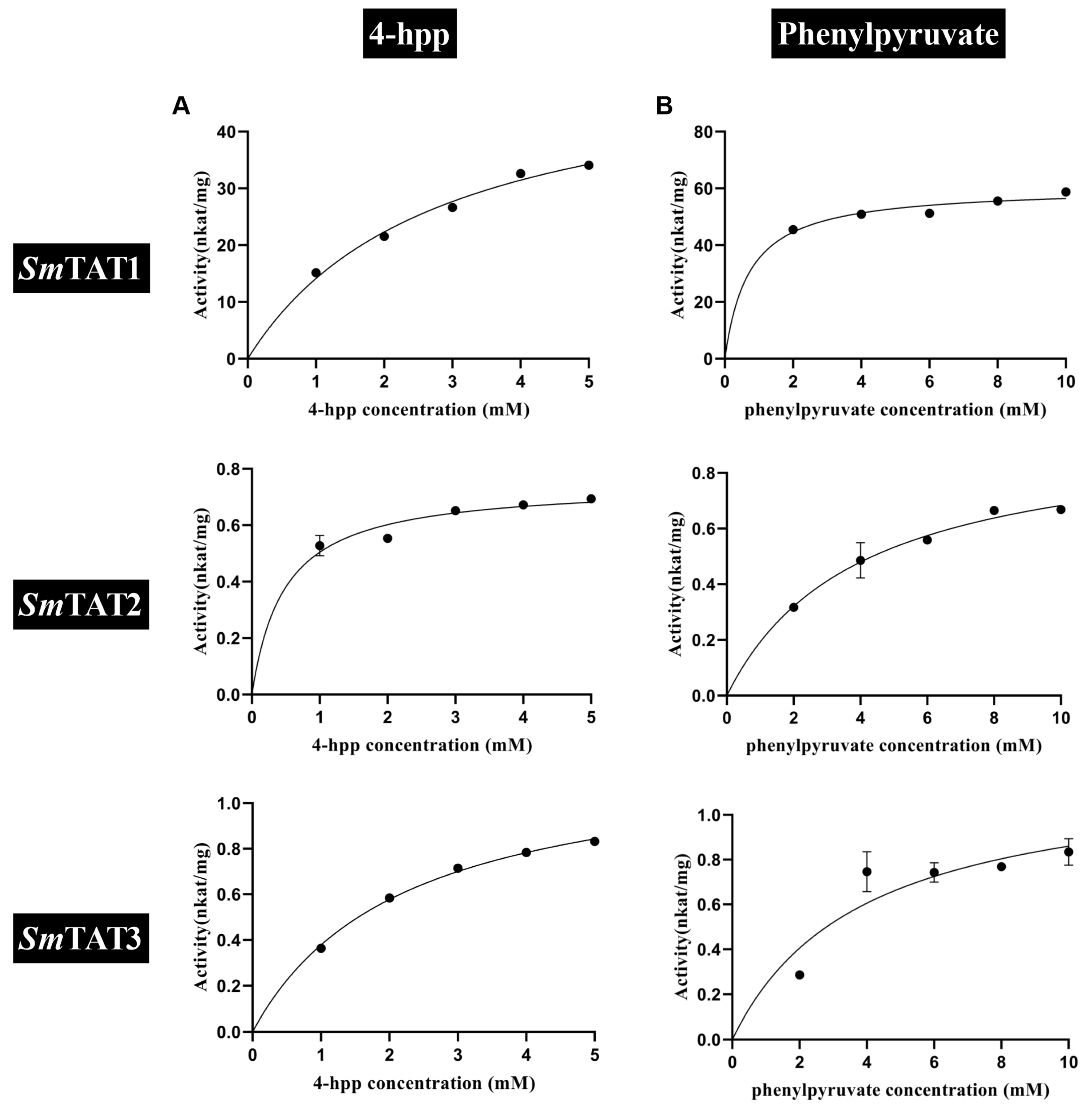
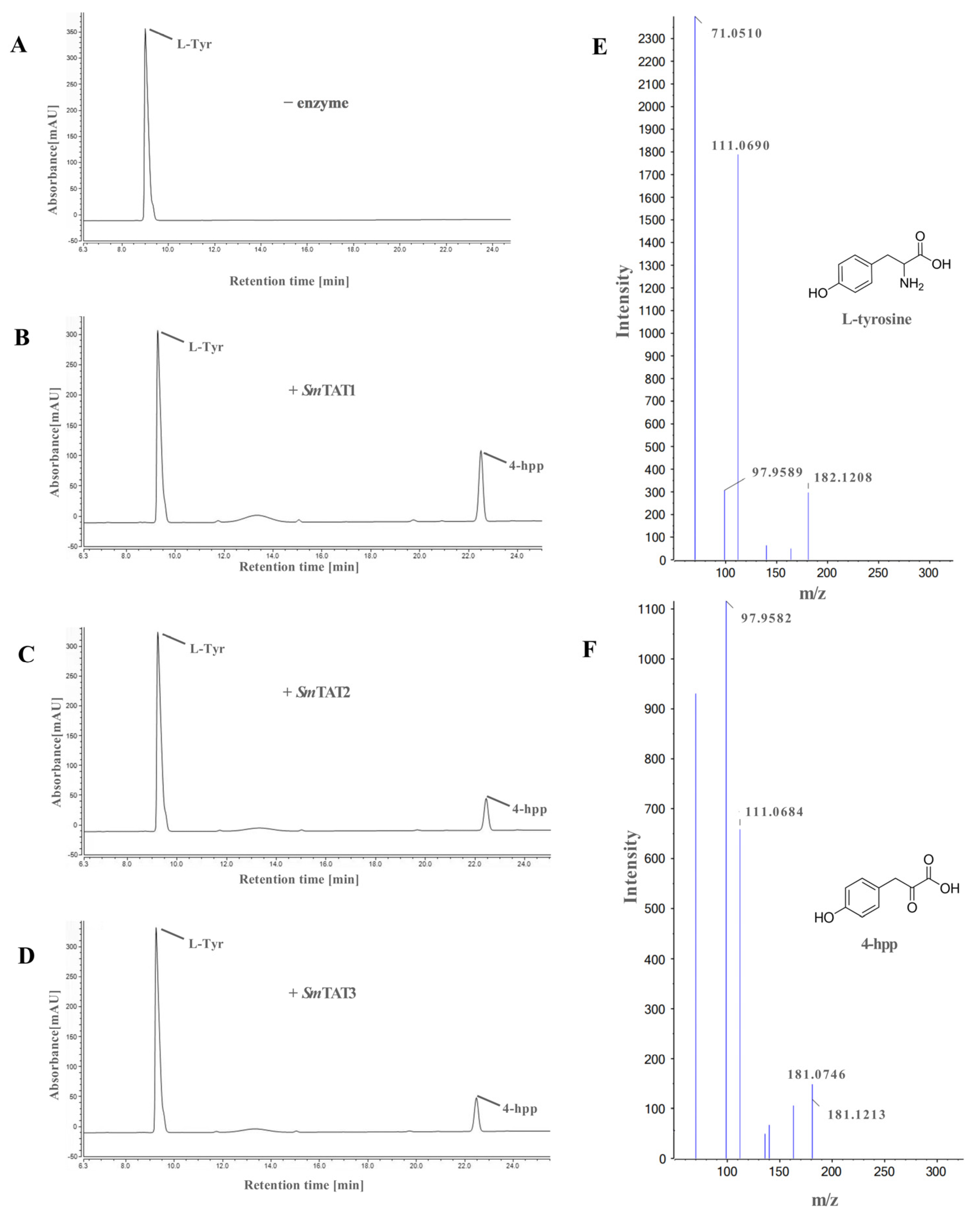
2.4. Prokaryotic Expression and Enzyme Assays of SmTATs
2.5. Transient Overexpression of SmTATs Increased the Content of 4-HPP in the Leaves of N. benthamiana
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Isolation of SmTAT1/2/3 and Sequence Analysis
4.3. Expression Analysis via qRT-PCR
4.4. Expression and Purification of SmTATs from Escherichia coli
4.5. In Vitro Enzyme Assays
4.6. Agrobacterium-Mediated Transient Expression in N. benthamiana
4.7. UPLC-QTOF/MS Analysis
4.8. Heterologous Expression of proSmHPPRs::GUS and GUS Staining
4.9. Subcellular Localization of SmTAT Proteins
4.10. Statistic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jung, I.; Kim, H.; Moon, S.; Lee, H.; Kim, B. Overview of Salvia miltiorrhiza as a potential therapeutic agent for various diseases: An update on efficacy and mechanisms of action. Antioxidants 2020, 9, 857. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zuo, Z.; Chow, M.S. Danshen: An overview of its chemistry, pharmacology, pharmacokinetics, and clinical use. J. Clin. Pharmacol. 2005, 45, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Guo, X.W.; Zheng, H.Z.; Li, D.P.; Jia, G.B.; Wang, J. Current progress of research on pharmacologic actions of salvianolic acid B. Chin. J. Integr. Med. 2012, 18, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Yun, C.; Gu, L.; Liu, J.; Yao, L.; Wang, W.; Wang, H. Melatonin enhances biomass, phenolic accumulation, and bioactivities of rosemary (Rosmarinus officinalis) in vitro shoots under UV-B stress. Physiol. Plant 2023, 16, e13956. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Sun, W.; Li, Q.; Li, Y.; Luo, H.; Song, J.; Sun, C.; Qian, J.; Zhu, Y.; Hayward, A.; et al. Genome-wide identification of phenolic acid biosynthetic genes in Salvia miltiorrhiza. Planta 2015, 241, 711–725. [Google Scholar] [CrossRef]
- Sasidharan, S.; Saudagar, P. Knockout of tyrosine aminotransferase gene by homologous recombination arrests growth and disrupts redox homeostasis in Leishmania Parasite. Parasitol. Res. 2022, 121, 3229–3241. [Google Scholar] [CrossRef]
- Mizukami, H.; Ellis, B.E. Rosmarinic acid formation and differential expression of tyrosine aminotransferase isoforms in Anchusa officinalis cell suspension cultures. Plant Cell Rep. 1991, 10, 321–324. [Google Scholar] [CrossRef]
- Lee, E.J.; Facchini, P.J. Tyrosine aminotransferase contributes to benzylisoquinoline alkaloid biosynthesis in opium poppy. Plant Physiol. 2011, 157, 1067–1078. [Google Scholar] [CrossRef]
- Zhang, S.; Yan, Y.; Wang, B. Selective responses of enzymes in the two parallel pathways of rosmarinic acid biosynthetic pathway to elicitors in Salvia miltiorrhiza hairy root cultures. J. Biosci. Bioeng. 2014, 117, 645–651. [Google Scholar] [CrossRef]
- Kim, Y.B.; Kim, J.K.; Uddin, M.R.; Xu, H.; Park, W.T.; Tuan, P.A.; Li, X.; Chung, E.; Lee, J.H.; Park, S.U. Metabolomics analysis and biosynthesis of rosmarinic acid in Agastache rugosa Kuntze treated with methyl jasmonate. PLoS ONE 2013, 8, e64199. [Google Scholar] [CrossRef]
- Wang, H.; Dong, Q.; Duan, D.; Zhao, S.; Li, M.; Van Nocker, S.; Ma, F.; Mao, K. Comprehensive genomic analysis of the TYROSINE AMINOTRANSFERASE (TAT) genes in apple (Malus domestica) allows the identification of MdTAT2 conferring tolerance to drought and osmotic stresses in plants. Plant Physiol. Biochem. 2018, 133, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Toda, K.; Maeda, H.A. Biochemical properties and subcellular localization of tyrosine aminotransferases in Arabidopsis thaliana. Phytochemistry 2016, 132, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Mehere, P.; Han, Q.; Lemkul, J.A.; Vavricka, C.J.; Robinson, H.; Bevan, D.R.; Li, J. Tyrosine aminotransferase: Biochemical and structural properties and molecular dynamics simulations. Protein Cell 2010, 1, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Koper, K.; Hataya, S.; Hall, A.G.; Takasuka, T.E.; Maeda, H.A. Biochemical characterization of plant aromatic aminotransferases. Methods Enzymol. 2023, 680, 35–83. [Google Scholar] [PubMed]
- Wang, M.; Toda, K.; Block, A.; Maeda, H.A. TAT1 and TAT2 tyrosine aminotransferases have both distinct and shared functions in tyrosine metabolism and degradation in Arabidopsis thaliana. J. Biol. Chem. 2019, 294, 3563–3576. [Google Scholar] [CrossRef]
- Antognoni, F.; Faudale, M.; Poli, F.; Biondi, S. Methyl jasmonate differentially affects tocopherol content and tyrosine amino transferase activity in cultured cells of Amaranthus caudatus and Chenopodium quinoa. Plant Biol. 2009, 11, 161–169. [Google Scholar] [CrossRef]
- Zhang, C.; Meng, S.; Li, M.; Zhao, Z. Transcriptomic insight into nitrogen uptake and metabolism of Populus simonii in response to drought and low nitrogen stresses. Tree Physiol. 2018, 38, 1672–1684. [Google Scholar] [CrossRef]
- Zhu, F.; Alseekh, S.; Koper, K.; Tong, H.; Nikoloski, Z.; Naake, T.; Liu, H.; Yan, J.; Brotman, Y.; Wen, W.; et al. Genome-wide association of the metabolic shifts underpinning dark-induced senescence in Arabidopsis. Plant Cell 2022, 34, 557–578. [Google Scholar] [CrossRef]
- Zhou, C.; Lin, C.; Xing, P.; Li, X.; Song, Z. SmGDB: Genome database of Salvia miltiorrhiza, an important TCM Plant. Genes Genom. 2022, 44, 699–707. [Google Scholar] [CrossRef]
- Liu, L.; Yang, D.; Liang, T.; Zhang, H.; He, Z.; Liang, Z. Phosphate starvation promoted the accumulation of phenolic acids by inducing the key enzyme genes in Salvia miltiorrhiza hairy roots. Plant Cell Rep. 2016, 35, 1933–1942. [Google Scholar] [CrossRef]
- Huang, B.; Yi, B.; Duan, Y.; Sun, L.; Yu, X.; Guo, J.; Chen, W. Characterization and expression profiling of tyrosine aminotransferase gene from Salvia miltiorrhiza (Dan-shen) in rosmarinic acid biosynthesis pathway. Mol. Biol. Rep. 2008, 35, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Habener, J.F. Regulation of polypeptide-hormone biosynthesis at the level of the genome. Am. J. Physiol. 1985, 249, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Zhang, Y.; Hua, W.P.; Wu, Y.C.; Jin, X.X.; Song, S.H.; Wang, Z.Z. Combination of transcriptomic and metabolomic analyses reveals a JAZ repressor in the jasmonate signaling pathway of Salvia miltiorrhiza. Sci. Rep. 2015, 5, 14048. [Google Scholar] [CrossRef]
- Hussain, H.; Fisher, D.I.; Abbott, W.M.; Roth, R.G.; Dickson, A.J. Use of a protein engineering strategy to overcome limitations in the production of “Difficult to Express” recombinant proteins. Biotechnol. Bioeng. 2017, 114, 2348–2359. [Google Scholar] [CrossRef] [PubMed]
- Jäger, V.D.; Kloss, R.; Grünberger, A.; Seide, S.; Hahn, D.; Karmainski, T.; Piqueray, M.; Embruch, J.; Longerich, S.; Mackfeld, U.; et al. Tailoring the properties of (catalytically)-active inclusion bodies. Microb. Cell Factories 2019, 18, 33. [Google Scholar] [CrossRef]
- Ru, M.; Wang, K.; Bai, Z.; Peng, L.; He, S.; Wang, Y.; Liang, Z. A tyrosine aminotransferase involved in rosmarinic acid biosynthesis in Prunella vulgaris L. Sci. Rep. 2017, 7, 4892. [Google Scholar] [CrossRef]
- Rippert, P.; Puyaubert, J.; Grisollet, D.; Derrier, L.; Matringe, M. Tyrosine and phenylalanine are synthesized within the plastids in Arabidopsis. Plant Physiol. 2009, 149, 1251–1260. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, L.; Gao, S.; Saechao, S.; Di, P.; Chen, J.; Chen, W. The c4h, tat, hppr and hppd genes prompted engineering of rosmarinic acid biosynthetic pathway in Salvia miltiorrhiza hairy root cultures. PLoS ONE 2011, 6, e29713. [Google Scholar] [CrossRef]
- Du, T.; Niu, J.; Su, J.; Li, S.; Guo, X.; Li, L.; Cao, X.; Kang, J. SmbHLH37 functions antagonistically with SmMYC2 in regulating jasmonate-mediated biosynthesis of phenolic acids in Salvia miltiorrhiza. Plant Sci. 2018, 9, 1720. [Google Scholar] [CrossRef]
- Li, L.; Wang, D.; Zhou, L.; Yu, X.; Yan, X.; Zhang, Q.; Li, B.; Liu, Y.; Zhou, W.; Cao, X.; et al. JA-responsive transcription factor SmMYB97 promotes phenolic acid and tanshinone accumulation in Salvia miltiorrhiza. J. Agric. Food Chem. 2020, 68, 14850–14862. [Google Scholar] [CrossRef]
- Yang, R.; Li, S.; Dong, S.; Wang, L.; Qin, H.; Zhan, H.; Wang, D.; Cao, X.; Xu, H. SmJAZ4 interacts with SmMYB111 or SmMYC2 to inhibit the synthesis of phenolic acids in Salvia miltiorrhiza. Plant Sci. 2023, 327, 111565. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.J.; Wu, Y.C.; Wang, S.Q.; Niu, J.F.; Cao, X.Y. SmbHLH53 is relevant to jasmonate signaling and plays dual roles in regulating the genes for enzymes in the pathway for salvianolic acid B biosynthesis in Salvia miltiorrhiza. Gene 2020, 756, 144920. [Google Scholar] [CrossRef] [PubMed]
- De Vandesompele, J.; Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2022, 3, RESEARCH0034. [Google Scholar]
- Yang, Q.Q.; Hua, W.P.; Zou, H.L.; Yang, J.X.; Wang, X.Z.; Zhang, T.; Wang, D.H.; Zhu, X.J.; Cao, X.Y. Overexpression of SmLAC25 promotes lignin accumulation and decreases salvianolic acid content in Salvia miltiorrhiza. Plant Sci. 2022, 325, 111462. [Google Scholar] [CrossRef]
- Prabhu, P.R.; Hudson, A.O. Identification and partial characterization of an L-tyrosine aminotransferase (TAT) from Arabidopsis thaliana. Biochem. Res. Int. 2010, 2010, 549572. [Google Scholar] [CrossRef]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef]
- Dong, S.; Kong, G.; Qutob, D.; Yu, X.; Tang, J.; Kang, J.; Dai, T.; Wang, H.; Gijzen, M.; Wang, Y. The NLP toxin family in Phytophthora sojae includes rapidly evolving groups that lack necrosis-inducing activity. Mol. Plant Microbe Interact. 2012, 25, 896–909. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. Cell Mol. Biol. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Gomes-Junior, R.A.; Moldes, C.A.; Delite, F.S.; Pompeu, G.B.; Gratão, P.L.; Mazzafera, P.; Lea, P.J.; Azevedo, R.A. Antioxidant metabolism of coffee cell suspension cultures in response to cadmium. Chemosphere 2006, 65, 1330. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable-Substrate | Co-Substrate | Km (mM) | Vmax (nmol s−1 mg−1) | kcat (s−1) | kcat/Km (mM−1 s−1) |
|---|---|---|---|---|---|
| SmTAT1 | |||||
| Tyr | α-KG | 0.31 ± 0.11 | 114.7 ± 5.41 | 19.83 ± 0.93 | 74.4 ± 29.4 |
| Phe | α-KG | 3.08 ± 0.99 | 72.69 ± 8.31 | 12.57 ± 1.43 | 4.68 ± 2.02 |
| 4-HPP | Glu | 2.77 ± 0.52 | 53.2 ± 4.61 | 9.21 ± 0.78 | 3.5 ± 0.94 |
| PPY | Glu | 0.71 ± 0.21 | 60.46 ± 2.48 | 10.45 ± 0.39 | 16.31 ± 4.62 |
| SmTAT2 | |||||
| Tyr | α-KG | 3.47 ± 1.69 | 0.81 ± 0.21 | 0.15 ± 0.03 | 0.06 ± 0.04 |
| Phe | α-KG | 6.93 ± 3.39 | 1.81 ± 0.47 | 0.33 ± 0.08 | 0.68 ± 0.24 |
| 4-HPP | Glu | 0.48 ± 0.16 | 7.47 ± 0.45 | 1.35 ± 0.08 | 3.23 ± 1.24 |
| PPY | Glu | 3.94 ± 0.99 | 0.95 ± 0.11 | 0.17 ± 0.02 | 0.05 ± 0.02 |
| SmTAT3 | |||||
| Tyr | α-KG | 2.73 ± 1.61 | 0.90 ± 0.26 | 0.16 ± 0.04 | 0.1 ± 0.08 |
| Phe | α-KG | 5.47 ± 3.23 | 2.02 ± 0.59 | 0.35 ± 0.11 | 0.12 ± 0.09 |
| 4-HPP | Glu | 2.22 ± 0.22 | 12.17 ± 0.51 | 2.12 ± 0.09 | 0.97 ± 0.14 |
| PPY | Glu | 3.91 ± 2.01 | 1.21 ± 0.26 | 0.21 ± 0.05 | 0.08 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, S.; Wang, L.; Qin, H.; Zhan, H.; Wang, D.; Cao, X. Expression Patterns and Functional Analysis of Three SmTAT Genes Encoding Tyrosine Aminotransferases in Salvia miltiorrhiza. Int. J. Mol. Sci. 2023, 24, 15575. https://doi.org/10.3390/ijms242115575
Dong S, Wang L, Qin H, Zhan H, Wang D, Cao X. Expression Patterns and Functional Analysis of Three SmTAT Genes Encoding Tyrosine Aminotransferases in Salvia miltiorrhiza. International Journal of Molecular Sciences. 2023; 24(21):15575. https://doi.org/10.3390/ijms242115575
Chicago/Turabian StyleDong, Shuai, Long Wang, Huiting Qin, Hongbin Zhan, Donghao Wang, and Xiaoyan Cao. 2023. "Expression Patterns and Functional Analysis of Three SmTAT Genes Encoding Tyrosine Aminotransferases in Salvia miltiorrhiza" International Journal of Molecular Sciences 24, no. 21: 15575. https://doi.org/10.3390/ijms242115575
APA StyleDong, S., Wang, L., Qin, H., Zhan, H., Wang, D., & Cao, X. (2023). Expression Patterns and Functional Analysis of Three SmTAT Genes Encoding Tyrosine Aminotransferases in Salvia miltiorrhiza. International Journal of Molecular Sciences, 24(21), 15575. https://doi.org/10.3390/ijms242115575