
Cordyceps militaris Solid Medium Extract Alleviates Lipoteichoic Acid-Induced MH-S Inflammation by Inhibiting TLR2/NF-κB/NLRP3 Pathways
Abstract
:1. Introduction
2. Results
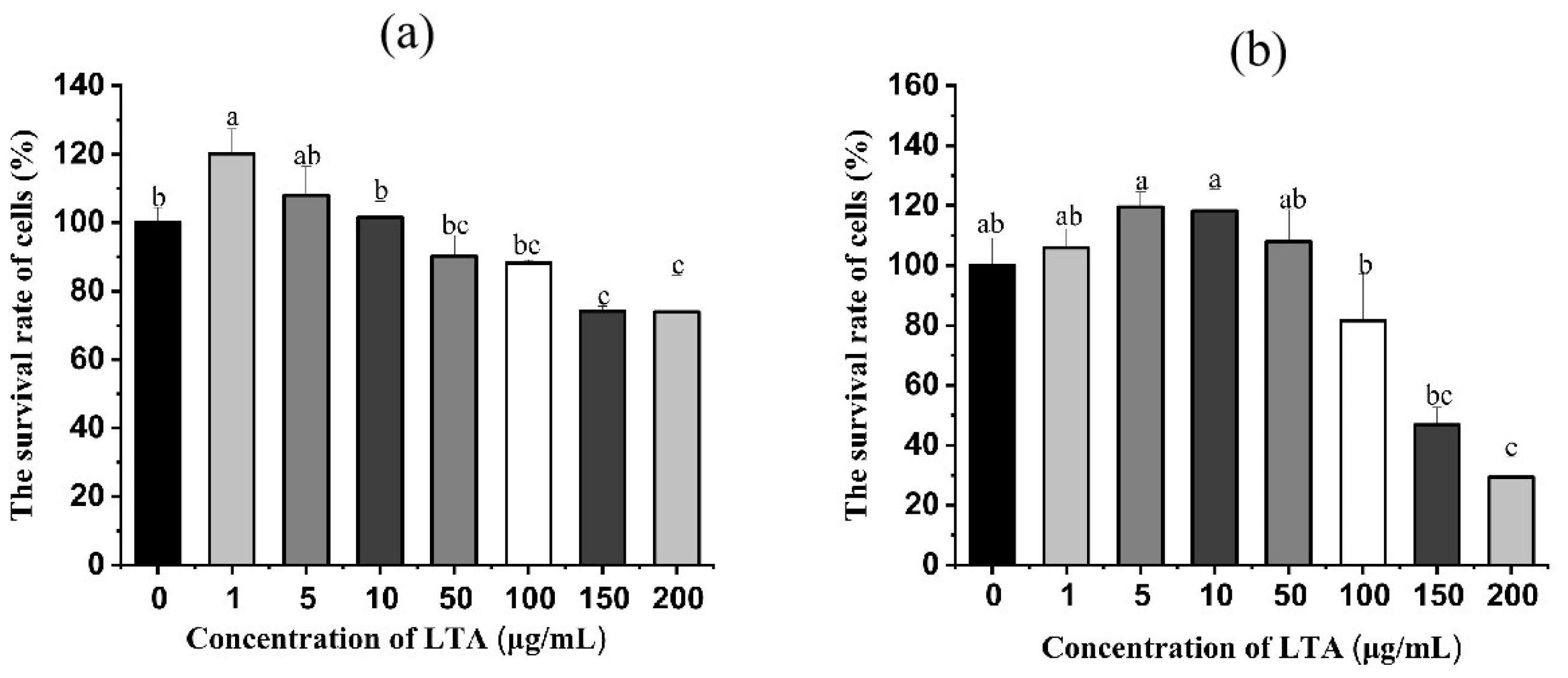
2.1. Effect of LTA on Survival Rate of MH-S Cells
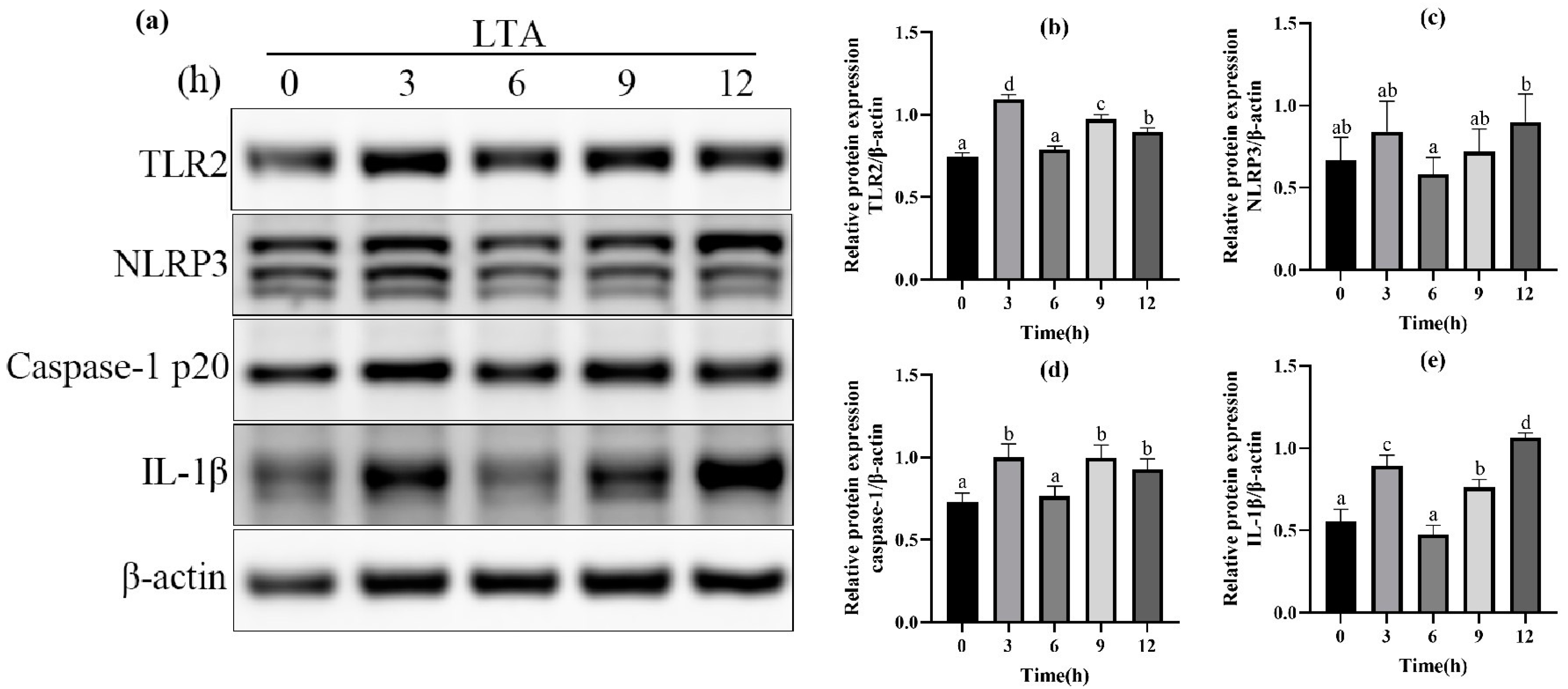
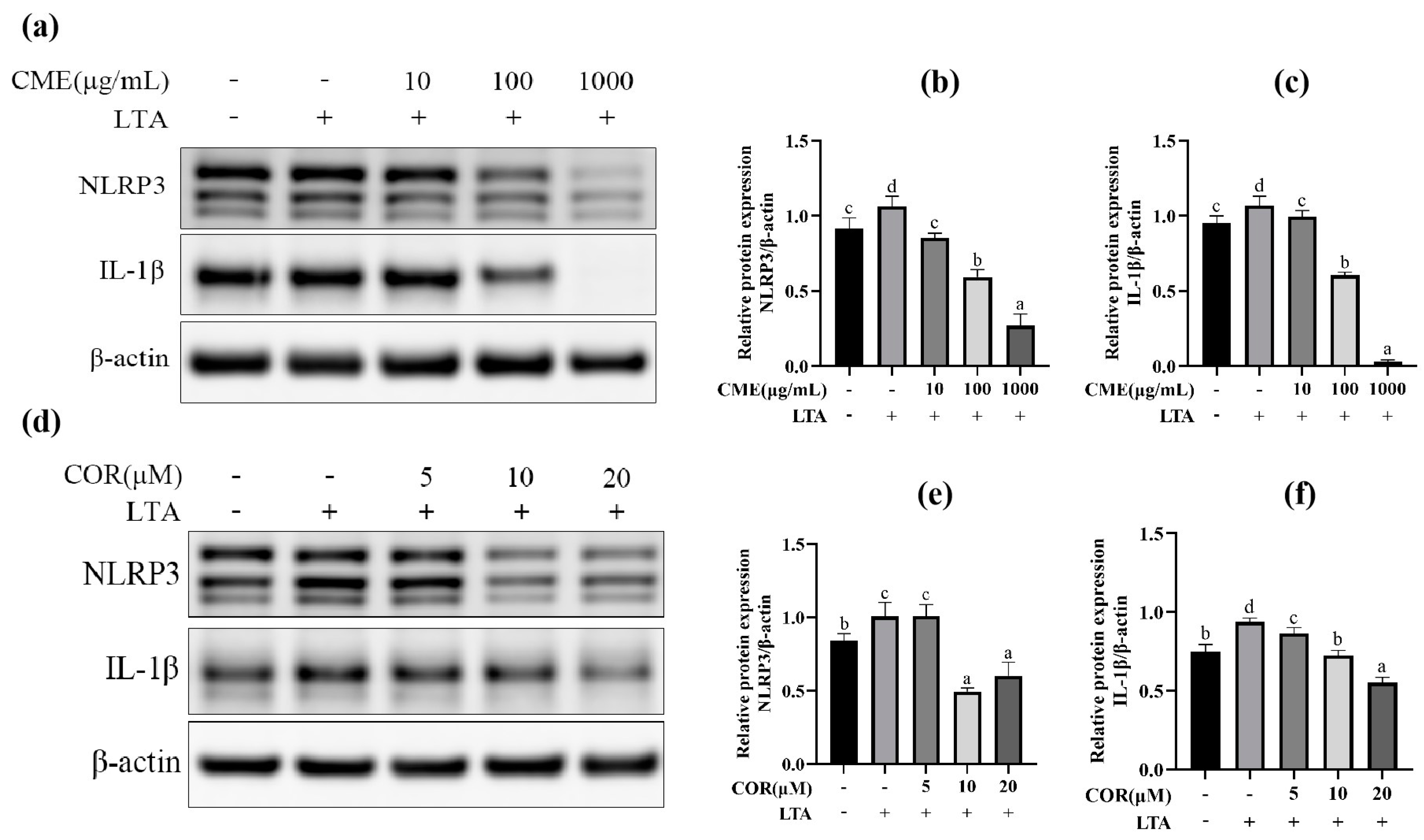
2.2. Effect of LTA Stimulation on Protein Expression
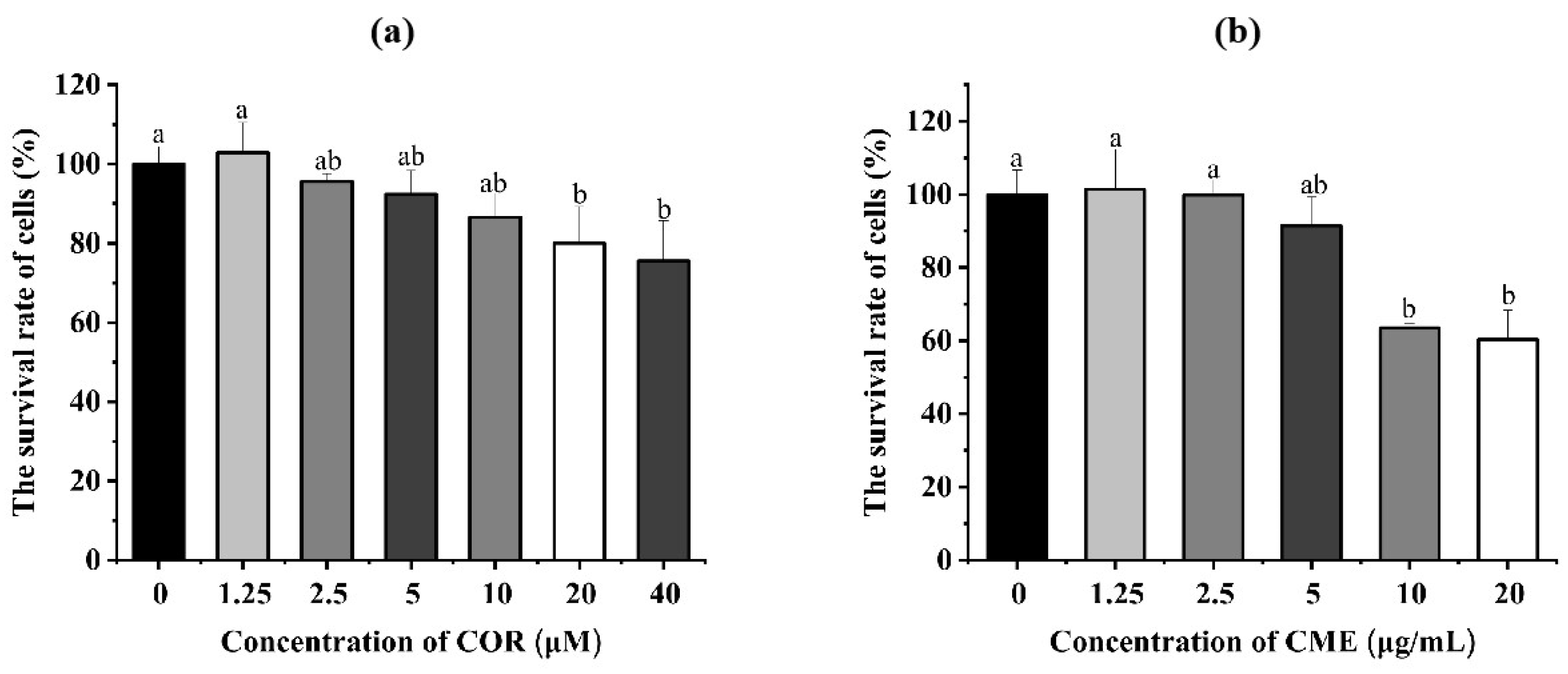
2.3. Effect of CME and COR on Survival Rate of MH-S Cells
2.4. Screening for Optimal Concentration of CME and COR
2.5. The Effect of CME and COR on Secretion of Inflammation Factors in LTA-Induced MH-S Cells
2.6. CME and COR Inhibit the Relative mRNA Expression of Inflammatory Factor in LTA-Induced MH-S Cells
2.7. CME Inhibited LTA-Induced Inflammation in MH-S Cells through TLR2/NF-κB Signal Pathway
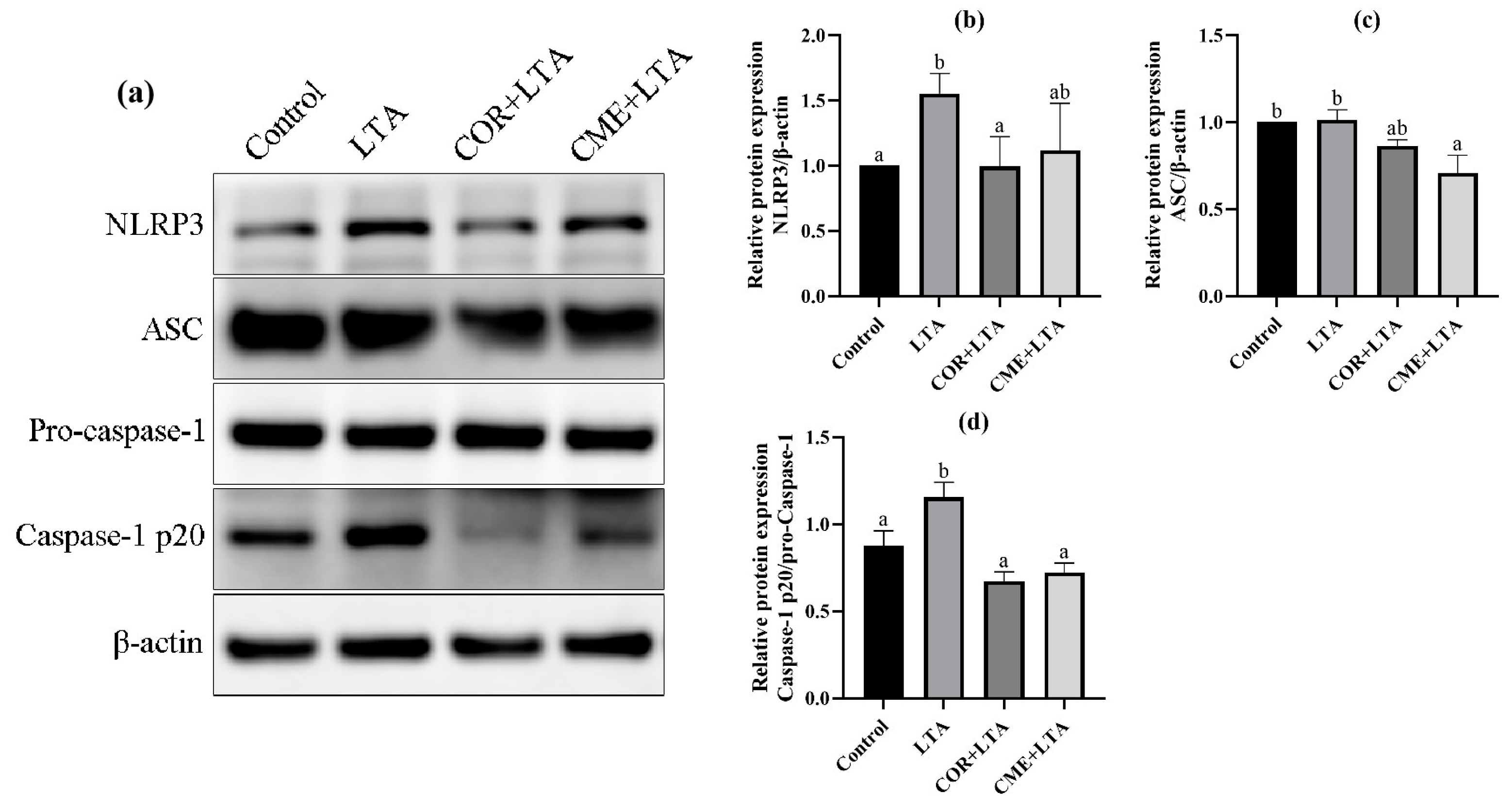
2.8. CME Inhibited LTA-Induced MH-S Cell Inflammatory Response through NLRP3
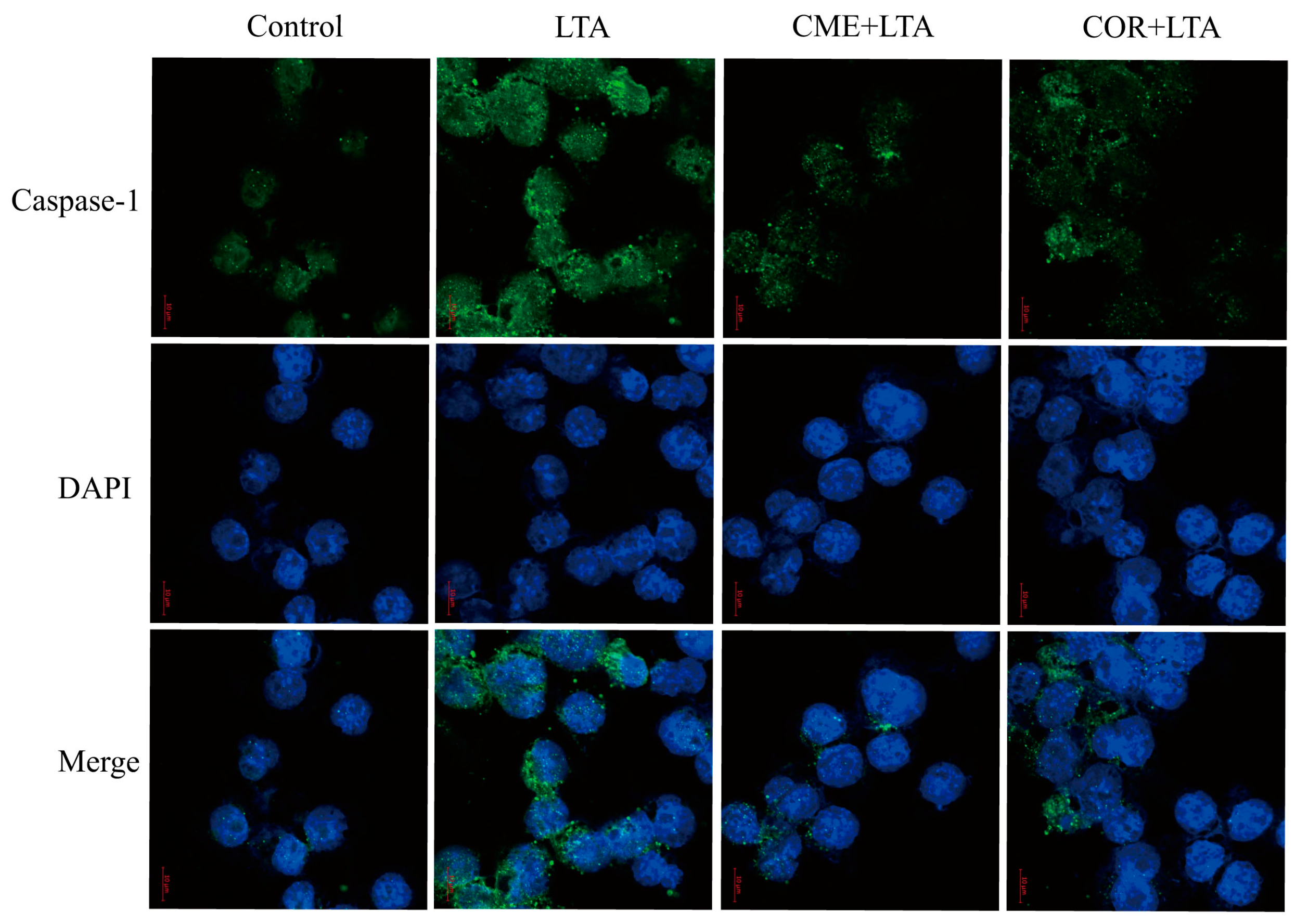
2.9. Effect of CME on Caspase-1 Activation of MH-S Cells Induced by LTA
3. Discussion
4. Materials and Methods
4.1. Reagent
4.2. The Methods of Extraction
4.3. Cell Culture
4.4. LTA Stimulates MH-S Cell Inflammation
4.5. Screening for Optimal Concentration of COR and CME
4.6. Experimental Grouping and Drug Treatment
4.7. ELISA Assay
4.8. ELISA Detection of Relative mRNA Expression by Real-Time PCR
4.9. WB Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CME | Cordyceps militaris solid medium extract |
| COR | Cordycepin |
| S. aureus | Staphylococcus aureus |
| LTA | Lipoteichoic acid |
| PAMP | Pathogen-associated molecules |
| DAMP | Damage-associated |
| TLR | Toll-like receptors |
| NLR | NOD-like receptors |
| ASC | Apoptosis-associated speck-like protein containing a CARD |
| pro-IL-1β | Pro-interleukinogen-1β |
| Nrf2 | Nuclearfactor erythroidderived 2-like 2 |
| HO-1. | Heme oxygenase-1 |
| AMPK | Amp-activated protein kinase |
References
- Hesser, A.R. Mechanism of Staphylococcus aureus Lipoteichoic Acid Length Regulation, Inhibition of Wall Teichoic Acid Biosynthesis, and Physiology of Teichoic Acids; Harvard University: Cambridge, MA, USA, 2020. [Google Scholar]
- Tsugami, Y.; Wakasa, H.; Kawahara, M.; Nishimura, T.; Kobayashi, K. Lipopolysaccharide and lipoteichoic acid influence milk production ability via different early responses in bovine mammary epithelial cells. Exp. Cell Res. 2021, 400, 112472. [Google Scholar] [CrossRef]
- Jang, J.; Kim, W.; Kim, K.; Chung, S.I.; Shim, Y.J.; Kim, S.M.; Yoon, Y. Lipoteichoic acid upregulates NF-κB and proinflammatory cytokines by modulating β-catenin in bronchial epithelial cells. Mol. Med. Rep. 2015, 12, 4720–4726. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Umemura, A.; Sanchez-Lopez, E.; Liang, S.; Shalapour, S.; Wong, J.; He, F.; Boassa, D.; Perkins, G.; Ali, S.R.; et al. NF-κB Restricts Inflammasome Activation via Elimination of Damaged Mitochondria. Cell 2016, 164, 896–910. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Xu, D.; Tian, Y.; Qu, X.; Sheng, M.; Lin, Y.; Ke, M.; Jiang, L.; Xia, Q.; Kaldas, F.M.; et al. Novel role of macrophage TXNIP-mediated CYLD-NRF2-OASL1 axis in stress-induced liver inflammation and cell death. JHEP Rep. Innov. Hepatol. 2022, 4, 100532. [Google Scholar] [CrossRef]
- Wu, B.Q.; Luo, J.M.; Wang, Y.H.; Shi, Y.F.; Liu, H.; Ba, J.H.; Zhang, T.T. Inhibitory effects of simvastatin on staphylococcus aureus lipoteichoic acid-induced inflammation in human alveolar macrophages. Clin. Exp. Med. 2014, 14, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, D.; Liu, T.; Wang, J.F.; Wu, J.X.; Zhao, J.P.; Xu, J.W.; Zhang, J.T.; Dong, L. FSTL1 aggravates OVA-induced inflammatory responses by activating the NLRP3/IL-1β signaling pathway in mice and macrophages. Inflamm. Res. 2021, 70, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jin, H.; Ye, D.; Wang, J.; Ao, X.; Dong, M.; Niu, W. Enterococcus faecalis Lipoteichoic Acid-induced NLRP3 Inflammasome via the Activation of the Nuclear Factor Kappa B Pathway. J. Endod. 2016, 42, 1093–1100. [Google Scholar] [CrossRef]
- Dominic, A.; Le, N.T.; Takahashi, M. Loop between NLRP3 Inflammasome and Reactive Oxygen Species. Antioxid. Redox Signal. 2022, 36, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Latz, E.; Xiao, T.S.; Stutz, A. Activation and regulation of the inflammasomes. Nat. Rev. Immunol. 2013, 13, 397–411. [Google Scholar] [CrossRef]
- Luo, M.; Yan, D.; Sun, Q.; Tao, J.; Xu, L.; Sun, H.; Zhao, H. Ginsenoside Rg1 attenuates cardiomyocyte apoptosis and inflammation via the TLR4/NF-kB/NLRP3 pathway. J. Cell. Biochem. 2020, 121, 2994–3004. [Google Scholar] [CrossRef]
- Jin, X.; Liu, M.Y.; Zhang, D.F.; Zhong, X.; Du, K.; Qian, P.; Yao, W.F.; Gao, H.; Wei, M.J. Baicalin mitigates cognitive impairment and protects neurons from microglia-mediated neuroinflammation via suppressing NLRP3 inflammasomes and TLR4/NF-κB signaling pathway. CNS Neurosci. Ther. 2019, 25, 575–590. [Google Scholar] [CrossRef]
- Wang, Q.; Ou, Y.; Hu, G.; Wen, C.; Yue, S.; Chen, C.; Xu, L.; Xie, J.; Dai, H.; Xiao, H.; et al. Naringenin attenuates non-alcoholic fatty liver disease by down-regulating the NLRP3/NF-κB pathway in mice. Br. J. Pharmacol. 2020, 177, 1806–1821. [Google Scholar] [CrossRef] [PubMed]
- Jo, E.; Jang, H.J.; Shen, L.; Yang, K.E.; Jang, M.S.; Huh, Y.H.; Yoo, H.S.; Park, J.; Jang, I.S.; Park, S.J. Cordyceps militaris Exerts Anticancer Effect on Non-Small Cell Lung Cancer by Inhibiting Hedgehog Signaling via Suppression of TCTN3. Integr. Cancer Ther. 2020, 19, 1534735420923756. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Wei, Y.; Song, P.; Li, Y.; Zhang, T.; Feng, Q.; Xu, G. Cordycepin inhibits LPS-induced acute lung injury by inhibiting inflammation and oxidative stress. Eur. J. Pharmacol. 2018, 818, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Naura, A.S.; Errami, Y.; Ju, J.; Boulares, A.H. Cordycepin blocks lung injury-associated inflammation and promotes BRCA1-deficient breast cancer cell killing by effectively inhibiting PARP. Mol. Med. 2011, 17, 893–900. [Google Scholar] [CrossRef]
- Jo, E.; Jang, H.J.; Yang, K.E.; Jang, M.S.; Huh, Y.H.; Yoo, H.S.; Park, J.S.; Jang, I.S.; Park, S.J. Cordyceps militaris induces apoptosis in ovarian cancer cells through TNF-α/TNFR1-mediated inhibition of NF-κB phosphorylation. BMC Complement. Med. Ther. 2020, 20, 1. [Google Scholar] [CrossRef]
- Liao, X.Z.; Gao, Y.; Zhao, H.W.; Zhou, M.; Chen, D.L.; Tao, L.T.; Guo, W.; Sun, L.L.; Gu, C.Y.; Chen, H.R.; et al. Cordycepin Reverses Cisplatin Resistance in Non-small Cell Lung Cancer by Activating AMPK and Inhibiting AKT Signaling Pathway. Front. Cell Dev. Biol. 2020, 8, 609285. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, J.; Su, Y.; Li, M.; Wen, J.; Li, S. Cordycepin induces M1/M2 macrophage polarization to attenuate the liver and lung damage and immunodeficiency in immature mice with sepsis via NF-κB/p65 inhibition. J. Pharm. Pharmacol. 2022, 74, 227–235. [Google Scholar] [CrossRef]
- Liu, Y.; Zuo, J.; Tao, Y.; Liu, W. Protective effect of Cordyceps polysaccharide on hydrogen peroxide-induced mitochondrial dysfunction in HL-7702 cells. Mol. Med. Rep. 2013, 7, 747–754. [Google Scholar] [CrossRef]
- Rupa, E.J.; Li, J.F.; Arif, M.H.; Yaxi, H.; Puja, A.M.; Chan, A.J.; Hoang, V.A.; Kaliraj, L.; Yang, D.C.; Kang, S.C. Cordyceps militaris Fungus Extracts-Mediated Nanoemulsion for Improvement Antioxidant, Antimicrobial, and Anti-Inflammatory Activities. Molecules 2020, 25, 5733. [Google Scholar] [CrossRef]
- Das, S.K.; Masuda, M.; Sakurai, A.; Sakakibara, M. Medicinal uses of the mushroom Cordyceps militaris: Current state and prospects. Fitoterapia 2010, 81, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wang, Y.; Liu, C.; Huang, Y.; He, L.; Cai, X.; Lu, J.; Liu, Y.; Wang, D. Cordyceps militaris fruit body extract ameliorates membranous glomerulonephritis by attenuating oxidative stress and renal inflammation via the NF-κB pathway. Food Funct. 2016, 7, 2006–2015. [Google Scholar] [CrossRef]
- Hsu, C.-H.; Sun, H.-L.; Sheu, J.-N.; Ku, M.-S.; Hu, C.-M.; Chan, Y.; Lue, K.-H. Effects of the immunomodulatory agent Cordyceps militaris on airway inflammation in a mouse asthma model. Pediatr. Neonatol. 2008, 49, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Tao, T.; Zhang, J.; Su, A.; Zhao, L.; Chen, H.; Hu, Q. Comparison of effects on colitis-associated tumorigenesis and gut microbiota in mice between Ophiocordyceps sinensis and Cordyceps militaris. Phytomed. Int. J. Phytother. Phytopharm. 2021, 90, 153653. [Google Scholar] [CrossRef]
- Huang, R.; Zhu, Z.; Wu, S.; Wang, J.; Chen, M.; Liu, W.; Huang, A.; Zhang, J.; Wu, Q.; Ding, Y. Polysaccharides from Cordyceps militaris prevent obesity in association with modulating gut microbiota and metabolites in high-fat diet-fed mice. Food Res. Int. 2022, 157, 111197. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Lin, J.; Guo, L.; Wang, X.; Tian, S.; Liu, C.; Zhao, Y.; Zhao, R. Advances in research on Cordyceps militaris degeneration. Appl. Microbiol. Biotechnol. 2019, 103, 7835–7841. [Google Scholar] [CrossRef]
- Ren, S.Y.; Zhao, C.Y.; Song, H.Y.; Zhao, H.L.; Sun, J.D. Optimum of polysaccharide distillation on scrap Cordyceps militaris medium. J. Chin. Med. Mater. 2008, 31, 342–343. [Google Scholar]
- Ni, H.; Zhou, X.H.; Li, H.H.; Huang, W.F. Column chromatographic extraction and preparation of cordycepin from Cordyceps militaris waster medium. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 2135–2141. [Google Scholar] [CrossRef]
- Wei, H.P.; Ye, X.L.; Zhang, H.Y.; Li, X.G.; Zhong, Y.J. Investigations on cordycepin production by solid culture of Cordyceps militaris. China J. Chin. Mater. Medica 2008, 33, 2159–2162. [Google Scholar]
- Li, C.; Yan, A.; Cai, C.; Liu, Z. Fast determination of adenosine and cordycepin in Cordyceps and its deserted solid medium. Chin. J. Chromatogr. 2012, 30, 711–715. [Google Scholar] [CrossRef]
- Chen, J.M.; Zheng, T.T.; Guo, L.Q.; Lin, Q.F.; Zheng, Q.W. Regulation of Cordyceps pupa solid fermentation on immune and intestinal flora in mice. Chin. J. Food Sci. 2022, 22, 76–87. (In Chinese) [Google Scholar]
- He, X.Z. Application of Solid Fermentation Extract of Cordyceps Pupa in Shampoo; Shanghai University of Applied Technology: Shanghai, China, 2022. (In Chinese) [Google Scholar]
- Cui, Y.R.; Qu, F.; Zhong, W.J.; Yang, H.H.; Zeng, J.; Huang, J.H.; Liu, J.; Zhang, M.Y.; Zhou, Y.; Guan, C.X. Beneficial effects of aloperine on inflammation and oxidative stress by suppressing necroptosis in lipopolysaccharide-induced acute lung injury mouse model. Phytomed. Int. J. Phytother. Phytopharm. 2022, 100, 154074. [Google Scholar] [CrossRef] [PubMed]
- Maguire, O.; O’Loughlin, K.; Minderman, H. Simultaneous assessment of NF-κB/p65 phosphorylation and nuclear localization using imaging flow cytometry. J. Immunol. Methods 2015, 423, 3–11. [Google Scholar] [CrossRef]
- Kapetanovic, R.; Jouvion, G.; Fitting, C.; Parlato, M.; Blanchet, C.; Huerre, M.; Cavaillon, J.M.; Adib-Conquy, M. Contribution of NOD2 to lung inflammation during Staphylococcus aureus-induced pneumonia. Microbes Infect. 2010, 12, 759–767. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-κB in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef]
- Ginsburg, I. Role of lipoteichoic acid in infection and inflammation. Lancet Infect. Dis. 2002, 2, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Jiang, K.; Wu, H.; Qiu, C.; Deng, G.; Peng, X. Polydatin reduces Staphylococcus aureus lipoteichoic acid-induced injury by attenuating reactive oxygen species generation and TLR2-NFκB signalling. J. Cell. Mol. Med. 2017, 21, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, N.G.; Kocan, M.; Schofield, L.; Pfleger, K.D.; Eriksson, E.M. Investigation of interactions between TLR2, MyD88 and TIRAP by bioluminescence resonance energy transfer is hampered by artefacts of protein overexpression. PLoS ONE 2018, 13, e0202408. [Google Scholar] [CrossRef]
- Lambrecht, B.N. Alveolar macrophage in the driver’s seat. Immunity 2006, 24, 366–368. [Google Scholar] [CrossRef]
- Kang, H.J.; Baik, H.W.; Kim, S.J.; Lee, S.G.; Ahn, H.Y.; Park, J.S.; Park, S.J.; Jang, E.J.; Park, S.W.; Choi, J.Y.; et al. Cordyceps militaris Enhances Cell-Mediated Immunity in Healthy Korean Men. J. Med. Food 2015, 18, 1164–1172. [Google Scholar] [CrossRef]
- Jeong, J.W.; Jin, C.Y.; Kim, G.Y.; Lee, J.D.; Park, C.; Kim, G.D.; Kim, W.J.; Jung, W.K.; Seo, S.K.; Choi, I.W.; et al. Anti-inflammatory effects of cordycepin via suppression of inflammatory mediators in BV2 microglial cells. Int. Immunopharmacol. 2010, 10, 1580–1586. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.S.; Choi, Y.J.; Kim, H.J.; Lee, J.Y.; Nam, B.H.; Lee, J.D.; Lee, S.W.; Seo, S.Y.; Jeong, M.H. The Anti-inflammatory Effects of Water Extract from Cordyceps militaris in Murine Macrophage. Mycobiology 2010, 38, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.N.; Zhang, X.B.; Chen, S.C.; Liu, C.X.; Li, D.W. Comprehensive extraction and separation of cordycepin, polysaccharide and polypeptide from Cordyceps pupa residue. Shaanxi Agric. Sci. 2021, 67, 32–37. (In Chinese) [Google Scholar]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition in the innate immune response. Biochem. J. 2009, 420, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Sim, J.R.; Yun, C.H.; Han, S.H. Lipoteichoic acids as a major virulence factor causing inflammatory responses via Toll-like receptor 2. Arch. Pharmacal Res. 2016, 39, 1519–1529. [Google Scholar] [CrossRef]
- Monie, T. Section 4-integrated innate immunity—Combining activation and effector functions. In The Innate Immune System; Academic Press: Cambridge, MA, USA, 2017; pp. 121–169. [Google Scholar]
- Avbelj, M.; Hafner-Bratkovič, I.; Lainšček, D.; Manček-Keber, M.; Peternelj, T.T.; Panter, G.; Treon, S.P.; Gole, B.; Potočnik, U.; Jerala, R. Cleavage-mediated regulation of Myd88 signaling by inflammasome-activated caspase-1. Front. Immunol. 2022, 12, 790258. [Google Scholar] [CrossRef]
- Melehani, J.H. Mechanisms and Consequences of Staphylococcus aureus Leukocidin AB-Mediated Activation of the Host NLRP3 Inflammasome; The University of North Carolina at Chapel Hill: Chapel Hill, NC, USA, 2016. [Google Scholar]
- Yang, J.; Li, Y.Z.; Hylemon, P.B.; Zhang, L.Y.; Zhou, H.P. Cordycepin inhibits LPS-induced inflammatory responses by modulating NOD-Like Receptor Protein 3 inflammasome activation. Biomed. Pharmacother. 2017, 95, 1777–1788. [Google Scholar] [CrossRef]
- Peng, Z.P.; Zheneg, J.Y.; Guo, Z.T.; Wang, L.; Zhang, K.; Zhang, K.; Wang, G.B.; Qiu, Z.Y.; Wang, X.Z.; Li, J.X. Extraction and determination of cordycepin from culture residues of Cordyceps pupa by hydrothermal reflux. Chin. J. Anim. Husb. Vet. Med. 2022, 49, 3333–3342. (In Chinese) [Google Scholar]
- Uchiyama, R.; Toyoda, E.; Maehara, M.; Wasai, S.; Omura, H.; Watanabe, M.; Sato, M. Effect of Platelet-Rich Plasma on M1/M2 Macrophage Polarization. Int. J. Mol. Sci. 2021, 22, 2336. [Google Scholar] [CrossRef]
- Tu, Y.; Li, X.; Fu, Y.; Chen, Y.; Fang, H.; Li, Y.; Gu, Y.; Zhang, J. Isocorydine Ameliorates IL-6 Expression in Bone Marrow-Derived Macrophages and Acute Lung Injury Induced by Lipopolysaccharide. Int. J. Mol. Sci. 2023, 24, 4629. [Google Scholar] [CrossRef]
- Han, Y.; Yuan, C.; Zhou, X.; Han, Y.; He, Y.; Ouyang, J.; Zhou, W.; Wang, Z.; Wang, H.; Li, G. Anti-Inflammatory Activity of Three Triterpene from Hippophae rhamnoides L. in Lipopolysaccharide-Stimulated RAW264.7 Cells. Int. J. Mol. Sci. 2021, 22, 12009. [Google Scholar] [CrossRef] [PubMed]
- Belali, O.M.; Ahmed, M.M.; Mohany, M.; Belali, T.M.; Alotaibi, M.M.; Al-Hoshani, A.; Al-Rejaie, S.S. LCZ696 Protects against Diabetic Cardiomyopathy-Induced Myocardial Inflammation, ER Stress, and Apoptosis through Inhibiting AGEs/NF-κB and PERK/CHOP Signaling Pathways. Int. J. Mol. Sci. 2022, 23, 1288. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) |
|---|---|
| GAPDH | Forward: TCATGAAACAAATGCATCTGG |
| Reverse: TCATGAAACAAATGCATCTGG | |
| IL-1β | Forward: GCAGGCAGTATCACTCATTGT |
| Reverse: GGCTTTTTTGTTGTTCATCTC | |
| IL-6 | Forward: GTTCTCTGGGAAATCGTGGA |
| Reverse: GCATTGGAAATTGGGCTAGG | |
| IL-10 | Forward: ACCTGGTAGAAGTGATGCCCCAGGCA |
| Reverse: CTATGCAGTTGATGAAGATGTCAAA | |
| IL-18 | Forward: GCAGTAATACGGAGCATAAA |
| Reverse: ATCCTTCACAGATAGGGTCA | |
| TNF-α | Forward: AAGGGAGAGTGGTCAGGTTGG |
| Reverse: CAGAGGTTCAGTGATGTAGCG | |
| TLR2 | Forward: ACGTTGGATGCCAAGTGCTGGG |
| Reverse: ACGTTGGATCCACCCTGAA | |
| MyD88 | Forward: CGGAACTTTTCGATGCCTTTAT |
| Reverse: CACACACAACTTAAGCCGATAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Peng, Z.; Wang, L.; Zhang, J.; Zhang, K.; Guo, Z.; Xu, G.; Li, J. Cordyceps militaris Solid Medium Extract Alleviates Lipoteichoic Acid-Induced MH-S Inflammation by Inhibiting TLR2/NF-κB/NLRP3 Pathways. Int. J. Mol. Sci. 2023, 24, 15519. https://doi.org/10.3390/ijms242115519
Wang X, Peng Z, Wang L, Zhang J, Zhang K, Guo Z, Xu G, Li J. Cordyceps militaris Solid Medium Extract Alleviates Lipoteichoic Acid-Induced MH-S Inflammation by Inhibiting TLR2/NF-κB/NLRP3 Pathways. International Journal of Molecular Sciences. 2023; 24(21):15519. https://doi.org/10.3390/ijms242115519
Chicago/Turabian StyleWang, Xiaoya, Zhiping Peng, Lei Wang, Jingyan Zhang, Kang Zhang, Zhiting Guo, Guowei Xu, and Jianxi Li. 2023. "Cordyceps militaris Solid Medium Extract Alleviates Lipoteichoic Acid-Induced MH-S Inflammation by Inhibiting TLR2/NF-κB/NLRP3 Pathways" International Journal of Molecular Sciences 24, no. 21: 15519. https://doi.org/10.3390/ijms242115519
APA StyleWang, X., Peng, Z., Wang, L., Zhang, J., Zhang, K., Guo, Z., Xu, G., & Li, J. (2023). Cordyceps militaris Solid Medium Extract Alleviates Lipoteichoic Acid-Induced MH-S Inflammation by Inhibiting TLR2/NF-κB/NLRP3 Pathways. International Journal of Molecular Sciences, 24(21), 15519. https://doi.org/10.3390/ijms242115519