


Reduced Expression of Septin7 Hinders Skeletal Muscle Regeneration
 , , , and
, , , and 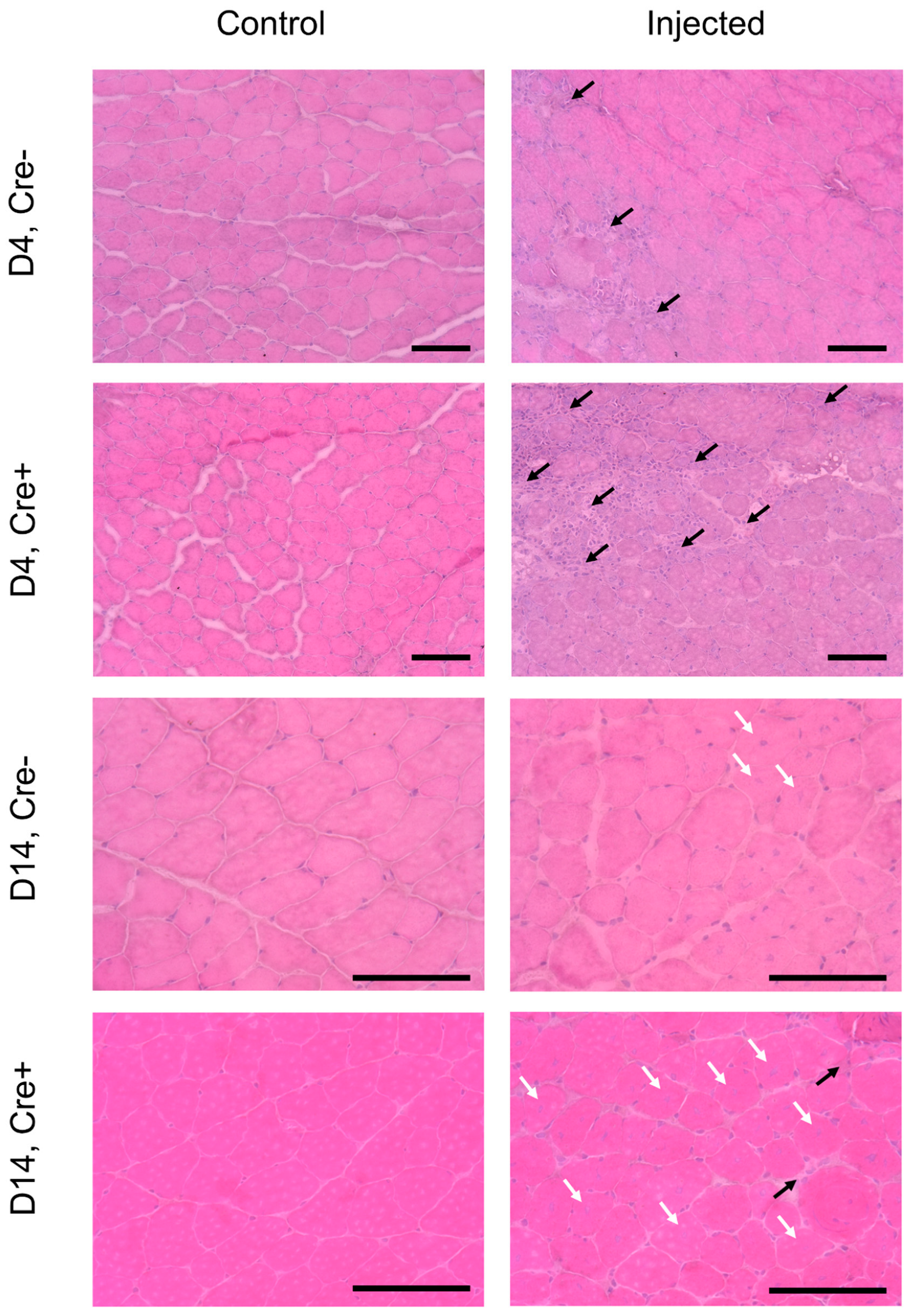{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
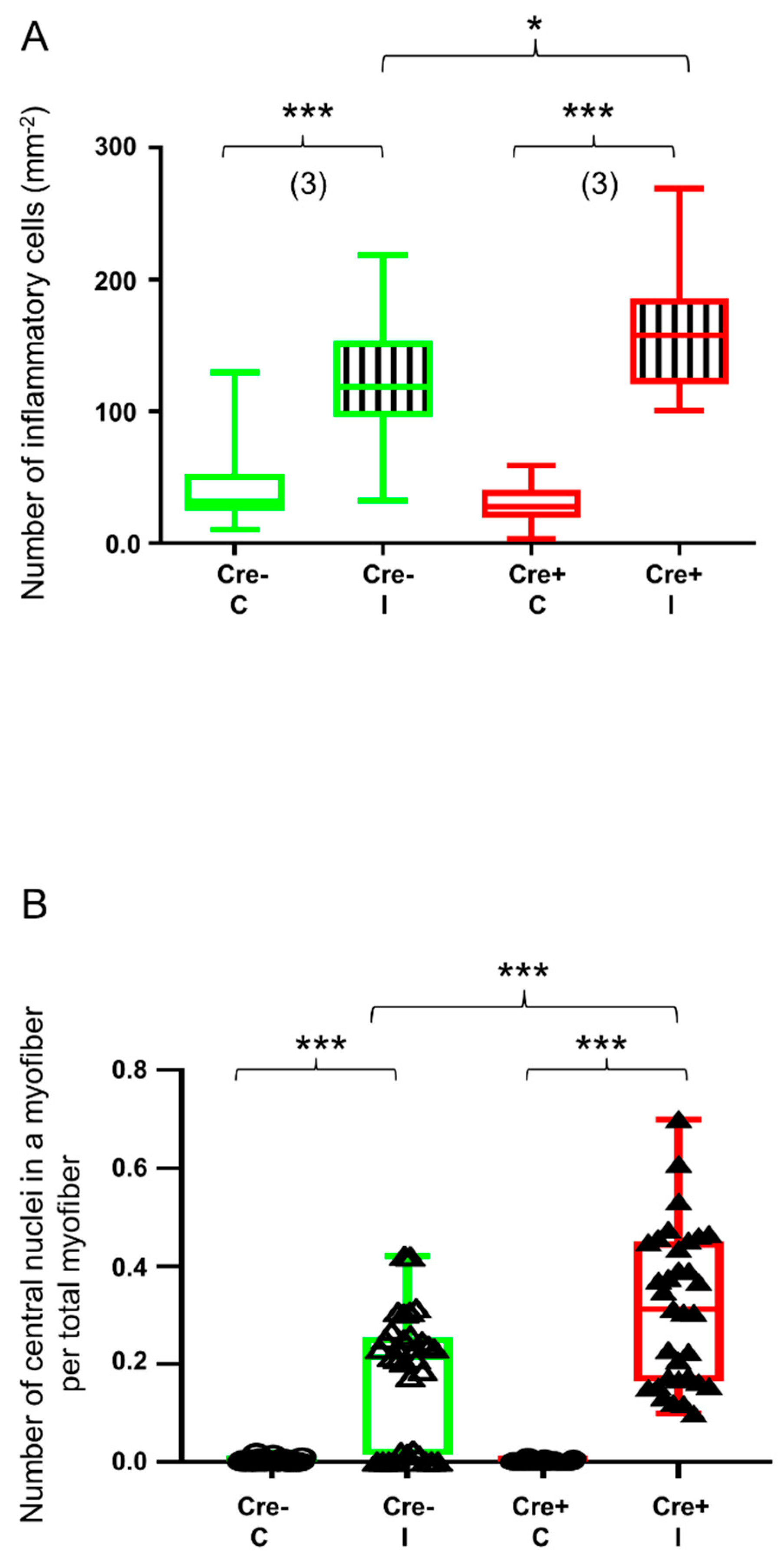
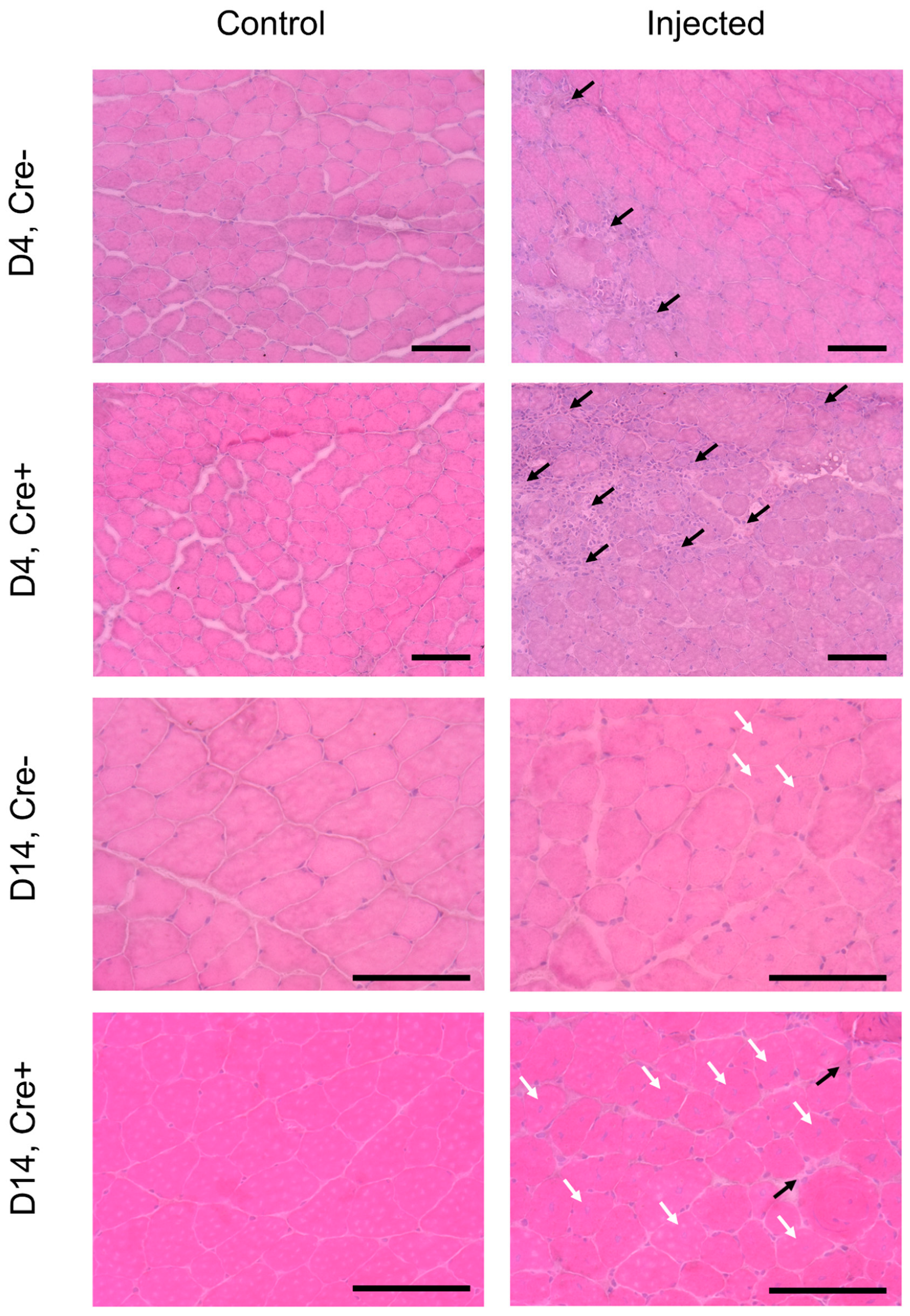
2.1. Skeletal Muscle Regeneration Is Hallmarked by Morphological Changes
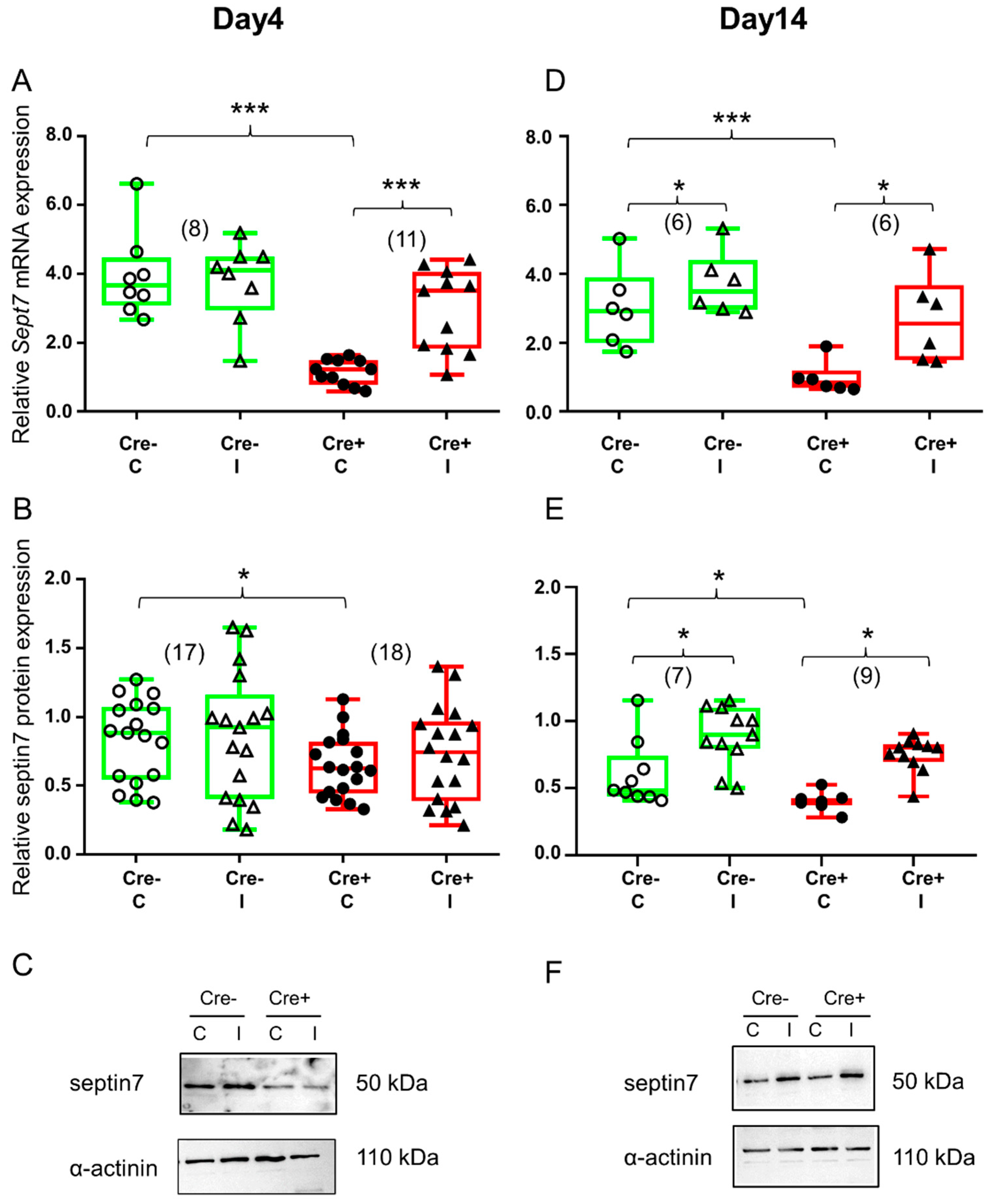
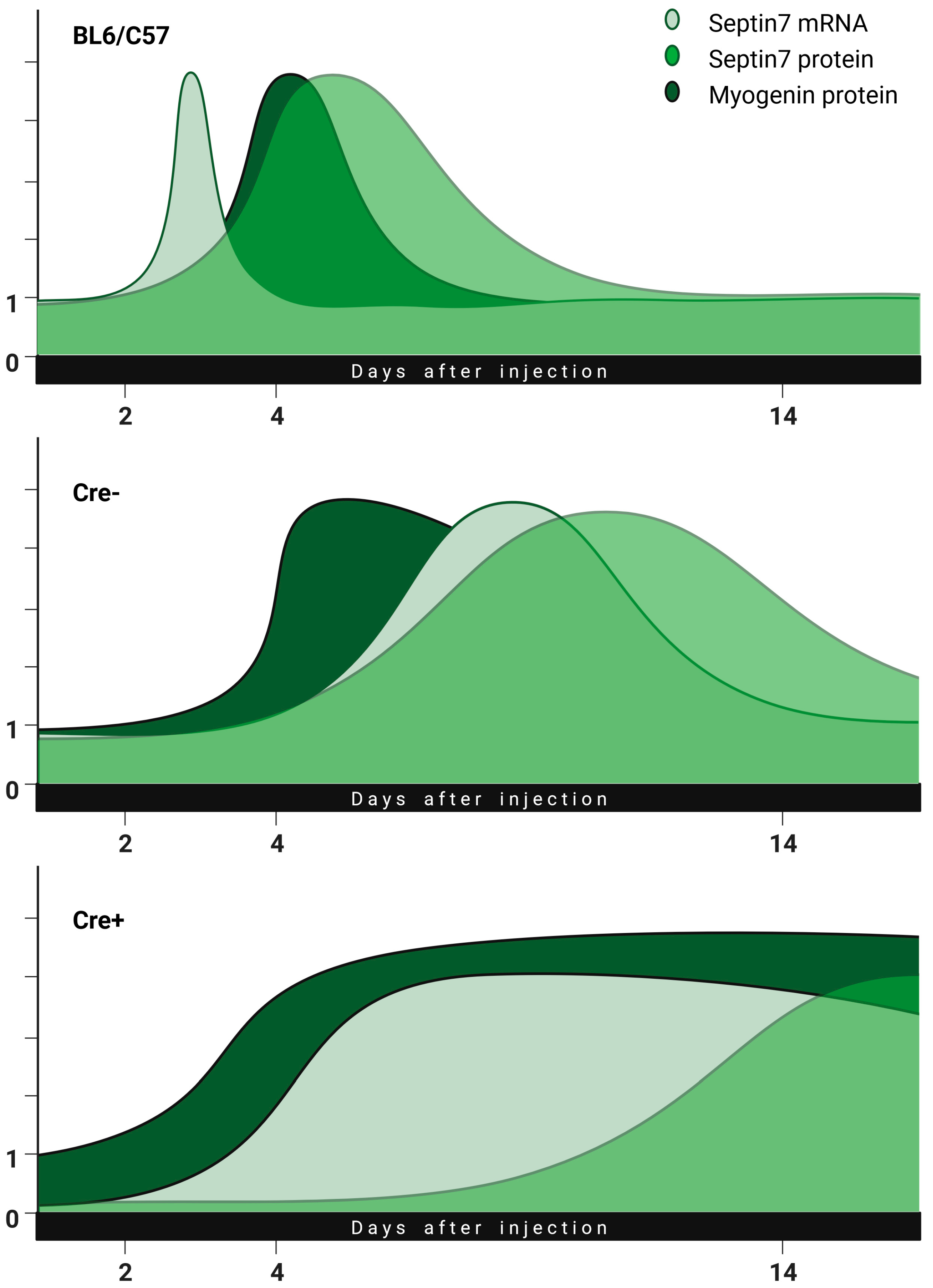
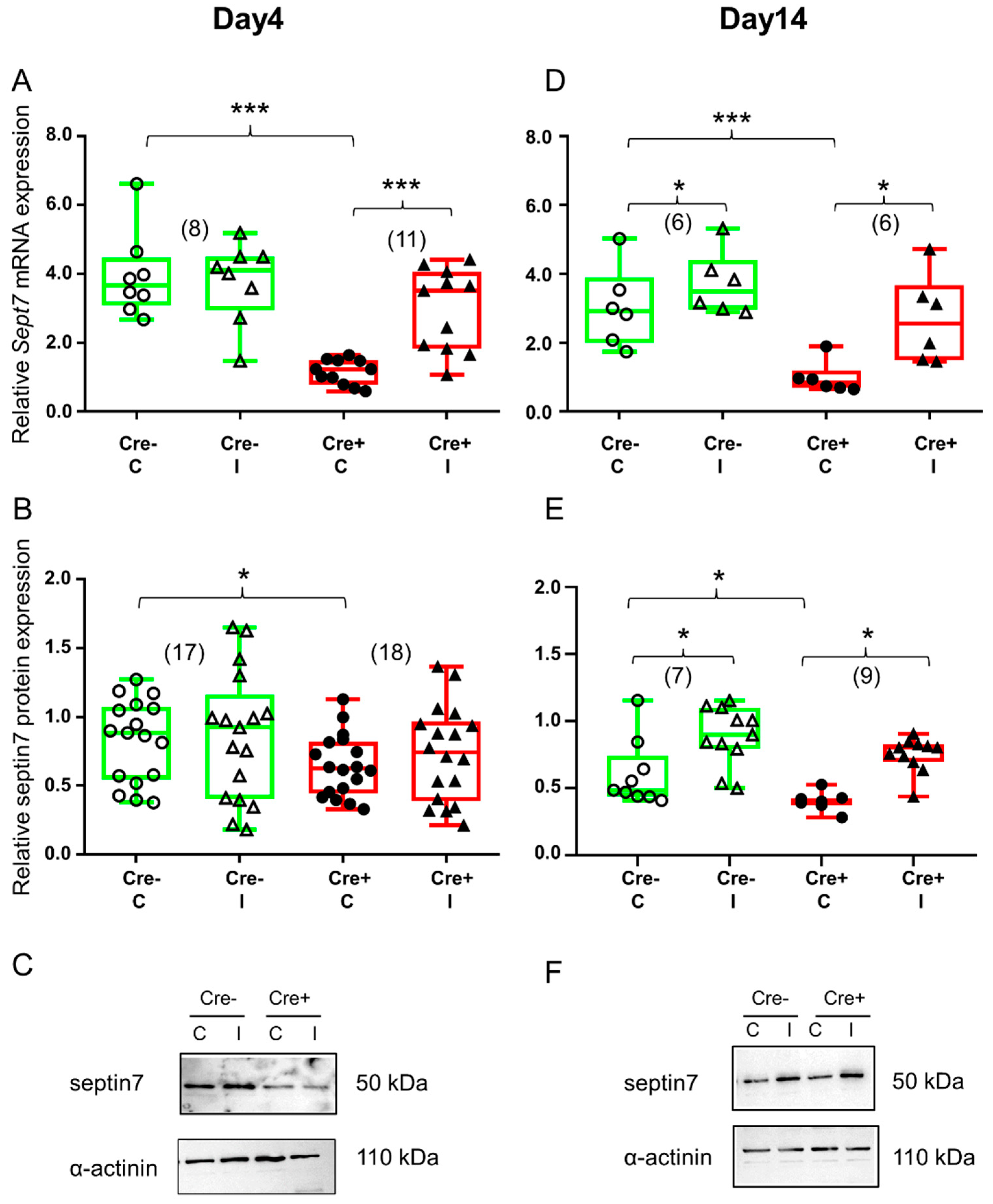
2.2. Skeletal Muscle Injury Induces Septin7 Upregulation
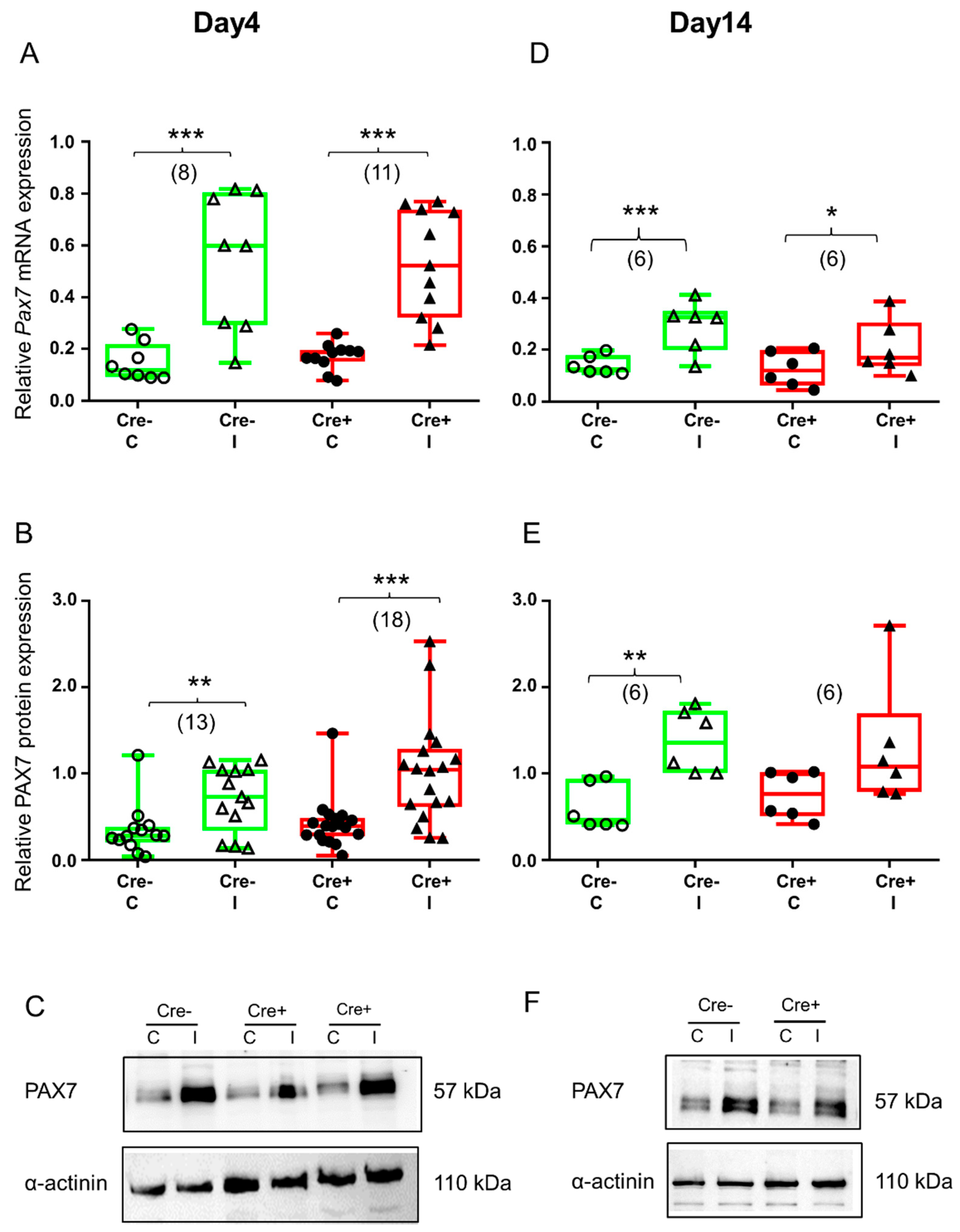
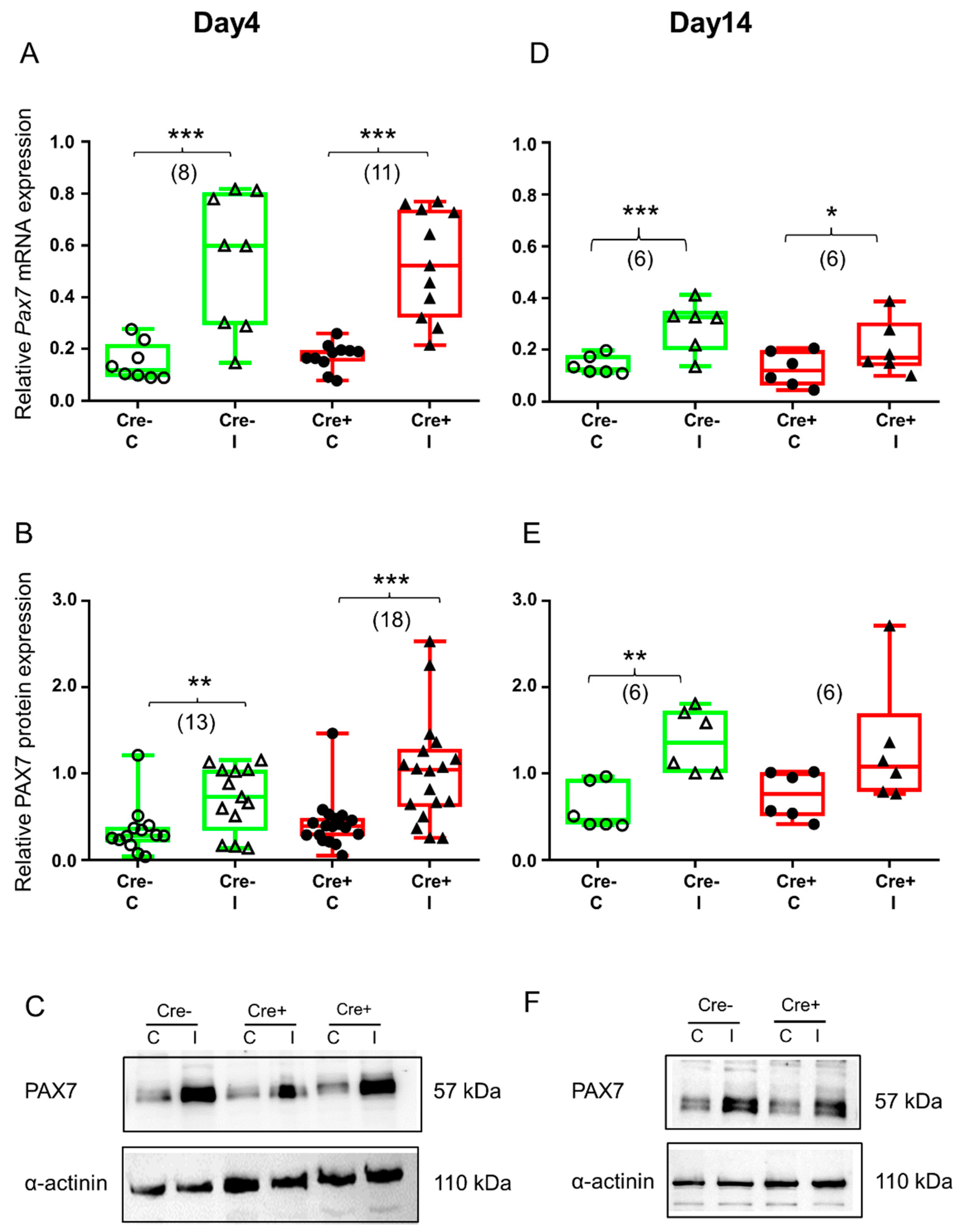
2.3. Upregulation of Transcription Factor PAX7 Accompanies Regeneration
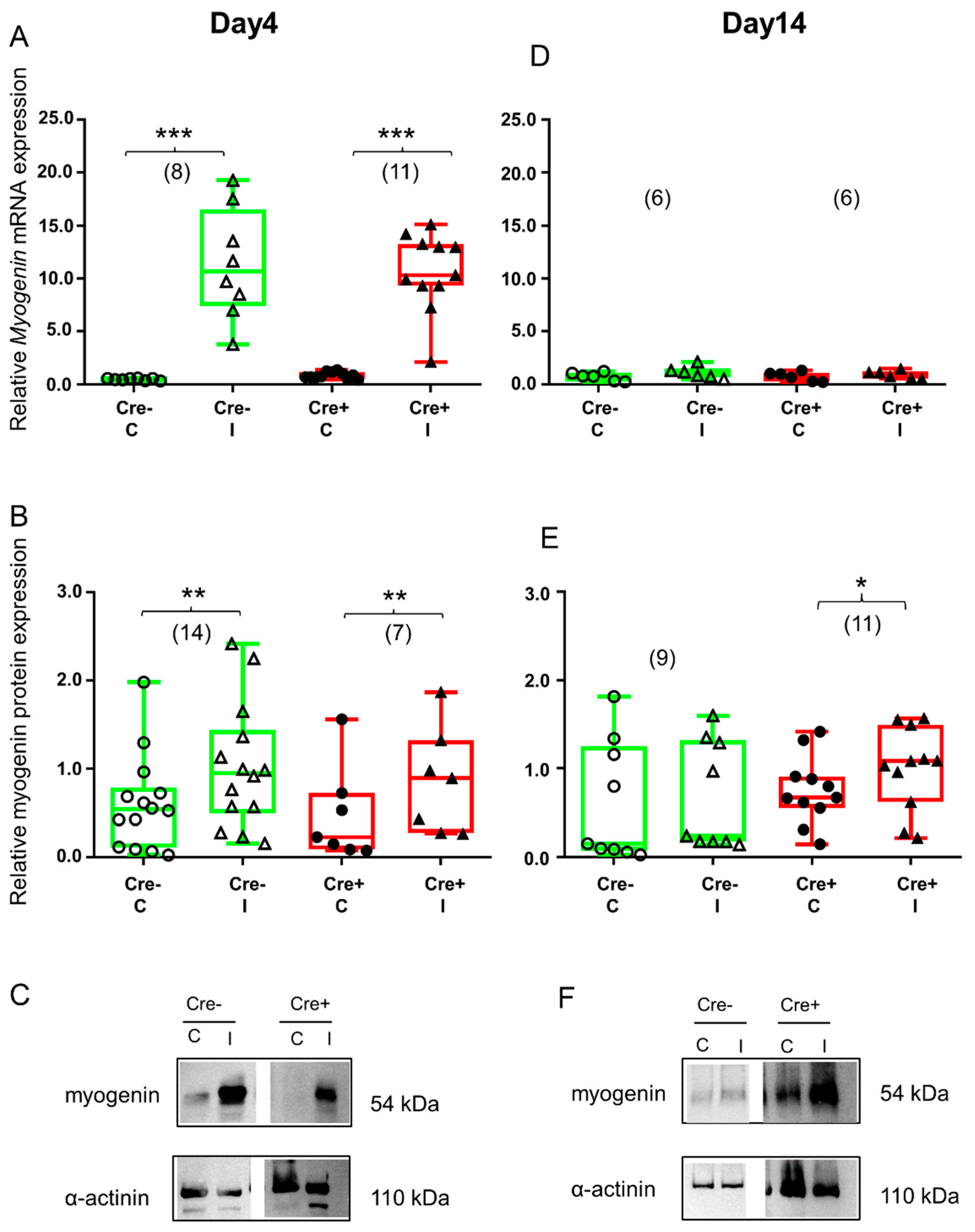
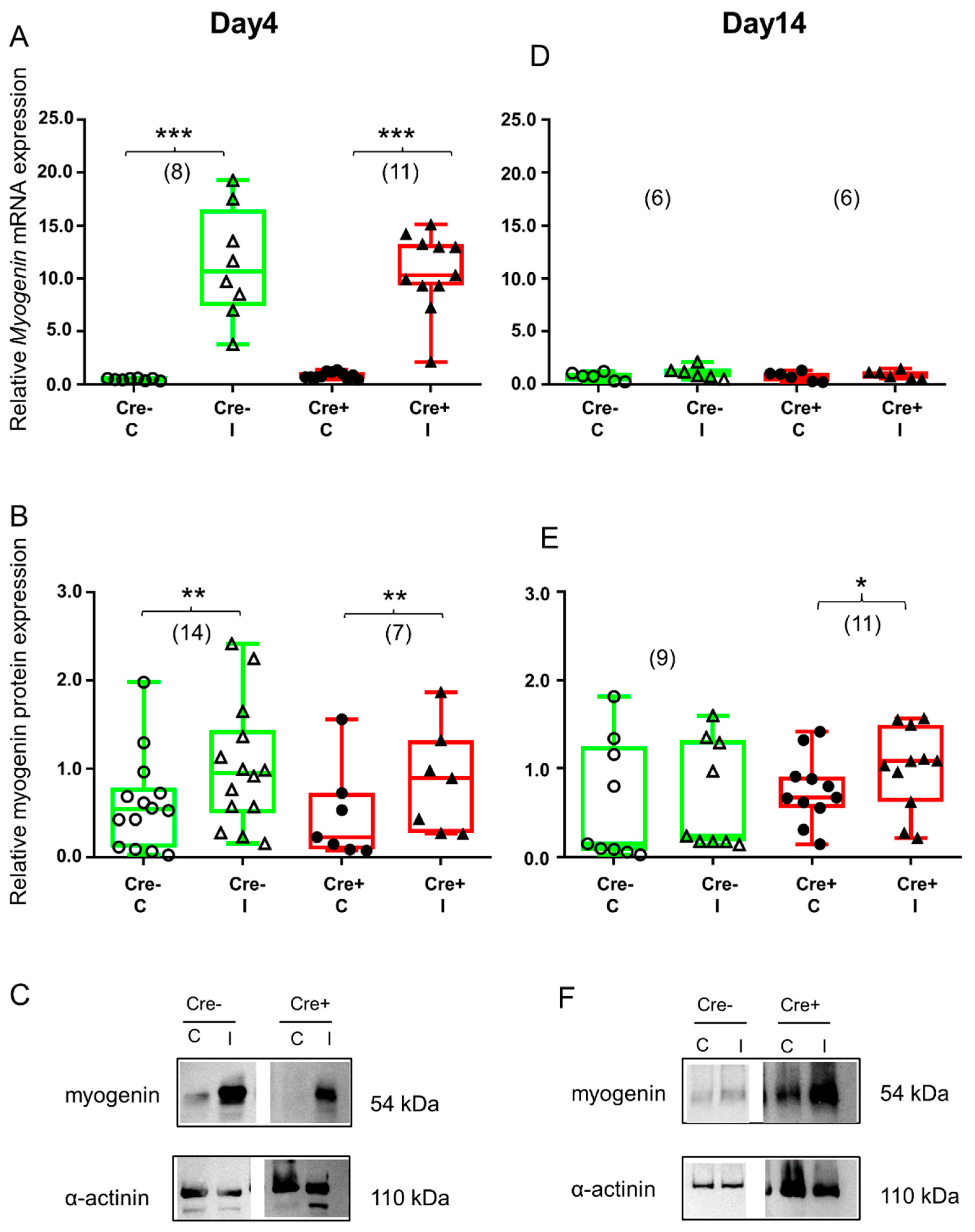
2.4. Transcription Factor Myogenin Is Upregulated during the Regeneration Phase
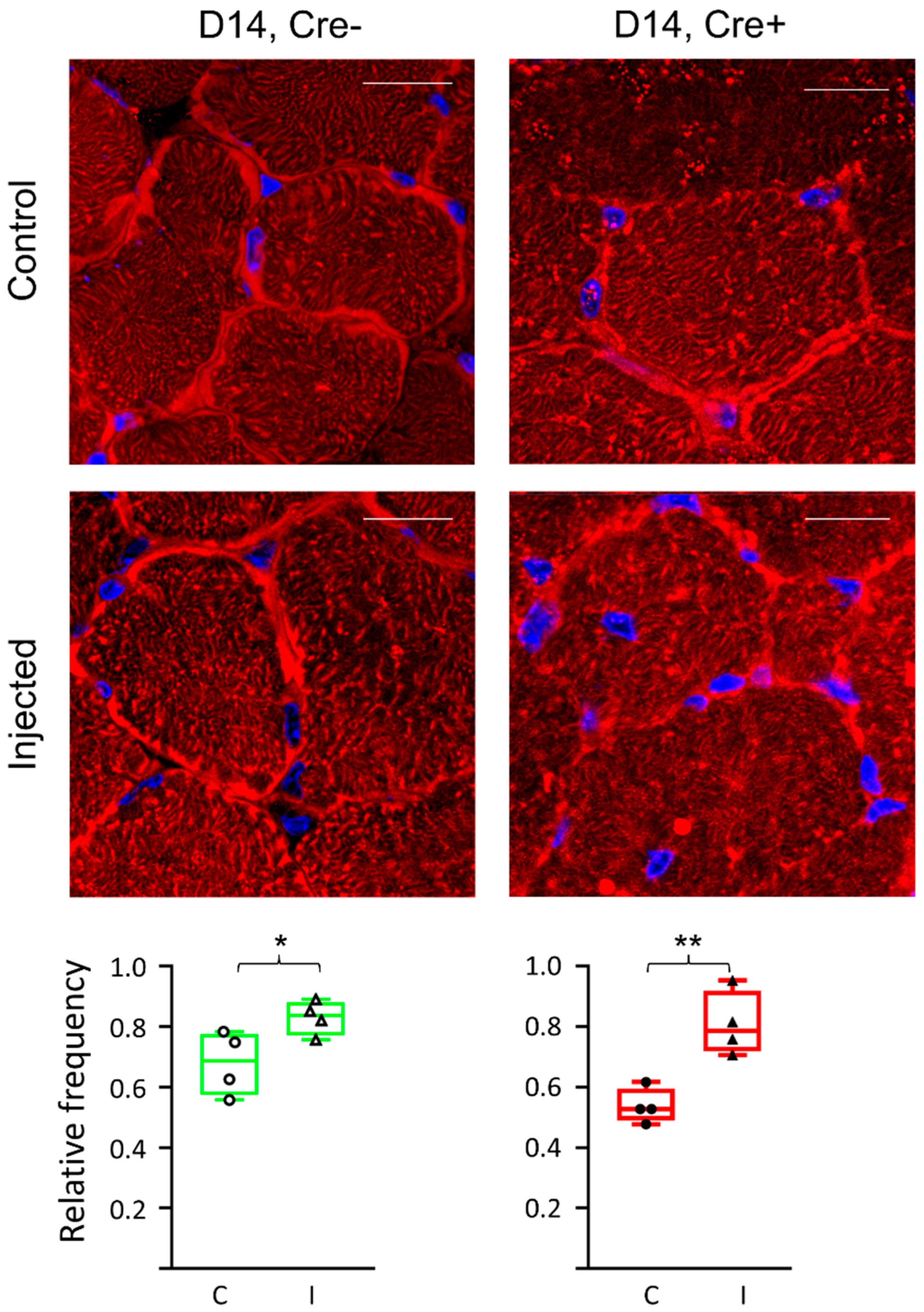
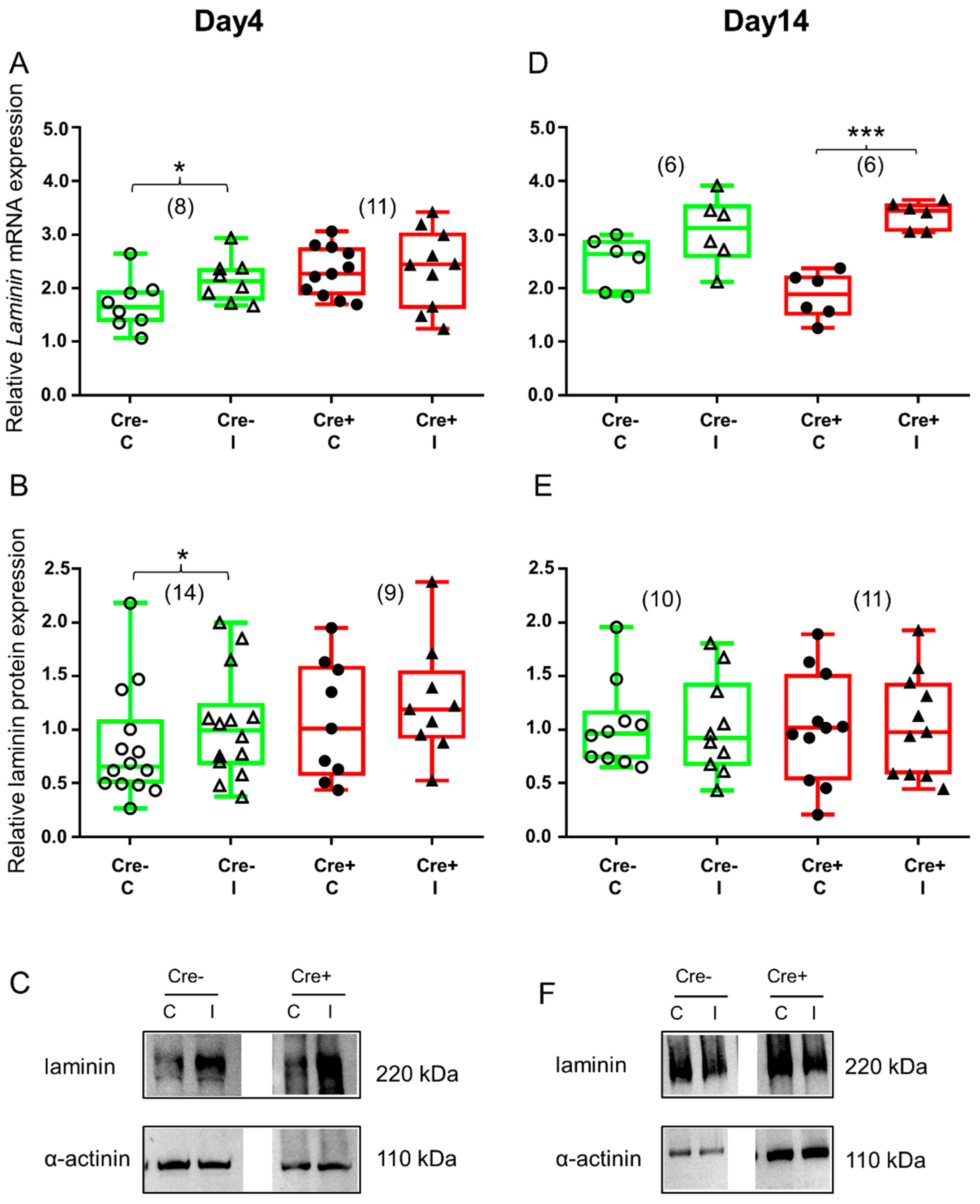
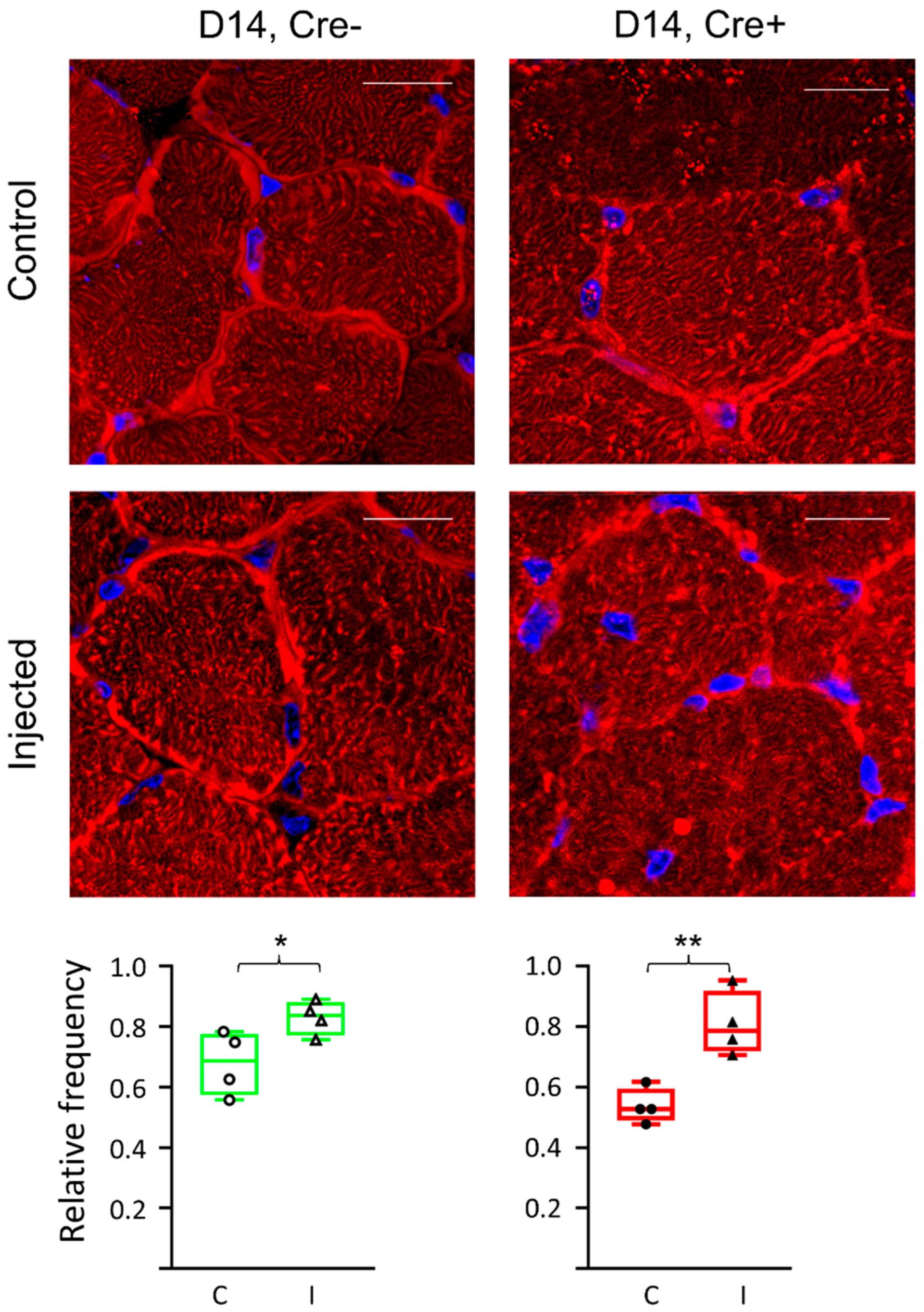
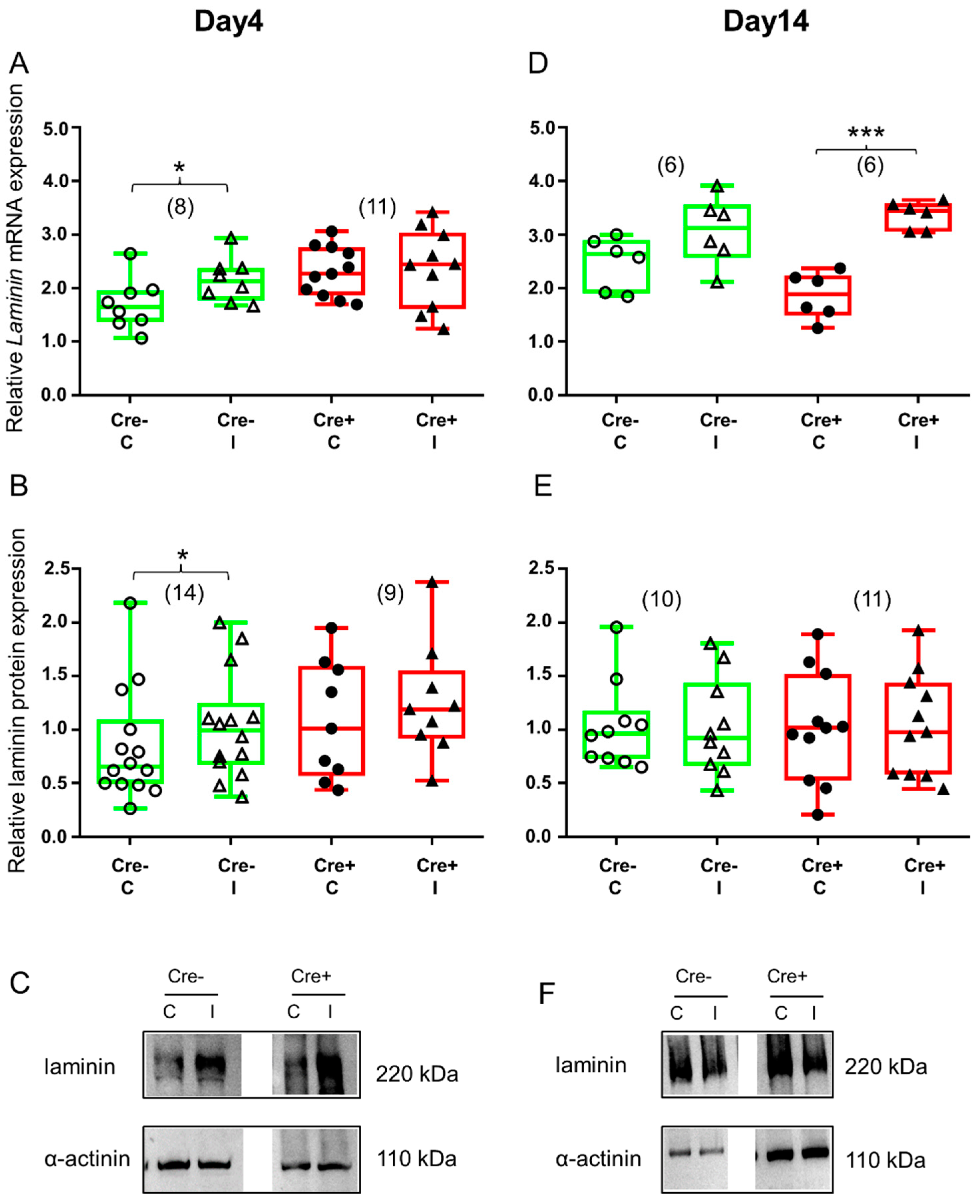
2.5. Differences in ECM Molecule Laminin Expression Reflects Changes in the Final Step of Regeneration
2.6. Partial Knock-Down of Sept7 Delays Regeneration as Reflected in Morphological Changes
3. Discussion
4. Materials and Methods
4.1. Animal Care
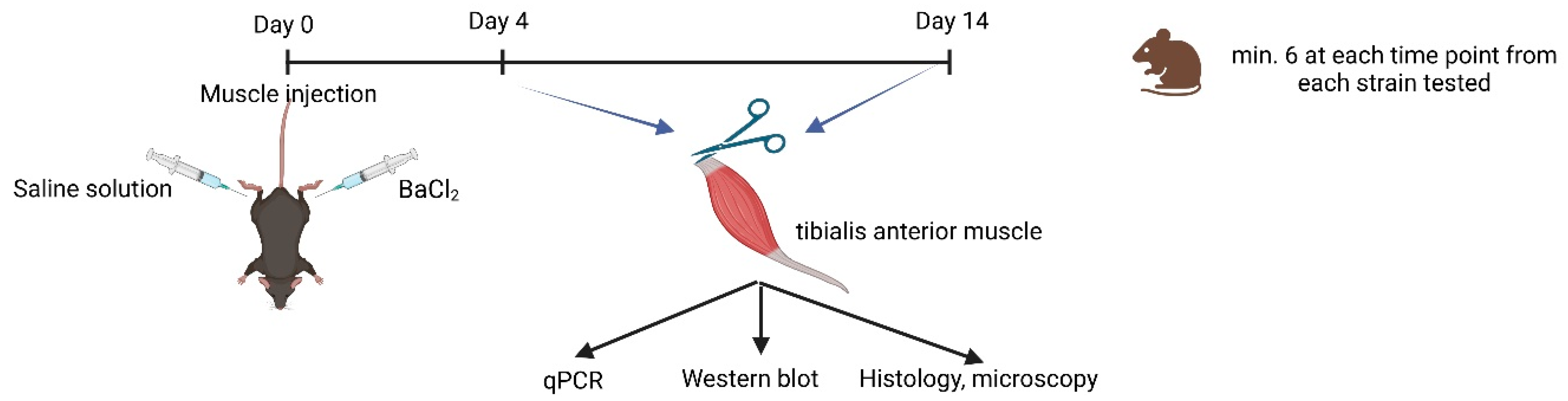
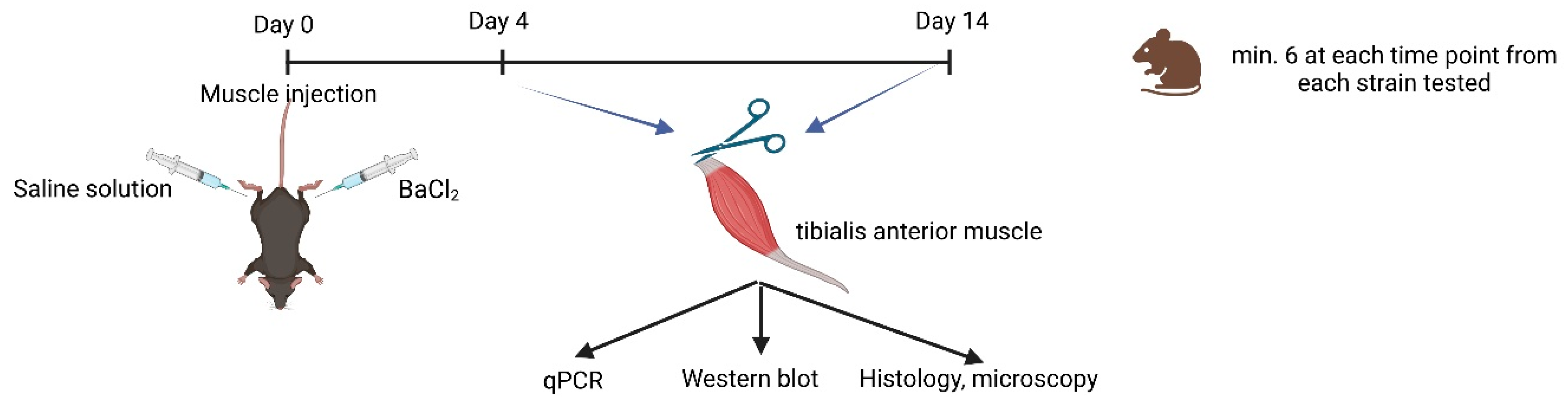
4.2. Muscle Regeneration
4.3. Experimental Design
4.4. Western Blotting
4.5. RNA Preparation, RT-PCR, and Quantitative Real-Time PCR
4.5.1. RNA Isolation
4.5.2. Reverse Transcription (RT) and Quantitative Polymerase Chain Reaction (qPCR)
4.5.3. Evaluation of qPCR Data
4.6. Immunolabeling and Confocal Imaging
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forcina, L.; Cosentino, M.; Musarò, A. Mechanisms Regulating Muscle Regeneration: Insights into the Interrelated and Time-Dependent Phases of Tissue Healing. Cells 2020, 9, 1297. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed]
- Carosio, S.; Berardinelli, M.G.; Aucello, M.; Musarò, A. Impact of ageing on muscle cell regeneration. Ageing Res. Rev. 2011, 10, 35–42. [Google Scholar] [CrossRef]
- Dulak, J. Many roles for Pax7. Cell Cycle 2017, 16, 21–22. [Google Scholar] [CrossRef]
- Le Moal, E.; Pialoux, V.; Juban, G.; Groussard, C.; Zouhal, H.; Chazaud, B.; Mounier, R. Redox Control of Skeletal Muscle Regeneration. Antioxid. Redox Signal. 2017, 27, 276–310. [Google Scholar] [CrossRef]
- Snijders, T.; Nederveen, J.P.; McKay, B.R.; Joanisse, S.; Verdijk, L.B.; van Loon, L.J.C.; Parise, G. Satellite cells in human skeletal muscle plasticity. Front. Physiol. 2015, 6, 283. [Google Scholar] [CrossRef]
- Ahrens, H.E.; Henze, H.; Schüler, S.C.; Schmidt, M.; Hüttner, S.S.; von Maltzahn, J. Analyzing satellite cell function during skeletal muscle regeneration by cardiotoxin injury and injection of self-delivering siRNA in vivo. J. Vis. Exp. 2019, e60194. [Google Scholar] [CrossRef]
- Mukund, K.; Subramaniam, S. Skeletal muscle: A review of molecular structure and function, in health and disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1462. [Google Scholar] [CrossRef]
- Hicks, M.R.; Pyle, A.D. The emergence of the stem cell niche. Trends Cell Biol. 2022, 33, 112–123. [Google Scholar] [CrossRef]
- Kann, A.P.; Hung, M.; Krauss, R.S. Cell–cell contact and signaling in the muscle stem cell niche. Curr. Opin. Cell Biol. 2021, 73, 78–83. [Google Scholar] [CrossRef]
- Seale, P.; Sabourin, L.A.; Girgis-Gabardo, A.; Mansouri, A.; Gruss, P.; Rudnicki, M.A. Pax7 is required for the specification of myogenic satellite cells. Cell 2000, 102, 777–786. [Google Scholar] [CrossRef]
- Von Maltzahn, J.; Jones, A.E.; Parks, R.J.; Rudnicki, M.A. Pax7 is critical for the normal function of satellite cells in adult skeletal muscle. Proc. Natl. Acad. Sci. USA 2013, 110, 16474–16479. [Google Scholar] [CrossRef]
- Addicks, G.C.; Brun, C.E.; Sincennes, M.C.; Saber, J.; Porter, C.J.; Francis Stewart, A.; Ernst, P.; Rudnicki, M.A. MLL1 is required for PAX7 expression and satellite cell self-renewal in mice. Nat. Commun. 2019, 10, 4256. [Google Scholar] [CrossRef]
- Buckingham, M.; Relaix, F. PAX3 and PAX7 as upstream regulators of myogenesis. Semin. Cell Dev. Biol. 2015, 44, 115–125. [Google Scholar] [CrossRef]
- Dueweke, J.J.; Awan, T.M.; Mendias, C.L. Regeneration of skeletal muscle after eccentric injury. J. Sport Rehabil. 2017, 26, 171–179. [Google Scholar] [CrossRef]
- Rayagiri, S.S.; Ranaldi, D.; Raven, A.; Mohamad Azhar, N.I.F.; Lefebvre, O.; Zammit, P.S.; Borycki, A.G. Basal lamina remodeling at the skeletal muscle stem cell niche mediates stem cell self-renewal. Nat. Commun. 2018, 9, 1075. [Google Scholar] [CrossRef]
- Baghdadi, M.B.; Tajbakhsh, S. Regulation and phylogeny of skeletal muscle regeneration. Dev. Biol. 2018, 433, 200–209. [Google Scholar] [CrossRef]
- Calve, S.; Hans-Georg, S. Biochemical and mechanical environment cooperatively regulate skeletal muscle regeneration. FASEB J. 2012, 26, 2538–2554. [Google Scholar] [CrossRef]
- Csapo, R.; Gumpenberger, M.; Wessner, B. Skeletal Muscle Extracellular Matrix—What Do We Know About Its Composition, Regulation, and Physiological Roles? A Narrative Review. Front. Physiol. 2020, 11, 253. [Google Scholar] [CrossRef]
- Marquardt, J.; Chen, X.; Bi, E. Architecture, remodeling, and functions of the septin cytoskeleton. Cytoskeleton 2019, 76, 7–14. [Google Scholar] [CrossRef]
- Addi, C.; Bai, J.; Echard, A. Actin, microtubule, septin and ESCRT filament remodeling during late steps of cytokinesis. Curr. Opin. Cell Biol. 2018, 50, 27–34. [Google Scholar] [CrossRef]
- Wang, X.; Fei, F.; Qu, J.; Li, C.; Li, Y.; Zhang, S. The role of septin 7 in physiology and pathological disease: A systematic review of current status. J. Cell. Mol. Med. 2018, 22, 3298–3307. [Google Scholar] [CrossRef] [PubMed]
- Akhmetova, K.A.; Chesnokov, I.N.; Fedorova, S.A. Functional Characterization of Septin Complexes. Mol. Biol. 2018, 52, 137–150. [Google Scholar] [CrossRef]
- DeRose, B.T.; Kelley, R.S.; Ravi, R.; Kokona, B.; Beld, J.; Spiliotis, E.T.; Padrick, S.B. Production and analysis of a mammalian septin hetero-octamer complex. Cytoskeleton 2020, 77, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Soroor, F.; Kim, M.S.; Palander, O.; Balachandran, Y.; Collins, R.F.; Benlekbir, S.; Rubinstein, J.L.; Trimble, W.S. Revised subunit order of mammalian septin complexes explains their in vitro polymerization properties. Mol. Biol. Cell 2021, 32, 289–300. [Google Scholar] [CrossRef]
- Sirajuddin, M.; Farkasovsky, M.; Hauer, F.; Kühlmann, D.; Macara, I.G.; Weyand, M.; Stark, H.; Wittinghofer, A. Structural insight into filament formation by mammalian septins. Nature 2007, 449, 311–315. [Google Scholar] [CrossRef]
- Dash, S.N.; Narumanchi, S.; Paavola, J.; Perttunen, S.; Wang, H.; Lakkisto, P.; Tikkanen, I.; Lehtonen, S. Sept7b is required for the subcellular organization of cardiomyocytes and cardiac function in zebrafish. Am. J. Physiol.-Heart Circ. Physiol. 2017, 312, H1085–H1095. [Google Scholar] [CrossRef]
- Perumal, N.; Straßburger, L.; Herzog, D.P.; Müller, M.B.; Pfeiffer, N.; Grus, F.H.; Manicam, C. Bioenergetic shift and actin cytoskeleton remodelling as acute vascular adaptive mechanisms to angiotensin II in murine retina and ophthalmic artery. Redox Biol. 2020, 34, 101597. [Google Scholar] [CrossRef]
- Angyal, Á.; Gönczi, M.; Ráduly, Z.; Szabó, L.; Dobrosi, N.; Dienes, B.; Csernoch, L. Septin 7 has an essential role in differentiation of C2C12 cells. J. Muscle Res. Cell Motil. 2019, 40, 242–243. [Google Scholar]
- Dobrosi, N.; Szabó, L.; Ráduly, Z.; Gönczi, M.; Kis, G.; Cseri, K.; Dienes, B. Septin 7 has no role in ec-coupling but severly modifies skeletal muscle architecture. J. Muscle Res. Cell Motil. 2019, 40, 250. [Google Scholar]
- Gönczi, M.; Ráduly, Z.; Szabó, L.; Fodor, J.; Telek, A.; Dobrosi, N.; Balogh, N.; Szentesi, P.; Kis, G.; Antal, M.; et al. Septin7 is indispensable for proper skeletal muscle architecture and function. Elife 2022, 11, e75863. [Google Scholar] [CrossRef] [PubMed]
- Zammit, P.S.; Partridge, T.A.; Yablonka-Reuveni, Z. The skeletal muscle satellite cell: The stem cell that came in from the cold. J. Histochem. Cytochem. 2006, 54, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, J.M.; García-González, E.G.; Brun, C.E.; Rudnicki, M.A. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Le Grand, F.; Rudnicki, M.A. Skeletal muscle satellite cells and adult myogenesis. Curr. Opin. Cell Biol. 2007, 19, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Conboy, I.M.; Conboy, M.J.; Wagers, A.J.; Girma, E.R.; Weismann, I.L.; Rando, T.A. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature 2005, 433, 760–764. [Google Scholar] [CrossRef]
- Wen, Y.; Dungan, C.M.; Mobley, C.B.; Valentino, T.; von Walden, F.; Murach, K.A. Nucleus Type-Specific DNA Methylomics Reveals Epigenetic “Memory” of Prior Adaptation in Skeletal Muscle. Function 2021, 2, zqab038. [Google Scholar] [CrossRef]
- Robinson, D.C.; Dilworth, F.J. Chapter Eight—Epigenetic Regulation of Adult Myogenesis. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 126, pp. 235–284. [Google Scholar]
- Wang, Y.X.; Feige, P.; Brun, C.E.; Hekmatnejad, B.; Dumont, N.A.; Renaud, J.M.; Faulkes, S.; Guindon, D.E.; Rudnicki, M.A. EGFR-Aurka Signaling Rescues Polarity and Regeneration Defects in Dystrophin-Deficient Muscle Stem Cells by Increasing Asymmetric Divisions. Cell Stem Cell 2019, 24, 419–432.e6. [Google Scholar] [CrossRef]
- Lukjanenko, L.; Jung, M.J.; Hegde, N.; Perruisseau-Carrier, C.; Migliavacca, E.; Rozo, M.; Karaz, S.; Jacot, G.; Schmidt, M.; Li, L.; et al. Loss of fibronectin from the aged stem cell niche affects the regenerative capacity of skeletal muscle in mice. Nat. Med. 2016, 22, 897–905. [Google Scholar] [CrossRef]
- Roman, W.; Pinheiro, H.; Pimentel, M.R.; Segalés, J.; Oliveira, L.M.; García-domínguez, E.; Gómez-cabrera, M.C.; Serrano, A.L.; Gomes, E.R.; Muñoz-cánoves, P. Muscle Repair after Physiological Damage Relies on Nuclear Migration for Cellular Reconstruction. Science 2021, 359, 355–359. [Google Scholar] [CrossRef]
- Estey, M.P.; Kim, M.S.; Trimble, W.S. Septins. Curr. Biol. 2011, 21, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Spiliotis, E.T.; Nakos, K. Cellular functions of actin- and microtubule-associated septins. Curr. Biol. 2021, 31, R651–R666. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kim, S.; Shin, H.; Uhm, C.S. Intercellular interaction observed by atomic force microscopy. Ultramicroscopy 2008, 108, 1148–1151. [Google Scholar] [CrossRef] [PubMed]
- Nowak, S.J.; Nahirney, P.C.; Hadjantonakis, A.K.; Baylies, M.K. Nap1-mediated actin remodeling is essential for mammalian myoblast fusion. J. Cell Sci. 2009, 122, 3282–3293. [Google Scholar] [CrossRef] [PubMed]
- Pajcini, K.V.; Pomerantz, J.H.; Alkan, O.; Doyonnas, R.; Blau, H.M. Myoblasts and macrophages share molecular components that contribute to cell-cell fusion. J. Cell Biol. 2008, 180, 1005–1019. [Google Scholar] [CrossRef]
- Boomershine, K.H. Colchicine-Induced Rhabdomyolysis. Ann. Pharm. 2002, 36, 824–826. [Google Scholar] [CrossRef]
- Wilbur, K.; Makowsky, M. Colchicine myotoxicity: Case reports and literature review. Pharmacotherapy 2004, 24, 1784–1792. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabó, L.; Telek, A.; Fodor, J.; Dobrosi, N.; Dócs, K.; Hegyi, Z.; Gönczi, M.; Csernoch, L.; Dienes, B. Reduced Expression of Septin7 Hinders Skeletal Muscle Regeneration. Int. J. Mol. Sci. 2023, 24, 13536. https://doi.org/10.3390/ijms241713536
Szabó L, Telek A, Fodor J, Dobrosi N, Dócs K, Hegyi Z, Gönczi M, Csernoch L, Dienes B. Reduced Expression of Septin7 Hinders Skeletal Muscle Regeneration. International Journal of Molecular Sciences. 2023; 24(17):13536. https://doi.org/10.3390/ijms241713536
Chicago/Turabian StyleSzabó, László, Andrea Telek, János Fodor, Nóra Dobrosi, Klaudia Dócs, Zoltán Hegyi, Mónika Gönczi, László Csernoch, and Beatrix Dienes. 2023. "Reduced Expression of Septin7 Hinders Skeletal Muscle Regeneration" International Journal of Molecular Sciences 24, no. 17: 13536. https://doi.org/10.3390/ijms241713536
APA StyleSzabó, L., Telek, A., Fodor, J., Dobrosi, N., Dócs, K., Hegyi, Z., Gönczi, M., Csernoch, L., & Dienes, B. (2023). Reduced Expression of Septin7 Hinders Skeletal Muscle Regeneration. International Journal of Molecular Sciences, 24(17), 13536. https://doi.org/10.3390/ijms241713536