
Combined Salivary Proteome Profiling and Machine Learning Analysis Provides Insight into Molecular Signature for Autoimmune Liver Diseases Classification

 , , , , , , ,
, , , , , , ,  , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Characteristics of the Participants
2.2. Saliva Sampling
2.3. RP-nanoHPLC-High Resolution ESI-MS and MS/MS Analysis and Protein Identification
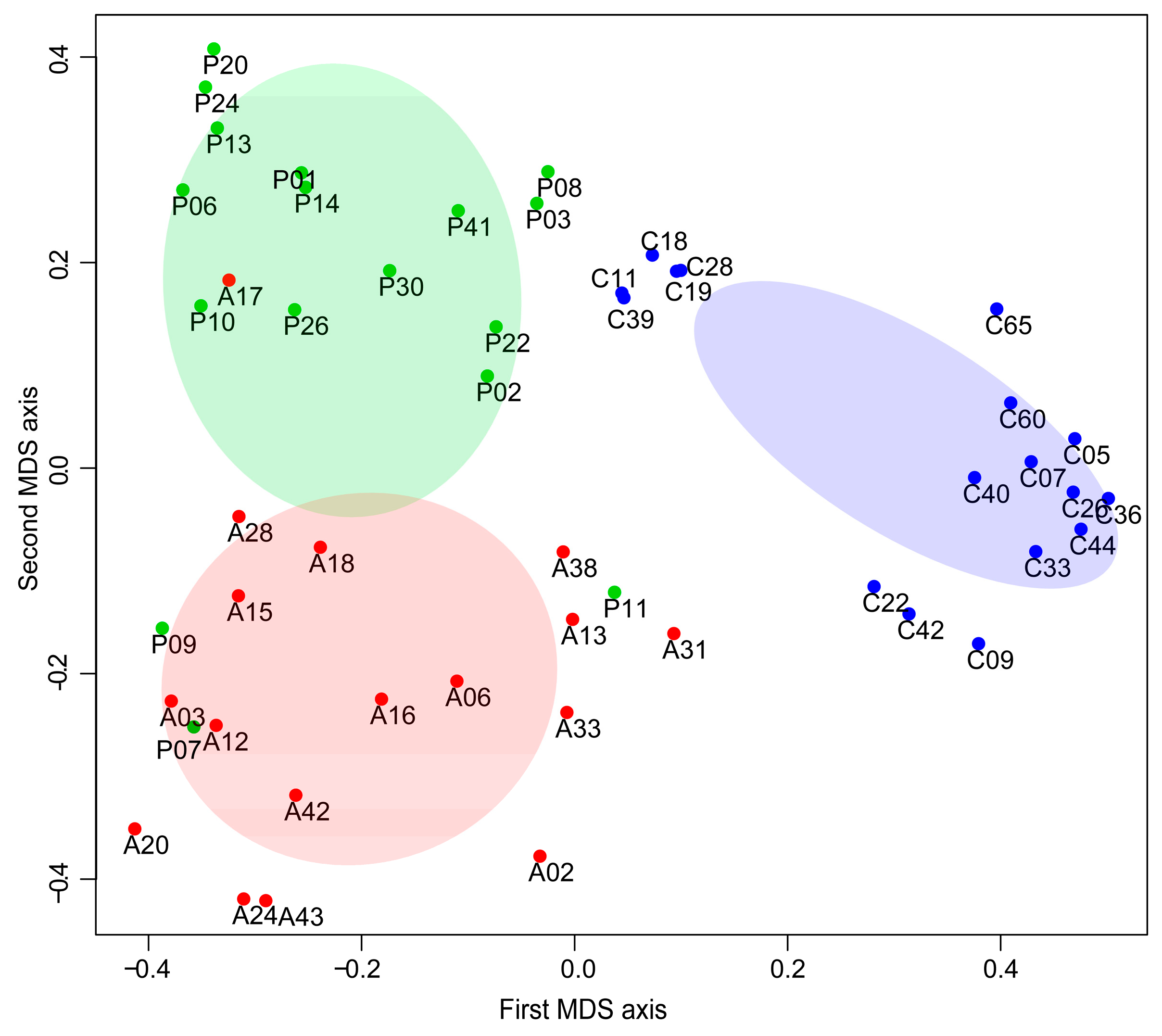
2.4. Subjecs Classification with Random Forest (RF) Analysis
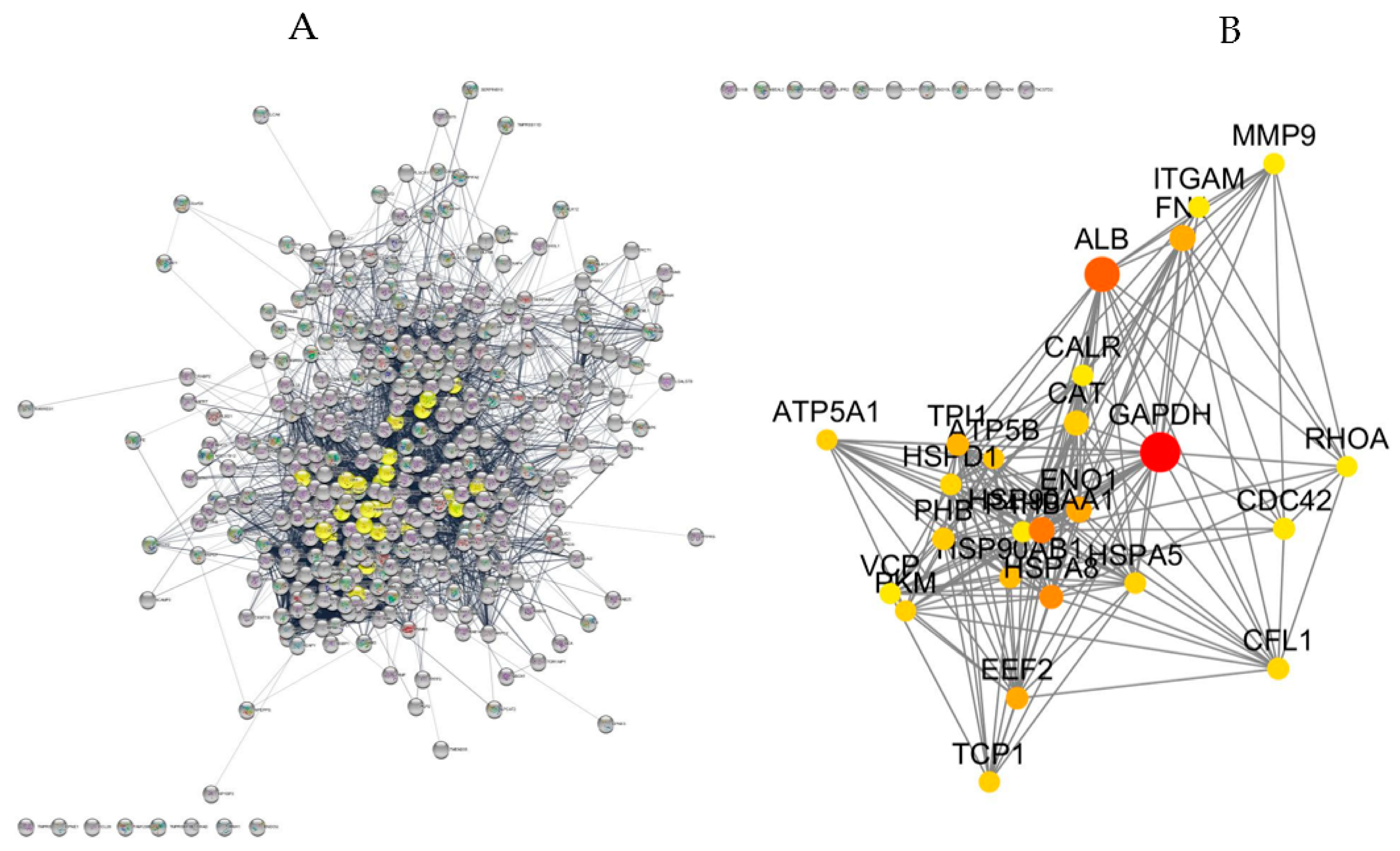
2.5. Protein-Protein Interaction Network, Topological Analysis and Pathway Enrichment
2.5.1. Proteins Commonly Found among AIHp, PBCp and HCs
2.5.2. Proteins with Varied Levels among AIHp, PBCp and HCs
3. Discussion
3.1. Topology-Based Functional Enrichment Analysis
3.2. Dysregulated Proteins in AIH and PBC Are Mainly Involved in Liver Fibrosis
3.3. Machine Learning Analysis of Proteomics Data
3.4. Study Limitation
4. Materials and Methods
4.1. Ethical Statement
4.2. Study Subjects and Clinical Studies
4.3. Sample Collection, Treatment and Acid-Insoluble Proteins Solubilization
4.4. SDS-PAGE, Bands Excision and Enzymatic Digestion
4.5. Nano-RP-HPLC-High Resolution ESI-MS/MS Analysis
4.6. Protein Identification and Quantitation
4.7. Data Analysis
4.8. Protein-Protein Interaction Network, Topological Analysis and Pathway Enrichment
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heneghan, M.A.; Yeoman, A.D.; Verma, S.; Smith, A.D.; Longhi, M.S. Autoimmune hepatitis. Lancet 2013, 382, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.P.; Czaja, A.J.; Gorham, J.D.; Krawitt, E.L.; Mieli-Vergani, G.; Vergani, D.; Vierling, J.M. American Association for the Study of Liver Diseases. Diagnosis and management of autoimmune hepatitis. Hepatology 2010, 51, 2193–2213. [Google Scholar] [CrossRef] [PubMed]
- Mack, C.L.; Adams, D.; Assis, D.N.; Kerkar, N.; Manns, M.P.; Mayo, M.J.; Vierling, J.M.; Alsawas, M.; Murad, M.H.; Czaja, A.J. Diagnosis and Management of Autoimmune Hepatitis in Adults and Children: 2019 Practice Guidance and Guidelines From the American Association for the Study of Liver Diseases. Hepatology 2020, 72, 671–722. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.P.; Lohse, A.W.; Vergani, D. Autoimmune hepatitis—Update 2015. J. Hepatol. 2015, 62, S100–S111. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. EASL Clinical Practice Guidelines: The diagnosis and management of patients with primary biliary cholangitis. J. Hepatol. 2017, 67, 145–172. [Google Scholar] [CrossRef] [PubMed]
- Lleo, A.; Leung, P.S.C.; Hirschfield, G.M.; Gershwin, E.M. The Pathogenesis of Primary Biliary Cholangitis: A Comprehensive Review. Semin. Liver Dis. 2020, 40, 34–48. [Google Scholar] [CrossRef]
- Chazouillères, O.; Wendum, D.; Serfaty, L.; Montembault, S.; Rosmorduc, O.; Poupon, R. Primary biliary cirrhosis-autoimmune hepatitis overlap syndrome: Clinical features and response to therapy. Hepatology 1998, 28, 296–301. [Google Scholar] [CrossRef]
- Czaja, A.J. Diagnosis and management of the overlap syndromes of autoimmune hepatitis. Can. J. Gastroenterol. 2013, 27, 417–423. [Google Scholar] [CrossRef]
- Schulz, L.; Sebode, M.; Weidemann, S.A.; Lohse, A.W. Variant syndromes of primary biliary cholangitis. Best Pract. Res. Clin. Gastroenterol. 2018, 34, 55–61. [Google Scholar] [CrossRef]
- Schiulthiß, C.; Steinmann, S.; Willscher, E.; Paschold, L.; Lohse, A.W.; Binder, M. Immune signatures in variant syndromes of primary biliary cholangitis and autoimmune hepatitis. Hepatol. Commun. 2023, 7, e0123. [Google Scholar] [CrossRef]
- Floreani, A.; Franceschet, I.; Cazzagon, N.; Spinazzè, A.; Buja, A.; Furlan, P.; Baldo, V.; Gershwin, M.E. Extrahepatic autoimmune conditions associated with primary biliary cirrhosis. Clin. Rev. Allergy Immunol. 2015, 48, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.W.; Yeong, T.; Lawrence, D.; Yeoman, A.D.; Verma, S.; Heneghan, M.A. Concurrent extrahepatic autoimmunity in autoimmune hepatitis: Implications for diagnosis, clinical course and long-term outcomes. Liver Int. 2017, 37, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Vierling, J.M. Diagnosis and treatment of autoimmune hepatitis. Curr. Gastroenterol. Rep. 2012, 14, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Norman, G.L.; Shums, Z.; Worman, H.J.; Krawitt, E.L.; Bizzaro, N.; Vergani, D.; Bogdanos, D.P.; Dalekos, G.N.; Milkiewicz, P.; et al. PBC screen: An IgG/IgA dual isotype ELISA detecting multiple mitochondrial and nuclear autoantibodies specific for primary biliary cirrhosis. J. Autoimmun. 2010, 35, 436–442. [Google Scholar] [CrossRef]
- Granito, A.; Muratori, L.; Muratori, P.; Pappas, G.; Guidi, M.; Cassani, F.; Volta, U.; Ferri, A.; Lenzi, M.; Bianchi, F.B. Antibodies to filamentous actin (F-actin) in type 1 autoimmune hepatitis. J. Clin. Pathol. 2006, 59, 280–284. [Google Scholar] [CrossRef]
- Gerosa, C.; Fanos, V.; Fanni, D.; Van Eyken, P.; Ambu, R.; Nemolato, S.; Floris, G.; Iacovidou, N.; Gibo, Y.; Nurchi, A.M.; et al. Liver Biopsy Interpretation in the Differential Diagnosis of Autoimmune Liver Disease in Children. J. Pediatr. Neonat. Individual. Med. 2013, 2, e020229. [Google Scholar] [CrossRef]
- Tan, D.; Goodman, Z.D. Liver biopsy in primary biliary cholangitis: Indications and interpretation. Clin. Liver Dis. 2018, 22, 579–588. [Google Scholar] [CrossRef]
- Contini, C.; Serrao, S.; Manconi, B.; Olianas, A.; Iavarone, F.; Bizzarro, A.; Masullo, C.; Castagnola, M.; Messana, I.; Diaz, G.; et al. Salivary Proteomics Reveals Significant Changes in Relation to Alzheimer’s Disease and Aging. J. Alzheimers Dis. 2022, 89, 605–622. [Google Scholar] [CrossRef]
- Sanna, M.; Firinu, D.; Manconi, P.E.; Pisanu, M.; Murgia, G.; Piras, V.; Castagnola, M.; Messana, I.; Del Giacco, S.; Cabras, T. The salivary proteome profile in patients affected by SAPHO syndrome characterized by a top-down RP-HPLC-ESI-MS platform. Mol. Biosyst. 2015, 11, 1552–1562. [Google Scholar] [CrossRef]
- Cabras, T.; Sanna, M.; Manconi, B.; Fanni, D.; Demelia, L.; Sorbello, O.; Iavarone, F.; Castagnola, M.; Faa, G.; Messana, I. Proteomic investigation of whole saliva in Wilson’s disease. J. Proteom. 2015, 128, 154–163. [Google Scholar] [CrossRef]
- Manconi, B.; Liori, B.; Cabras, T.; Vincenzoni, F.; Iavarone, F.; Lorefice, L.; Cocco, E.; Castagnola, M.; Messana, I.; Olianas, A. Top-down proteomic profiling of human saliva in multiple sclerosis patients. J. Proteom. 2018, 187, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Boroumand, M.; Olianas, A.; Cabras, T.; Manconi, B.; Fanni, D.; Faa, G.; Desiderio, C.; Messana, I.; Castagnola, M. Saliva, a bodily fluid with recognized and potential diagnostic applications. J. Sep. Sci. 2021, 44, 3677–3690. [Google Scholar] [CrossRef] [PubMed]
- Rao, B.; Lou, J.; Lu, H.; Liang, H.; Li, J.; Zhou, H.; Fan, Y.; Zhang, H.; Sun, Y.; Zou, Y.; et al. Oral Microbiome Characteristics in Patients with Autoimmune Hepatitis. Front. Cell Infect. Microbiol. 2021, 11, 656674. [Google Scholar] [CrossRef] [PubMed]
- Messana, I.; Cabras, T.; Pisano, E.; Sanna, M.T.; Olianas, A.; Manconi, B.; Pellegrini, M.; Paludetti, G.; Scarano, E.; Fiorita, A.; et al. Trafficking and postsecretory events responsible for the formation of secreted human salivary peptides. Mol. Cell Proteom. 2008, 7, 911–926. [Google Scholar] [CrossRef]
- Ghallab, N.A. Diagnostic potential and future directions of biomarkers in gingival crevicular fluid and saliva of periodontal diseases: Review of the current evidence. Arch. Oral Biol. 2018, 87, 115–124. [Google Scholar] [CrossRef]
- Iavarone, F.; Olianas, A.; Patini, R.; Gallenzi, P.; Di Tonno, L.; Desiderio, C.; Cabras, T.; Manconi, B.; Vincenzoni, F.; Cordaro, M.; et al. Top down proteomic analysis of gingival crevicular fluid in deciduous, exfoliating and permanent teeth in children. J. Proteom. 2020, 226, 103890. [Google Scholar] [CrossRef] [PubMed]
- Grassl, N.; Kulak, N.A.; Pichler, G.; Geyer, P.E.; Jung, J.; Schubert, S.; Sinitcyn, P.; Cox, J.; Mann, M. Ultra-deep and quantitative saliva proteome reveals dynamics of the oral microbiome. Genome Med. 2016, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Olianas, A.; Guadalupi, G.; Cabras, T.; Contini, C.; Serrao, S.; Iavarone, F.; Castagnola, M.; Messana, I.; Onali, S.; Chessa, L.; et al. Top-Down Proteomics Detection of Potential Salivary Biomarkers for Autoimmune Liver Diseases Classification. Int. J. Mol. Sci. 2023, 24, 959. [Google Scholar] [CrossRef]
- Joy, M.P.; Brock, A.; Ingber, D.E.; Huang, S. High-betweenness proteins in the yeast protein interaction network. J. Biomed. Biotechnol. 2005, 2005, 96–103. [Google Scholar] [CrossRef]
- Nithya, C.; Kiran, M.; Nagarajaram, H.A. Dissection of hubs and bottlenecks in a protein-protein interaction network. Comput. Biol. Chem. 2023, 102, 107802. [Google Scholar] [CrossRef]
- Shibuya, A.; Ikewaki, N. High serum glyceraldehyde-3-phosphate dehydrogenase levels in patients with liver cirrhosis. Hepatol. Res. 2002, 22, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Domenicali, M.; Baldassarre, M.; Giannone, F.A.; Naldi, M.; Mastroroberto, M.; Biselli, M.; Laggetta, M.; Patrono, D.; Bertucci, C.; Bernardi, M.; et al. Posttranscriptional changes of serum albumin: Clinical and prognostic significance in hospitalized patients with cirrhosis. Hepatology 2014, 60, 1851–1860. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, G.; Garcia-Tsao, G.; Pagliaro, L. Natural history and prognostic indicators of survival in cirrhosis: A systematic review of 118 studies. J. Hepatol. 2006, 44, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.R.; Verdelho Machado, M. New Insights About Albumin and Liver Disease. Ann. Hepatol. 2018, 17, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y. Discovering disease-genes by topological features in human protein-protein interaction network. Bioinformatics 2006, 22, 2800–2805. [Google Scholar] [CrossRef] [PubMed]
- Gieseck, R.L., III; Ramalingam, T.R.; Hart, K.M.; Vannella, K.M.; Cantu, D.A.; Lu, W.Y.; Ferreira-González, S.; Forbes, S.J.; Vallier, L.; Wynn, T.A. Interleukin-13 Activates Distinct Cellular Pathways Leading to Ductular Reaction, Steatosis, and Fibrosis. Immunity 2016, 45, 145–158. [Google Scholar] [CrossRef]
- Webb, G.J.; Siminovitch, K.A.; Hirschfield, G.M. The immunogenetics of primary biliary cirrhosis: A comprehensive review. J. Autoimmun. 2015, 64, 42–52. [Google Scholar] [CrossRef]
- Liaskou, E.; Patel, S.R.; Webb, G.; Bagkou Dimakou, D.; Akiror, S.; Krishna, M.; Mells, G.; Jones, D.E.; Bowman, S.J.; Barone, F.; et al. Increased sensitivity of Treg cells from patients with PBC to low dose IL-12 drives their differentiation into IFN-γ secreting cells. J. Autoimmun. 2018, 94, 143–155. [Google Scholar] [CrossRef]
- Povero, D.; Busletta, C.; Novo, E.; di Bonzo, L.V.; Cannito, S.; Paternostro, C.; Parola, M. Liver fibrosis: A dynamic and potentially reversible process. Histol. Histopathol. 2010, 25, 1075–1091. [Google Scholar] [CrossRef]
- Vinken, M.; Papeleu, P.; Snykers, S.; De Rop, E.; Henkens, T.; Chipman, J.K.; Rogiers, V.; Vanhaecke, T. Involvement of cell junctions in hepatocyte culture functionality. Crit. Rev. Toxicol. 2006, 36, 299–318. [Google Scholar] [CrossRef]
- Zhou, L.; Pradhan-Sundd, T.; Poddar, M.; Singh, S.; Kikuchi, A.; Stolz, D.B.; Shou, W.; Li, Z.; Nejak-Bowen, K.N.; Monga, S.P. Mice with Hepatic Loss of the Desmosomal Protein γ-Catenin Are Prone to Cholestatic Injury and Chemical Carcinogenesis. Am. J. Pathol. 2015, 185, 3274–3289. [Google Scholar] [CrossRef]
- Nakagawa, H.; Hikiba, Y.; Hirata, Y.; Font-Burgada, J.; Sakamoto, K.; Hayakawa, Y.; Taniguchi, K.; Umemura, A.; Kinoshita, H.; Sakitani, K.; et al. Loss of liver E-cadherin induces sclerosing cholangitis and promotes carcinogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Dubash, A.D.; Kam, C.Y.; Aguado, B.A.; Patel, D.M.; Delmar, M.; Shea, L.D.; Green, K.J. Plakophilin-2 loss promotes TGF-β1/p38 MAPK-dependent fibrotic gene expression in cardiomyocytes. J. Cell Biol. 2016, 212, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Radwan, N.A.; Ahmed, N.S. The diagnostic value of arginase-1 immunostaining in differentiating hepatocellular carcinoma from metastatic carcinoma and cholangiocarcinoma as compared to HepPar-1. Diagn. Pathol. 2012, 7, 149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wu, Z.; Salas, S.S.; Aguilar, M.M.; Trillos-Almanza, M.C.; Buist-Homan, M.; Moshage, H. Arginase 1 expression is increased during hepatic stellate cell activation and facilitates collagen synthesis. J. Cell Biochem. 2023, 124, 808–817. [Google Scholar] [CrossRef]
- Ogino, K.; Kubo, M.; Takahashi, H.; Zhang, R.; Zou, Y.; Fujikura, Y. Anti-inflammatory effect of arginase inhibitor and corticosteroid on airway allergic reactions in a Dermatophogoides farinae-induced NC/Nga mouse model. Inflammation 2013, 36, 141–151. [Google Scholar] [CrossRef]
- Hijazi, N.; Rockey, D.C.; Shi, Z. The cellular microenvironment and cytoskeletal actin dynamics in liver fibrogenesis. Biocell 2022, 46, 2003–2007. [Google Scholar] [CrossRef]
- Xu, Y.; Duan, J.; Ji, W.; Liu, C.; Li, X.; Wu, Q.; Gao, C.; Su, C. A novel matrine derivative, WM130, inhibits activation and movement of human hepatic stellate LX-2 cells by targeting cofilin 1. Cytotechnology 2022, 74, 613–622. [Google Scholar] [CrossRef]
- Yao, B.; Li, Y.; Chen, T.; Niu, Y.; Wang, Y.; Yang, Y.; Wei, X.; Liu, Q.; Tu, K. Hypoxia-induced cofilin 1 promotes hepatocellular carcinoma progression by regulating the PLD1/AKT pathway. Clin. Transl. Med. 2021, 11, e366. [Google Scholar] [CrossRef]
- Witke, W.; Sharpe, A.H.; Hartwig, J.H.; Azuma, T.; Stossel, T.P.; Kwiatkowski, D.J. Hemostatic, inflammatory, and fibroblast responses are blunted in mice lacking gelsolin. Cell 1995, 81, 41–51. [Google Scholar] [CrossRef]
- Aigelsreiter, A.; Janig, E.; Sostaric, J.; Pichler, M.; Unterthor, D.; Halasz, J.; Lackner, C.; Zatloukal, K.; Denk, H. Clusterin expression in cholestasis, hepatocellular carcinoma and liver fibrosis. Histopathology 2009, 54, 561–570. [Google Scholar] [CrossRef]
- Seo, H.Y.; Lee, S.H.; Lee, J.H.; Kang, Y.N.; Choi, Y.K.; Hwang, J.S.; Park, K.G.; Jang, B.K.; Kim, M.K. Clusterin Attenuates Hepatic Fibrosis by Inhibiting Hepatic Stellate Cell Activation and Downregulating the Smad3 Signaling Pathway. Cells 2019, 8, 1442. [Google Scholar] [CrossRef]
- Jimenez-Rivera, C.; Jolin-Dahel, K.S.; Fortinsky, K.J.; Gozdyra, P.; Benchimol, E.I. International incidence and outcomes of biliary atresia. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 344–354. [Google Scholar] [CrossRef]
- Sahin, M.; Yetim, A.; Ates, F. The role of tacstd-2 level in hepatitis C patients (controlled clinical research). Libyan J. Med. 2019, 14, 1622363. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Miotti, S.; Stella, M.; Klein, C.E.; Fornaro, M.; Menard, S.; Colnaghi, M.I. Biochemical characterization of Trop-2, a cell surface molecule expressed by human carcinomas: Formal proof that the monoclonal antibodies T16 and MOv-16 recognize Trop-2. Hybridoma 1992, 11, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Sekhar, V.; Pollicino, T.; Diaz, G.; Engle, R.E.; Alayli, F.; Melis, M.; Kabat, J.; Tice, A.; Pomerenke, A.; Altan-Bonnet, N.; et al. Infection with hepatitis C virus depends on TACSTD2, a regulator of claudin-1 and occludin highly downregulated in hepatocellular carcinoma. PLoS Pathog. 2018, 14, e1006916. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.J.; Shen, S.L.; Li, S.Q.; Hua, Y.P.; Hu, W.J.; Guo, B.; Peng, B.G. Hornerin promotes tumor progression and is associated with poor prognosis in hepatocellular carcinoma. BMC Cancer 2018, 18, 815. [Google Scholar] [CrossRef] [PubMed]
- Henry, J.; Hsu, C.Y.; Haftek, M.; Nachat, R.; de Koning, H.D.; Gardinal-Galera, I.; Hitomi, K.; Balica, S.; Jean-Decoster, C.; Schmitt, A.M.; et al. Hornerin is a component of the epidermal cornified cell envelopes. FASEB J. 2011, 25, 1567–1576. [Google Scholar] [CrossRef]
- Arteel, G.E.; Naba, A. The liver matrisome—Looking beyond collagens. JHEP Rep. 2020, 2, 100115. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Betrapally, N.S.; Hylemon, P.B.; Heuman, D.M.; Daita, K.; White, M.B.; Unser, A.; Thacker, L.R.; Sanyal, A.J.; Kang, D.J.; et al. Salivary microbiota reflects changes in gut microbiota in cirrhosis with hepatic encephalopathy. Hepatology 2015, 62, 1260–1271. [Google Scholar] [CrossRef]
- Elghannam, M.T.; Hassanien, M.H.; Ameen, Y.A.; Turky, E.A.; Elattar, G.M.; ElRay, A.A.; Eltalkawy, M.D. Oral microbiota and liver diseases. Clin. Nutr. ESPEN 2023, 54, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Beamer, L.J.; Carroll, S.F.; Eisenberg, D. The BPI/LBP family of proteins: A structural analysis of conserved regions. Protein Sci. 1998, 7, 906–914. [Google Scholar] [CrossRef]
- Lv, L.; Jiang, H.; Chen, X.; Wang, Q.; Wang, K.; Ye, J.; Li, Y.; Fang, D.; Lu, Y.; Yang, L.; et al. The Salivary Microbiota of Patients With Primary Biliary Cholangitis Is Distinctive and Pathogenic. Front. Immunol. 2021, 12, 713647. [Google Scholar] [CrossRef] [PubMed]
- Nandula, S.R.; Huxford, I.; Wheeler, T.T.; Aparicio, C.; Gorr, S.U. The parotid secretory protein BPIFA2 is a salivary surfactant that affects lipopolysaccharide action. Exp. Physiol. 2020, 105, 1280–1292. [Google Scholar] [CrossRef] [PubMed]
- Sallustio, F.; Stasi, A.; Curci, C.; Divella, C.; Picerno, A.; Franzin, R.; De Palma, G.; Rutigliano, M.; Lucarelli, G.; Battaglia, M.; et al. Renal progenitor cells revert LPS-induced endothelial-to-mesenchymal transition by secreting CXCL6, SAA4, and BPIFA2 antiseptic peptides. FASEB J. 2019, 33, 10753–10766. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Komori, A.; Nakamura, M.; Takii, Y.; Kamihira, T.; Shimoda, S.; Mori, T.; Fujiwara, S.; Koyabu, M.; Taniguchi, K.; et al. Human intrahepatic biliary epithelial cells function in innate immunity by producing IL-6 and IL-8 via the TLR4-NF-kappaB and -MAPK signaling pathways. Liver Int. 2006, 26, 467–476. [Google Scholar] [CrossRef]
- Bazil, V.; Strominger, J.L. Shedding as a mechanism of down-modulation of CD14 on stimulated human monocytes. J. Immunol. 1991, 147, 1567–1574. [Google Scholar] [CrossRef]
- Umemura, T.; Sekiguchi, T.; Joshita, S.; Yamazaki, T.; Fujimori, N.; Shibata, S.; Ichikawa, Y.; Komatsu, M.; Matsumoto, A.; Shums, Z.; et al. Association between serum soluble CD14 and IL-8 levels and clinical outcome in primary biliary cholangitis. Liver Int. 2017, 37, 897–905. [Google Scholar] [CrossRef]
- Karlmark, K.R.; Tacke, F.; Dunay, I.R. Monocytes in health and disease—Minireview. Eur. J. Microbiol. Immunol. 2012, 2, 97–102. [Google Scholar] [CrossRef]
- Boberg, K.M. Prevalence and epidemiology of autoimmune hepatitis. Clin. Liver Dis. 2002, 6, 635–647. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape StringApp: Network Analysis and Visualization of Proteomics Data. J. Proteome Res. 2019, 18, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Assenov, Y.; Ramírez, F.; Schelhorn, S.E.; Lengauer, T.; Albrecht, M. Computing topological parameters of biological networks. Bioinformatics 2008, 24, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Shen, H.; Zhang, L.G.; Liu, J.; Cao, X.G.; Yao, A.L.; Kang, S.S.; Gao, W.X.; Han, H.; Cao, F.H.; et al. Construction and analysis of protein-protein interaction networks based on proteomics data of prostate cancer. Int. J. Mol. Med. 2016, 37, 1576–1586. [Google Scholar] [CrossRef]
- Yu, H.; Kim, P.M.; Sprecher, E.; Trifonov, V.; Gerstein, M. The importance of bottlenecks in protein networks: Correlation with gene essentiality and expression dynamics. PLoS Comput. Biol. 2007, 3, e59. [Google Scholar] [CrossRef]
- Benjamini, Y.; and Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Statist. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Kursa, M.B.; Jankowski, A.; Rudnicki, W.R. Boruta—A system for feature selection. Fundam. Inform. 2010, 101, 271–285. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | AIHp | PBCp | |
|---|---|---|---|
| Age, average (range) | Years | 60.6 (40–83) | 63.0 (52–83) |
| Gender, n (%) | Female | 15 (88.3%) | 17 (100%) |
| BMI, average (range) | Kg/m2 | 26.46 (17.58–36.13) | 24.63 (19.33–30.27) |
| Cirrhosis, n (%) | 4 (23.5%) | 4 (23.5%) | |
| Histological stage n (%) | I–II | 5 (29.4%) | 11 (64.7%) |
| III–IV | 12 (70.6%) | 6 (35.3%) | |
| Positivity to autoantibodies, n (%) | ANA | 12 (70.6%) | 8 (47.1%) |
| SMA | 10 (58.8%) | 3 (17.6%) | |
| LKM | 1 (5.9%) | 2 (11.8%) | |
| AMA | 0 (0%) | 16 (94.1%) | |
| AST, median (range) | IU/L | 25.0 (14–57) | 28.0 (17–65) |
| ALT, median (range) | IU/L | 20.0 (8–46) | 28.0 (17–133) |
| GGT, median (range) | IU/L | 30.0 (0–138) | 68.0 (14–452) |
| ALP, median (range) | IU/L | 64.5 (28–216) | 118.0 (65–415) |
| IgG, median (range) | g/dL | 1.4 (0.69–2.51) | 1.4 (1.1–2.3) |
| Albumine, median (range) | g/dL | 3.9 (3.24–4.76) | 3.9 (2.8–4.4) |
| Prothrombin time, median (range) | INR | 0.93 (0.92–1.03) | 1.0 (0.91–1.61) |
| TB, median (range) | mg/dL | 0.69 (0.3–1.46) | 0.61 (0.28–2.95) |
| Platelets, median (range) | 109/L | 199.0 (91–402) | 223 (46–418) |
| Pharmacological treatment (% treated) | AZA (only or + UDCA and/or Steroids) | 58.8% | |
| Steroids | 11.8% | ||
| UDCA (only or + steroids) | 23.5% | 100% | |
| Naïve | 5.9% |
| Proteins | HCs vs. AIHp | HCs vs. PBCp | AIHp vs. PBCp | HCs vs. AIHp vs. PBCp | |
|---|---|---|---|---|---|
| Uniprot Code | Description | Mann Whitney | Mann Whitney | Mann Whitney | Kruskal Wallis |
| p-Value | p-Value | p-Value | p-Value | ||
| P05089 | Arginase * | 0.0003 AIH < HC | 0.0730 | 0.0061 | 0.0006 |
| P06396 | Gelsolin | 0.2313 | 0.0015 PBC > HC | 0.0653 | 0.0077 |
| P08571 | Monocyte differentiation antigen * | 0.0256 | 0.0001 PBC > HC | 0.1474 | 0.0005 |
| P09758 | Tumor-associated calcium signal transducer 2 | 0.0187 | 0.0010 PBC > HC | 0.5352 | 0.0035 |
| P10909 | Clusterin * | 0.0540 | 0.0002 PBC > HC | 0.0777 | 0.0009 |
| P14923 | Junction plakoglobin | 0.0007 AIH < HC | 0.2313 | 0.0287 | 0.0034 |
| P15924 | Desmoplakin * | 0.0007 AIH < HC | 0.3057 | 0.0177 | 0.0030 |
| P22626 | Heterogeneous nuclear ribonucleoproteins A2/B1 * | 0.0036 | 0.0006 PBC > HC | 0.1899 | 0.0009 |
| P23528 | Cofilin-1 * | 0.0679 | 0.0007 PBC > HC | 0.5177 | 0.0062 |
| P52790 | Hexokinase-3 | 0.0013 AIH < HC | 0.0987 | 0.0819 | 0.0053 |
| Q08554 | Desmocollin-1 * | <0.0001 AIH < HC | 0.0064 | 0.0114 | <0.0001 |
| Q86YZ3 | Hornerin * | <0.0001 AIH < HC | <0.0001 PBC < HC | 0.4824 | <0.0001 |
| Q8N4F0 | BPI fold-containing family B member 2 | 0.0145 | 0.0007 PBC > HC | 0.5629 | 0.0035 |
| Q96DR5 | BPI fold-containing family A member 2 | 0.0007 AIH > HC | 0.0216 | 0.5861 | 0.0044 |
| Predicted Class | |||||
|---|---|---|---|---|---|
| HCs | AIHp | PBCp | OOB Error (%) | ||
| True class | HCs | 17 | 0 | 0 | 0 |
| AIHp | 0 | 16 | 1 | 6 | |
| PBCp | 0 | 2 | 15 | 12 | |
| Term ID | Term | Term Size | Enriched Terms | FDR * | Associated Genes |
|---|---|---|---|---|---|
| Gene Ontology Biological Process | |||||
| GO:0071310 | Cellular response to organic substance | 2369 | 21 | 5.99 × 10−12 | GAPDH|CAT|ATP5B|EEF2|TCP1|PKM|CALR|HSPA5|P4HB|HSP90AA1|FN1|VCP|HSP90AB1|MMP9|HSPD1|CDC42|RHOA|HSPA8|CFL1|ITGAM|PHB |
| GO:0006457 | Protein folding | 213 | 9 | 6.53 × 10−09 | TCP1|CALR|HSPA5|P4HB|HSP90AA1|VCP|HSP90AB1|HSPD1|HSPA8 |
| GO:0046034 | ATP metabolic process | 204 | 8 | 1.12× 10−07 | GAPDH|TPI1|ENO1|ATP5B|PKM|VCP|ATP5A1|HSPA8 |
| GO:0043312 | Neutrophil degranulation | 484 | 10 | 1.42 × 10−07 | CAT|EEF2|PKM|HSP90AA1|VCP|HSP90AB1|MMP9|RHOA|HSPA8|ITGAM |
| GO:0051702 | Interaction with symbiont | 93 | 6 | 7.16 × 10−07 | GAPDH|FN1|HSPD1|HSPA8|CFL1|PHB |
| Reactome pathways | |||||
| HSA-168256 | Immune System | 1956 | 17 | 1.13 × 10−08 | CAT|EEF2|TCP1|PKM|CALR|HSPA5|P4HB|HSP90AA1|FN1|VCP|HSP90AB1|MMP9|CDC42|RHOA|HSPA8|CFL1|ITGAM |
| HSA-3371556 | Cellular response to heat stress | 89 | 5 | 4.23 × 10−05 | HSPA5|HSP90AA1|VCP|HSP90AB1|HSPA8 |
| HSA-5336415 | Uptake and function of diphtheria toxin | 6 | 3 | 4.76 × 10−05 | EEF2|HSP90AA1|HSP90AB1 |
| HSA-6785807 | Interleukin-4 and Interleukin-13 signaling | 107 | 5 | 7.35 × 10−05 | HSP90AA1|FN1|MMP9|HSPA8|ITGAM |
| HSA-9020591 | Interleukin-12 signaling | 46 | 4 | 1.10 × 10−04 | TCP1|P4HB|CDC42|CFL1 |
| Term ID | Term | Term Size | Enriched Terms | FDR | Associated Genes |
|---|---|---|---|---|---|
| Gene Ontology Biological Process | |||||
| GO:0045055 | Regulated exocytosis | 697 | 12 | 5.56 × 10−08 | TUBA4A|DSC1|IQGAP1|HK3|CD14|CLU|ARG1|HRNR|GSN|DSP|JUP|SERPINB6 |
| GO:0044419 | Interspecies interaction between organisms | 1899 | 12 | 1.60 × 10−04 | BPIFB2|CANX|BPIFA2|IQGAP1|RPS3|CD14|CLU|RPS3A|HNRNPA2B1|ARG1|GSN|CFL1 |
| GO:0060429 | Epithelium development | 1109 | 9 | 0.0011 | DSC1|IQGAP1|SPRR1B|HRNR|TACSTD2|PGK1|DSP|JUP|CFL1 |
| GO:0070268 | Cornification | 113 | 4 | 0.0036 | DSC1|SPRR1B|DSP|JUP |
| GO:2001235 | Positive regulation of apoptotic signaling pathway | 180 | 4 | 0.0162 | RPS3|CLU|GSN|YWHAZ |
| GO:0071345 | Cellular response to cytokine stimulus | 1013 | 7 | 0.0289 | CANX|RPS3|HNRNPA2B1|ARG1|GSN|YWHAZ|CFL1 |
| GO:0030155 | Regulation of cell adhesion | 712 | 6 | 0.0328 | IQGAP1|RPS3|ARG1|TACSTD2|GSN|JUP |
| Reactome pathways | |||||
| HSA-168256 | Immune System | 1956 | 18 | 4.76 × 10−11 | BPIFB2|CANX|TUBA4A|BPIFA2|DSC1|IQGAP1|HK3|CD14|CLU|HNRNPA2B1|ARG1|HRNR|GSN|DSP|JUP|YWHAZ|CFL1|SERPINB6 |
| HSA-6809371 | Formation of the cornified envelope | 127 | 4 | 0.0083 | DSC1|SPRR1B|DSP|JUP |
| HSA-109581 | Apoptosis | 173 | 4 | 0.0159 | CD14|GSN|DSP|YWHAZ |
| HSA-447115 | Interleukin-12 family signaling | 56 | 3 | 0.0159 | CANX|HNRNPA2B1|CFL1 |
| HSA-6803157 | Antimicrobial peptides | 87 | 3 | 0.0338 | BPIFB2|BPIFA2|CLU |
| HSA-76002 | Platelet activation, signaling and aggregation | 260 | 4 | 0.0461 | TUBA4A|CLU|YWHAZ|CFL1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guadalupi, G.; Contini, C.; Iavarone, F.; Castagnola, M.; Messana, I.; Faa, G.; Onali, S.; Chessa, L.; Vitorino, R.; Amado, F.; et al. Combined Salivary Proteome Profiling and Machine Learning Analysis Provides Insight into Molecular Signature for Autoimmune Liver Diseases Classification. Int. J. Mol. Sci. 2023, 24, 12207. https://doi.org/10.3390/ijms241512207
Guadalupi G, Contini C, Iavarone F, Castagnola M, Messana I, Faa G, Onali S, Chessa L, Vitorino R, Amado F, et al. Combined Salivary Proteome Profiling and Machine Learning Analysis Provides Insight into Molecular Signature for Autoimmune Liver Diseases Classification. International Journal of Molecular Sciences. 2023; 24(15):12207. https://doi.org/10.3390/ijms241512207
Chicago/Turabian StyleGuadalupi, Giulia, Cristina Contini, Federica Iavarone, Massimo Castagnola, Irene Messana, Gavino Faa, Simona Onali, Luchino Chessa, Rui Vitorino, Francisco Amado, and et al. 2023. "Combined Salivary Proteome Profiling and Machine Learning Analysis Provides Insight into Molecular Signature for Autoimmune Liver Diseases Classification" International Journal of Molecular Sciences 24, no. 15: 12207. https://doi.org/10.3390/ijms241512207
APA StyleGuadalupi, G., Contini, C., Iavarone, F., Castagnola, M., Messana, I., Faa, G., Onali, S., Chessa, L., Vitorino, R., Amado, F., Diaz, G., Manconi, B., Cabras, T., & Olianas, A. (2023). Combined Salivary Proteome Profiling and Machine Learning Analysis Provides Insight into Molecular Signature for Autoimmune Liver Diseases Classification. International Journal of Molecular Sciences, 24(15), 12207. https://doi.org/10.3390/ijms241512207