
CFTR Modulates Hypothalamic Neuron Excitability to Maintain Female Cycle


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. CFTR-Inhibition Evokes Electrical Pulses in Primary Rat Hypothalamic Neurons In Vitro
2.2. CFTR-Inhibition Induces Cl−-Dependent Ca2+ Responses in Primary Rat Hypothalamic Neurons and Glia Cells In Vitro
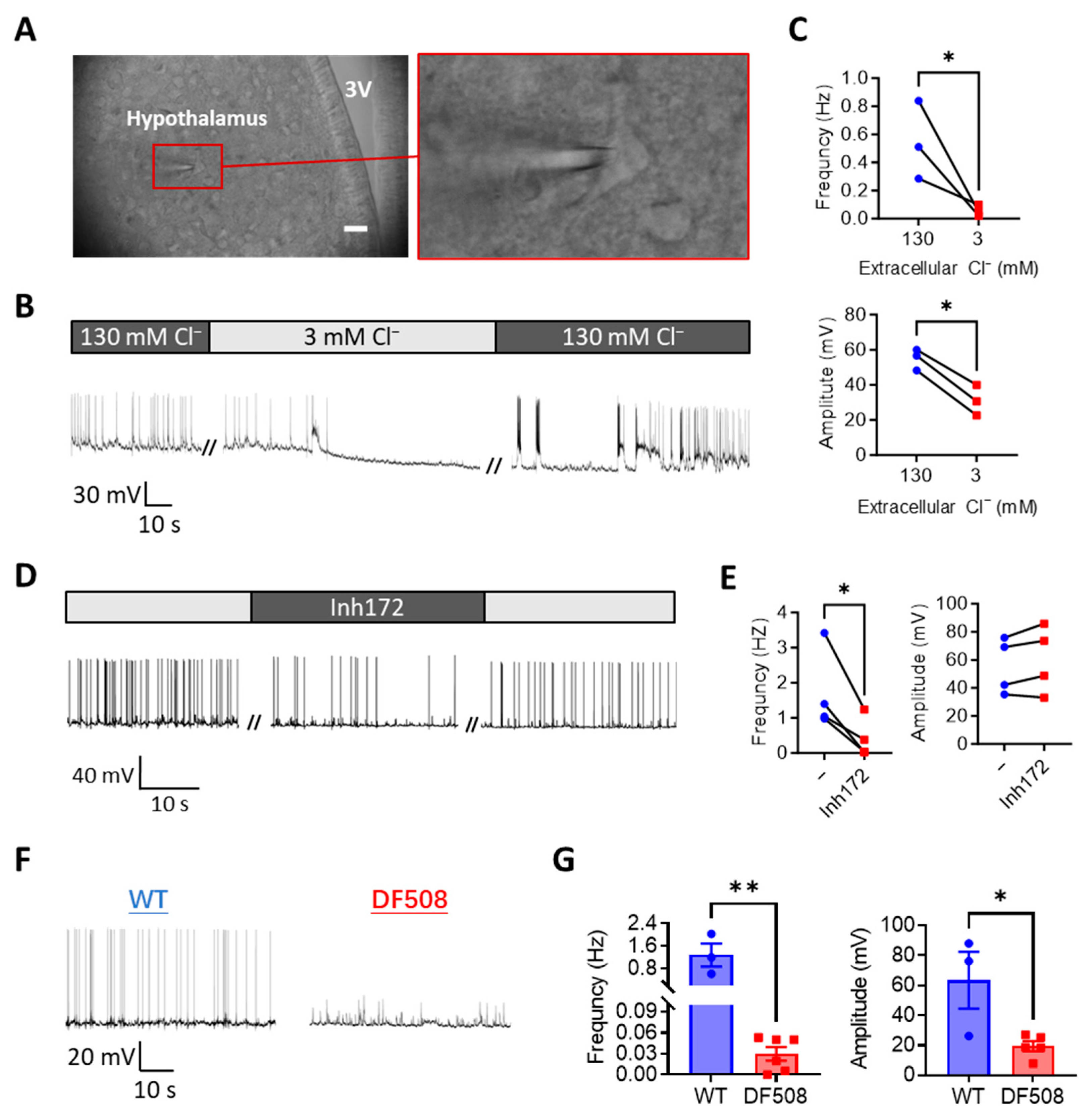
2.3. CFTR Deficiency or Cl− Removal Impairs Hypothalamic Electrical Firing in Mouse Brain Slices Ex Vivo
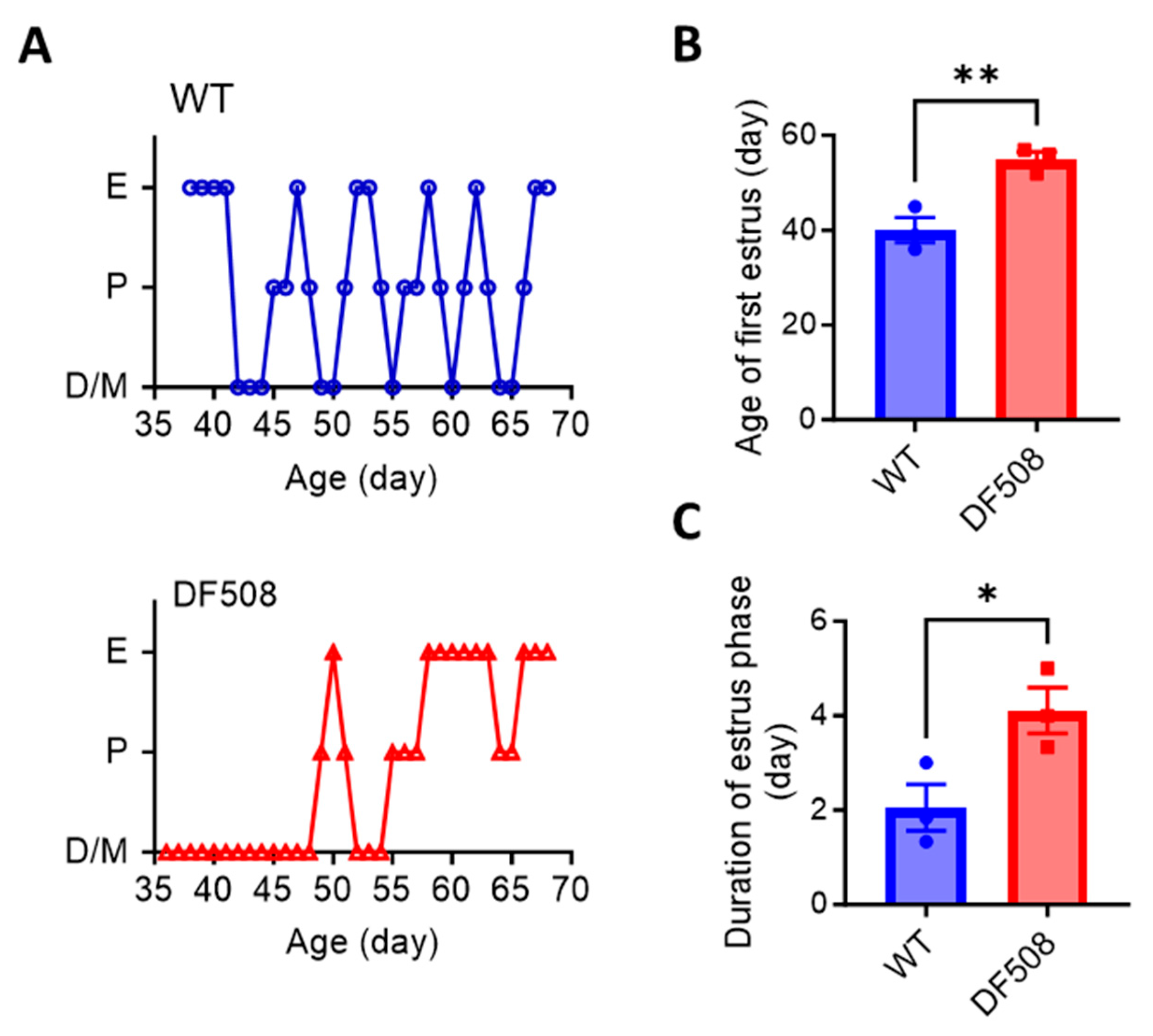
2.4. CFTR Deficiency Disturbs Mouse Estrus Cycle and Hypothalamic Neuron Activities In Vivo
3. Discussion
4. Methods and Materials
4.1. Animals
4.2. Primary Cultures of Hypothalamic Neurons
4.3. Patch-Clamp
4.4. Ca2+ Imaging
4.5. Brain Slice Patch-Clamp
4.6. Vaginal Cytology
4.7. Stereotaxic Surgery
4.8. Fiber Photometry
4.9. Immunofluorescence Staining
4.10. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Schreiber, R.; Nitschke, R.; Greger, R.; Kunzelmann, K. The cystic fibrosis transmembrane conductance regulator activates aquaporin 3 in airway epithelial cells. J. Biol. Chem. 1999, 274, 11811–11816. [Google Scholar] [CrossRef] [PubMed]
- Wuchu, F.; Ma, X.; Que, Y.; Chen, J.; Ruan, Y.C. Biphasic regulation of CFTR expression by ENaC in epithelial cells: The involvement of Ca2+-modulated cAMP production. Front. Cell Dev. Biol. 2022, 10, 781762. [Google Scholar] [CrossRef] [PubMed]
- Rich, D.P.; Anderson, M.P.; Gregory, R.J.; Cheng, S.H.; Paul, S.; Jefferson, D.M.; McCann, J.D.; Klinger, K.W.; Smith, A.E.; Welsh, M.J. Expression of cystic fibrosis transmembrane conductance regulator corrects defective chloride channel regulation in cystic fibrosis airway epithelial cells. Nature 1990, 347, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Klimova, B.; Kuca, K.; Novotny, M.; Maresova, P. Cystic Fibrosis Revisited—A Review Study. Med. Chem. 2017, 13, 102–109. [Google Scholar] [CrossRef]
- Houwen, R.H.; van der Doef, H.P.; Sermet, I.; Munck, A.; Hauser, B.; Walkowiak, J.; Robberecht, E.; Colombo, C.; Sinaasappel, M.; Wilschanski, M. Defining DIOS and constipation in cystic fibrosis with a multicentre study on the incidence, characteristics, and treatment of DIOS. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 38–42. [Google Scholar] [CrossRef]
- Ruan, Y.C.; Wang, Y.; Da Silva, N.; Kim, B.; Diao, R.Y.; Hill, E.; Brown, D.; Chan, H.C.; Breton, S. CFTR interacts with ZO-1 to regulate tight junction assembly and epithelial differentiation via the ZONAB pathway. J. Cell Sci. 2014, 127, 4396–4408. [Google Scholar] [CrossRef]
- Goertchen, P.; Nizze, H.; Gould, N.S.; Gould, V.E.; Oberholzer, M.; Heitz, P.U.; Löhr, M.; Klöoppel, G. Cystic fibrosis associated islet changes may provide a basis for diabetes. Virchows Arch. 1989, 414, 179–185. [Google Scholar] [CrossRef]
- Noronha, R.M.; Calliari, L.E.P.; Damaceno, N.; Muramatu, L.H.; Monte, O. Update on diagnosis and monitoring of cystic fibrosis-related diabetes mellitus (CFRD). Arq. Bras. Endocrinol. Metabol. 2011, 55, 613–621. [Google Scholar] [CrossRef]
- Dif, F.; Marty, C.; Baudoin, C.; de Vernejoul, M.-C.; Levi, G. Severe osteopenia in CFTR-null mice. Bone 2004, 35, 595–603. [Google Scholar] [CrossRef]
- Paradis, J.; Wilke, M.; Haston, C.K. Osteopenia in Cftr-deltaF508 mice. J. Cyst Fibros. 2010, 9, 239–245. [Google Scholar] [CrossRef]
- Guo, J.H.; Chen, H.; Ruan, Y.C.; Zhang, X.L.; Zhang, X.H.; Fok, K.L.; Tsang, L.L.; Yu, M.K.; Huang, W.Q.; Sun, X.; et al. Glucose-induced electrical activities and insulin secretion in pancreatic islet β-cells are modulated by CFTR. Nat. Commun. 2014, 5, 4420. [Google Scholar] [CrossRef]
- Tandon, O.P.; Chintala, R. Hypothalamo-pituitary-gonadal axis in control of female reproductive cycle. Indian J. Physiol. Pharmacol. 2001, 45, 395–407. [Google Scholar]
- Mulberg, A.E.; Resta, L.P.; Wiedner, E.B.; Altschuler, S.M.; Jefferson, D.M.; Broussard, D.L. Expression and localization of the cystic fibrosis transmembrane conductance regulator mRNA and its protein in rat brain. J. Clin. Investig. 1995, 96, 646–652. [Google Scholar] [CrossRef]
- Johannesson, M.; Bogdanovic, N.; Nordqvist, A.C.; Hjelte, L.; Schalling, M. Cystic fibrosis mRNA expression in rat brain: Cerebral cortex and medial preoptic area. Neuroreport 1997, 8, 535–539. [Google Scholar] [CrossRef]
- Mulberg, A.E.; Weyler, R.T.; Altschuler, S.M.; Hyde, T.M. Cystic fibrosis transmembrane conductance regulator expression in human hypothalamus. Neuroreport 1998, 9, 141–144. [Google Scholar] [CrossRef]
- Rogan, M.P.; Reznikov, L.R.; Pezzulo, A.A.; Gansemer, N.D.; Samuel, M.; Prather, R.S.; Zabner, J.; Fredericks, D.C.; McCray, P.B.; Welsh, M.J.; et al. Pigs and humans with cystic fibrosis have reduced insulin-like growth factor 1 (IGF1) levels at birth. Proc. Natl. Acad. Sci. USA 2010, 107, 20571–20575. [Google Scholar] [CrossRef]
- Weyler, R.T.; Yurko-Mauro, K.A.; Rubenstein, R.; Kollen, W.J.; Reenstra, W.; Altschuler, S.M.; Egan, M.; Mulberg, A.E. CFTR is functionally active in GnRH-expressing GT1-7 hypothalamic neurons. Am. J. Physiol. 1999, 277, C563–C571. [Google Scholar] [CrossRef]
- Shwachman, H.; Kulczycki, L.L.; Khaw, K.T. A report on sixty-five patients over 17 years of age. Pediatrics 1965, 36, 689–699. [Google Scholar] [CrossRef]
- Stern, R.C.; Boat, T.F.; Doershuk, C.F.; Tucker, A.S.; Primiano, F.P.; Matthews, L.W. Course of cystic fibrosis in 95 patients. J. Pediatr. 1976, 89, 406–411. [Google Scholar] [CrossRef]
- Umławska, W.; Sands, D.; Zielińska, A. Age of menarche in girls with cystic fibrosis. Folia Histochem. Cytobiol. 2010, 48, 185–190. [Google Scholar] [CrossRef]
- Goldsweig, B.; Kaminski, B.; Sidhaye, A.; Blackman, S.M.; Kelly, A. Puberty in cystic fibrosis. J. Cyst. Fibros. 2019, 18, S88–S94. [Google Scholar] [CrossRef] [PubMed]
- Reznikov, L.R. Cystic Fibrosis and the Nervous System. Chest 2017, 151, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Cupello, A. Neuronal transmembrane chloride electrochemical gradient: A key player in GABA A receptor activation physiological effect. Amino Acids 2003, 24, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Maitra, R.; Sivashanmugam, P.; Warner, K. A Rapid membrane potential assay to monitor CFTR function and inhibition. SLAS Discov. Adv. Sci. Drug Discov. 2013, 18, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Ostroumov, A.; Simonetti, M.; Nistri, A. Cystic fibrosis transmembrane conductance regulator modulates synaptic chloride ho-meostasis in motoneurons of the rat spinal cord during neonatal development. Dev. Neurobiol. 2011, 71, 253–268. [Google Scholar] [CrossRef]
- Akita, T.; Fukuda, A. Intracellular Cl− dysregulation causing and caused by pathogenic neuronal activity. Pflügers Arch. -Eur. J. Physiol. 2020, 472, 977–987. [Google Scholar] [CrossRef]
- Raheja, G.; Gill, K.D. Calcium homeostasis and dichlorvos induced neurotoxicity in rat brain. Mol. Cell. Biochem. 2002, 232, 13–18. [Google Scholar] [CrossRef]
- Chen, T.-W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 2013, 499, 295–300. [Google Scholar] [CrossRef]
- Zhang, L.-W.; Bi, A.-L.; Li, Q.; Bi, H.-S. Application of fiber photometry in neuroscience research. Sheng Li Xue Bao Acta Physiol. Sin. 2021, 73, 306–314. [Google Scholar]
- Parnet, P.; Lenoir, V.; Palkovits, M.; Kerdelhué, B. Estrous cycle variations in gonadotropin-releasing hormone, substance P and beta-endorphin contents in the median eminence, the arcuate nucleus and the medial preoptic nucleus in the rat: A detailed analysis of proestrus changes. J. Neuroendocr. 1990, 2, 291–296. [Google Scholar] [CrossRef]
- Clarke, I.J.; Thomas, G.B.; Yao, B.; Cummins, J.T. GnRH secretion throughout the ovine estrous cycle. Neuroendocrinology 1987, 46, 82–88. [Google Scholar] [CrossRef]
- Jin, R.; Hodges, C.A.; Drumm, M.L.; Palmert, M.R. The cystic fibrosis transmembrane conductance regulator (Cftr) modulates the timing of puberty in mice. J. Med. Genet. 2006, 43, e29. [Google Scholar] [CrossRef]
- Hughan, K.S.; Daley, T.; Rayas, M.S.; Kelly, A.; Roe, A. Female reproductive health in cystic fibrosis. J. Cyst. Fibros. 2019, 18, S95–S104. [Google Scholar] [CrossRef]
- Habibullah, M.M. The role of CFTR channel in female infertility. Hum. Fertil. 2022, 2022, 1–10. [Google Scholar] [CrossRef]
- Chan, H.C.; Ruan, Y.C.; He, Q.; Chen, M.H.; Chen, H.; Xu, W.M.; Chen, W.Y.; Xie, C.; Zhang, X.H.; Zhou, Z. The cystic fibrosis transmembrane conductance regulator in reproductive health and disease. J. Physiol. 2009, 587, 2187–2195. [Google Scholar] [CrossRef]
- Chan, H.C.; Chen, H.; Ruan, Y.; Sun, T. Physiology and pathophysiology of the epithelial barrier of the female reproductive tract: Role of ion channels. Adv. Exp. Med. Biol. 2012, 763, 193–217. [Google Scholar]
- Chen, H.; Guo, J.H.; Lu, Y.C.; Ding, G.L.; Yu, M.K.; Tsang, L.L.; Fok, K.L.; Liu, X.M.; Zhang, X.H.; Chung, Y.W.; et al. Impaired CFTR-dependent amplification of FSH-stimulated estrogen production in cystic fibrosis and PCOS. J. Clin. Endocrinol. Metab. 2012, 97, 923–932. [Google Scholar] [CrossRef]
- Ting, J.T.; Lee, B.R.; Chong, P.; Soler-Llavina, G.; Cobbs, C.; Koch, C.; Zeng, H.; Lein, E. Preparation of Acute Brain Slices Using an Optimized N-Methyl-D-glucamine Protective Recovery Method. J. Vis. Exp. 2018, 132, 53825. [Google Scholar]
- Byers, S.L.; Wiles, M.V.; Dunn, S.L.; Taft, R.A. Mouse estrous cycle identification tool and images. PLoS ONE 2012, 7, e35538. [Google Scholar] [CrossRef]
- Felicio, L.S.; Nelson, J.F.; Finch, C.E. Longitudinal studies of estrous cyclicity in aging C57BL/6J mice: II. cessation of cyclicity and the duration of persistent vaginal cornification 1. Biol. Reprod. 1984, 31, 446–453. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Que, Y.; Chen, J.; Sun, L.; Guo, J.; Ruan, Y.C. CFTR Modulates Hypothalamic Neuron Excitability to Maintain Female Cycle. Int. J. Mol. Sci. 2023, 24, 12572. https://doi.org/10.3390/ijms241612572
Wu Y, Que Y, Chen J, Sun L, Guo J, Ruan YC. CFTR Modulates Hypothalamic Neuron Excitability to Maintain Female Cycle. International Journal of Molecular Sciences. 2023; 24(16):12572. https://doi.org/10.3390/ijms241612572
Chicago/Turabian StyleWu, Yong, Yanting Que, Junjiang Chen, Lei Sun, Jinghui Guo, and Ye Chun Ruan. 2023. "CFTR Modulates Hypothalamic Neuron Excitability to Maintain Female Cycle" International Journal of Molecular Sciences 24, no. 16: 12572. https://doi.org/10.3390/ijms241612572
APA StyleWu, Y., Que, Y., Chen, J., Sun, L., Guo, J., & Ruan, Y. C. (2023). CFTR Modulates Hypothalamic Neuron Excitability to Maintain Female Cycle. International Journal of Molecular Sciences, 24(16), 12572. https://doi.org/10.3390/ijms241612572