
Mitoquinone Alleviates Donation after Cardiac Death Kidney Injury during Hypothermic Machine Perfusion in Rat Model

 , ,
, ,
Abstract
:1. Introduction
2. Results
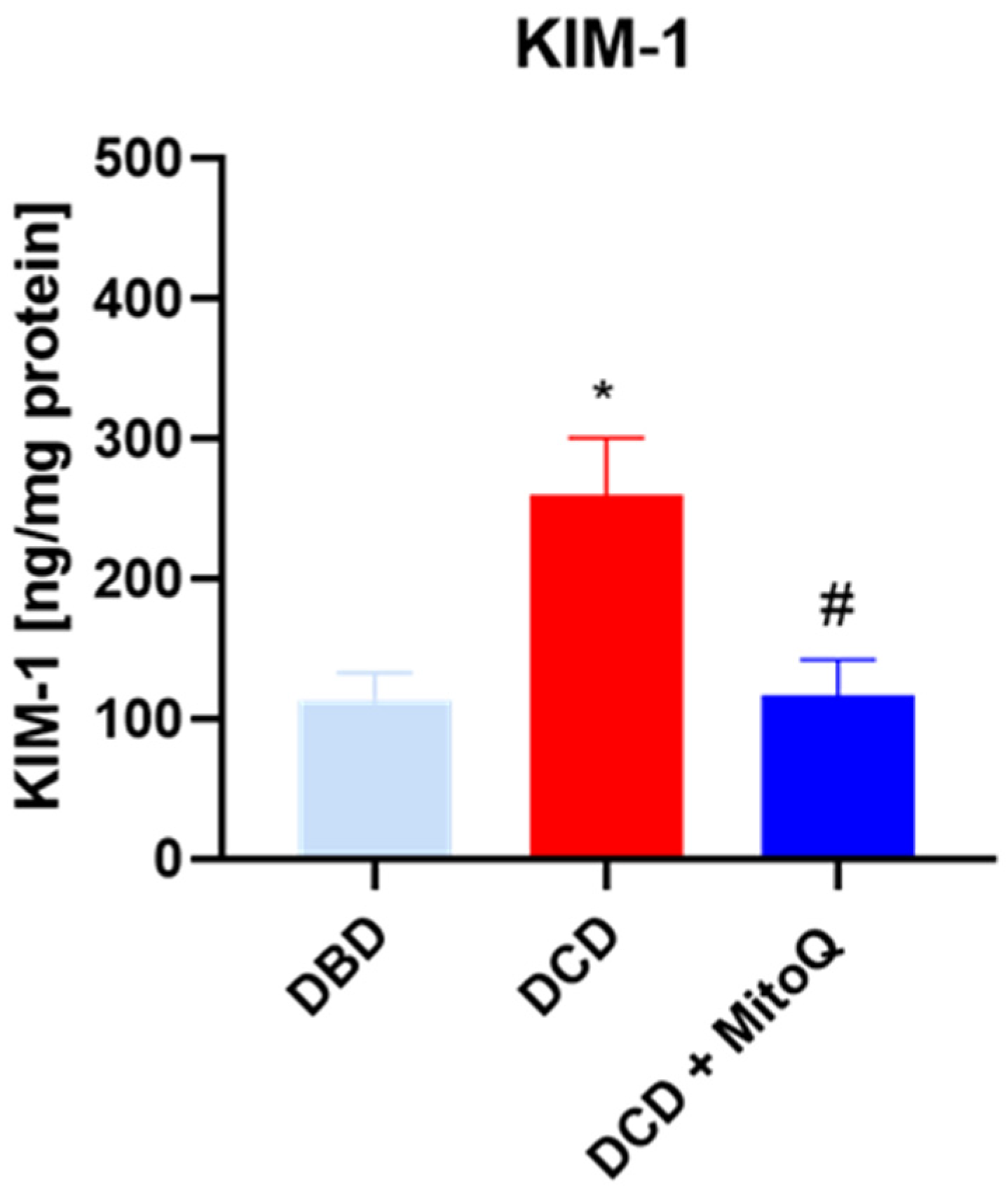
2.1. The Influence of MitoQ Administration on the Level of Kidney Injury Markers
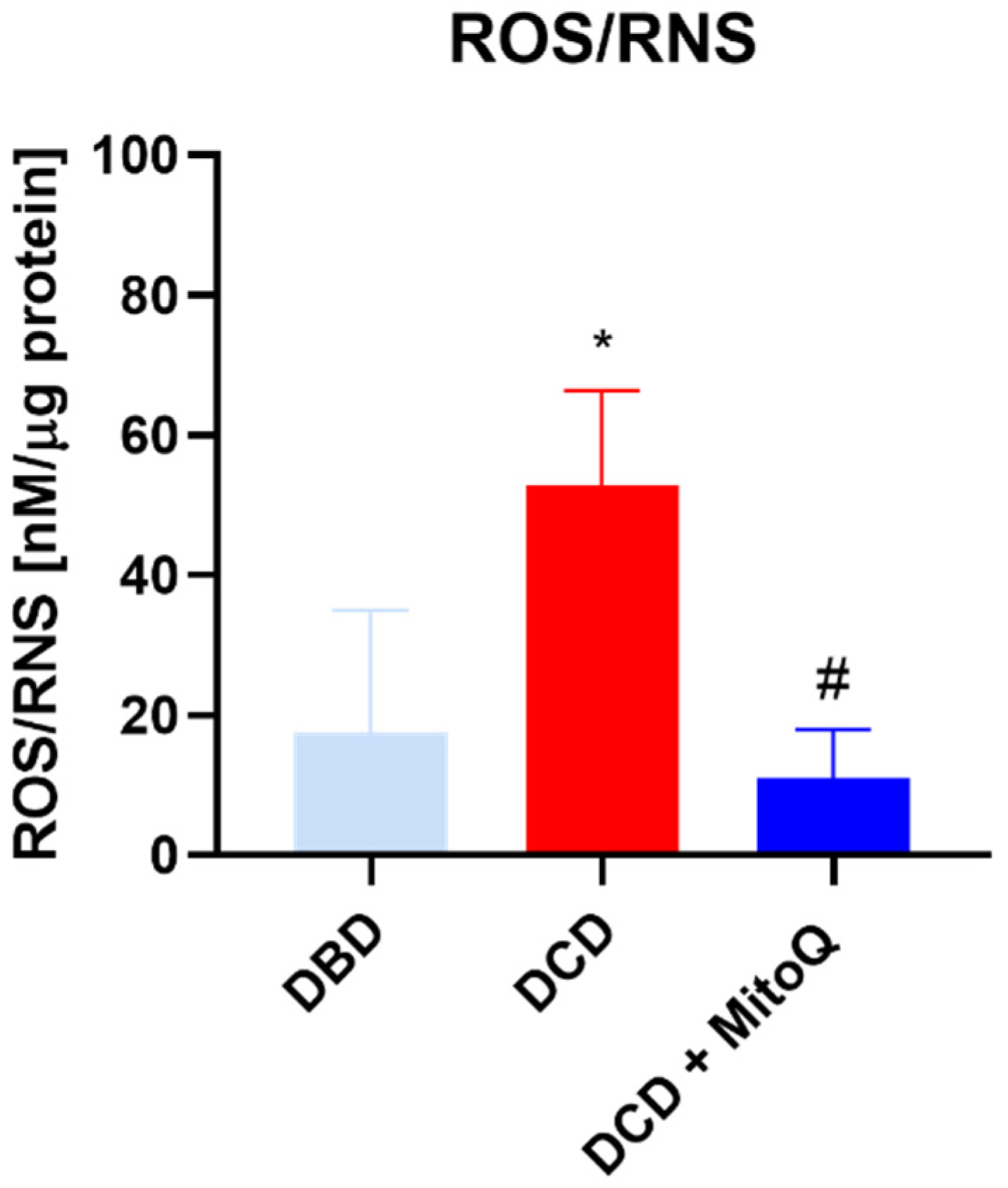
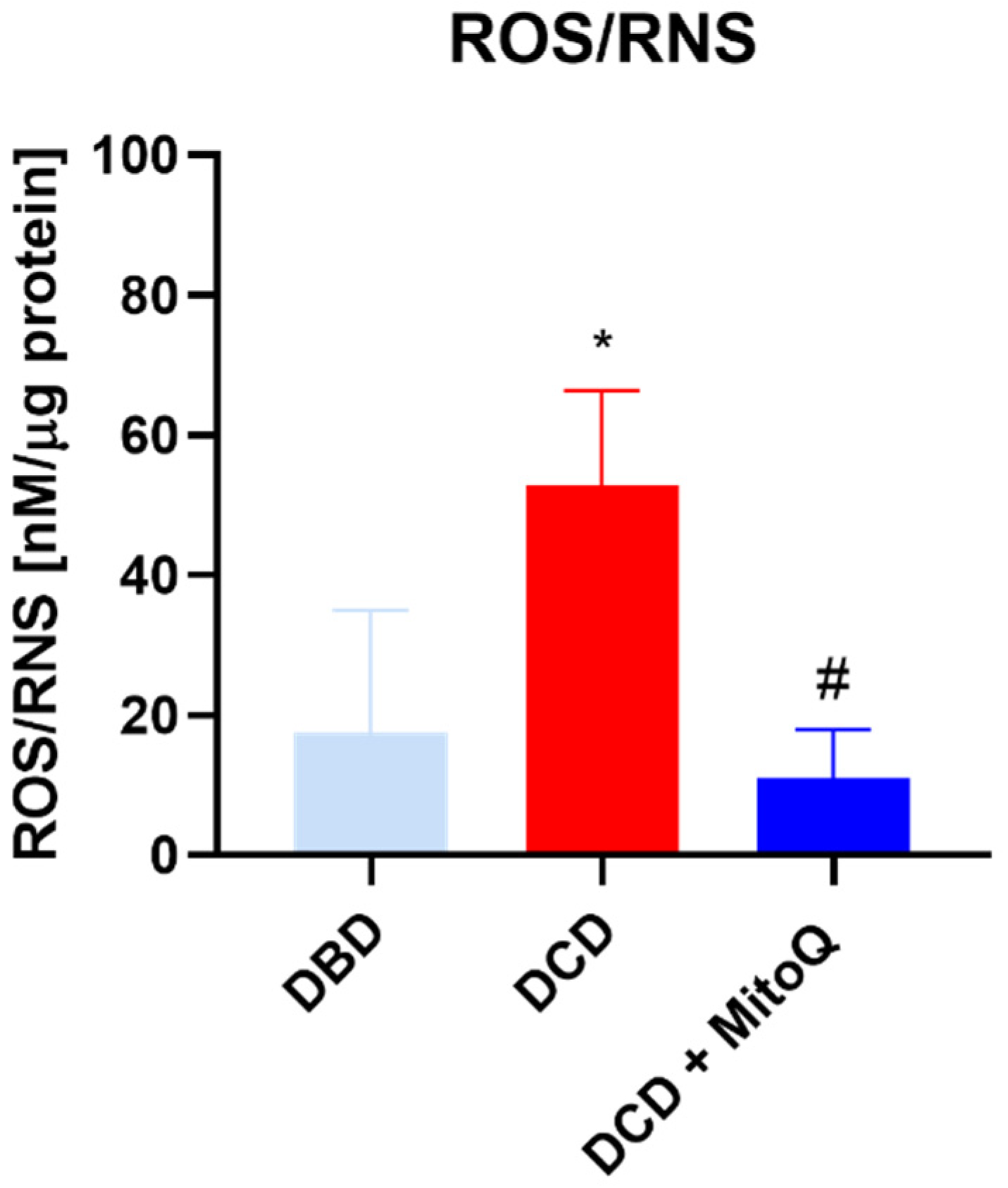
2.2. Oxidative Status of Perfused Kidneys
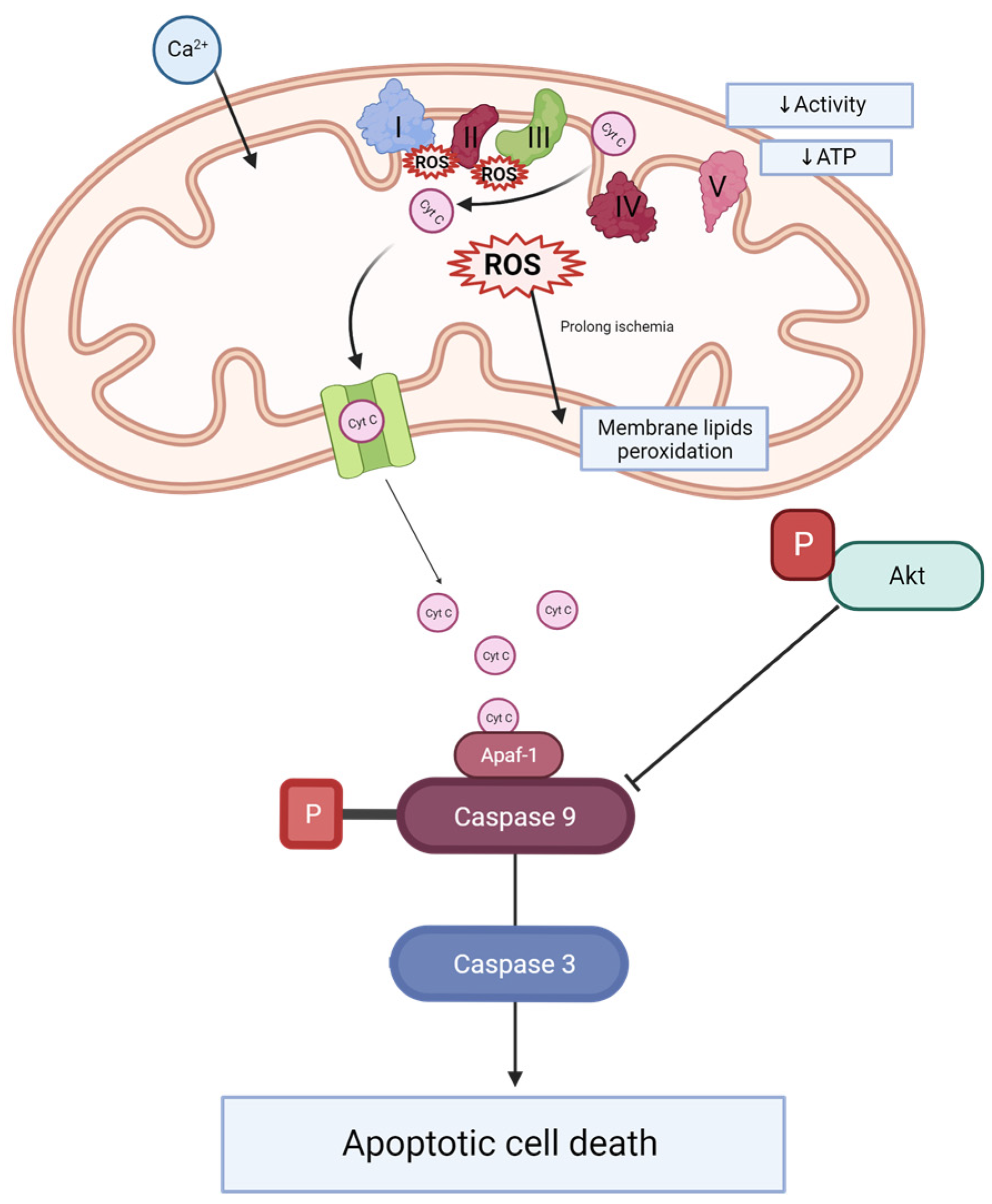
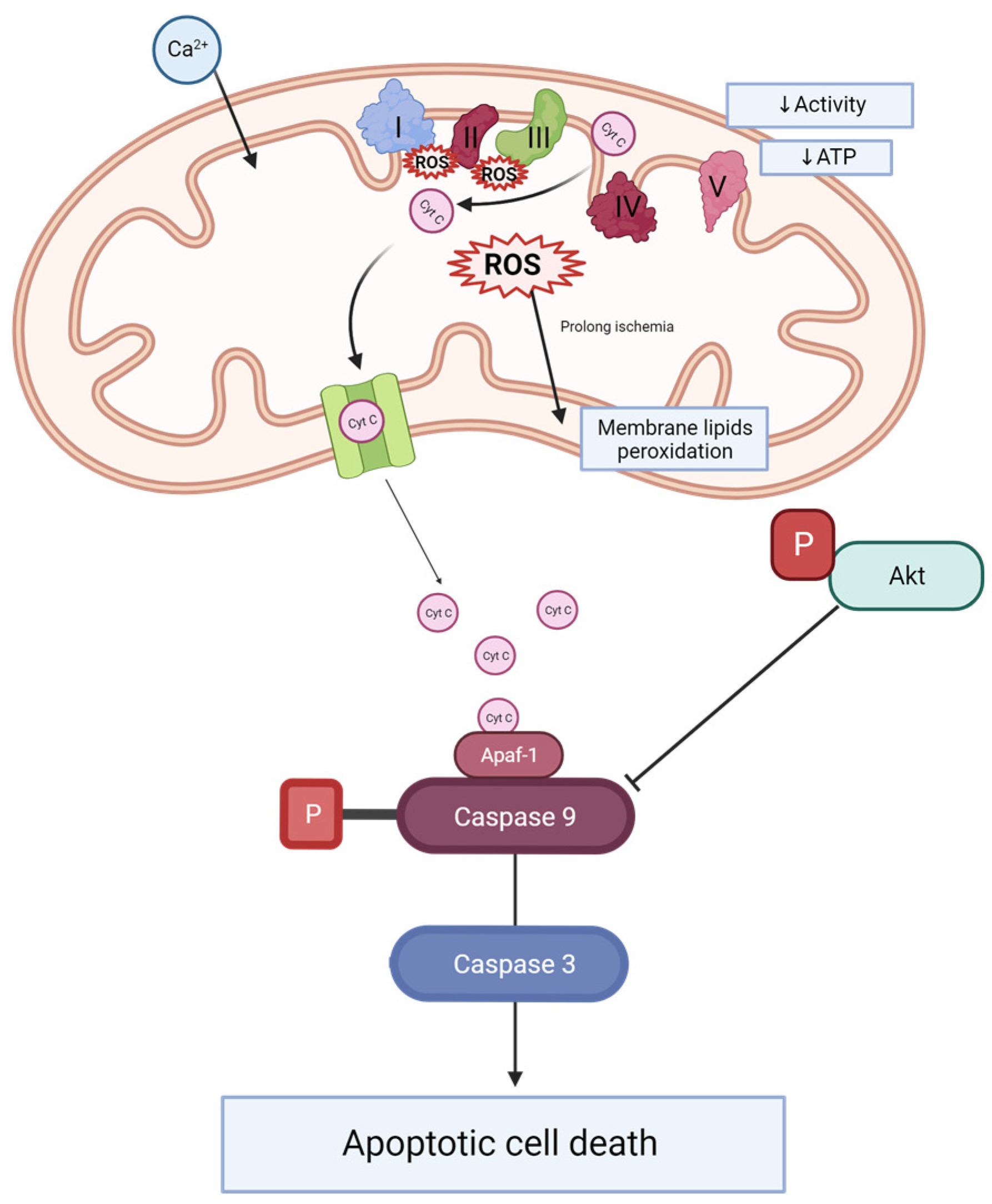
2.3. Caspase 3 and Caspase 9 Concentrations Are Reduced after MitoQ Administration
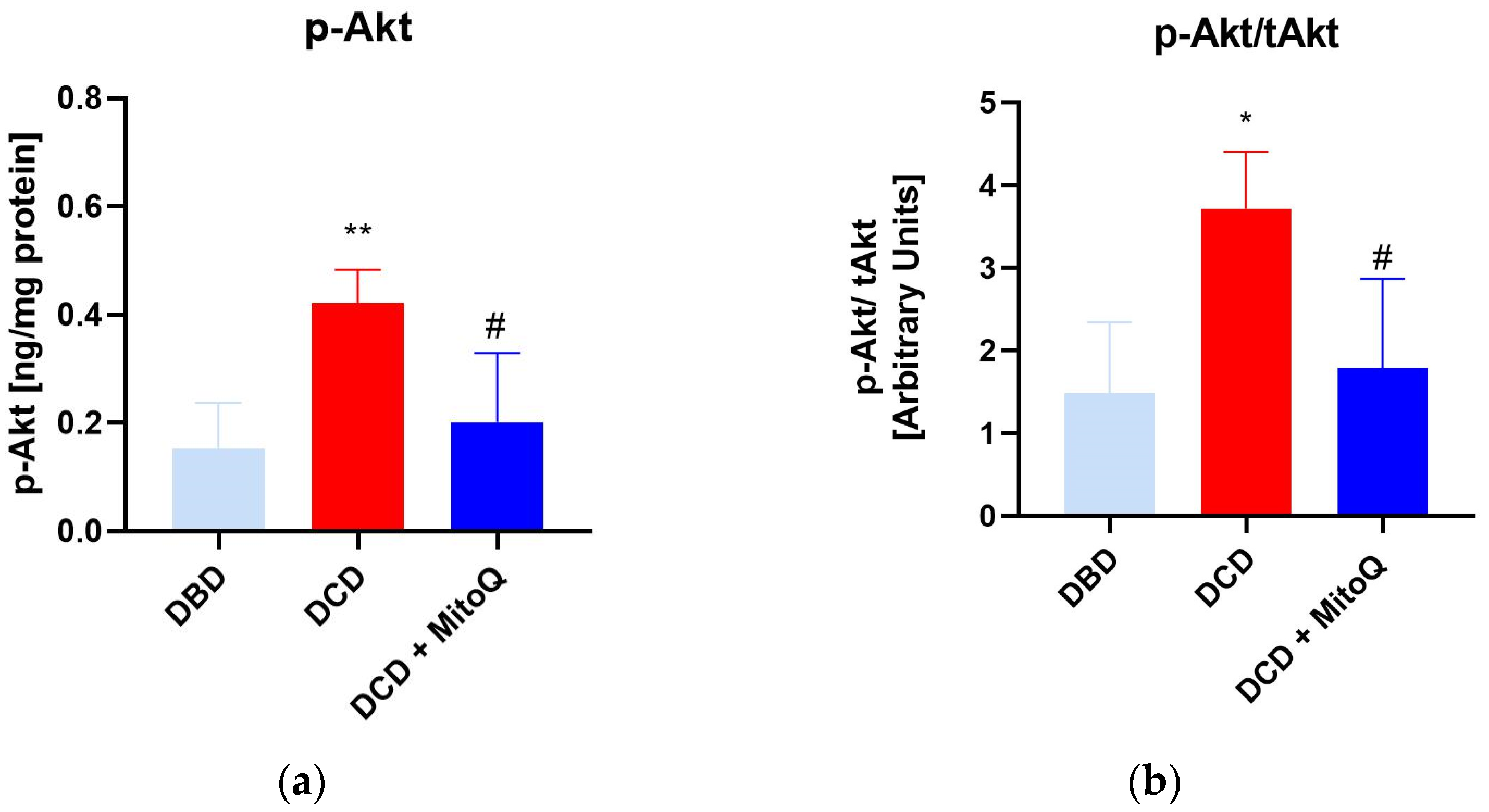
2.4. MitoQ Mitigate the Ratio of Phosphorylated Akt and Total Akt in Kidney Tissue
2.5. MitoQ Mitigate the Ratio of Phosphorylated Akt and Total Akt in Kidney Tissue
3. Discussion
4. Material and Methods
4.1. Reagents
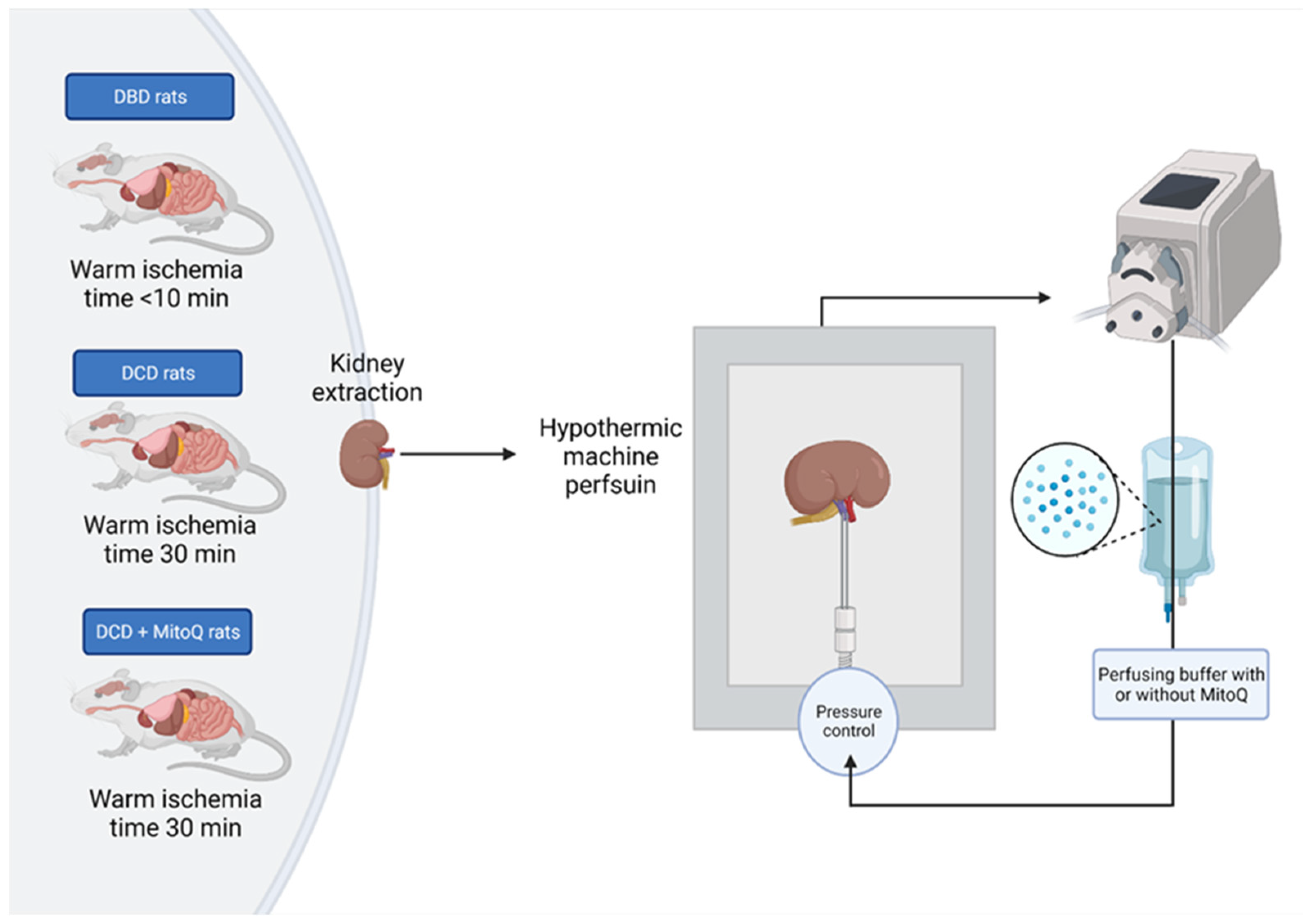
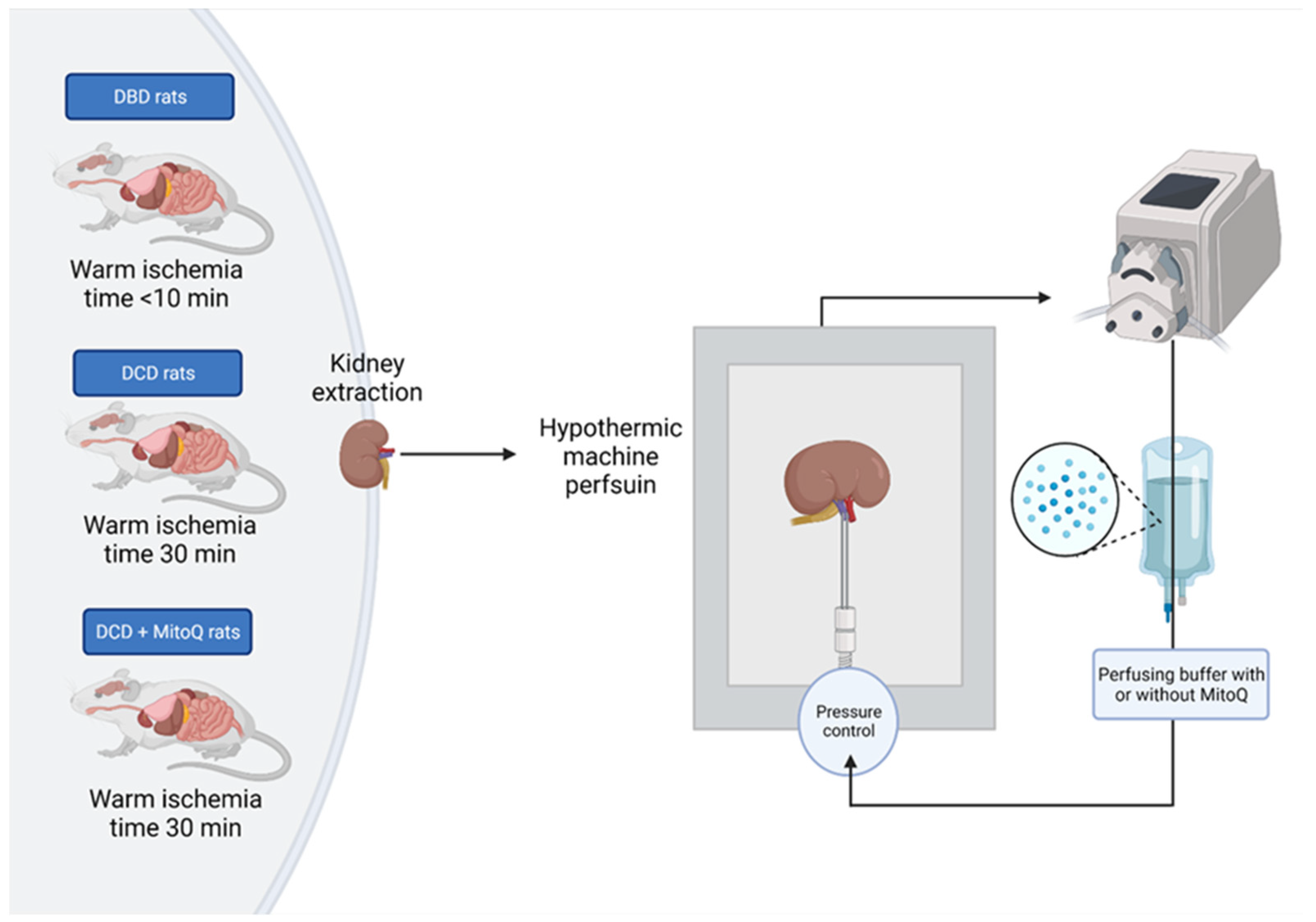
4.2. Experimental Animals
4.3. Hypothermic Machine Perfusion of Rat Kidney
4.4. Preparation of Kidney Homogenates
4.5. Assessment of Kidney Injury Marker (KIM-1)
4.6. Detection of Total Protein Concentration
4.7. Assessments of ROS/RNS in the Kidney Tissue
4.8. Mitochondria Isolation
4.9. Measurements of Cytochrome C Oxidase Activity and Outer Mitochondrial Membrane Integrity
4.10. Caspase 3 and Caspase 9 Concentration Measurement
4.11. Total Akt and Phosphorylated Akt Concentration
4.12. Statistical Analysis
5. Study Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tonelli, M.; Wiebe, N.; Knoll, G.; Bello, A.; Browne, S.; Jadhav, D.; Klarenbach, S.; Gill, J. Systematic Review: Kidney Transplantation Compared With Dialysis in Clinically Relevant Outcomes. Am. J. Transplant. 2011, 11, 2093–2109. [Google Scholar] [CrossRef] [PubMed]
- van Heurn, L.E.; Talbot, D.; Nicholson, M.L.; Akhtar, M.Z.; Sanchez-Fructuoso, A.I.; Weekers, L.; Barrou, B. Recommendations for donation after circulatory death kidney transplantation in Europe. Transpl. Int. 2016, 29, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, R.P.; Shapter, O.; Aitken, E.; Stevenson, K.; Shiels, P.G.; Kingsmore, D.B. Has the Expansion in Extended Criteria Deceased Donors Led to a Different Type of Delayed Graft Function and Poorer Outcomes? Transplant. Proc. 2018, 50, 3160–3164. [Google Scholar] [CrossRef]
- Krzywonos-Zawadzka, A.; Franczak, A.; Moser, M.A.J.; Olejnik, A.; Sawicki, G.; Bil-Lula, I. Pharmacological Protection of Kidney Grafts from Cold Perfusion-Induced Injury. Biomed. Res. Int. 2019, 2019, 9617087. [Google Scholar] [CrossRef]
- Mezzolla, V.; Pontrelli, P.; Fiorentino, M.; Stasi, A.; Pesce, F.; Franzin, R.; Rascio, F.; Grandaliano, G.; Stallone, G.; Infante, B.; et al. Emerging biomarkers of delayed graft function in kidney transplantation. Transplant. Rev. 2021, 35, 100629. [Google Scholar] [CrossRef]
- Radajewska, A.; Krzywonos-Zawadzka, A.; Bil-Lula, I. Recent Methods of Kidney Storage and Therapeutic Possibilities of Transplant Kidney. Biomedicines 2022, 10, 1013. [Google Scholar] [CrossRef]
- Radajewska, A.; Szyller, J.; Niewiadomska, J.; Noszczyk-Nowak, A.; Bil-Lula, I. Punica granatum L. Polyphenolic Extract as an Antioxidant to Prevent Kidney Injury in Metabolic Syndrome Rats. Oxid. Med. Cell. Longev. 2023, 2023, 6144967. [Google Scholar] [CrossRef]
- Daenen, K.; Andries, A.; Mekahli, D.; Van Schepdael, A.; Jouret, F.; Bammens, B. Oxidative stress in chronic kidney disease. Pediatr. Nephrol. 2019, 34, 975–991. [Google Scholar] [CrossRef]
- Liakopoulos, V.; Roumeliotis, S.; Gorny, X.; Eleftheriadis, T.; Mertens, P.R. Oxidative Stress in Patients Undergoing Peritoneal Dialysis: A Current Review of the Literature. Oxid. Med. Cell. Longev. 2017, 2017, 3494867. [Google Scholar] [CrossRef]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The Role of Oxidative Stress in Physiopathology and Pharmacological Treatment with Pro- and Antioxidant Properties in Chronic Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 2082145. [Google Scholar] [CrossRef]
- Fonseca, I.; Reguengo, H.; Almeida, M.; Dias, L.; Martins, L.S.; Pedroso, S.; Santos, J.; Lobato, L.; Henriques, A.C.; Mendonça, D. Oxidative stress in kidney transplantation: Malondialdehyde is an early predictive marker of graft dysfunction. Transplantation 2014, 97, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Li, J.; Yu, Y.; Wei, Q.; Deng, W.; Yu, L. l-carnitine attenuates oxidant injury in HK-2 cells via ROS-mitochondria pathway. Regul. Pept. 2010, 161, 58–66. [Google Scholar] [CrossRef]
- La Manna, G.; Lanci, N.; Della Bella, E.; Comai, G.; Cappuccilli, M.L.; Nisi, K.; Todeschini, P.; Carretta, E.; Scolari, M.P.; Stefoni, S. Reduction of oxidative damage reflects a better kidney transplantation outcome. Am. J. Nephrol. 2011, 34, 496–504. [Google Scholar] [CrossRef]
- James, A.M.; Sharpley, M.S.; Manas, A.R.B.; Frerman, F.E.; Hirst, J.; Smith, R.A.; Murphy, M.P. Interaction of the Mitochondria-targeted Antioxidant MitoQ with Phospholipid Bilayers and Ubiquinone Oxidoreductases. J. Biol. Chem. 2007, 282, 14708–14718. [Google Scholar] [CrossRef] [PubMed]
- Antipodean Pharmaceuticals, Inc. A Double-Blind, Parallel, Randomized Comparison of Two Doses of MitoQ and Placebo for the Treatment of Patients with Raised Liver Enzymes Due to Hepatitis C. 2008. Available online: https://clinicaltrials.gov/study/NCT00433108 (accessed on 5 September 2023).
- A Study to Compare MitoQ and Placebo to Treat Non-alcoholic Fatty Liver Disease (NAFLD)—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01167088 (accessed on 12 September 2023).
- Hostrup, M. Mitochondrial Dysfunction and Insulin Resistance in Skeletal Muscle. 2021. Available online: https://clinicaltrials.gov/study/NCT04558190 (accessed on 5 September 2023).
- Imperial College London. Examining the Effects of Mitochondrial Oxidative Stress in DCM. 2022. Available online: https://clinicaltrials.gov/study/NCT05410873 (accessed on 5 September 2023).
- University of Colorado, Denver. Mitochondrial Targeted Antioxidant Supplementation and Diastolic Function in Healthy Men and Women. 2022. Available online: https://clinicaltrials.gov/study/NCT03586414 (accessed on 11 September 2023).
- Dare, A.J.; Bolton, E.A.; Pettigrew, G.J.; Bradley, J.A.; Saeb-Parsy, K.; Murphy, M.P. Protection against renal ischemia–reperfusion injury in vivo by the mitochondria targeted antioxidant MitoQ. Redox Biol. 2015, 5, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Allan, L.A.; Clarke, P.R. Apoptosis and autophagy: Regulation of caspase-9 by phosphorylation. FEBS J. 2009, 276, 6063–6073. [Google Scholar] [CrossRef] [PubMed]
- Hamed, M.; Logan, A.; Gruszczyk, A.V.; Beach, T.E.; James, A.M.; Dare, A.J.; Barlow, A.; Martin, J.; Georgakopoulos, N.; Gane, A.M.; et al. Mitochondria-targeted antioxidant MitoQ ameliorates ischaemia–reperfusion injury in kidney transplantation models. Br. J. Surg. 2021, 108, 1072–1081. [Google Scholar] [CrossRef]
- Gane, E.J.; Weilert, F.; Orr, D.W.; Keogh, G.F.; Gibson, M.; Lockhart, M.M.; Frampton, C.M.; Taylor, K.M.; Smith, R.A.; Murphy, M.P. The mitochondria-targeted anti-oxidant mitoquinone decreases liver damage in a phase II study of hepatitis C patients. Liver Int. 2010, 30, 1019–1026. [Google Scholar] [CrossRef]
- Hall, A.M.; Schuh, C.D. Mitochondria as therapeutic targets in acute kidney injury. Curr. Opin. Nephrol. Hypertens. 2016, 25, 355–362. [Google Scholar] [CrossRef]
- Qiao, X.; Chen, X.; Wu, D.; Ding, R.; Wang, J.; Hong, Q.; Shi, S.; Li, J.; Xie, Y.; Lu, Y.; et al. Mitochondrial pathway is responsible for aging-related increase of tubular cell apoptosis in renal ischemia/reperfusion injury. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 830–839. [Google Scholar] [CrossRef]
- Mitchell, T.; Rotaru, D.; Saba, H.; Smith, R.A.J.; Murphy, M.P.; MacMillan-Crow, L.A. The Mitochondria-Targeted Antioxidant Mitoquinone Protects against Cold Storage Injury of Renal Tubular Cells and Rat Kidneys. J. Pharmacol. Exp. Ther. 2011, 336, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.A.; Arcand, S.; Lin, H.B.; Wojnarowicz, C.; Sawicka, J.; Banerjee, T.; Luo, Y.; Beck, G.R.; Luke, P.P.; Sawicki, G. Protection of the Transplant Kidney from Preservation Injury by Inhibition of Matrix Metalloproteinases. PLoS ONE 2016, 11, e0157508. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Youle, R.J. The Role of Mitochondria in Apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef]
- Ojano-Dirain, C.P.; Antonelli, P.J. Prevention of gentamicin-induced apoptosis with the mitochondria-targeted antioxidant mitoquinone. Laryngoscope 2012, 122, 2543–2548. [Google Scholar] [CrossRef]
- Turkseven, S.; Bolognesi, M.; Brocca, A.; Pesce, P.; Angeli, P.; Di Pascoli, M. Mitochondria-targeted antioxidant mitoquinone attenuates liver inflammation and fibrosis in cirrhotic rats. Am. J. Physiol.-Gastrointest. Liver Physiol. 2020, 318, G298–G304. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Xiang, T.; Qiu, Q.; Leung, J.; Xu, J.; Zhou, W.; Hu, Q.; Lan, J.; Liu, Z.; Zhong, Z.; et al. Aldehyde dehydrogenase 2 regulates autophagy via the Akt-mTOR pathway to mitigate renal ischemia-reperfusion injury in hypothermic machine perfusion. Life Sci. 2020, 253, 117705. [Google Scholar] [CrossRef]
- Du, X.; Zhang, P.; Fu, H.; Ahsan, H.M.; Gao, J.; Chen, Q. Smart mitochondrial-targeted cancer therapy: Subcellular distribution, selective TrxR2 inhibition accompany with declined antioxidant capacity. Int. J. Pharm. 2019, 555, 346–355. [Google Scholar] [CrossRef]
- Franke, T.F.; Hornik, C.P.; Segev, L.; Shostak, G.A.; Sugimoto, C. PI3K/Akt and apoptosis: Size matters. Oncogene 2003, 22, 8983–8998. [Google Scholar] [CrossRef]
- Fathy, M.; Abdel-Latif, R.; Abdelgwad, Y.M.; Othman, O.A.; Abdel-Razik, A.R.H.; Dandekar, T.; Othman, E.M. Nephroprotective potential of eugenol in a rat experimental model of chronic kidney injury; targeting NOX, TGF-β, and Akt signaling. Life Sci. 2022, 308, 120957. [Google Scholar] [CrossRef]
- Lan, A.; Du, J. Potential role of Akt signaling in chronic kidney disease. Nephrol. Dial. Transplant. 2015, 30, 385–394. [Google Scholar] [CrossRef]
- Böttinger, E.P. TGF-β in Renal Injury and Disease. Semin. Nephrol. 2007, 27, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Ghosh Choudhury, G.; Abboud, H.E. Tyrosine phosphorylation-dependent PI 3 kinase/Akt signal transduction regulates TGFβ-induced fibronectin expression in mesangial cells. Cell. Signal. 2004, 16, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Conduit, S.E.; Davies, E.M.; Ooms, L.M.; Gurung, R.; McGrath, M.J.; Hakim, S.; Cottle, D.L.; Smyth, I.M.; Dyson, J.M.; Mitchell, C.A. AKT signaling promotes DNA damage accumulation and proliferation in polycystic kidney disease. Hum. Mol. Genet. 2020, 29, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Duan, Z.; Yu, T.; Xu, J.; Liu, L. Inhibiting Cytochrome C Oxidase Leads to Alleviated Ischemia Reperfusion Injury. Korean Circ. J. 2017, 47, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Hüttemann, M.; Helling, S.; Sanderson, T.H.; Sinkler, C.; Samavati, L.; Mahapatra, G.; Varughese, A.; Lu, G.; Liu, J.; Ramzan, R.; et al. Regulation of mitochondrial respiration and apoptosis through cell signaling: Cytochrome c oxidase and cytochrome c in ischemia/reperfusion injury and inflammation. Biochim. Biophys. Acta 2012, 1817, 598–609. [Google Scholar] [CrossRef]
- Sanderson, T.H.; Wider, J.M.; Lee, I.; Reynolds, C.A.; Liu, J.; Lepore, B.; Tousignant, R.; Bukowski, M.J.; Johnston, H.; Fite, A.; et al. Inhibitory modulation of cytochrome c oxidase activity with specific near-infrared light wavelengths attenuates brain ischemia/reperfusion injury. Sci. Rep. 2018, 8, 3481. [Google Scholar] [CrossRef]
- See, Y.P.; Weisel, R.D.; Mickle, D.A.; Teoh, K.H.; Wilson, G.J.; Tumiati, L.C.; Mohabeer, M.K.; Madonik, M.M.; Axford-Gatley, R.A.; Salter, D.R. Prolonged hypothermic cardiac storage for transplantation. The effects on myocardial metabolism and mitochondrial function. J. Thorac. Cardiovasc. Surg. 1992, 104, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, T.; Bonventre, J.V.; Bailly, V.; Wei, H.; Hession, C.A.; Cate, R.L.; Sanicola, M. Kidney injury molecule-1 (KIM-1), a putative epithelial cell adhesion molecule containing a novel immunoglobulin domain, is up-regulated in renal cells after injury. J. Biol. Chem. 1998, 273, 4135–4142. [Google Scholar] [CrossRef]
- Thompson, D.A.; Ferguson-Miller, S. Lipid and subunit III depleted cytochrome c oxidase purified by horse cytochrome c affinity chromatography in lauryl maltoside. Biochemistry 1983, 22, 3178–3187. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Product | Source | Identifier |
|---|---|---|
| Aprotinin | Roche (Basel, Switzerland) | 10236624001 |
| Bovine Serum Albumin | Sigma (Saint Louis, MO, USA) | A3294-100G |
| Buprenorphine | Orion Pharma, 0.3 mg/ml (Warsaw, Poland) | N/A |
| CaCl2 | Sigma (Saint Louis, MO, USA) | C1016 |
| Dithiothreitol | Roche (Basel, Switzerland) | 10708984001 |
| DMSO | Origen (Austin, Texas) | CP-50 |
| EDTA | Chempur (Piekary Śląskie, Poland) | 118798103 |
| Glucose | Chempur (Piekary Śląskie, Poland) | 114595600 |
| KCl | Chempur (Piekary Śląskie, Poland) | 117397402 |
| KH2PO4 | Sigma (Saint Louis, MO, USA) | P0662 |
| Leupeptin | Sigma (Saint Louis, MO, USA) | L8511 |
| MgSO4 | Sigma (Saint Louis, MO, USA) | M7506 |
| Mitoquinone | Abcam (Cambridge, United Kingdom) | ab285406 |
| NaCl | POCH (Gliwice, Poland) | BA4121116 |
| NaHCO3 | Chempur (Piekary Śląskie, Poland) | 428105306 |
| DPBS | GibcoTM (Thermo Fisher Scientific, Inc., Waltham, MA, USA) | A2644601 |
| Sodium pentobarbital | Biowet, (133,3 mg + 26,7 mg)/ml (Puławy, Poland) | N/A |
| Soybean trypsin inhibitor | Sigma (Saint Louis, MO, USA) | P8340 |
| Sucrose | Sigma (Saint Louis, MO, USA) | S0389 |
| Tris-HCl | Roche (Basel, Switzerland) | 10812846001 |
| Triton X-100 | Sigma (Saint Louis, MO, USA) | X100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radajewska, A.; Szyller, J.; Krzywonos-Zawadzka, A.; Olejnik, A.; Sawicki, G.; Bil-Lula, I. Mitoquinone Alleviates Donation after Cardiac Death Kidney Injury during Hypothermic Machine Perfusion in Rat Model. Int. J. Mol. Sci. 2023, 24, 14772. https://doi.org/10.3390/ijms241914772
Radajewska A, Szyller J, Krzywonos-Zawadzka A, Olejnik A, Sawicki G, Bil-Lula I. Mitoquinone Alleviates Donation after Cardiac Death Kidney Injury during Hypothermic Machine Perfusion in Rat Model. International Journal of Molecular Sciences. 2023; 24(19):14772. https://doi.org/10.3390/ijms241914772
Chicago/Turabian StyleRadajewska, Anna, Jakub Szyller, Anna Krzywonos-Zawadzka, Agnieszka Olejnik, Grzegorz Sawicki, and Iwona Bil-Lula. 2023. "Mitoquinone Alleviates Donation after Cardiac Death Kidney Injury during Hypothermic Machine Perfusion in Rat Model" International Journal of Molecular Sciences 24, no. 19: 14772. https://doi.org/10.3390/ijms241914772
APA StyleRadajewska, A., Szyller, J., Krzywonos-Zawadzka, A., Olejnik, A., Sawicki, G., & Bil-Lula, I. (2023). Mitoquinone Alleviates Donation after Cardiac Death Kidney Injury during Hypothermic Machine Perfusion in Rat Model. International Journal of Molecular Sciences, 24(19), 14772. https://doi.org/10.3390/ijms241914772