
Broadening the Substrate Specificity of Cellobiose Phosphorylase from Clostridium thermocellum for Improved Transformation of Cellodextrin to Starch


Abstract
:1. Introduction
2. Results
2.1. Modification of the Catalytic Loop of CtCBP
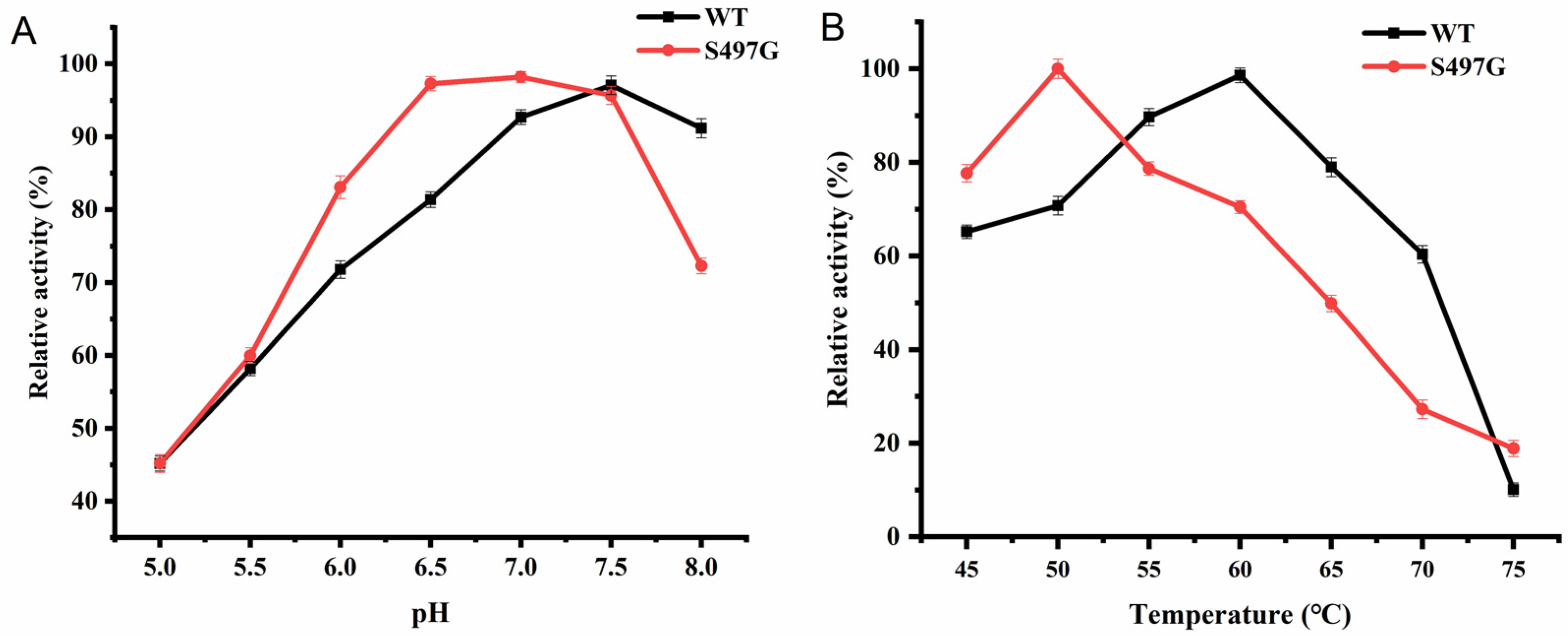
2.2. Catalytic Properties of CtCBP Variant S497G
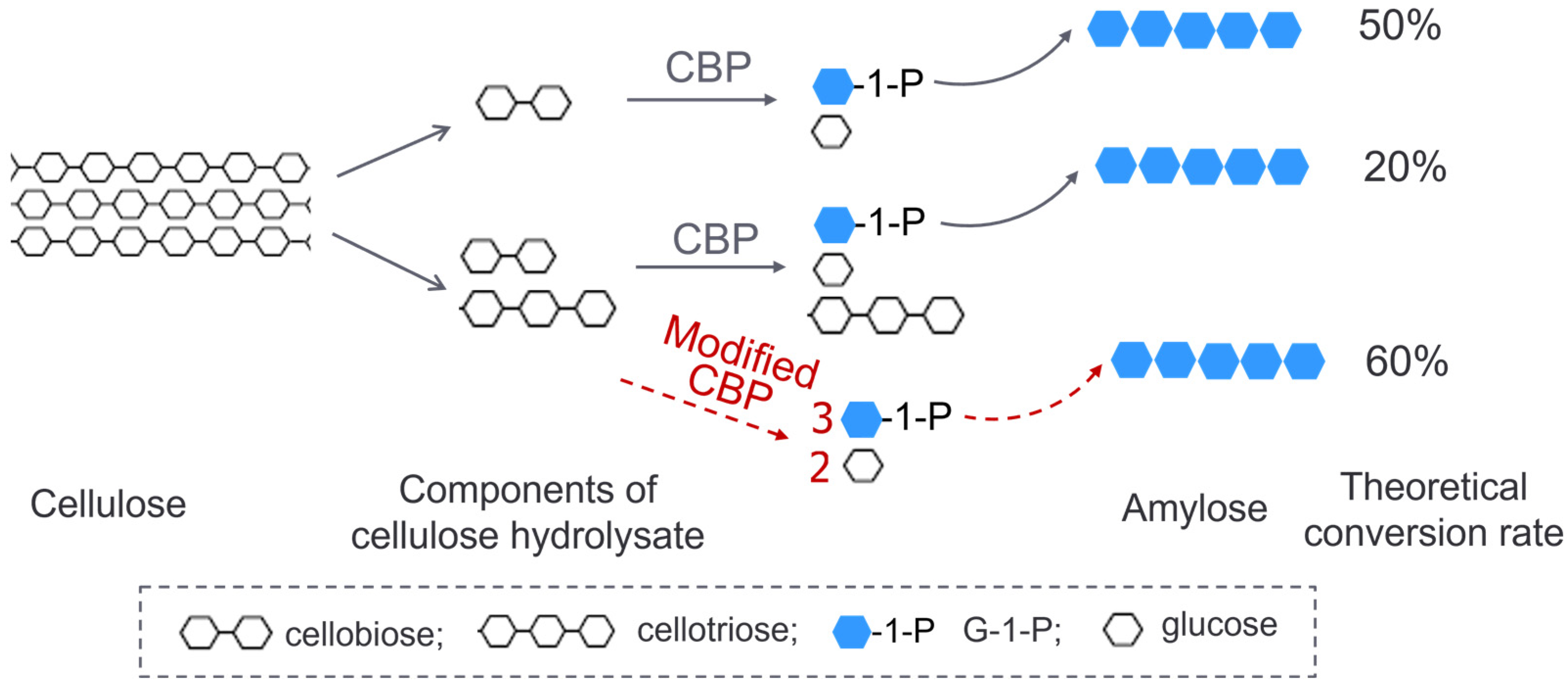
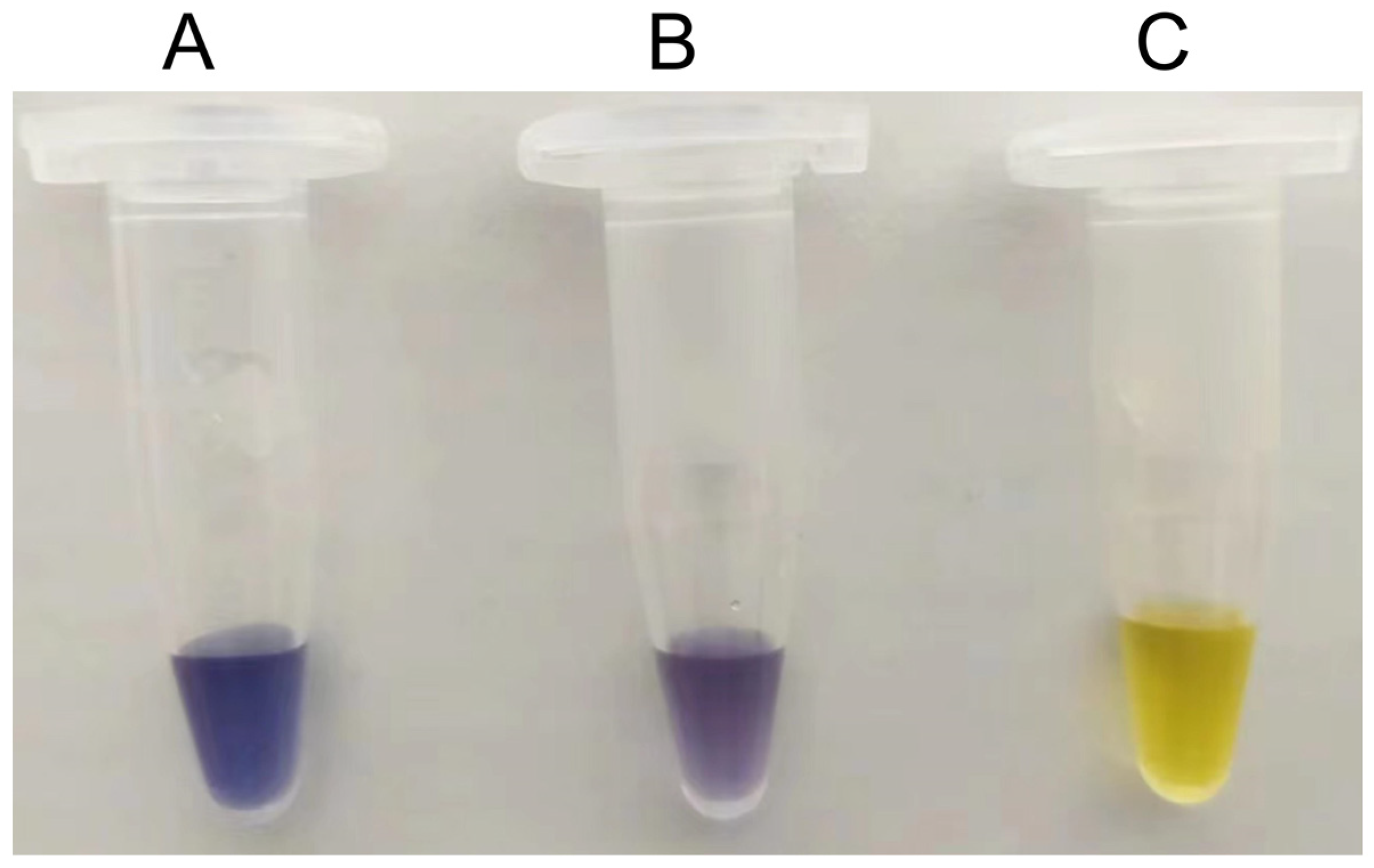
2.3. Transformation of Cellodextrin to Amylose
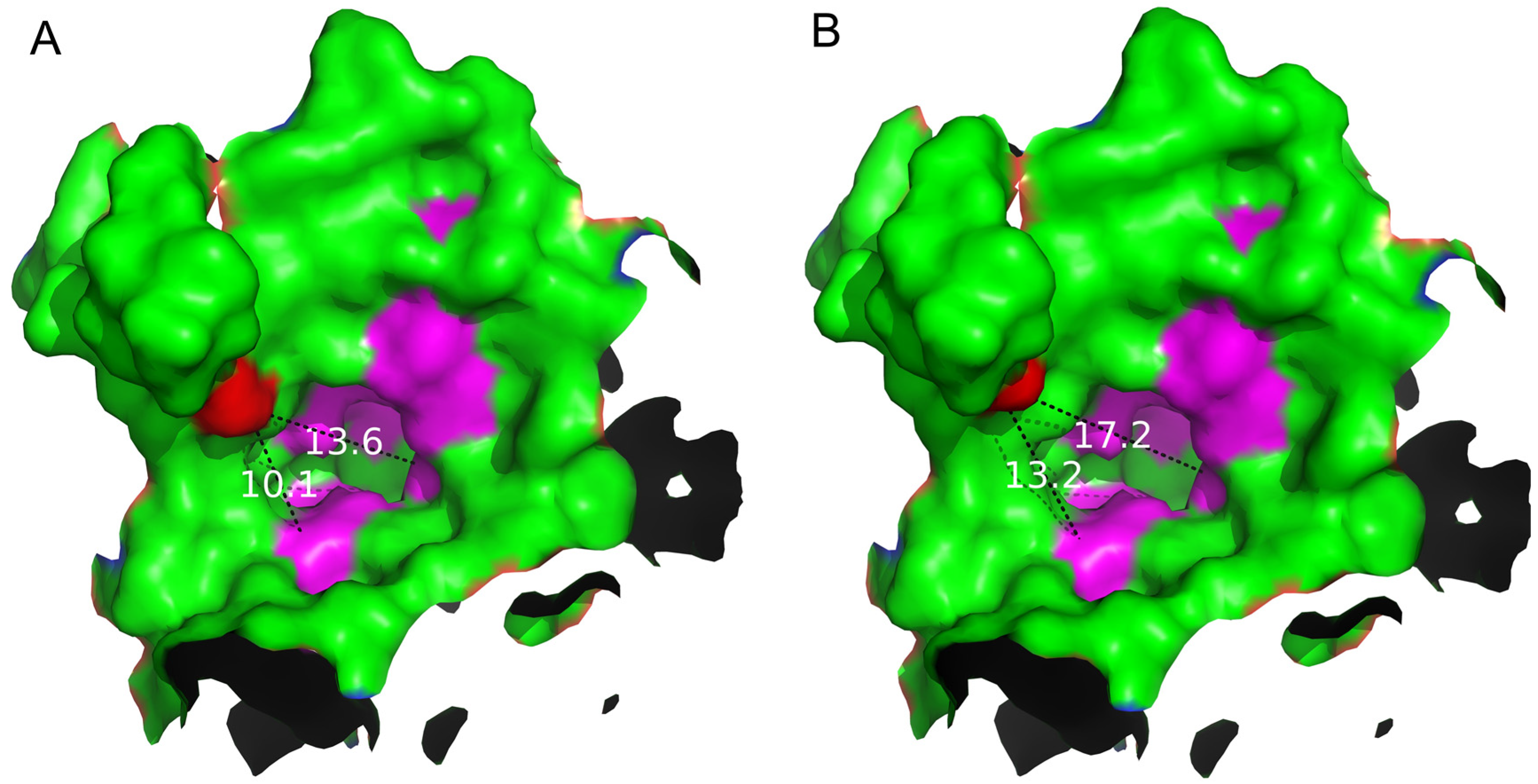
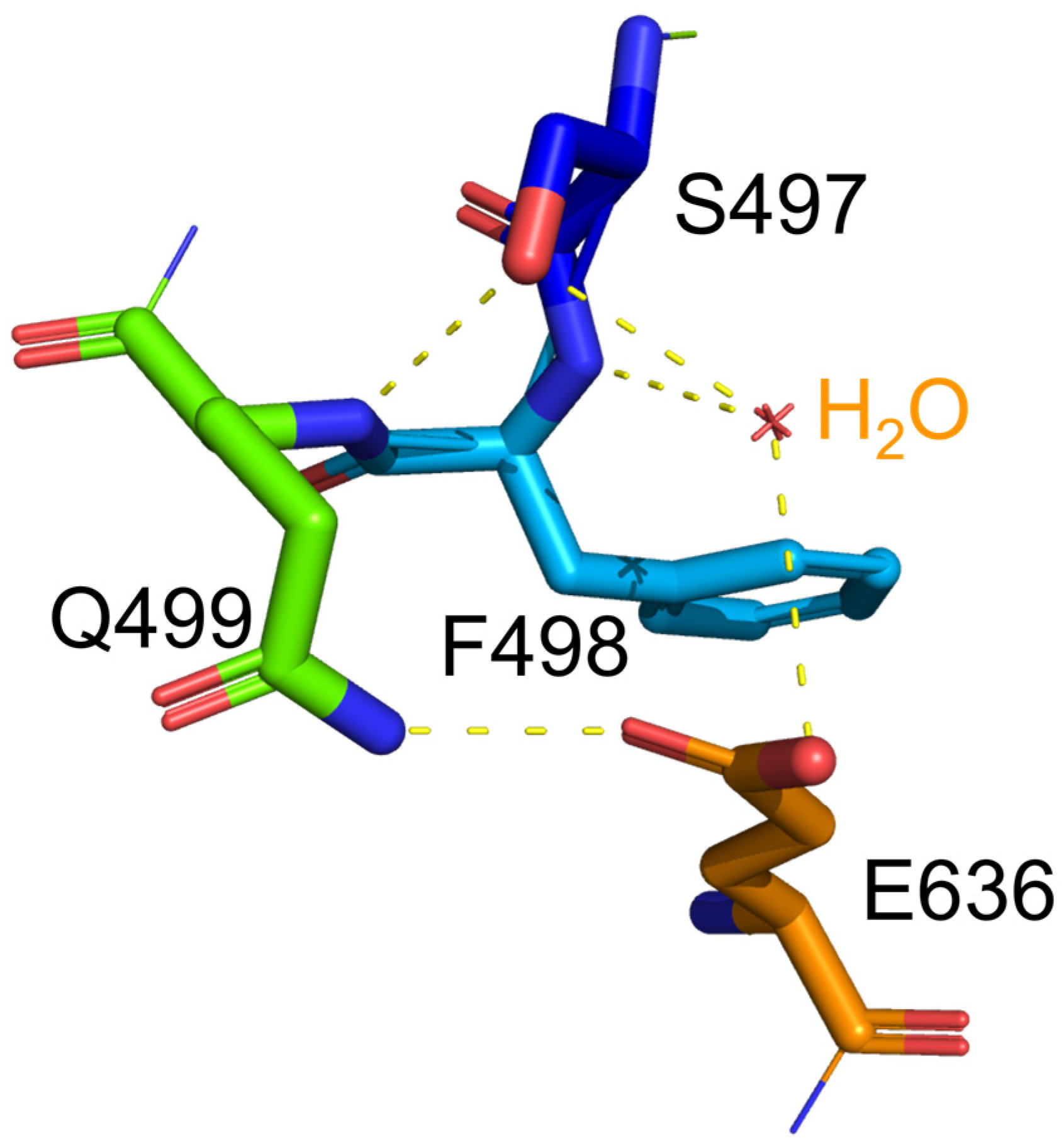
2.4. Structure Basis for Properties of S497G Variant
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Mutagenesis of CtCBP
4.3. Expression and Purification
4.4. Enzyme Assays
4.5. Kinetic Analysis
4.6. Transformation of Cellodextrin to Amylose
4.7. Homology Modeling and Structure Comparison
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanaka, K.; Kawaguchi, T.; Imada, Y.; Ooi, T.; Arai, M. Purification and properties of cellobiose phosphorylase from Clostridium thermocellum. J. Ferment. Bioeng. 1995, 79, 212–216. [Google Scholar] [CrossRef]
- Doudoroff, M. [28] Disaccharide phosphorylases. Methods Enzymol. 1955, 1, 225–231. [Google Scholar] [CrossRef]
- Hüttermann, A.; Volger, C. Cellobiose Phosphorylase in Fomes annosus. Nat. New Biol. 1973, 245, 64. [Google Scholar] [CrossRef] [PubMed]
- Reichenbecher, M.; Lottspeich, F.; Bronnenmeier, K. Purification and Properties of a Cellobiose Phosphorylase (CepA) and a Cellodextrin Phosphorylase (CepB) from the Cellulolytic Thermophile Clostridium Stercorarium. JBIC J. Biol. Inorg. Chem. 1997, 247, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Yernool, D.A.; Mccarthy, J.K.; Eveleigh, D.E.; Bok, J.D. Cloning and characterization of the glucooligosaccharide catabolic pathway beta-glucan glucohydrolase and cellobiose phosphorylase in the marine hyperthermophile Thermotoga neapolitana. J. Bacteriol. 2000, 182, 5172–5179. [Google Scholar] [CrossRef] [PubMed]
- Nidetzky, B.; Eis, C.; Albert, M. Role of non-covalent enzyme-substrate interactions in the reaction catalysed by cellobiose phosphorylase from Cellulomonas uda. Biochem. J. 2000, 351, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Hamura, K.; Saburi, W.; Abe, S.; Morimoto, N.; Taguchi, H.; Mori, H.; Matsui, H. Enzymatic Characteristics of Cellobiose Phosphorylase from Ruminococcus albus NE1 and Kinetic Mechanism of Unusual Substrate Inhibition in Reverse Phosphorolysis. Biosci. Biotechnol. Biochem. 2012, 76, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.-J.; Galazka, J.M.; Oh, E.J.; Kordić, V.; Kim, H.; Jin, Y.-S.; Cate, J.H. Energetic benefits and rapid cellobiose fermentation by Saccharomyces cerevisiae expressing cellobiose phosphorylase and mutant cellodextrin transporters. Metab. Eng. 2013, 15, 134–143. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Chen, H.; Myung, S.; Sathitsuksanoh, N.; Ma, H.; Zhang, X.-Z.; Li, J.; Zhang, Y.-H.P. Enzymatic transformation of nonfood biomass to starch. Proc. Natl. Acad. Sci. USA 2013, 110, 7182–7187. [Google Scholar] [CrossRef] [PubMed]
- Bianchetti, C.M.; Elsen, N.L.; Fox, B.G.; Phillips, G.N. The structure of Cellobiose phosphorylase from Clostridium thermocellum in complex with phosphate. Acta Crystallogr. Sect. F 2011, 67, 1345–1349. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.C.; Pergolizzi, G.; Stevenson, C.E.; Lawson, D.M.; Nepogodiev, S.A.; Field, R.A. Cellodextrin phosphorylase from Ruminiclostridium thermocellum: X-ray crystal structure and substrate specificity analysis. Carbohydr. Res. 2017, 451, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Ishida, A.; Watanabe, G.; Oshikawa, M.; Ajioka, I.; Muraoka, T. Glycine Substitution Effects on the Supramolecular Morphology and Rigidity of Cell-Adhesive Amphiphilic Peptides. Chem.—A Eur. J. 2019, 25, 13523–13530. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, M.; Igarashi, K.; Kimura, S.; Wada, M.; Kitaoka, M.; Samejima, M. Synthesis of highly ordered cellulose II in vitro using cellodextrin phosphorylase. Carbohydr. Res. 2009, 344, 2468–2473. [Google Scholar] [CrossRef] [PubMed]
- De Groeve, M.R.M.; Remmery, L.; Van Hoorebeke, A.; Stout, J.; Desmet, T.; Savvides, S.N.; Soetaert, W. Construction of cellobiose phosphorylase variants with broadened acceptor specificity towards anomerically substituted glucosides. Biotechnol. Bioeng. 2010, 107, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Ubiparip, Z.; Moreno, D.S.; Beerens, K.; Desmet, T. Engineering of cellobiose phosphorylase for the defined synthesis of cellotriose. Appl. Microbiol. Biotechnol. 2020, 104, 8327–8337. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hou, H.; Li, Y.; Yang, S.; Lin, H.; Chen, H. Fusion of cellobiose phosphorylase and potato alpha-glucan phosphorylase facilitates substrate channeling for enzymatic conversion of cellobiose to starch. Prep. Biochem. Biotechnol. 2021, 52, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 245–248. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Mao, G.T.; Fan, H.Y.; Song, A.D.; Zhang, Y.-H.P.; Chen, H.G. Biochemical properties of GH94 cellodextrin phos-phorylase THA_1941 from a thermophilic eubacterium Thermosipho africanus TCF52B with cellobiose phosphorylase activity. Sci. Rep. 2017, 7, 4849. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Replaced Fragments of CtCBP | Introduced Fragments of CtCDP | Specific Activity | |
|---|---|---|---|---|
| Xylose (μmol/min/mg) | Cellobiose (μmol/h/mg) | |||
| WT | / | / | 2.95 ± 0.08 | 2.28 ± 0.04 |
| CBP-Δ1 | 465–510 (46 aa) | 606–663 (58 aa) | ND | ND |
| CBP-Δ2 | 484–510 (27 aa) | 627–663 (37 aa) | ND | ND |
| CBP-Δ3 | 486–510 (25 aa) | 627–663 (37 aa) | 0.11 ± 0.01 | 0.58 ± 0.01 |
| CBP-Δ4 | 491–510 (17 aa) | 632–660 (29 aa) | 0.15 ± 0.02 | 3.20 ± 0.11 |
| CBP-Δ5 | 491–494 (4 aa) | 632–650 (19 aa) | 0.20 ± 0.01 | 4.74 ± 0.15 |
| Enzyme | Specific Activity | |
|---|---|---|
| Xylose (μmol/min/mg) | Cellobiose (μmol/h/mg) | |
| WT | 2.91 ± 0.07 | 2.02 ± 0.05 |
| CBP-GGGG | 0.09 ± 0.01 | 0.34 ± 0.02 |
| CBP-AAAA | 0.09 ± 0.01 | 0.16 ± 0.01 |
| S497G | 3.84 ± 0.12 | 4.67 ± 0.14 |
| Enzyme | Xylose | Cellobiose | ||||
|---|---|---|---|---|---|---|
| Km (mM) | kcat (s−1) | kcat/Km (s−1mM−1) | Km (mM) | kcat (s−1) | kcat/Km (s−1mM−1) | |
| WT | 38.26 ± 1.40 | 3.25 ± 0.13 | 0.09 | 10.32 ± 1.12 | 0.34 ± 0.03 | 0.03 |
| S497G | 6.94 ± 0.38 | 6.39 ± 0.15 | 0.92 | 5.36 ± 0.12 | 0.43 ± 0.02 | 0.08 |
| Enzyme | Cellobiose | Cellotriose | ||||
|---|---|---|---|---|---|---|
| Km (mM) | kcat (s−1) | kcat/Km (s−1mM−1) | Km (mM) | kcat (s−1) | kcat/Km (s−1mM−1) | |
| WT | 8.47 ± 0.45 | 7.09 ± 0.46 | 0.84 | 8.65 ± 0.22 | 0.67 ± 0.05 | 0.08 |
| S497G | 6.62 ± 0.13 | 7.41 ± 0.35 | 1.12 | 3.48 ± 0.18 | 1.52 ± 0.10 | 0.44 |
| Enzyme | Reaction System | Amylose Yield (%) |
|---|---|---|
| WT | Cellobiose | 13.5 ± 0.3 b |
| S497G | Cellobiose | 15.0 ± 0.2 a |
| WT | Mixed cellodextrin | 11.7 ± 0.4 b |
| S497G | Mixed cellodextrin | 16.2 ± 0.2 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Li, Y.; Lin, H.; Mao, G.; Long, X.; Liu, X.; Chen, H. Broadening the Substrate Specificity of Cellobiose Phosphorylase from Clostridium thermocellum for Improved Transformation of Cellodextrin to Starch. Int. J. Mol. Sci. 2023, 24, 14452. https://doi.org/10.3390/ijms241914452
Zhang Y, Li Y, Lin H, Mao G, Long X, Liu X, Chen H. Broadening the Substrate Specificity of Cellobiose Phosphorylase from Clostridium thermocellum for Improved Transformation of Cellodextrin to Starch. International Journal of Molecular Sciences. 2023; 24(19):14452. https://doi.org/10.3390/ijms241914452
Chicago/Turabian StyleZhang, Yuanyuan, Yapeng Li, Hui Lin, Guotao Mao, Xiang Long, Xinyu Liu, and Hongge Chen. 2023. "Broadening the Substrate Specificity of Cellobiose Phosphorylase from Clostridium thermocellum for Improved Transformation of Cellodextrin to Starch" International Journal of Molecular Sciences 24, no. 19: 14452. https://doi.org/10.3390/ijms241914452
APA StyleZhang, Y., Li, Y., Lin, H., Mao, G., Long, X., Liu, X., & Chen, H. (2023). Broadening the Substrate Specificity of Cellobiose Phosphorylase from Clostridium thermocellum for Improved Transformation of Cellodextrin to Starch. International Journal of Molecular Sciences, 24(19), 14452. https://doi.org/10.3390/ijms241914452