


Application of Nanotechnology in Plant Genetic Engineering

 ,
, 
Abstract
:1. Introduction
2. Conventional Genetic Transformation Methods
2.1. Gene Gun-Mediated Transformation
2.2. Agrobacterium-Mediated Transformation
2.3. Electroporation
2.4. PEG Delivery System
3. Nanoparticle-Mediated Biomolecule Delivery
3.1. Type of Nanoparticles
3.2. Methods for Nanomaterial Preparation
3.3. Transportation of NPs in Plants
4. Application
4.1. Application of Nanotechnology in Plant Genetic Transformation
4.2. Application of Nanotechnology in Gene Editing
5. Conclusions
6. Prospective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clapp, J.; Moseley, W.G. This food crisis is different: COVID-19 and the fragility of the neoliberal food security order. J. Peasant Stud. 2020, 47, 1393–1417. [Google Scholar] [CrossRef]
- Squire, H.J.; Tomatz, S.; Voke, E.; González-Grandío, E.; Landry, M. The emerging role of nanotechnology in plant genetic engineering. Nat. Rev. Bioeng. 2023, 1, 314–328. [Google Scholar] [CrossRef]
- Demirer, G.S.; Silva, T.N.; Jackson, C.T.; Thomas, J.B.; Ehrhardt, D.W.; Rhee, S.Y.; Mortimer, J.C.; Landry, M.P. Nanotechnology to advance CRISPR-Cas genetic engineering of plants. Nat. Nanotechnol. 2021, 16, 243–250. [Google Scholar] [CrossRef]
- Ogata, T.; Ishizaki, T.; Fujita, M.; Fujita, Y. CRISPR/Cas9-targeted mutagenesis of OsERA1 confers enhanced responses to abscisic acid and drought stress and increased primary root growth under nonstressed conditions in rice. PLoS ONE 2020, 15, e0243376. [Google Scholar] [CrossRef]
- Qiu, Z.; Kang, S.; He, L.; Zhao, J.; Zhang, S.; Hu, J.; Zeng, D.; Zhang, G.; Dong, G.; Gao, Z. The newly identified heat-stress sensitive albino 1 gene affects chloroplast development in rice. Plant Sci. 2018, 267, 168–179. [Google Scholar] [CrossRef]
- Cunningham, F.J.; Goh, N.S.; Demirer, G.S.; Matos, J.L.; Landry, M.P. Nanoparticle-mediated delivery towards advancing plant genetic engineering. Trends Biotechnol. 2018, 36, 882–897. [Google Scholar] [CrossRef]
- Wang, J.W.; Cunningham, F.J.; Goh, N.S.; Boozarpour, N.N.; Pham, M.; Landry, M.P. Nanoparticles for protein delivery in planta. Curr. Opin. Plant Biol. 2021, 60, 102052. [Google Scholar] [CrossRef]
- Wang, J.; Fu, A.; Xu, M.; Wang, Y. Research progress of gene gun bombardment in genetic transformation. Genom. Appl. Biol 2018, 37, 459–468. [Google Scholar]
- Lv, Z.; Jiang, R.; Chen, J.; Chen, W. Nanoparticle-mediated gene transformation strategies for plant genetic engineering. Plant J. 2020, 104, 880–891. [Google Scholar] [CrossRef]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Voke, E.; Pinals, R.L.; Goh, N.S.; Landry, M.P. In Planta Nanosensors: Understanding biocorona formation for functional design. ACS Sens. 2021, 6, 2802–2814. [Google Scholar] [CrossRef] [PubMed]
- Raj, S.N.; Anooj, E.; Rajendran, K.; Vallinayagam, S. A comprehensive review on regulatory invention of nano pesticides in agricultural nano formulation and food system. J. Mol. Struct. 2021, 1239, 130517. [Google Scholar]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munné-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Goh, N.S.; Wang, J.; Demirer, G.S.; Butrus, S.; Park, S.-J.; Landry, M.P. Nanoparticle cellular internalization is not required for RNA delivery to mature plant leaves. Nat. Nanotechnol. 2021, 17, 197–205. [Google Scholar] [CrossRef]
- Shou, H.; Frame, B.R.; Whitham, S.A.; Wang, K. Assessment of transgenic maize events produced by particle bombardment or Agrobacterium-mediated transformation. Mol. Breed. 2004, 13, 201–208. [Google Scholar] [CrossRef]
- Wang, J.W.; Grandio, E.G.; Newkirk, G.M.; Demirer, G.S.; Butrus, S.; Giraldo, J.P.; Landry, M.P. Nanoparticle-mediated genetic engineering of plants. Mol. Plant 2019, 12, 1037–1040. [Google Scholar] [CrossRef]
- Michielse, C.B.; Hooykaas, P.J.; van den Hondel, C.A.; Ram, A.F. Agrobacterium-mediated transformation as a tool for functional genomics in fungi. Curr. Genet. 2005, 48, 1–17. [Google Scholar] [CrossRef]
- Gustafson, V.; Mallubhotla, S.; MacDonnell, J.; Sanyal-Bagchi, M.; Chakravarty, B.; Wang-Pruski, G.; Rothwell, C.; Audy, P.; De Koeyer, D.; Siahbazi, M. Transformation and plant regeneration from leaf explants of Solanum tuberosum L. cv.‘Shepody’. Plant Cell Tiss. Org. 2006, 85, 361–366. [Google Scholar] [CrossRef]
- Kumar, S.; Raj, S.; Sharma, A.; Varma, H. Genetic transformation and development of Cucumber mosaic virus resistant transgenic plants of Chrysanthemum morifolium cv. Kundan. Sci. Hortic. 2012, 134, 40–45. [Google Scholar] [CrossRef]
- Du, N.; Pijut, P.M. Agrobacterium-mediated transformation of Fraxinus pennsylvanica hypocotyls and plant regeneration. Plant Cell Rep. 2009, 28, 915–923. [Google Scholar] [CrossRef]
- Song, J.; Lu, S.; Chen, Z.-Z.; Lourenco, R.; Chiang, V.L. Genetic transformation of Populus trichocarpa genotype Nisqually-1: A functional genomic tool for woody plants. Plant Cell Physiol. 2006, 47, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Visser, R.; Jacobsen, E.; Hesseling-Meinders, A.; Schans, M.; Witholt, B.; Feenstra, W. Transformation of homozygous diploid potato with an Agrobacterium tumefaciens binary vector system by adventitious shoot regeneration on leaf and stem segments. Plant Mol. Biol. 1989, 12, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Kiryushkin, A.S.; Ilina, E.L.; Guseva, E.D.; Pawlowski, K.; Demchenko, K.N. Hairy CRISPR: Genome editing in plants using hairy root transformation. Plants 2022, 11, 51. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Zhong, Y.; Huang, Z.; Yan, H.; Yuanda, L.; Jiang, B.; Zhong, G. Citrus cell suspension culture establishment, maintenance, efficient transformation and regeneration to complete transgenic plant. Plants 2021, 10, 664. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.-S.; Ge, X.-L.; Wang, R.; Yang, H.-F.; Bai, Y.-E.; Guo, Y.-H.; Zhang, J.; Lu, M.-Z.; Zhao, S.-T.; Wang, L.-Q. An efficient agrobacterium-mediated transformation method for hybrid poplar 84K (Populus alba × P. glandulosa) using calli as explants. Int. J. Mol. Sci. 2022, 23, 2216. [Google Scholar] [CrossRef]
- Zhou, G.-Y.; Weng, J.; Zeng, Y.; Huang, J.; Qian, S.; Liu, G. Introduction of exogenous DNA into cotton embryos. In Methods Enzymology; Academic Press: Cambridge, MA, USA, 1983; Volume 101, pp. 433–481. [Google Scholar]
- Ahmar, S.; Mahmood, T.; Fiaz, S.; Mora-Poblete, F.; Shafique, M.S.; Chattha, M.S.; Jung, K.H. Advantage of nanotechnology-based genome editing system and its application in crop improvement. Front. Plant Sci. 2021, 12, 663849. [Google Scholar] [CrossRef]
- Ali, A.; Bang, S.W.; Chung, S.-M.; Staub, J.E. Plant transformation via pollen tube-mediated gene transfer. Plant. Mol. Biol. Rep. 2014, 33, 742–747. [Google Scholar] [CrossRef]
- Maher, M.F.; Nasti, R.A.; Vollbrecht, M.; Starker, C.G.; Clark, M.D.; Voytas, D.F. Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 2020, 38, 84–89. [Google Scholar] [CrossRef]
- Zhang, H.; Demirer, G.S.; Zhang, H.; Ye, T.; Goh, N.S.; Aditham, A.J.; Cunningham, F.J.; Fan, C.; Landry, M.P. DNA nanostructures coordinate gene silencing in mature plants. Proc. Natl. Acad. Sci. USA 2019, 116, 7543–7548. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, H.; Demirer, G.S.; González-Grandío, E.; Fan, C.; Landry, M.P. Engineering DNA nanostructures for siRNA delivery in plants. Nat. Protoc. 2020, 15, 3064–3087. [Google Scholar] [CrossRef]
- Landry, M.P.; Mitter, N. How nanocarriers delivering cargos in plants can change the GMO landscape. Nat. Nanotechnol. 2019, 14, 512–514. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhu, X.; Yu, Y.; Li, C.; Zhang, Z.; Wang, F. Nanotechnology strategies for plant genetic engineering. Adv. Mater. 2022, 34, e2106945. [Google Scholar] [CrossRef] [PubMed]
- Kuzma, J.; Kokotovich, A. Renegotiating GM crop regulation: Targeted gene-modification technology raises new issues for the oversight of genetically modified crops. EMBO Rep. 2011, 12, 883–888. [Google Scholar] [CrossRef]
- Ichim, M.C. The more favorable attitude of the citizens toward GMOs supports a new regulatory framework in the European Union. GM Crops Food 2021, 12, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hu, H. High-efficiency nuclear transformation of the diatom Phaeodactylum tricornutum by electroporation. Mar. Genom. 2014, 16, 63–66. [Google Scholar] [CrossRef]
- Dong, Z.Z.; Chang, L.Q. Recent electroporation-based systems for intracellular molecule delivery. Nanotech. Precis. Eng. 2021, 4, 045001. [Google Scholar] [CrossRef]
- Ozyigit, I.I. Gene transfer to plants by electroporation: Methods and applications. Mol. Biol. Rep. 2020, 47, 3195–3210. [Google Scholar] [CrossRef]
- Batista Napotnik, T.; Miklavcic, D. In vitro electroporation detection methods—An overview. Bioelectrochemistry 2018, 120, 166–182. [Google Scholar] [CrossRef]
- Wu, S.; Zhu, H.; Liu, J.; Yang, Q.; Shao, X.; Bi, F.; Hu, C.; Huo, H.; Chen, K.; Yi, G. Establishment of a PEG-mediated protoplast transformation system based on DNA and CRISPR/Cas9 ribonucleoprotein complexes for banana. BMC Plant Biol. 2020, 20, 425. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, L.; Shen, M.; Liu, J.; Li, Y.; Xu, S.; Chen, L.; Shi, G.; Ding, Z. Establishment of an efficient Polyethylene Glycol (PEG)-mediated transformation system in Pleurotus eryngii var. ferulae using comprehensive optimization and multiple endogenous promoters. J. Fungi 2022, 8, 186. [Google Scholar] [CrossRef]
- Modena, M.M.; Rühle, B.; Burg, T.P.; Wuttke, S. Nanoparticle characterization: What to measure? Adv. Mater. 2019, 31, 1901556. [Google Scholar] [CrossRef]
- Gad, M.A.; Li, M.-J.; Ahmed, F.K.; Almoammar, H. Nanomaterials for gene delivery and editing in plants: Challenges and future perspective. In Multifunctional Hybrid Nanomaterials Sustainable Agri-Food Ecosystems; Elsevier: Amsterdam, The Netherlands, 2020; pp. 135–153. [Google Scholar]
- Guo, C.; Anwar, M.; Mei, R.; Li, X.; Zhao, D.; Jiang, Y.; Zhuang, J.; Liu, C.; Wang, C.; Hu, Z. Establishment and optimization of PEG-mediated protoplast transformation in the microalga Haematococcus pluvialis. J. Appl. Phycol. 2022, 34, 1595–1605. [Google Scholar] [CrossRef]
- Hubbard, J.D.; Lui, A.; Landry, M.P. Multiscale and multidisciplinary approach to understanding nanoparticle transport in plants. Curr. Opin. Chem. Eng. 2020, 30, 135–143. [Google Scholar] [CrossRef]
- Zhao, X.; Meng, Z.; Wang, Y.; Chen, W.; Sun, C.; Cui, B.; Cui, J.; Yu, M.; Zeng, Z.; Guo, S.; et al. Pollen magnetofection for genetic modification with magnetic nanoparticles as gene carriers. Nat. Plants 2017, 3, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.A.; Mohamed, A.E.-M.A.; Abd-Elsalam, K.A. Magnetic nanoparticles in plant protection: Promises and risks. Magn. Nanostructures Env. Agric. Appl. 2019, 2019, 225–246. [Google Scholar]
- Wang, Z.P.; Zhang, Z.B.; Zheng, D.Y.; Zhang, T.T.; Li, X.L.; Zhang, C.; Yu, R.; Wei, J.H.; Wu, Z.Y. Efficient and genotype independent maize transformation using pollen transfected by DNA-coated magnetic nanoparticles. J. Integr. Plant Biol. 2022, 64, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Demirer, G.S.; Zhang, H.; Goh, N.S.; Gonzalez-Grandio, E.; Landry, M.P. Carbon nanotube-mediated DNA delivery without transgene integration in intact plants. Nat. Protoc. 2019, 14, 2954–2971. [Google Scholar] [CrossRef] [PubMed]
- Demirer, G.S.; Zhang, H.; Matos, J.L.; Goh, N.; Cunningham, F.; Sung, Y.; Chang, R.; Aditham, A.J.; Chio, L.; Cho, M.-J.; et al. High aspect ratio nanomaterials enable delivery of functional genetic material without DNA integration in mature plants. Nat. Nanotechnol. 2018, 14, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.M.; Belisle, M.; Frommer, W.B. The evolving landscape around genome editing in agriculture: Many countries have exempted or move to exempt forms of genome editing from GMO regulation of crop plants. EMBO Rep. 2020, 21, e50680. [Google Scholar] [CrossRef]
- Baig, N.; Kammakakam, I.; Falath, W. Nanomaterials: A review of synthesis methods, properties, recent progress, and challenges. Mater. Adv. 2021, 2, 1821–1871. [Google Scholar] [CrossRef]
- Banerjee, A.; Sarkar, A.; Acharya, K.; Chakraborty, N. Nanotechnology: An emerging hope in crop improvement. Lett. Appl. NanoBioScience 2021, 10, 2784–2803. [Google Scholar] [CrossRef]
- Marchiol, L. Synthesis of metal nanoparticles in living plants. Ital. J. Agron. 2012, 7, e37. [Google Scholar] [CrossRef]
- Khan, F.; Shariq, M.; Asif, M.; Siddiqui, M.A.; Malan, P.; Ahmad, F. Green nanotechnology: Plant-mediated nanoparticle synthesis and application. Nanomatererials 2022, 12, 673. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Bulut, O.; Some, S.; Mandal, A.K.; Yilmaz, M.D. Green synthesis of silver nanoparticles: Biomolecule-nanoparticle organizations targeting antimicrobial activity. RSC Adv. 2019, 9, 2673–2702. [Google Scholar] [CrossRef]
- Hu, P.; An, J.; Faulkner, M.M.; Wu, H.; Li, Z.; Tian, X.; Giraldo, J.P. Nanoparticle charge and size control foliar delivery efficiency to plant cells and organelles. ACS Nano 2020, 14, 7970–7986. [Google Scholar] [CrossRef]
- Avellan, A.; Yun, J.; Zhang, Y.; Spielman-Sun, E.; Unrine, J.M.; Thieme, J.; Li, J.; Lombi, E.; Bland, G.; Lowry, G.V. Nanoparticle size and coating chemistry control foliar uptake pathways, translocation, and leaf-to-rhizosphere transport in wheat. ACS Nano 2019, 13, 5291–5305. [Google Scholar] [CrossRef]
- Sanzari, I.; Leone, A.; Ambrosone, A. Nanotechnology in plant science: To make a long story short. Front. Bioeng. Biotech. 2019, 7, 120. [Google Scholar] [CrossRef]
- Wang, X.; Xie, H.; Wang, P.; Yin, H. Nanoparticles in plants: Uptake, transport and physiological activity in leaf and root. Materials 2023, 16, 3097. [Google Scholar] [CrossRef]
- Jassby, D.; Su, Y.; Adeleye, A.S.; Kim, C.; Ashworth, V.; Rolshausen, P.; Roper, C.; White, J.C. Delivery, uptake, fate, and transport of engineered nanoparticles in plants: A critical review and data analysis. Env. Sci. Nano 2019, 6, 2311–2331. [Google Scholar]
- Ali, S.; Mehmood, A.; Khan, N.; Tan, B. Uptake, translocation, and consequences of nanomaterials on plant growth and stress adaptation. J. Nanomater. 2021, 2021, 6677616. [Google Scholar] [CrossRef]
- Zhu, J.; Li, J.; Shen, Y.; Liu, S.; Zeng, N.; Zhan, X.; White, J.C.; Gardea-Torresdey, J.; Xing, B. Mechanism of zinc oxide nanoparticle entry into wheat seedling leaves. Env. Sci. Nano 2020, 7, 3901–3913. [Google Scholar] [CrossRef]
- Avellan, A.; Yun, J.; Morais, B.P.; Clement, E.T.; Rodrigues, S.M.; Lowry, G.V. Critical review: Role of inorganic nanoparticle properties on their foliar uptake and in planta translocation. Env. Sci. Technol. 2021, 55, 13417–13431. [Google Scholar] [CrossRef]
- Lew, T.T.S.; Wong, M.H.; Kwak, S.Y.; Sinclair, R.; Koman, V.B.; Strano, M.S. Rational design principles for the transport and subcellular distribution of nanomaterials into plant protoplasts. Small 2018, 14, e1802086. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.J.; Wang, H.; Yan, B.; Zheng, H.; Jiang, Y.; Miranda, O.R.; Rotello, V.M.; Xing, B.; Vachet, R.W. Effect of surface charge on the uptake and distribution of gold nanoparticles in four plant species. Env. Sci. Technol. 2012, 46, 12391–12398. [Google Scholar] [CrossRef]
- Khan, M.R.; Adam, V.; Rizvi, T.F.; Zhang, B.; Ahamad, F.; Josko, I.; Zhu, Y.; Yang, M.; Mao, C. Nanoparticle-plant interactions: Two-way traffic. Small 2019, 15, e1901794. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.T.E.; Choi, C.H.J. Enabling transgenic plant cell–derived biomedicines with nanotechnology. Adv. NanoBiomed. Res. 2020, 1, 2000028. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, J.; Zhan, X.; Li, A.; White, J.C.; Gardea-Torresdey, J.L.; Xing, B. Role of charge and size in the translocation and distribution of zinc oxide particles in wheat cells. ACS Sustain. Chem. Eng. 2021, 9, 11556–11564. [Google Scholar] [CrossRef]
- Zhang, P.; Guo, Z.; Zhang, Z.; Fu, H.; White, J.C.; Lynch, I. Nanomaterial transformation in the soil-plant system: Implications for food safety and application in agriculture. Small 2020, 16, e2000705. [Google Scholar] [CrossRef]
- Demirer, G.S.; Zhang, H.; Goh, N.S.; Chang, R.; Landry, M.P. Carbon nanocarriers deliver siRNA to intact plant cells for efficient gene knockdown. Sci. Adv. 2019, 6, eaaz0495. [Google Scholar] [CrossRef]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 16207. [Google Scholar] [CrossRef]
- Ahmed, S.; Gao, X.; Jahan, M.A.; Adams, M.; Wu, N.; Kovinich, N. Nanoparticle-based genetic transformation of Cannabis sativa. J. Biotechnol. 2021, 326, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Martin-Ortigosa, S.; Peterson, D.J.; Valenstein, J.S.; Lin, V.S.-Y.; Trewyn, B.G.; Lyznik, L.A.; Wang, K. Mesoporous silica nanoparticle-mediated intracellular Cre protein delivery for maize genome editing via loxP site excision. Plant Physiol. 2014, 164, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Torney, F.; Trewyn, B.G.; Lin, V.S.-Y.; Wang, K. Mesoporous silica nanoparticles deliver DNA and chemicals into plants. Nat. Nanotechnol. 2007, 2, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.; Maitra, A.; Abdin, M.; Akmal, M.; Arora, I.; Samim, M. Calcium phosphate nanoparticle mediated genetic transformation in plants. J. Mater. Chem. 2012, 22, 3500–3507. [Google Scholar] [CrossRef]
- Burlaka, O.M.; Pirko, Y.V.; Yemets, A.I.; Blume, Y.B. Application of carbon nanotubes for plant genetic transformation. In Proceedings of the Nanocomposites, Nanophotonics, Nanobiotechnology, and Applications: Selected Proceedings of the Second FP7 Conference and International Summer School Nanotechnology: From Fundamental Research to Innovations, Bukovel, Ukraine, 25 August–1 September 2013; Springer: Berlin/Heidelberg, Germany, 2015; pp. 233–255. [Google Scholar]
- Chang, F.P.; Kuang, L.Y.; Huang, C.A.; Jane, W.N.; Hung, Y.; Yue-ie, C.H.; Mou, C.Y. A simple plant gene delivery system using mesoporous silica nanoparticles as carriers. J. Mater. Chem. B 2013, 1, 5279–5287. [Google Scholar] [CrossRef]
- Hajiahmadi, Z.; Shirzadian-Khorramabad, R.; Kazemzad, M.; Sohani, M.M. Enhancement of tomato resistance to Tuta absoluta using a new efficient mesoporous silica nanoparticle-mediated plant transient gene expression approach. Sci. Hortic. 2019, 243, 367–375. [Google Scholar] [CrossRef]
- Mosa, K.A.; El-Naggar, M.; Ramamoorthy, K.; Alawadhi, H.; Elnaggar, A.; Wartanian, S.; Ibrahim, E.; Hani, H. Copper nanoparticles induced genotoxicty, oxidative stress, and changes in superoxide dismutase (SOD) gene expression in cucumber (Cucumis sativus) plants. Front. Plant Sci. 2018, 9, 872. [Google Scholar] [CrossRef]
- Martin-Ortigosa, S.; Valenstein, J.S.; Lin, V.S.Y.; Trewyn, B.G.; Wang, K. Gold functionalized mesoporous silica nanoparticle mediated protein and DNA codelivery to plant cells via the biolistic method. Adv. Funct. Mater. 2012, 22, 3576–3582. [Google Scholar] [CrossRef]
- Hao, Y.; Yang, X.; Shi, Y.; Song, S.; Xing, J.; Marowitch, J.; Chen, J.; Chen, J. Magnetic gold nanoparticles as a vehicle for fluorescein isothiocyanate and DNA delivery into plant cells. Botany 2013, 91, 457–466. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, S.P.; Zhu, Z.H.; Liu, J.S. Experimental evaluation on high temperature rheological properties of various fiber modified asphalt binders. J. Cent. South Univ. Technol. 2008, 15, 135–139. [Google Scholar] [CrossRef]
- Pasupathy, K.; Lin, S.; Hu, Q.; Luo, H.; Ke, P.C. Direct plant gene delivery with a poly (amidoamine) dendrimer. Biotechnol. J. Healthc. Nutr. Technol. 2008, 3, 1078–1082. [Google Scholar] [CrossRef] [PubMed]
- Golestanipour, A.; Nikkhah, M.; Aalami, A.; Hosseinkhani, S. Gene delivery to tobacco root cells with single-walled carbon nanotubes and cell-penetrating fusogenic peptides. Mol. Biotechnol. 2018, 60, 863–878. [Google Scholar] [CrossRef] [PubMed]
- Finiuk, N.; Buziashvili, A.; Burlaka, O.; Zaichenko, A.; Mitina, N.; Miagkota, O.; Lobachevska, O.; Stoika, R.; Blume, Y.; Yemets, A. Investigation of novel oligoelectrolyte polymer carriers for their capacity of DNA delivery into plant cells. Plant Cell Tiss. Org. 2017, 131, 27–39. [Google Scholar] [CrossRef]
- Kwak, S.-Y.; Lew, T.T.S.; Sweeney, C.J.; Koman, V.B.; Wong, M.H.; Bohmert-Tatarev, K.; Snell, K.D.; Seo, J.S.; Chua, N.-H.; Strano, M.S. Chloroplast-selective gene delivery and expression in planta using chitosan-complexed single-walled carbon nanotube carriers. Nat. Nanotechnol. 2019, 14, 447–455. [Google Scholar] [CrossRef]
- Newkirk, G.M.; de Allende, P.; Jinkerson, R.E.; Giraldo, J.P. Nanotechnology approaches for chloroplast biotechnology advancements. Front. Plant Sci. 2021, 12, 691295. [Google Scholar] [CrossRef] [PubMed]
- Jat, S.K.; Bhattacharya, J.; Sharma, M.K. Nanomaterial based gene delivery: A promising method for plant genome engineering. J. Mater. Chem. B 2020, 8, 4165–4175. [Google Scholar] [CrossRef]
- Grohmann, L.; Keilwagen, J.; Duensing, N.; Dagand, E.; Hartung, F.; Wilhelm, R.; Bendiek, J.; Sprink, T. Detection and identification of genome editing in plants: Challenges and opportunities. Front. Plant Sci. 2019, 10, 236. [Google Scholar] [CrossRef]
- Tröder, S.E.; Zevnik, B. History of genome editing: From meganucleases to CRISPR. Lab. Anim. 2021, 56, 60–68. [Google Scholar] [CrossRef]
- Baltes, N.J.; Voytas, D.F. Enabling plant synthetic biology through genome engineering. Trends Biotechnol. 2015, 33, 120–131. [Google Scholar] [CrossRef]
- Becker, S.; Boch, J. TALE and TALEN genome editing technologies. Gene Genome Ed. 2021, 2, 100007. [Google Scholar] [CrossRef]
- Mao, Y.; Botella, J.R.; Liu, Y.; Zhu, J.-K. Gene editing in plants: Progress and challenges. Natl. Sci. Rev. 2019, 6, 421–437. [Google Scholar] [CrossRef]
- Van Eck, J. Applying gene editing to tailor precise genetic modifications in plants. J. Biol. Chem. 2020, 295, 13267–13276. [Google Scholar] [CrossRef] [PubMed]
- Nandy, D.; Maity, A.; Mitra, A.K. Target-specific gene delivery in plant systems and their expression: Insights into recent developments. J. Biosci. 2020, 45, 30. [Google Scholar] [CrossRef]
- Sukegawa, S.; Saika, H.; Toki, S. Plant genome editing: Ever more precise and wide reaching. Plant. J. 2021, 106, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Cheng, Q.; Min, Y.L.; Olson, E.N.; Siegwart, D.J. Systemic nanoparticle delivery of CRISPR-Cas9 ribonucleoproteins for effective tissue specific genome editing. Nat. Commun. 2020, 11, 3232. [Google Scholar] [CrossRef]
- Duan, L.; Ouyang, K.; Xu, X.; Xu, L.; Wen, C.; Zhou, X.; Qin, Z.; Xu, Z.; Sun, W.; Liang, Y. Nanoparticle delivery of CRISPR/Cas9 for genome editing. Front. Genet. 2021, 12, 673286. [Google Scholar] [CrossRef]
- Mout, R.; Ray, M.; Yesilbag Tonga, G.; Lee, Y.W.; Tay, T.; Sasaki, K.; Rotello, V.M. Direct cytosolic delivery of CRISPR/Cas9-Ribonucleoprotein for efficient gene editing. ACS Nano 2017, 11, 2452–2458. [Google Scholar] [CrossRef]
- McCarty, N.S.; Graham, A.E.; Studená, L.; Ledesma-Amaro, R. Multiplexed CRISPR technologies for gene editing and transcriptional regulation. Nat. Commun. 2020, 11, 1281. [Google Scholar] [CrossRef]
- Izuegbunam, C.L.; Wijewantha, N.; Wone, B.; Ariyarathne, M.A.; Sereda, G.; Wone, B.W.M. A nano-biomimetic transformation system enables in planta expression of a reporter gene in mature plants and seeds. Nanoscale Adv. 2021, 3, 3240–3250. [Google Scholar] [CrossRef]
- Rao, X.; Zhao, H.; Shao, C.; Yi, C. Characterizing off-target effects of genome editors. Curr. Opin. Biomed. Eng. 2023, 28, 100480. [Google Scholar] [CrossRef]
- Yuan, G.; Martin, S.; Hassan, M.M.; Tuskan, G.A.; Yang, X. PARA: A new platform for the rapid assembly of gRNA arrays for multiplexed CRISPR technologies. Cells 2022, 11, 2467. [Google Scholar] [CrossRef] [PubMed]
- Libbrecht, M.W.; Noble, W.S. Machine learning applications in genetics and genomics. Nat. Rev. Genet. 2015, 16, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Emmert-Streib, F.; Yang, Z.; Feng, H.; Tripathi, S.; Dehmer, M. An introductory review of deep learning for prediction models with big data. Front. Artif. Intell. 2020, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Bertho, L.; Schmidt, K.; Schmidtke, J.; Brants, I.; Cantón, R.F.; Novillo, C.; Head, G. Results from ten years of post-market environmental monitoring of genetically modified MON 810 maize in the European Union. PLoS ONE 2020, 15, e0217272. [Google Scholar] [CrossRef]
- Mishra, S.; Sundaram, B. Fate, transport, and toxicity of nanoparticles: An emerging pollutant on biotic factors. Process Saf. Env. 2023, 174, 595–607. [Google Scholar] [CrossRef]
- Crossa, J.; Jarquín, D.; Franco, J.; Pérez-Rodríguez, P.; Burgueño, J.; Saint-Pierre, C.; Vikram, P.; Sansaloni, C.; Petroli, C.; Akdemir, D.; et al. Genomic prediction of gene bank wheat landraces. G3 Genes Genomes Genet. 2016, 6, 1819–1834. [Google Scholar] [CrossRef]
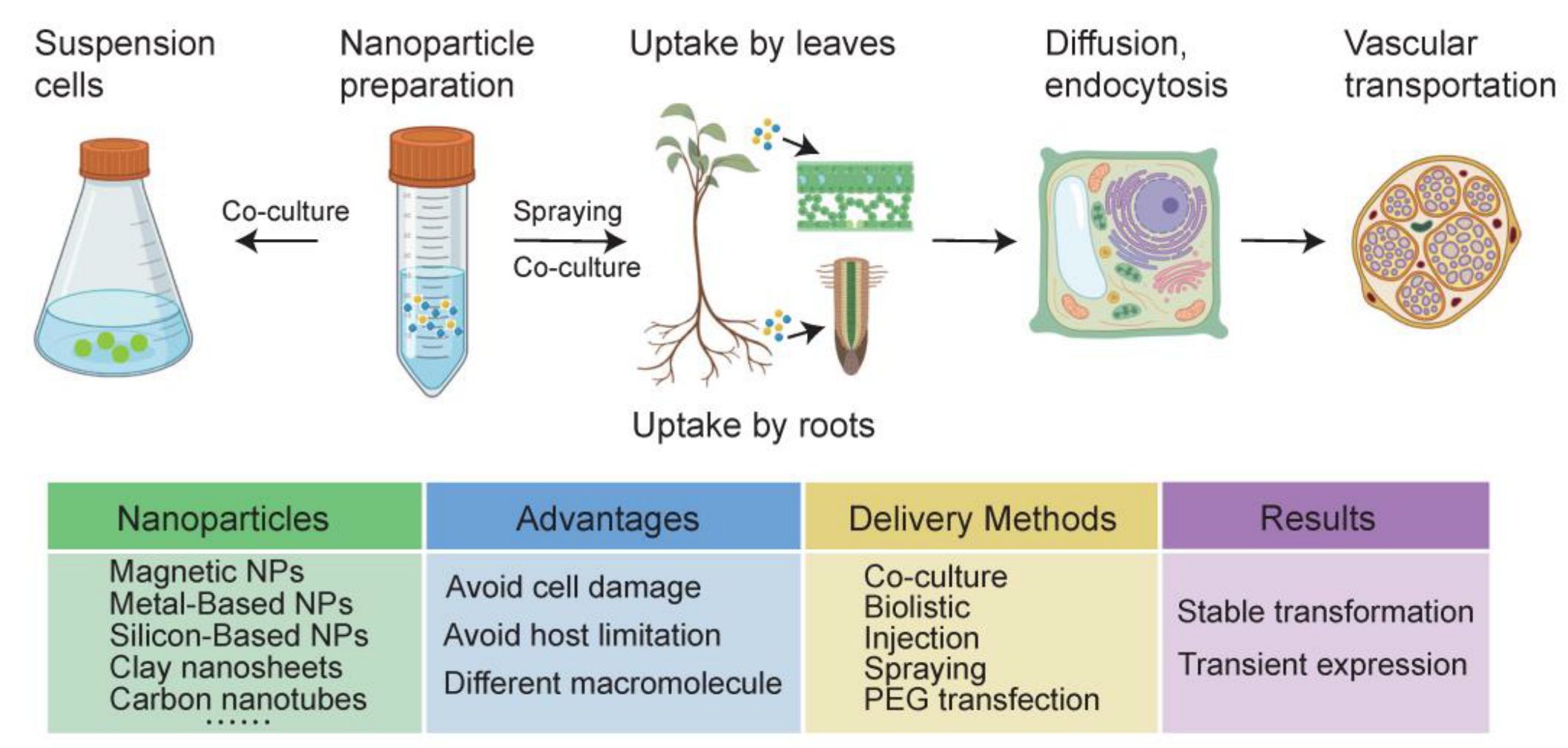
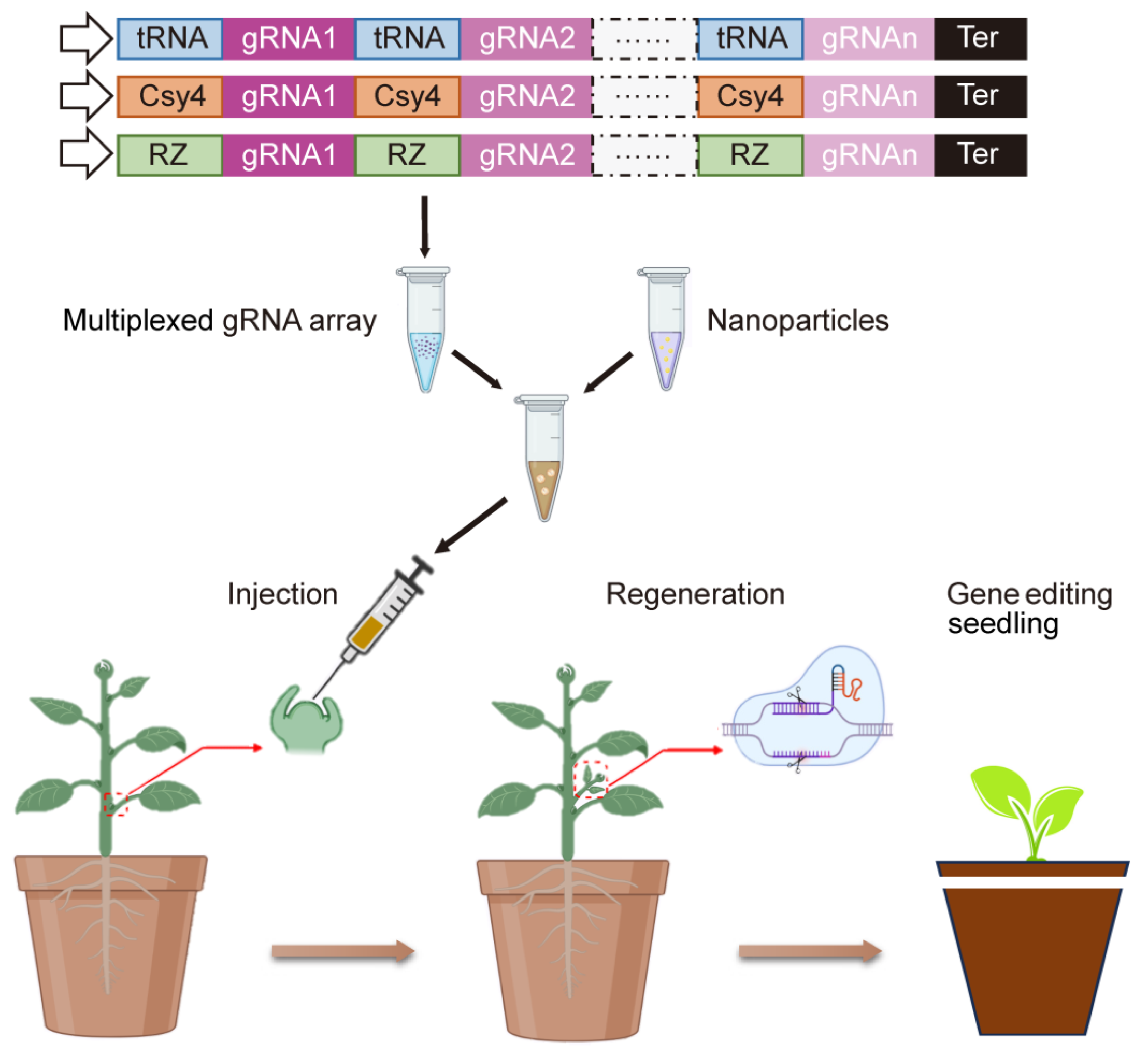

{kind=link}
{kind=link}
| Target Plant Species and Tissues | Nanoparticle Type | Cargo | Delivery Method | Stable Transformation/ Transient Expression | References |
|---|---|---|---|---|---|
| N. benthamiana (leaves) | Layered dihydroxide clay nanosheets | Double-stranded RNA | Spraying | Transient expression | [72] |
| N. benthamiana | Carbon nanotube | GFP plasmid | Injection | Transient expression | [49] |
| E. sativa | Carbon nanotube | ||||
| T. aestivum | Carbon nanotube | ||||
| C. sativa | PEI-Au@SiO2 | Transcription factors | Infiltration | Transient expression | [73] |
| Z. mays (calli) | Cationic fluorescence nanoparticle | Protein | Biolistic | Stable transformation | [74] |
| N. tabacum (cotyledons) | Gold capped MSNs | GFP plasmid; chemical expression inducer | Biolistic | Transient expression | [75] |
| B. juncea (hypocotyl explants) | Calcium phosphate NPs (CaPNPs) | β-glucuronidase (GUS) plasmid | Passive diffusion | Stable transformation | [76] |
| N. tabacum (protoplasts and leaf explants) | Organically functionalized CNTs | YFP plasmid | Co-culture | Stable transformation | [77] |
| A. thaliana (roots) | Organically functionalized MSNs | mCherry plasmid | Passive diffusion | Transient expression | [78] |
| E. sativa, N. benthamiana, T. aestivum and G. hirsutum (leaves) | Polymer-functionalized CNTs | GFP plasmid; siRNA for transgenic silencing | Infiltration | Transient expression | [50] |
| S. lycopersicum (leaves) | Mesoporous silica nanoparticles (MSNs) | β-glucuronidase (GUS) plasmid | Spraying and injection | Transient expression | [79] |
| C. sativus (root) | Copper nanoparticles (CuNP) | - | Co-culture | - | [80] |
| A. cepa (epidermis cells) | Gold-plated MSNs | GFP and mCherry plasmids; GFP protein | Biolistic | Transient expression | [81] |
| B. napus L. var. Jet Neuf (protoplasts and walled cell suspension) | Magnetic gold NPs | FITC molecules and β-glucuronidase (GUS) plasmid | External magnetic field | Transient expression | [82] |
| D. zingiberensis (calli suspension) | Poly-L-lysine-coated starch NPs | GFP plasmid | Sonoporation | Transient expression | [83] |
| A. stolonifera L. (calli suspension) | Polyamidoamine (PAMAM) dendrimer NPs | GFP plasmid | Co-culture | Transient expression | [84] |
| N. tabacum var. Virginia (root cells) | Single-walled carbon nanotube | GFP plasmid | Co-culture | Transient expression | [85] |
| N. tabacum and C. purpureus (protoplasts) | Dimethylaminoethyl methacrylate (DMAEM) polymer NPs | YFP and GFP plasmids | PEG transfection | Stable transformation and transient expression | [86] |
| G. hirsutum (pollen) | Magnetic Fe3O4 NPs | Selectable marker gene plasmids | External magnetic field | Stable transformation | [46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, K.; Xu, C.; Li, T.; Ma, H.; Gong, J.; Li, X.; Sun, X.; Hu, X. Application of Nanotechnology in Plant Genetic Engineering. Int. J. Mol. Sci. 2023, 24, 14836. https://doi.org/10.3390/ijms241914836
Wu K, Xu C, Li T, Ma H, Gong J, Li X, Sun X, Hu X. Application of Nanotechnology in Plant Genetic Engineering. International Journal of Molecular Sciences. 2023; 24(19):14836. https://doi.org/10.3390/ijms241914836
Chicago/Turabian StyleWu, Kexin, Changbin Xu, Tong Li, Haijie Ma, Jinli Gong, Xiaolong Li, Xuepeng Sun, and Xiaoli Hu. 2023. "Application of Nanotechnology in Plant Genetic Engineering" International Journal of Molecular Sciences 24, no. 19: 14836. https://doi.org/10.3390/ijms241914836
APA StyleWu, K., Xu, C., Li, T., Ma, H., Gong, J., Li, X., Sun, X., & Hu, X. (2023). Application of Nanotechnology in Plant Genetic Engineering. International Journal of Molecular Sciences, 24(19), 14836. https://doi.org/10.3390/ijms241914836