
Potential of Human Hemoglobin as a Source of Bioactive Peptides: Comparative Study of Enzymatic Hydrolysis with Bovine Hemoglobin and the Production of Active Peptide α137–141

 , , and
, , and
Abstract
1. Introduction
2. Results
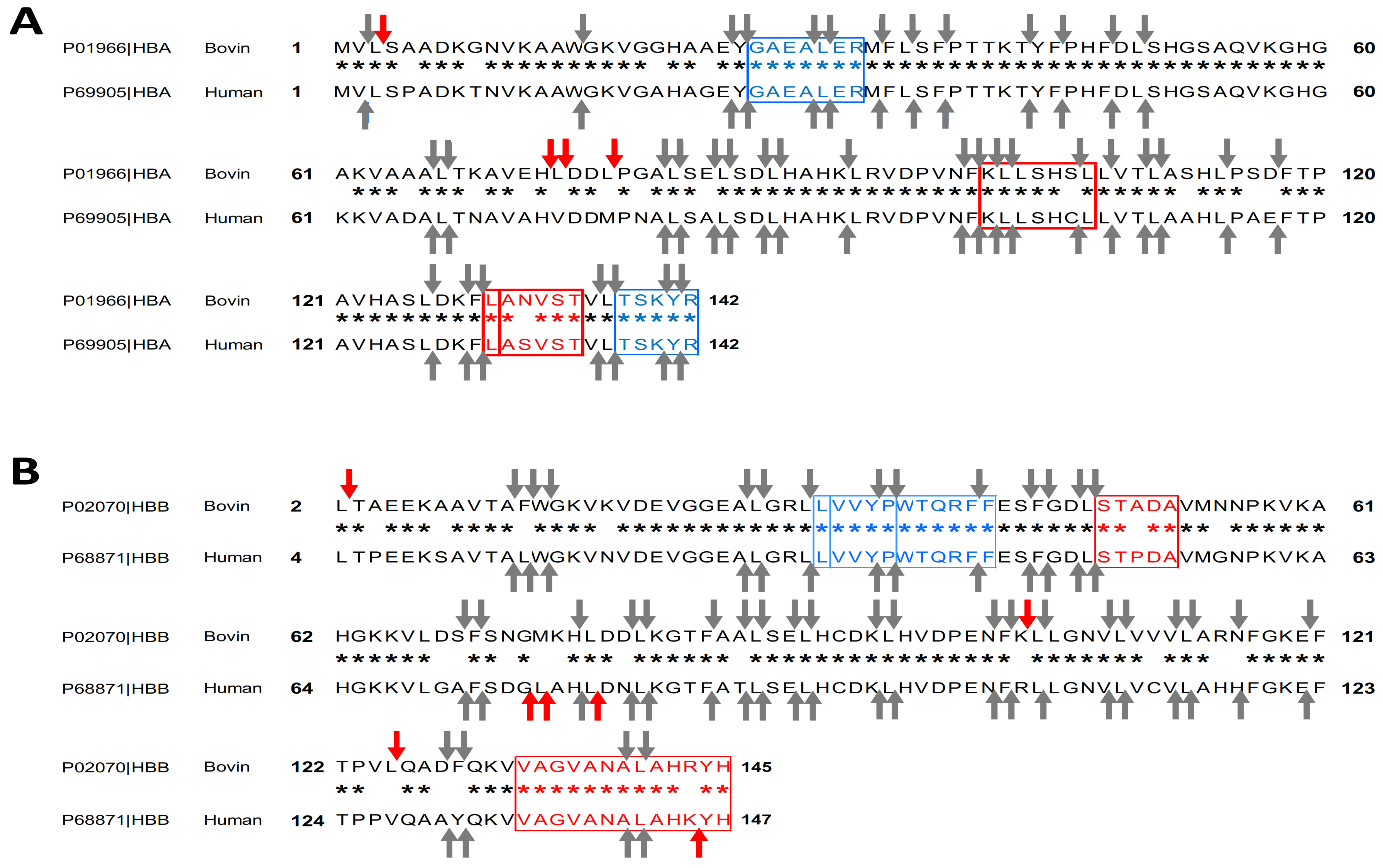
2.1. In Silico Comparative Analysis of Bovine (Bos taurus) and Human (Homo sapiens) Hemoglobin
2.2. Bioinformatics Approach to Predict Peptides Derived from the Pepsic Hydrolysis of Bovine Hemoglobin Bos taurus and Human Hemoglobin Homo sapiens
2.3. Enzymatic Kinetics and Mechanism of Action Involved
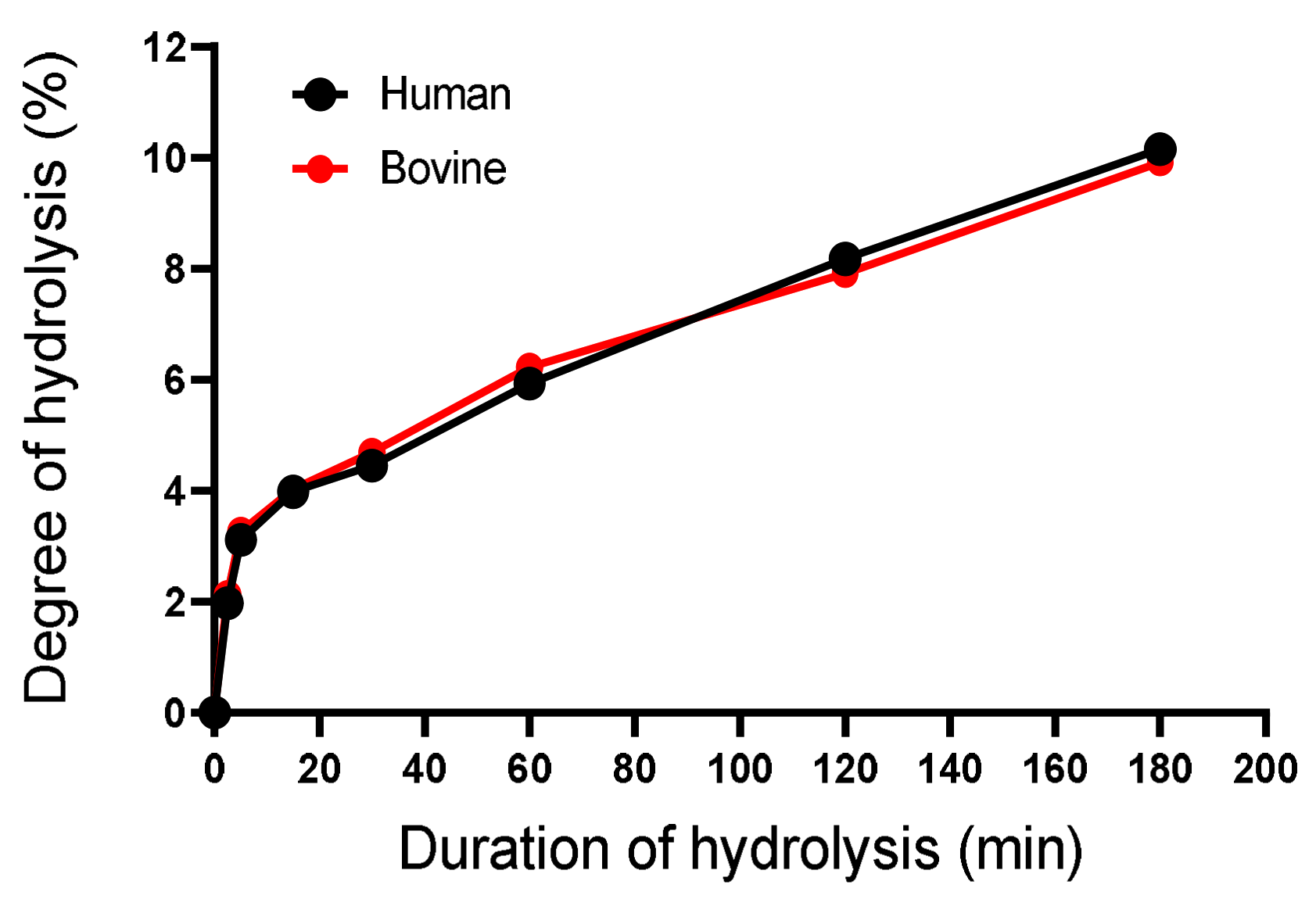
2.3.1. Determination of the Degree of Hydrolysis
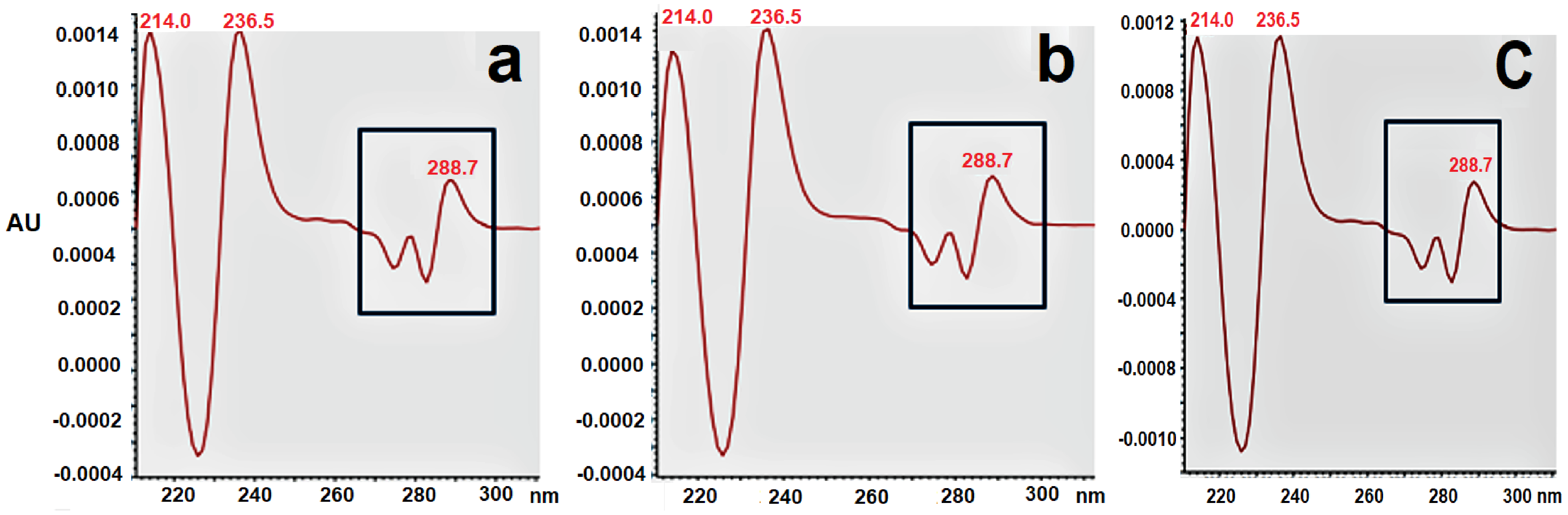
2.3.2. Identification of α137–141 by Waters Software
2.3.3. Reaction Mechanism of the Enzymatic Hydrolysis of Human Hemoglobin by Pepsin
2.4. Obtaining the Active Peptide α137–141 during the Peptide Hydrolysis of Human and Bovine Hemoglobin
2.4.1. Effect of Increasing the Initial Concentration of Bovine Hemoglobin on the Enzymatic Obtention of Active Peptide α137–141
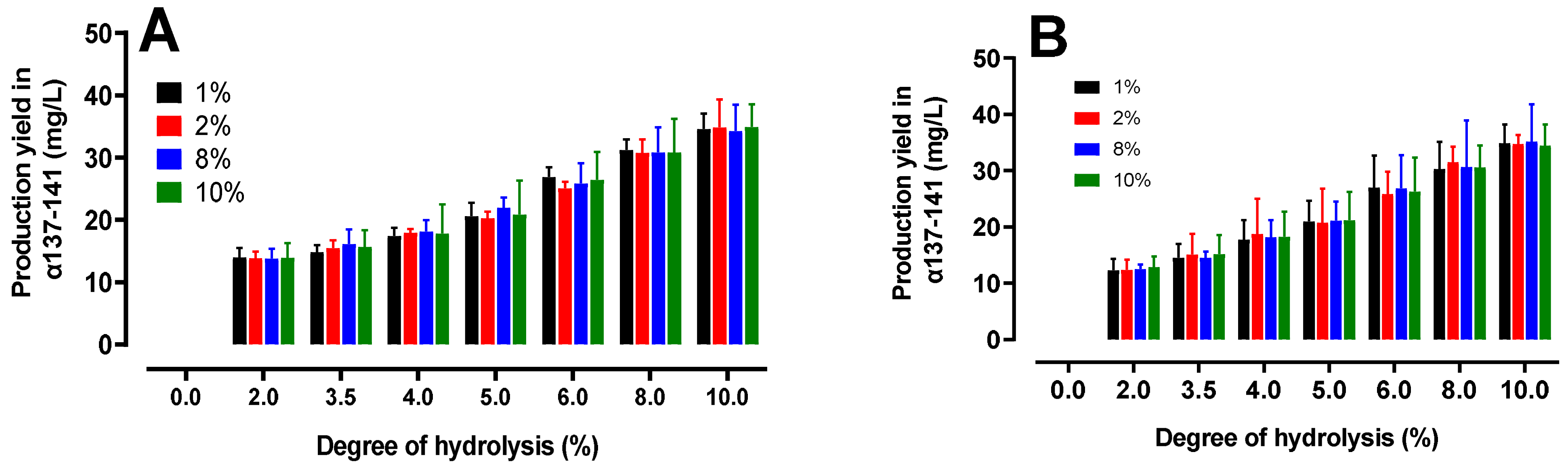
2.4.2. Quantification of the Production of the Active Peptide α137–141
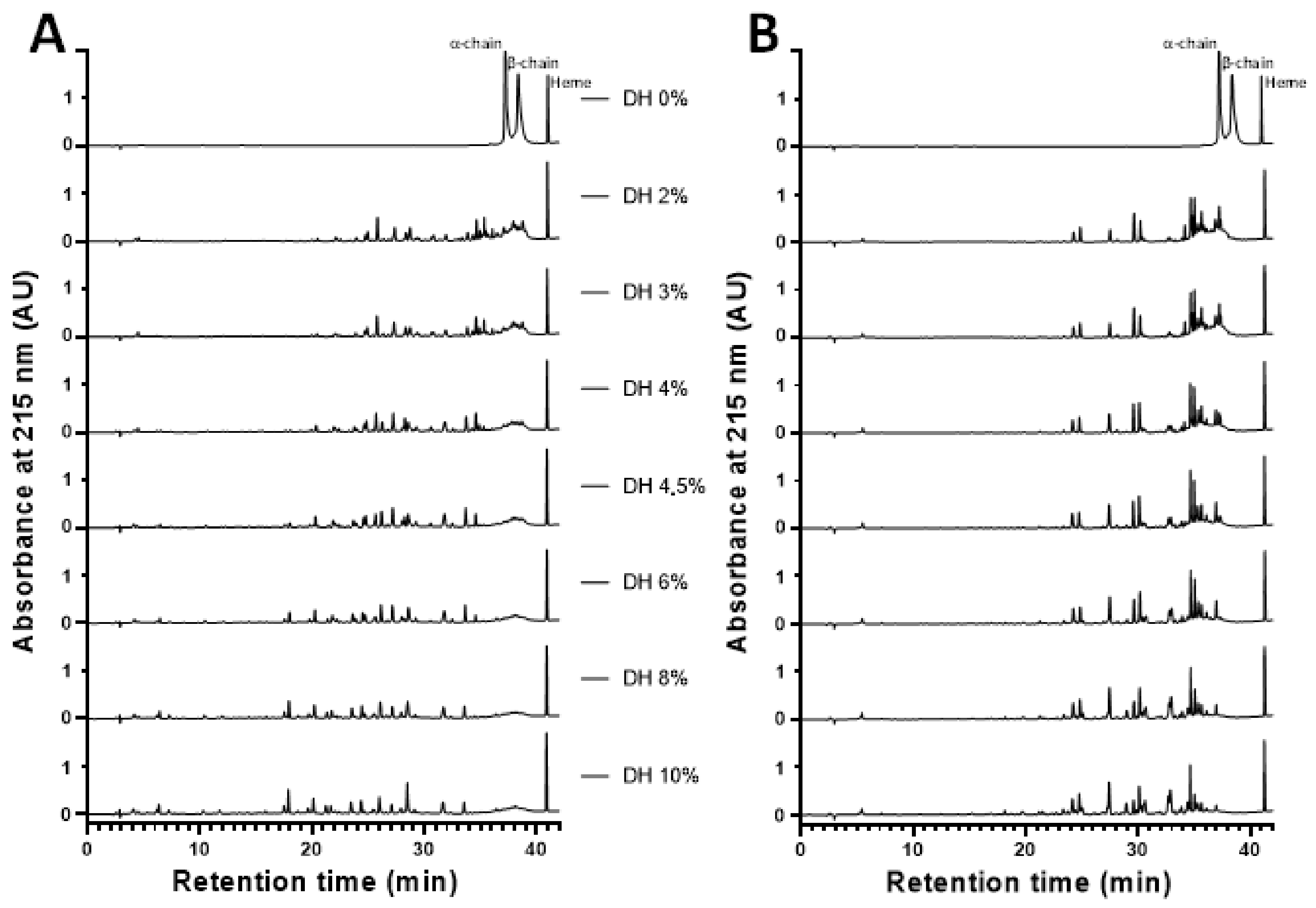
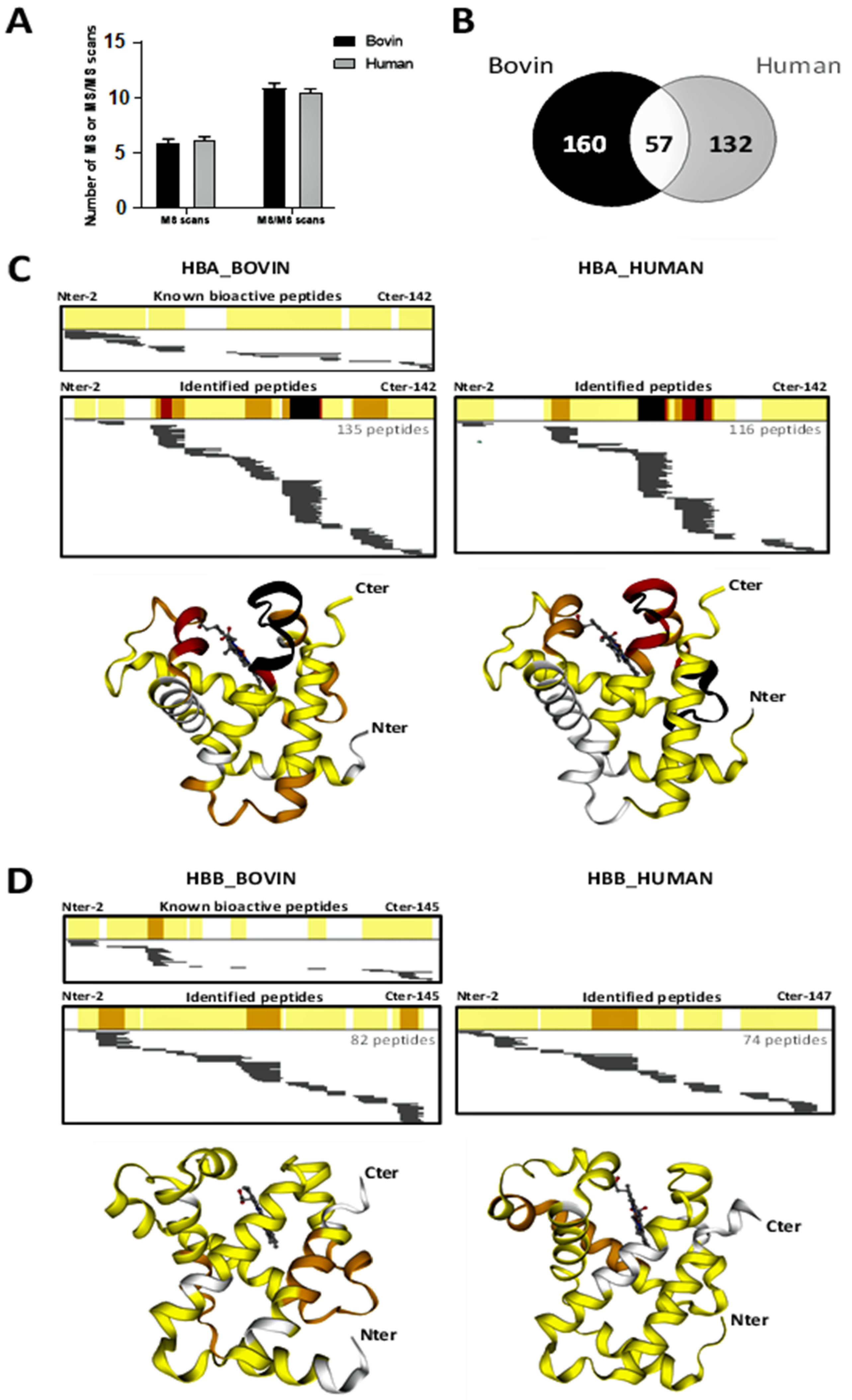
2.5. Peptidomics Approach to Characterizing the Peptide Populations
2.6. Focus on Bioactive Peptides from Bovine and Human Hemoglobin Hydrolysis
3. Discussion
3.1. Reaction Mechanism and Production of the Active Peptide α137–141 during the Enzymatic Hydrolysis of Human and Bovine Hemoglobin
3.2. Application of the Peptidomics Approach to the Study of Peptide Populations
4. Materials and Methods
4.1. Comparison of Peptide Sequences by Bioinformatics Approach
4.2. Prediction of Clipping Sites by Bioinformatics Approach
4.3. Materials: Reagents, Solvents and Standards Used
- -
- Purified bovine hemoglobin powder (H2625), dark brown, and purified human hemoglobin (H7379), dark red, were obtained from Sigma-Aldrich (St. Louis, MI, USA). The hemoglobin were stored at 4 °C before use.
- -
- Pepsin, a lyophilized powder sourced from porcine gastric mucosa, purchased from Sigma-Aldrich (P6887). Its activity was measured at 3250 AU/mg protein using a protocol established by the supplier. To maintain the stability of the pepsin, it was stored at a −20 °C.
4.4. Preparation of Hydrolysates
4.4.1. Preparation of the Stock Solution
4.4.2. Hydrolysis Process
4.5. Determination of the Degree of Bovine and Human Hemoglobin Hydrolysis
4.6. Analysis of the Peptide Hydrolysate by RP-UPLC and RP-HPLC
4.6.1. Hardware, Software, and Protocol Used
Analysis by RP-UPLC
4.6.2. Identification of the α137–141Peptide
4.6.3. Quantification of α137–141 in Hydrolysates
4.7. Peptidomics Approach
4.8. Two-Dimensional (2D) and Tri-Dimensional (3D) Heatmaps
4.9. RP-UPLC Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gatnau, R.; Polo, J.; Robert, E. Plasma protein antimicrobial substitution at negligible risk, Feed Manufacturing in the Mediterranean Region. Improv. Saf. Feed Food 2001, 54, 141–150. [Google Scholar]
- Mora, L.; Reig, M.; Toldrá, F. Bioactive peptides generated from meat industry by-products. Food Res. Int. 2014, 65, 344–349. [Google Scholar]
- Gómez-Juárez, C.; Castellanos, R.; Ponce-Noyola, T.; Calderón, V.; Figueroa, J. Protein recovery from slaughterhouse wastes. Bioresour. Technol. 1999, 70, 129–133. [Google Scholar] [CrossRef]
- Bah, C.S.; Bekhit, A.E.-D.A.; Carne, A.; McConnell, M.A. Slaughterhouse blood: An emerging source of bioactive compounds. Compr. Rev. Food Sci. Food Saf. 2013, 12, 314–331. [Google Scholar] [CrossRef]
- Toldrá, F.; Aristoy, M.-C.; Mora, L.; Reig, M. Innovations in value-addition of edible meat by-products. Meat Sci. 2012, 92, 290–296. [Google Scholar] [CrossRef]
- Hsieh, Y.-H.P.; Ofori, J.A. Blood-derived products for human consumption. Revel. Sci. 2011, 1, 14–21. [Google Scholar]
- Sasakawa, S. Studies on Hemoglobin VI. Amino Acid Compositions of the Fractionated Bovine Globin α and β. J. Biochem. 1961, 50, 345–351. [Google Scholar] [CrossRef]
- Lignot, B.; Froidevaux, R.; Nedjar-Arroume, N.; Guillochon, D. Solvent effect on kinetics of appearance of neokyotorphin, VV-haemorphin-4 and a bradykinin-potentiating peptide in the course of peptic hydrolysis of bovine haemoglobin. Biotechnol. Appl. Biochem. 1999, 30, 201–207. [Google Scholar]
- Zhao, Q.; Molina, P.; Piot, J.M. Peptic peptide mapping by HPLC, on line with photodiode array detection, of a hemoglobin hydrolysate produced at pilot-plant scale from an ultrafiltration process. J. Liq. Chromatogr. Relat. Technol. 1997, 20, 1717–1739. [Google Scholar] [CrossRef]
- Vercaigne-Marko, D.; Kosciarz, E.; Nedjar-Arroume, N.; Guillochon, D. Improvement of Staphylococcus aureus-V8-protease hydrolysis of bovine haemoglobin by its adsorption on to a solid phase in the presence of SDS: Peptide mapping and obtention of two haemopoietic peptides. Biotechnol. Appl. Biochem. 2000, 31, 127–134. [Google Scholar] [CrossRef]
- Adje, E.Y. Hydrolyse Ménagée de L’hémoglobine Bovine par la Pepsine Porcine en Mélanges Hydroalcooliques et Obtention D’une Nouvelle Famille de Peptides Antimicrobiens. Ph.D. Thesis, Université de Lille, Villeneuve-d’Ascq, France, 2010. [Google Scholar]
- Daoud, R.; Dubois, V.; Bors-Dodita, L.; Nedjar-Arroume, N.; Krier, F.; Chihib, N.-E.; Mary, P.; Kouach, M.; Briand, G.; Guillochon, D. New antibacterial peptide derived from bovine hemoglobin. Peptides 2005, 26, 713–719. [Google Scholar] [CrossRef]
- Nedjar-Arroume, N.; Dubois-Delval, V.; Adje, E.Y.; Traisnel, J.; Krier, F.; Mary, P.; Kouach, M.; Briand, G.; Guillochon, D. Bovine hemoglobin: An attractive source of antibacterial peptides. Peptides 2008, 29, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Choisnard, L.; Froidevaux, R.; Nedjar-Arroume, N.; Lignot, B.; Vercaigne-Marko, D.; Krier, F.; Dhulster, P.; Guillochon, D. Kinetic study of the appearance of an anti-bacterial peptide in the course of bovine haemoglobin peptic hydrolysis. Biotechnol. Appl. Biochem. 2002, 36, 187–194. [Google Scholar] [CrossRef]
- Przybylski, R.; Firdaous, L.; Châtaigné, G.; Dhulster, P.; Nedjar, N. Production of an antimicrobial peptide derived from slaughterhouse by-product and its potential application on meat as preservative. Food Chem. 2016, 211, 306–313. [Google Scholar] [CrossRef] [PubMed]
- De Melo, J.G.; de Sousa Araújo, T.A.V. Thijan Nobre de Almeida e Castro, D. Lyra de Vasconcelos Cabral, M. Do Desterro Rodrigues, S. Carneiro do Nascimento, E.L. Cavalcanti de Amorim, U.P. De Albuquerque, Antiproliferative activity, antioxidant capacity and tannin content in plants of semi-arid northeastern Brazil. Molecules 2010, 15, 8534–8542. [Google Scholar]
- Sakanaka, S.; Tachibana, Y.; Okada, Y. Preparation and antioxidant properties of extracts of Japanese persimmon leaf tea (kakinoha-cha). Food Chem. 2005, 89, 569–575. [Google Scholar] [CrossRef]
- Chen, L.; Xiong, Z.; Sun, L.; Yang, J.; Jin, Q. VFDB 2012 update: Toward the genetic diversity and molecular evolution of bacterial virulence factors. Nucleic Acids Res. 2012, 40, D641–D645. [Google Scholar] [CrossRef]
- Saadi, S.; Saari, N.; Anwar, F.; Hamid, A.A.; Ghazali, H.M. Recent advances in food biopeptides: Production, biological functionalities and therapeutic applications. Biotechnol. Adv. 2015, 33, 80–116. [Google Scholar] [CrossRef]
- Linderstrom-Lang, K. Les phases initiales de la dégradation des protéines par les enzymes. Bull. Société Chim. Biol. 1953, 35, 100–116. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Huang, X.; Miller, W. A time-efficient, linear-space local similarity algorithm. Adv. Appl. Math. 1991, 12, 337–357. [Google Scholar] [CrossRef]
- Takagi, H.; Shiomi, H.; Fukui, K.; Hayashi, K.; Kiso, Y.; Kitagawa, K. Isolation of a novel analgesic pentapeptide, neo-kyotorphin, from bovine brain. Life Sci. 1982, 31, 1733–1736. [Google Scholar] [CrossRef]
- Ivanov, V.; Andrei, T.; Karelin, A.; Mikhaleva, I.; Vaskovsky, B.V.; Sviryaev, V.L.; Nazimov, I.V. Isolation, structure and properties of endogenous peptides. Russ. J. Bioorganic Chem. 1992, 18, 1271–1311. [Google Scholar]
- Piot, J.-M.; Zhao, Q.; Guillochon, D.; Ricart, G.; Thomas, D. Isolation and characterization of two opioid peptides from a bovine hemoglobin peptic hydrolysate. Biochem. Biophys. Res. Commun. 1992, 189, 101–110. [Google Scholar] [CrossRef]
- Abou-Diab, M.; Thibodeau, J.; Deracinois, B.; Flahaut, C.; Fliss, I.; Dhulster, P.; Nedjar, N.; Bazinet, L. Bovine hemoglobin enzymatic hydrolysis by a new eco-efficient process—Part II, production of bioactive peptides. Membranes 2020, 10, 268. [Google Scholar] [CrossRef] [PubMed]
- Caron, J.; Cudennec, B.; Domenger, D.; Belguesmia, Y.; Flahaut, C.; Kouach, M.; Lesage, J.; Goossens, J.-F.; Dhulster, P.; Ravallec, R. Simulated GI digestion of dietary protein: Release of new bioactive peptides involved in gut hormone secretion. Food Res. Int. 2016, 89, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Catiau, L.; Traisnel, J.; Delval-Dubois, V.; Chihib, N.-E.; Guillochon, D.; Nedjar-Arroume, N. Minimal antimicrobial peptidic sequence from hemoglobin alpha-chain: KYR. Peptides 2011, 32, 633–638. [Google Scholar] [CrossRef]
- Albuquerque, W.; Ghezellou, P.; Lee, K.-Z.; Schneider, Q.; Gross, P.; Kessel, T.; Omokungbe, B.; Spengler, B.; Vilcinskas, A.; Zorn, H. Peptidomics as a Tool to Assess the Cleavage of Wine Haze Proteins by Peptidases from Drosophila suzukii Larvae. Biomolecules 2023, 13, 451. [Google Scholar] [CrossRef]
- Zhao, Q.; Sannier, F.; Piot, J.M. Kinetics of appearance of four hemorphins from bovine hemoglobin peptic hydrolysates by HPLC coupled with photodiode array detection, Biochimica et Biophysica Acta (BBA)-Protein Structure and Molecular. Enzymology 1996, 1295, 73–80. [Google Scholar]
- Kagawa, K.; Matsutaka, H.; Fukuhama, C.; Watanabe, Y.; Fujino, H. Globin digest; acidic protease hydrolysate, inhibits dietary hypertriglyceridemia and Val-Val-Tyr-Pro, one of its constituents, possesses most superior effect. Life Sci. 1996, 58, 1745–1755. [Google Scholar] [CrossRef]
- Zhao, Q.; Sannier, F.; Ricart, G.; Piot, J.M. A rapid detection and identification of hemorphins released from bovine hemoglobin enzymatic hydrolysis by use of HPLC coupled with photodiode array detector. J. Liq. Chromatogr. Relat. Technol. 1995, 18, 93–103. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, M.; Zhang, J.; Zhao, D.; Sun, J. Peptidomics insights into the interplay between the pre-digestion effect of mixed starters and the digestive pattern of sausage proteins. Food Res. Int. 2022, 162, 111963. [Google Scholar] [CrossRef] [PubMed]
- Dubois, V.; Nedjar-Arroume, N.; Guillochon, D. Influence of pH on the appearance of active peptides in the course of peptic hydrolysis of bovine haemoglobin. Prep. Biochem. Biotechnol. 2005, 35, 85–102. [Google Scholar] [CrossRef] [PubMed]
- Barkhudaryan, N.; Oberthuer, W.; Lottspeich, F.; Galoyan, A. Structure of hypothalamic coronaro-constrictory peptide factors. Neurochem. Res. 1992, 17, 1217–1221. [Google Scholar] [CrossRef]
- Blishchenko, E.Y.; Mernenko, O.A.; Mirkina, I.I.; Satpaev, D.K.; Ivanov, V.S.; Tchikin, L.D.; Ostrovsky, A.G.; Karelin, A.A.; Ivanov, V.T. Tumor cell cytolysis mediated by valorphin, an opioid-like fragment of hemoglobin β-chain. Peptides 1997, 18, 79–85. [Google Scholar] [CrossRef]
- Zouari, O.; Przybylski, R.; Hannioui, M.; Sion, L.; Dhulster, P.; Nedjar-Arroume, N. High added-value co-product: The porcine cruor is an attractive source of active peptides. J. Nutr. Health Food Sci 2020, 7, 1–9. [Google Scholar]
- Zouari, O.; Deracinois, B.; Flahaut, C.; Przybylski, R.; Nedjar, N. Poultry cruor hydrolysate is a new potential source of hemoglobin: Obtaining of active peptides. Res. Sq. 2023. preprint. [Google Scholar] [CrossRef]
- Nicolas, P.; Mor, A. Peptides as weapons against microorganisms in the chemical defense system of vertebrates. Annu. Rev. Microbiol. 1995, 49, 277–304. [Google Scholar] [CrossRef]
- Powers, J.-P.S.; Hancock, R.E. The relationship between peptide structure and antibacterial activity. Peptides 2003, 24, 1681–1691. [Google Scholar] [CrossRef]
- Mak, P.; Wójcik, K.; Wicherek, Ł.; Suder, P.; Dubin, A. Antibacterial hemoglobin peptides in human menstrual blood. Peptides 2004, 25, 1839–1847. [Google Scholar] [CrossRef]
- Parish, C.A.; Jiang, H.; Tokiwa, Y.; Berova, N.; Nakanishi, K.; McCabe, D.; Zuckerman, W.; Xia, M.M.; Gabay, J.E. Broad-spectrum antimicrobial activity of hemoglobin. Bioorganic Med. Chem. 2001, 9, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Dunn, B.M. Structure and mechanism of the pepsin-like family of aspartic peptidases. Chem. Rev. 2002, 102, 4431–4458. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Spellman, D.; McEvoy, E.; O’cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Church, F.C.; Porter, D.H.; Catignani, G.L.; Swaisgood, H.E. An o-phthalaldehyde spectrophotometric assay for proteinases. Anal. Biochem. 1985, 146, 343–348. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Activity | Position | Sequence | Monoisotopic Molecular (Da) | Bovine Hemoglobin | Human Hemoglobin |
|---|---|---|---|---|---|
| Antimicrobial | α34–46 | LSFPTTKTYFPHF | 1584.787 | + | − |
| α33–46 | FLSFPTTKTYFPHF | 1731.855 | + | − | |
| α37–46 | PTTKTYFPHF | 1237.602 | + | + | |
| α36–45 | FPTTKTYFPH | 1237.602 | + | + | |
| α137–141 | TSKYR | 653.339 | + | + | |
| α133–141 | STVLTSKYR | 1053.571 | + | + | |
| α99–105 | KLLSHSL | 796.470 | + | KLLSHCL | |
| α100–105 | LLSHSL | 668.375 | + | − | |
| α99–106 | KLLSHSLL | 909.554 | + | KLLSHCLL | |
| Hematopoietic | α76–82 | LPGALSE | 685.354 | + | MPNALSA |
| β115–122 | RNFGKEFT | 997.487 | + | HHFGKEFT | |
| Opioid | α137–141 | TSKYR | 653.339 | + | + |
| β32–40 | VVYPWTQRF | 1194.608 | + | + | |
| β31–40 | LVVYPWTQRF | 1307.692 | + | + | |
| β31–37 | LVVYPWT | 876.464 | + | − | |
| Analgesic and Potentiator of bradykinin | α129–134 | LANVST | 603.312 | + | LASVST |
| β129–134 | QKVVAG | 600.349 | + | − | |
| dipeptidyl-peptidase Inhibitor | α130–134 | ANVST | 490.228 | + | ASVST |
| β6–10 | KAAVT | 488.285 | + | KSAVT | |
| ACE inhibition | β129–135 | KVVAGVA | 642.395 | + | + |
| Antihypertensive | α99–105 | KLLSHSL | 796.470 | + | KLLSHCL |
| Antioxidant | α137–141 | TSKYR | 653.339 | + | + |
| Bacterial growth stimulator | β48–52 | STADA | 463.180 | + | STPDA |
| Anticancer | β33–39 | VVYPWTQ | 891.438 | + | + |
| Position | Sequence | Monoisotopic Molecular (Da) | Bovine Hemoglobin | Human Hemoglobin | Ref |
|---|---|---|---|---|---|
| The first family is situated on the N-terminal end of the α-chain, with the active portion found between residues 1 and 23. | |||||
| α1–40 | VLSPADKTNVKAAWGKVGAHAGEYGAEALERMFLSFPTTK | 4249 | − | + | b |
| α1–33 | VLSPADKTNVKAAWGKVGAHAGEYGAEALERMF | 3474 | − | + | b |
| α1–32 | VLSPADKTNVKAAWGKVGAHAGEYGAEALERM | 3327 | + | + | a |
| VLSAADKGNVKAAWGKVGGHAAEYGAEALERM (bovine) | 3257 | + | − | ||
| α1–31 | VLSPADKTNVKAAWGKVGAHAGEYGAEALER | 3196 | − | + | b |
| α1–29 | VLSPADKTNVKAAWGKVGAHAGEYGAEAL | 2911 | − | + | b |
| VLSAADKGNVKAAWGKVGGHAAEYGAEAL (bovine) | 2841 | + | − | a | |
| α1–28 | VLSAADKGNVKAAWGKVGGHAAEYGAEA | 2728 | + | − | a |
| α1–27 | VLSAADKGNVKAAWGKVGGHAAEYGAE | 2656 | + | − | a |
| α1–23 | VLSAADKGNVKAAWGKVGGHAAE | 2237 | + | + | a |
| α1–20 | VLSPADKTNVKAAWGKVGAH | 2949 | − | + | b |
| α18–44 | VGAHAGEYGAEALERMFLSFPTTKTYF | 2994 | − | + | b |
| A second family of peptides located between residues 32 and 98, with an active sequence between residues 36 and 46. | |||||
| α32–41 | FLSFPTTKTY | 1204 | − | + | c |
| α33–46 | FLSFPTTKTYFPHF | 1731 | + | + | a/e |
| α34–46 | LSFPTTKTYFPHF | 1585 | + | + | a/e |
| α36–45 | FPTTKTYFPH | 1238 | + | + | a/e |
| α37–46 | PTTKTYFPHF | 1238 | + | + | a/e |
| α33–98 | FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVNF | 7151 | + | − | a |
| α33–97 | FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVN | 7004 | + | − | a |
| α34–98 | LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVNF | 7004 | + | − | a |
| α36–97 | SFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVN | 6744 | + | − | a |
| α37–98 | PTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSELSDLHAHKLRVDPVNF | 6657 | + | − | a |
| α33–83 | FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSEL | 5422 | + | − | a |
| α34–83 | LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAALTKAVEHLDDLPGALSEL | 5274 | + | − | a |
| α33–66 | FLSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAAL | 3632 | + | − | a |
| α34–66 | LSFPTTKTYFPHFDLSHGSAQVKGHGAKVAAAL | 3484 | + | − | a |
| α35–56 | SFPTTKTYFPHFDLSHGSAQVK | 2495 | − | + | b |
| α35–80 | SFPTTKTYFPHFDLSHGSAQVKGHGKKVADALTNAVAHVDDMPNAL | 4922 | − | + | b |
| α35–77 | SFPTTKTYFPHFDLSHGSAQVKGHGKKVADALTNAVAHVDMP | 4624 | − | + | b |
| The third family is located on the c-terminal side of the α-chain. | |||||
| α110–131 | AAHLPAEFTPAVHASLDKFLAS | 2293 | + | − | a |
| α107–141 | VTLASHLPSDFTPAVHASLDKFLANVSTVLTSKYR | 3788 | + | − | a |
| α107–136 | VTLASHLPSDFTPAVHASLDKFLANVSTVL | 3152 | + | − | a |
| α107–133 | VTLASHLPSDFTPAVHASLDKFLANVS | 2838 | + | − | a |
| α133–141 | STVLTSKYR | 1055 | + | + | a/e |
| α137–141 | TSKYR | 654 | + | + | a/e |
| α99–105 | KLLSHSL (bovine) | 796 | + | − | a |
| KLLSHCL | 813 | − | + | e | |
| α100–105 | LLSHSL | 668 | + | − | a |
| α99–106 | KLLSHSLL | 910 | + | − | a |
| KLLSHCLL | 926 | − | + | e | |
| The last family of peptides is located in the c terminal region of the β chain of hemoglobin | |||||
| β1–30 | MLTAEEKAAVTAFWGKVKVDEVGGEALGRL (bovine) | 3176 | + | + | a |
| MVH LTPEEKSA VTALWGKVNVDEVGGEALG | 3137 | ||||
| β1–55 | MVHLTPEEKSAVTALWGKVNVDEVGGEALGRLLVVYPWTQRFFESFGDLSTPDAV | 6063 | − | + | d |
| β56–146 | MGNPKVKAHGKKVLGAFSDGLAHLDNLKGTFATLSELHCDKLHVDPENFRLLGNVLVCVLAHHFGKEFTPPVQAAYQKVVAGVANALAHKY | 9815 | − | + | d |
| β116–146 | LLGNVLVCVLAHHFGKEFTPPVQAAYQKVVAGVANALAHKY | 4375 | − | + | d |
| β56–72 | MGNPKVKAHGKKVLGAF | 1782 | − | + | d |
| β43–83 | RFFESFGDLSTPDAVMGNPKVKAHGKKVLGAFSDGLAHLDNLK | 4616 | − | + | b |
| β111–146 | LVCVLAHHFGKEFTPPVQAAYQKVVAGVANALAHKY | 3878 | − | + | b |
| β115–146 | LAHHFGKEFTPPVQAAYQKVVAGVANALAHKY | 3463 | − | + | b |
| β114–145 | ARNFGKEFTPVLQADFQKVVAGVANALAHRYH | 3556 | + | − | a |
| β121–145 | FTPVLQADFQKVVAGVANALAHRYH | 2753 | − | + | b |
| β126–145 | QADFQKVVAGVANALAHRYH | 2196 | + | − | b |
| β1–13 | MLTAEEKAAVTAF | 1381 | + | − | a |
| β140–145 | LAHRYH | 795 | + | − | a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Outman, A.; Deracinois, B.; Flahaut, C.; Diab, M.A.; Gressier, B.; Eto, B.; Nedjar, N. Potential of Human Hemoglobin as a Source of Bioactive Peptides: Comparative Study of Enzymatic Hydrolysis with Bovine Hemoglobin and the Production of Active Peptide α137–141. Int. J. Mol. Sci. 2023, 24, 11921. https://doi.org/10.3390/ijms241511921
Outman A, Deracinois B, Flahaut C, Diab MA, Gressier B, Eto B, Nedjar N. Potential of Human Hemoglobin as a Source of Bioactive Peptides: Comparative Study of Enzymatic Hydrolysis with Bovine Hemoglobin and the Production of Active Peptide α137–141. International Journal of Molecular Sciences. 2023; 24(15):11921. https://doi.org/10.3390/ijms241511921
Chicago/Turabian StyleOutman, Ahlam, Barbara Deracinois, Christophe Flahaut, Mira Abou Diab, Bernard Gressier, Bruno Eto, and Naïma Nedjar. 2023. "Potential of Human Hemoglobin as a Source of Bioactive Peptides: Comparative Study of Enzymatic Hydrolysis with Bovine Hemoglobin and the Production of Active Peptide α137–141" International Journal of Molecular Sciences 24, no. 15: 11921. https://doi.org/10.3390/ijms241511921
APA StyleOutman, A., Deracinois, B., Flahaut, C., Diab, M. A., Gressier, B., Eto, B., & Nedjar, N. (2023). Potential of Human Hemoglobin as a Source of Bioactive Peptides: Comparative Study of Enzymatic Hydrolysis with Bovine Hemoglobin and the Production of Active Peptide α137–141. International Journal of Molecular Sciences, 24(15), 11921. https://doi.org/10.3390/ijms241511921