
Gender-Specific Differences in Human Vertebral Bone Marrow Clot
 , , , ,
, , , ,  ,
,  , ,
, ,  ,
,  ,
, 

Abstract
1. Introduction
1.1. Vertebral Bone Marrow Clot
1.2. Gender Differences
2. Results
2.1. Enzyme-Linked Immunosorbent Assay (ELISA)
2.2. Cell Morphology and Viability
2.3. Flow Cytometry
2.4. Population-Doubling Time (PDT)
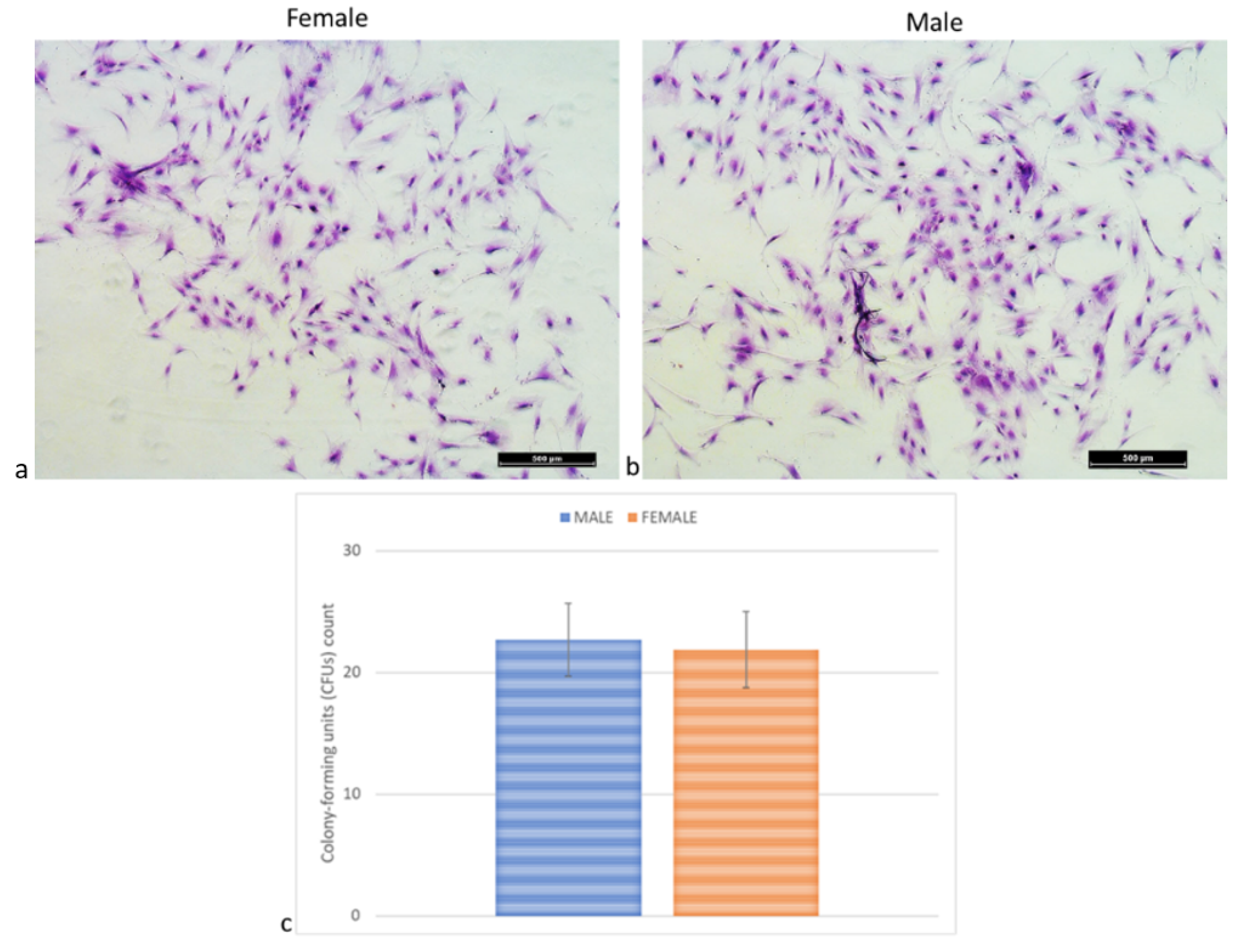
2.5. Colony-Forming Units (CFUs) Assay
2.6. Osteogenic, Adipogenic, and Chondrogenic Differentiation Ability
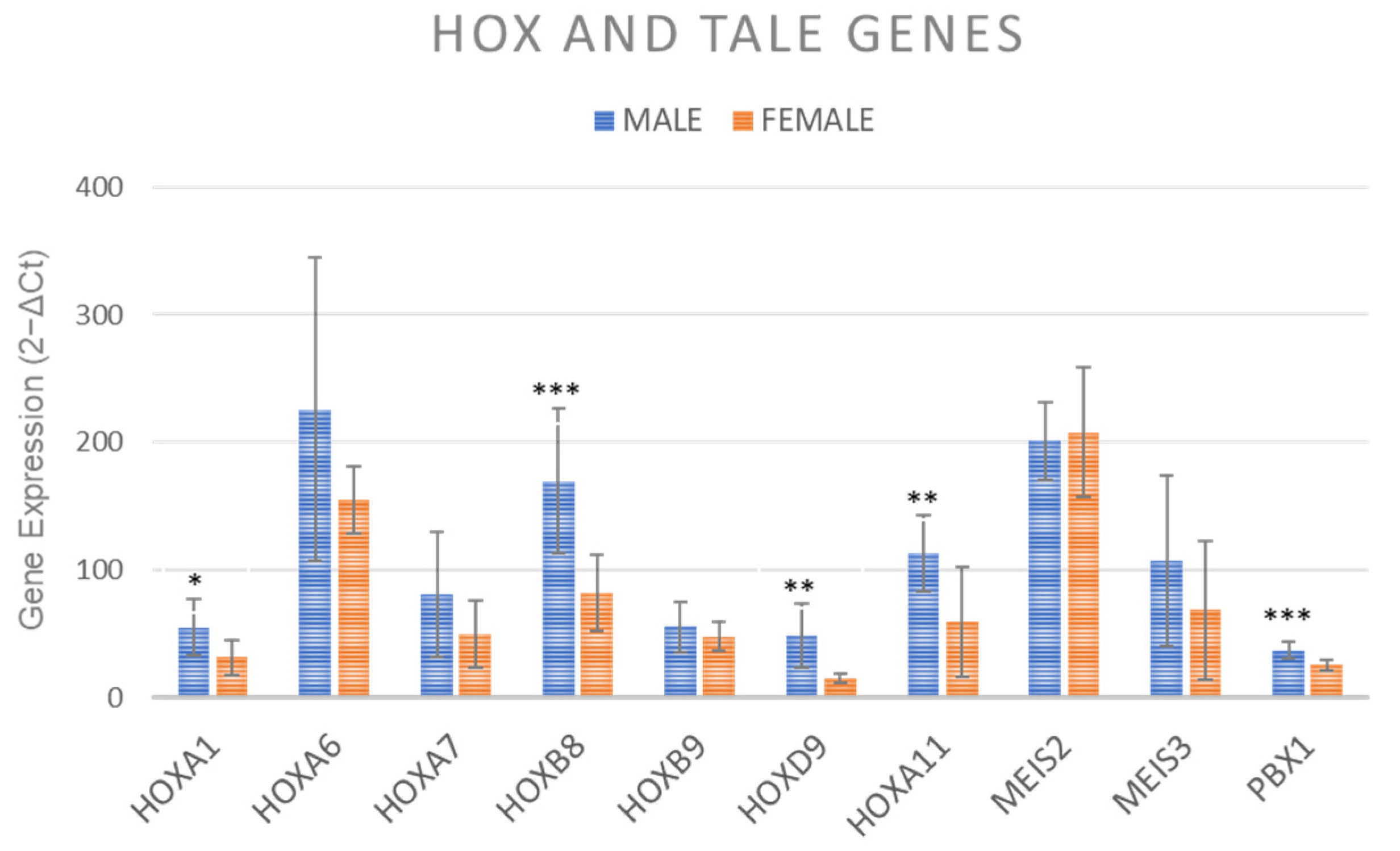
2.7. Gene Expression of HOX and TALE Genes
2.8. Senescence Associated Factors Expression
3. Discussion
Gender-Related Differences in vBMA Clot
4. Materials and Methods
4.1. Isolation and Culture of Human BMA Clot
4.2. Enzyme-Linked Immunosorbent Assay (ELISA)
4.3. Cell Morphology and Viability
4.4. Flow Cytometry
4.5. Population-Doubling Time
4.6. Colony-Forming Assay
4.7. Osteogenic, Adipogenic, and Chondrogenic Differentiation Ability
4.8. Staining
4.9. Gene Expression
4.10. Gene Expression of HOX and TALE Genes
4.11. Senescence Associated Factors Expression
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khashan, M.; Inoue, S.; Berven, S.H. Cell based therapies as compared to autologous bone grafts for spinal arthrodesis. Spine 2013, 38, 1885–1891. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, C.E.; Lee, K.B.; Keorochana, G.; Tzeng, S.T.; Yoo, J.H.; Wang, J.C. Bone morphogenetic protein-2 and bone marrow aspirate with allograft as alternatives to autograft in instrumented revision posterolateral lumbar spinal fusion: A minimum two-year follow-up study. Spine 2010, 35, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Hart, R.; Komzák, M.; Okál, F.; Náhlík, D.; Jajtner, P.; Puskeiler, M. Allograft alone versus allograft with bone marrow concentrate for the healing of the instrumented posterolateral lumbar fusion. Spine J. 2014, 14, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.C.; Tsai, T.T.; Fu, T.S.; Lai, P.L.; Chen, L.H.; Chen, W.J. A comparison of posterolateral lumbar fusion comparing autograft, autogenous laminectomy bone with bone marrow aspirate, and calcium sulphate with bone marrow aspirate: A prospective randomized study. Spine 2009, 34, 2715–2719. [Google Scholar] [CrossRef]
- Schottel, P.C.; Warner, S.J. Role of Bone Marrow Aspirate in Orthopedic Trauma. Orthop. Clin. N. Am. 2017, 48, 311–321. [Google Scholar] [CrossRef]
- Baksh, D.; Zandstra, P.W.; Davies, J.E. A non-contact suspension culture approach to the culture of osteogenic cells derived from a CD49elow subpopulation of human bone marrow-derived cells. Biotechnol. Bioeng. 2007, 98, 1195–1208. [Google Scholar] [CrossRef]
- Yasuhara, S.; Yasunaga, Y.; Hisatome, T.; Ishikawa, M.; Yamasaki, T.; Tabata, Y.; Ochi, M. Efficacy of bone marrow mononuclear cells to promote bone regeneration compared with isolated CD34+ cells from the same volume of aspirate. Artif. Organs 2010, 34, 594–599. [Google Scholar] [CrossRef]
- Goel, A.; Sangwan, S.S.; Siwach, R.C.; Ali, A.M. Percutaneous bone marrow grafting for the treatment of tibial non-union. Injury 2005, 36, 203–206. [Google Scholar] [CrossRef]
- Braly, H.L.; O′Connor, D.P.; Brinker, M.R. Percutaneous autologous bone marrow injection in the treatment of distal meta-diaphyseal tibial nonunions and delayed unions. J. Orthop. Trauma. 2013, 27, 527–533. [Google Scholar] [CrossRef]
- Kassem, M.S. Percutaneous autogenous bone marrow injection for delayed union or non union of fractures after internal fixation. Acta Orthop. Belg. 2013, 79, 711–717. [Google Scholar]
- Singh, A.K.; Sinha, A. Percutaneous autologous bone marrow injections for delayed or non-union of bones. J. Orthop. Surg. 2013, 21, 267. [Google Scholar] [CrossRef] [PubMed]
- Salamanna, F.; Cepollaro, S.; Contartese, D.; Giavaresi, G.; Barbanti Brodano, G.; Griffoni, C.; Gasbarrini, A.; Fini, M. Biological Rationale for the Use of Vertebral Whole Bone Marrow in Spinal Surgery. Spine 2018, 43, 1401–1410. [Google Scholar] [CrossRef] [PubMed]
- Salamanna, F.; Contartese, D.; Giavaresi, G.; Sicuro, L.; Barbanti Brodano, G.; Gasbarrini, A.; Fini, M. A Rationale for the Use of Clotted Vertebral Bone Marrow to Aid Tissue Regeneration Following Spinal Surgery. Sci. Rep. 2020, 10, 4115. [Google Scholar] [CrossRef] [PubMed]
- Salamanna, F.; Contartese, D.; Borsari, V.; Pagani, S.; Barbanti Brodano, G.; Griffoni, C.; Ricci, A.; Gasbarrini, A.; Fini, M. Two Hits for Bone Regeneration in Aged Patients: Vertebral Bone Marrow Clot as a Biological Scaffold and Powerful Source of Mesenchymal Stem Cells. Front. Bioeng. Biotechnol. 2022, 9, 807679. [Google Scholar] [CrossRef]
- Katsara, O.; Mahaira, L.G.; Iliopoulou, E.G.; Moustaki, A.; Antsaklis, A.; Loutradis, D.; Stefanidis, K.; Baxevanis, C.N.; Papamichail, M.; Perez, S.A. Effects of donor age, gender, and in vitro cellular aging on the phenotypic, functional, and molecular characteristics of mouse bone marrow-derived mesenchymal stem cells. Stem Cells Dev. 2011, 20, 1549–1561. [Google Scholar] [CrossRef]
- Strube, P.; Mehta, M.; Baerenwaldt, A.; Trippens, J.; Wilson, C.J.; Ode, A.; Perka, C.; Duda, G.N.; Kasper, G. Sex-specific compromised bone healing in female rats might be associated with a decrease in mesenchymal stem cell quantity. Bone 2009, 45, 1065–1072. [Google Scholar] [CrossRef]
- Tschon, M.; Contartese, D.; Pagani, S.; Borsari, V.; Fini, M. Gender and Sex Are Key Determinants in Osteoarthritis Not Only Confounding Variables. A Systematic Review of Clinical Data. J. Clin. Med. 2021, 10, 3178. [Google Scholar] [CrossRef]
- Contartese, D.; Tschon, M.; De Mattei, M.; Fini, M. Sex Specific Determinants in Osteoarthritis: A Systematic Review of Preclinical Studies. Int. J. Mol. Sci. 2020, 21, 3696. [Google Scholar] [CrossRef]
- Zaim, M.; Karaman, S.; Cetin, G.; Isik, S. Donor age and long-term culture affect differentiation and proliferation of human bone marrow mesenchymal stem cells. Ann. Hematol. 2012, 91, 1175–1186. [Google Scholar] [CrossRef]
- Corsi, K.A.; Pollett, J.B.; Phillippi, J.A.; Usas, A.; Li, G.; Huard, J. Osteogenic potential of postnatal skeletal muscle-derived stem cells is influenced by donor sex. J. Bone Miner. Res. 2007, 22, 1592–1602. [Google Scholar] [CrossRef]
- Rodríguez, J.P.; Montecinos, L.; Ríos, S.; Reyes, P.; Martínez, J. Mesenchymal stem cells from osteoporotic patients produce a type I collagen-deficient extracellular matrix favoring adipogenic differentiation. J. Cell Biochem. 2000, 79, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Sun, Y.; Zhang, Y. MiR-133 is Involved in Estrogen Deficiency-Induced Osteoporosis through Modulating Osteogenic Differentiation of Mesenchymal Stem Cells. Med. Sci. Monit. 2015, 21, 1527–1534. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yeung, D.K.W.; Griffith, J.F.; Antonio, G.E.; Lee, F.K.H.; Woo, J.; Leung, P.C. Osteoporosis is associated with increased marrow fat content and decreased marrow fat unsaturation: A proton MR spectroscopy study. J. Magn. Reson. Imaging 2005, 22, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.Z.; Yang, L.; Meng, G.L.; Lin, Y.S.; Wei, B.Y.; Fan, J.; Hu, H.M.; Liu, Y.W.; Chen, S.; Zhang, J.K.; et al. Estrogen improves the proliferation and differentiation of hBMSCs derived from postmenopausal osteoporosis through notch signaling pathway. Mol. Cell Biochem. 2014, 392, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells—current trends and future prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef]
- Nas, A.V.; Guhathakurta, D.; Wang, S.S.; Yehya, N.; Horvath, S.; Zhang, B.; Ingram-Drake, L.; Chaudhuri, G.; Schadt, E.E.; Drake, T.A.; et al. Elucidating the role of gonadal hormones in sexually dimorphic gene coexpression networks. Endocrinology 2009, 150, 1235–1249. [Google Scholar] [CrossRef]
- Williams, T.M.; Carroll, S.B. Genetic and molecular insights into the development and evolution of sexual dimorphism. Nat. Rev. Genet. 2009, 10, 797–804. [Google Scholar] [CrossRef]
- Palta, S.; Saroa, R.; Palta, A. Overview of the coagulation system. Indian. J. Anaesth. 2014, 58, 515–523. [Google Scholar] [CrossRef]
- Muschler, G.F.; Nitto, H.; Matsukura, Y.; Boehm, C.; Valdevit, A.; Kambic, H.; Davros, W.; Powell, K.; Easley, K. Spine fusion using cell matrix composites enriched in bone marrow-derived cells. Clin. Orthop. Relat. Res. 2003, 407, 102–118. [Google Scholar] [CrossRef]
- Assoian, R.K.; Komoriya, A.; Meyers, C.A.; Miller, D.M.; Sporn, M.B. Transforming growth factor-beta in human platelets. Identification of a major storage site, purification, and characterization. J. Biol. Chem. 1983, 258, 7155–7160. [Google Scholar] [CrossRef]
- Seyedin, S.M.; Thomas, T.C.; Thompson, A.Y.; Rosen, D.M.; Piez, K.A. Purification and characterization of two cartilage-inducing factors from bovine demineralized bone. Proc. Natl. Acad. Sci. USA 1985, 82, 2267–2271. [Google Scholar] [CrossRef]
- Nakashima, M.; Hari Reddi, A. The application of bone morphogenetic proteins to dental tissue engineering. Nat. Biotechnol. 2003, 21, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Yazawa, M.; Kishi, K.; Nakajima, H.; Nakajima, T. Expression of bone morphogenetic proteins during mandibular distraction osteogenesis in rabbits. J. Oral. Maxillofac. Surg. 2003, 61, 587–592. [Google Scholar] [CrossRef]
- Caplan, A.I. Review: Mesenchymal stem cells: Cell-based reconstructive therapy in orthopedics. Tissue Eng. 2005, 11, 1198–1211. [Google Scholar] [CrossRef]
- Bielby, R.; Jones, E.; McGonagle, D. The role of mesenchymal stem cells in maintenance and repair of bone. Injury 2007, 38 (Suppl. S1), S26–S32. [Google Scholar] [CrossRef]
- Wright, E.M.; Snopek, B.; Koopman, P. Seven new members of the Sox gene family expressed during mouse development. Nucleic Acids Res. 1993, 21, 744. [Google Scholar] [CrossRef] [PubMed]
- Picchi, J.; Trombi, L.; Spugnesi, L.; Barachini, S.; Maroni, G.; Barbanti Brodano, G.; Boriani, S.; Valtieri, M.; Petrini, M.; Magli, M.C. HOX and TALE signatures specify human stromal stem cell populations from different sources. J. Cell Physiol. 2013, 228, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ceballos, E.; Chambon, P.; Gudas, L.J. Differences in gene expression between wild type and Hoxa1 knockout embryonic stem cells after retinoic acid treatment or leukemia inhibitory factor (LIF) removal. J. Biol. Chem. 2005, 280, 16484–16498. [Google Scholar] [CrossRef]
- Rux, D.R.; Song, J.Y.; Swinehart, I.T.; Pineault, K.M.; Schlientz, A.J.; Trulik, K.G.; Goldstein, S.A.; Kozloff, K.M.; Lucas, D.; Wellik, D.M. Regionally Restricted Hox Function in Adult Bone Marrow Multipotent Mesenchymal Stem/Stromal Cells. Dev. Cell 2016, 39, 653–666. [Google Scholar] [CrossRef]
- Rux, D.R.; Song, J.Y.; Pineault, K.M.; Mandair, G.S.; Swinehart, I.T.; Schlientz, A.J.; Garthus, K.N.; Goldstein, S.A.; Kozloff, K.M.; Wellik, D.M. Hox11 Function Is Required for Region-Specific Fracture Repair. J. Bone Miner. Res. 2017, 32, 1750–1760. [Google Scholar] [CrossRef]
- Song, J.Y.; Pineault, K.M.; Dones, J.M.; Raines, R.T.; Wellik, D.M. Hox genes maintain critical roles in the adult skeleton. Proc. Natl. Acad. Sci. USA 2020, 117, 7296–7304. [Google Scholar] [CrossRef] [PubMed]
- Seifert, A.; Werheid, D.F.; Knapp, S.M.; Tobiasch, E. Role of Hox genes in stem cell differentiation. World J. Stem Cells 2015, 7, 583–595. [Google Scholar] [CrossRef] [PubMed]
- DiMartino, J.F.; Selleri, L.; Traver, D.; Firpo, M.T.; Rhee, J.; Warnke, R.; O′Gorman, S.; Weissman, I.L.; Cleary, M.L. The Hox cofactor and proto-oncogene Pbx1 is required for maintenance of definitive hematopoiesis in the fetal liver. Blood 2001, 98, 618–626. [Google Scholar] [CrossRef]
- Kim, H.Y.; Nelson, C.M. Extracellular matrix and cytoskeletal dynamics during branching morphogenesis. Organogenesis 2012, 8, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Manley, N.R.; Selleri, L.; Brendolan, A.; Gordon, J.; Cleary, M.L. Abnormalities of caudal pharyngeal pouch development in Pbx1 knockout mice mimic loss of Hox3 paralogs. Dev. Biol. 2004, 276, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, C.A.; Selleri, L.; Cleary, M.L. Pbx1 is essential for adrenal development and urogenital differentiation. Genesis 2003, 37, 123–130. [Google Scholar] [CrossRef]
- Schnabel, C.A.; Godin, R.E.; Cleary, M.L. Pbx1 regulates nephrogenesis and ureteric branching in the developing kidney. Dev. Biol. 2003, 254, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Selleri, L.; Depew, M.J.; Jacobs, Y.; Chanda, S.K.; Tsang, K.Y.; Cheah, K.S.; Rubenstein, J.L.; O′Gorman, S.; Cleary, M.L. Requirement for Pbx1 in skeletal patterning and programming chondrocyte proliferation and differentiation. Development 2001, 128, 3543–3557. [Google Scholar] [CrossRef]
- Moens, C.B.; Selleri, L. Hox cofactors in vertebrate development. Dev. Biol. 2006, 291, 193–206. [Google Scholar] [CrossRef]
- Forsberg, E.C.; Prohaska, S.S.; Katzman, S.; Heffner, G.C.; Stuart, J.M.; Weissman, I.L. Differential expression of novel potential regulators in hematopoietic stem cells. PLoS Genet. 2005, 1, e28. [Google Scholar] [CrossRef]
- Kiel, M.J.; Iwashita, T.; Yilmaz, O.H.; Morrison, S.J. Spatial differences in hematopoiesis but not in stem cells indicate a lack of regional patterning in definitive hematopoietic stem cells. Dev. Biol. 2005, 283, 29–39. [Google Scholar] [CrossRef]
- Johnson, P.M.; Knewtson, K.E.; Hodge, J.G.; Lehtinen, J.M.; Trofimoff, A.S.; Fritz, D.J.; Robinson, J.L. Surfactant location and internal phase volume fraction dictate emulsion electrospun fiber morphology and modulate drug release and cell response. Biomater. Sci. 2021, 9, 1397–1408. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, P.M. Gender differences in osteoporosis and fractures. Clin. Orthop. Relat. Res. 2011, 469, 1900–1905. [Google Scholar] [CrossRef] [PubMed]
- Beck, T.J.; Ruff, C.B.; Shaffer, R.A.; Betsinger, K.; Trone, D.W.; Brodine, S.K. Stress fracture in military recruits: Gender differences in muscle and bone susceptibility factors. Bone 2000, 27, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.A.; Bishop, A.T.; Moran, S.L.; Shin, A.Y. The outcomes and complications of 1,2-intercompartmental supraretinacular artery pedicled vascularized bone grafting of scaphoid nonunions. J. Hand Surg. Am. 2006, 31, 387–396. [Google Scholar] [CrossRef]
- David, V.; Lafage-Proust, M.H.; Laroche, N.; Christian, A.; Ruegsegger, P.; Vico, L. Two-week longitudinal survey of bone architecture alteration in the hindlimb-unloaded rat model of bone loss: Sex differences. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E440-7. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.T.; Lollar, P. Contribution of A1 subunit residue Q316 in thrombin-activated factor VIII to A2 subunit dissociation. Biochemistry 2007, 46, 9737–9742. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GENE | Primer Forward | Primer Reverse | Annealing Temperature |
|---|---|---|---|
| GAPDH | 5′-TGGTATCGTGGAAGGACTCA-3′ | 5′-GCAGGGATGATGTTCTGGA-3′ | 56 °C |
| ACAN | 5′-TCGAGGACAGCGAGGCC-3′ | 5′-TCGAGGGTGTAGCGTGTAGAGA-3′ | 60 °C |
| SOX 9 | 5′-GAGCAGACGCACATCTC-3′ | 5′-CCTGGGATTGCCCCGA-3′ | 60 °C |
| COL2A1 | QuantiTect Primer Assay (Qiagen) Hs_COL2A1_1_SG | 55 °C | |
| COL1A1 | QuantiTect Primer Assay (Qiagen) Hs_COL1A1_1_SG | 55 °C | |
| ALPL | QuantiTect Primer Assay (Qiagen) Hs_ALPL_1_SG | 55 °C | |
| BGLAP | QuantiTect Primer Assay (Qiagen) Hs_BGLAP_1_SG | 55 °C | |
| RUNX2 | QuantiTect Primer Assay (Qiagen) Hs_RUNX2_1_SG | 55 °C | |
| PPARg | QuantiTect Primer Assay (Qiagen) Hs_PPARG_1_SG | 55 °C | |
| ADIPOQ | QuantiTect Primer Assay (Qiagen) Hs_ADIPOQ_1_SG | 55 °C | |
| IL1A | QuantiTect Primer Assay (Qiagen) Hs_IL1A_1_SG | 55 °C | |
| IL 1B | QuantiTect Primer Assay (Qiagen) Hs_IL1B_1_SG | 55 °C | |
| IL 6 | QuantiTect Primer Assay (Qiagen) Hs_IL6_1_SG | 55 °C | |
| CCL4L1 (MIP-1β) | QuantiTect Primer Assay (Qiagen) Hs_CCL4L1_4_SG | 55 °C | |
| CXCL2 (MIP-2α) | QuantiTect Primer Assay (Qiagen) Hs_CXCL2_1_SG | 55 °C | |
| KL (Klotho) | QuantiTect Primer Assay (Qiagen) Hs_KL_1_SG | 55 °C | |
| TNFα | QuantiTect Primer Assay (Qiagen) Hs_TNF_1_SG | 55 °C | |
| MCP-1 | QuantiTect Primer Assay (Qiagen) Hs_CCL2_1_SG | 55 °C | |
| IL8 | 5′-ATGACTTCCAAGCTGGCCGTG-3′ | 5′-TTATGAATTCTCAGCCCTCTTCAAAAACTTCTC-3′ | 60 °C |
| HOXA1 | Hs_HOXA1_1_SG | 55 °C 20″ | |
| HOXA6 | Hs_HOXA6_1_SG | 55 °C 20″ | |
| HOXA7 | Hs_HOXA7_2_SG | 55 °C 20″ | |
| HOXA11 | Hs_HOXA11_2_SG | 55 °C 20″ | |
| HOXB8 | Hs_HOXB8_2_SG | 55 °C 20″ | |
| HOXB9 | Hs_HOXB9_1_SG | 55 °C 20″ | |
| HOXD9 | Hs_HOXD9_1_SG | 55 °C 20″ | |
| HOXD10 | Hs_HOXD10_1_SG | 55 °C 20″ | |
| MEIS2 | Hs_MEIS2_1_SG | 55 °C 20″ | |
| MEIS3 | Hs_MEIS3_1_SG | 55 °C 20″ | |
| PBX1 | Hs_PBX1_1_SG | 55 °C 20″ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salamanna, F.; Contartese, D.; Borsari, V.; Pagani, S.; Sartori, M.; Tschon, M.; Griffoni, C.; Giavaresi, G.; Tedesco, G.; Barbanti Brodano, G.; et al. Gender-Specific Differences in Human Vertebral Bone Marrow Clot. Int. J. Mol. Sci. 2023, 24, 11856. https://doi.org/10.3390/ijms241411856
Salamanna F, Contartese D, Borsari V, Pagani S, Sartori M, Tschon M, Griffoni C, Giavaresi G, Tedesco G, Barbanti Brodano G, et al. Gender-Specific Differences in Human Vertebral Bone Marrow Clot. International Journal of Molecular Sciences. 2023; 24(14):11856. https://doi.org/10.3390/ijms241411856
Chicago/Turabian StyleSalamanna, Francesca, Deyanira Contartese, Veronica Borsari, Stefania Pagani, Maria Sartori, Matilde Tschon, Cristiana Griffoni, Gianluca Giavaresi, Giuseppe Tedesco, Giovanni Barbanti Brodano, and et al. 2023. "Gender-Specific Differences in Human Vertebral Bone Marrow Clot" International Journal of Molecular Sciences 24, no. 14: 11856. https://doi.org/10.3390/ijms241411856
APA StyleSalamanna, F., Contartese, D., Borsari, V., Pagani, S., Sartori, M., Tschon, M., Griffoni, C., Giavaresi, G., Tedesco, G., Barbanti Brodano, G., Gasbarrini, A., & Fini, M. (2023). Gender-Specific Differences in Human Vertebral Bone Marrow Clot. International Journal of Molecular Sciences, 24(14), 11856. https://doi.org/10.3390/ijms241411856