
Epigenome-Wide Associations of Placental DNA Methylation and Behavioral and Emotional Difficulties in Children at 3 Years of Age

 ,
,  ,
,  , , , ,
, , , ,
Abstract
1. Introduction
2. Results
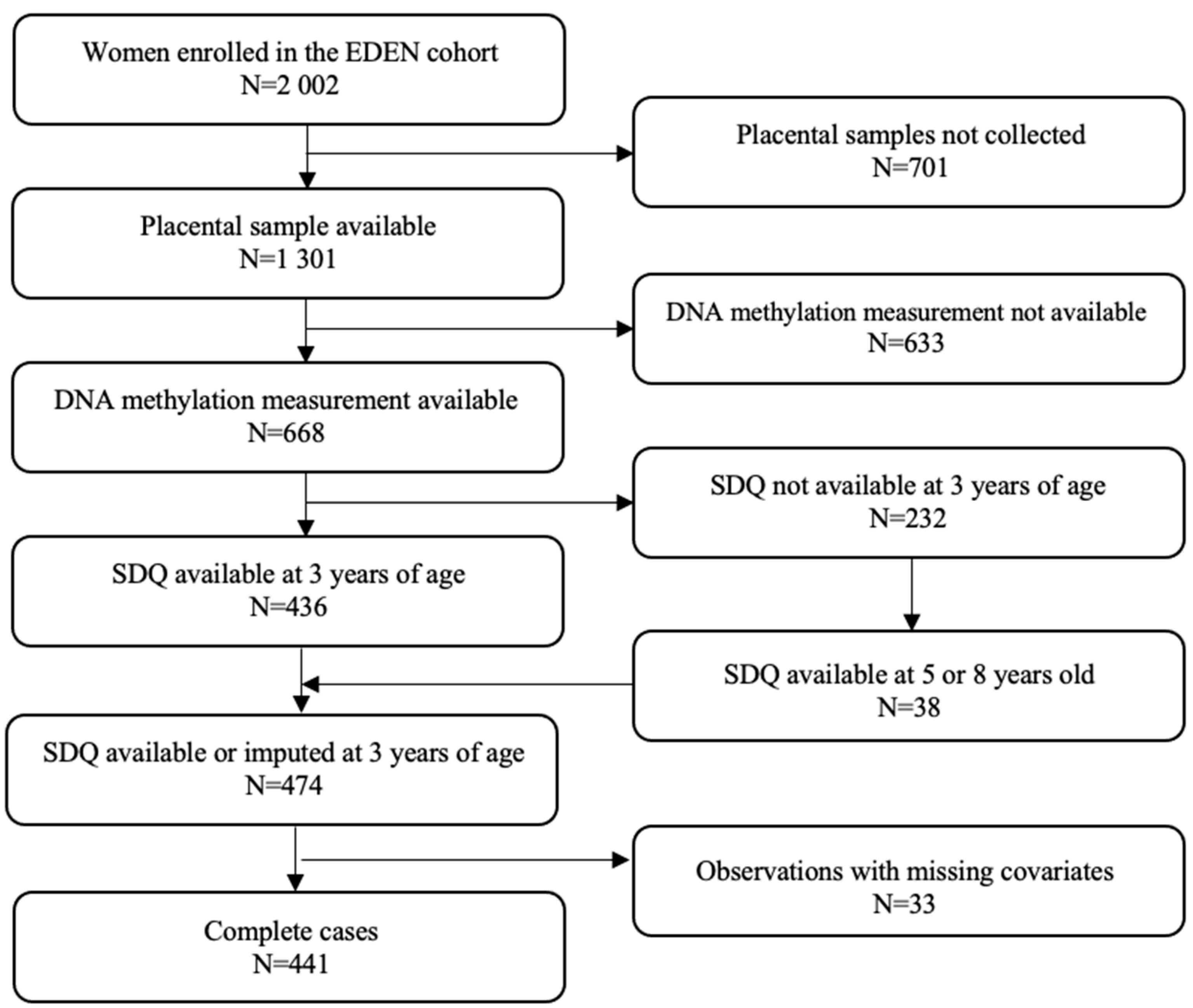
2.1. Characteristics of Study Participants
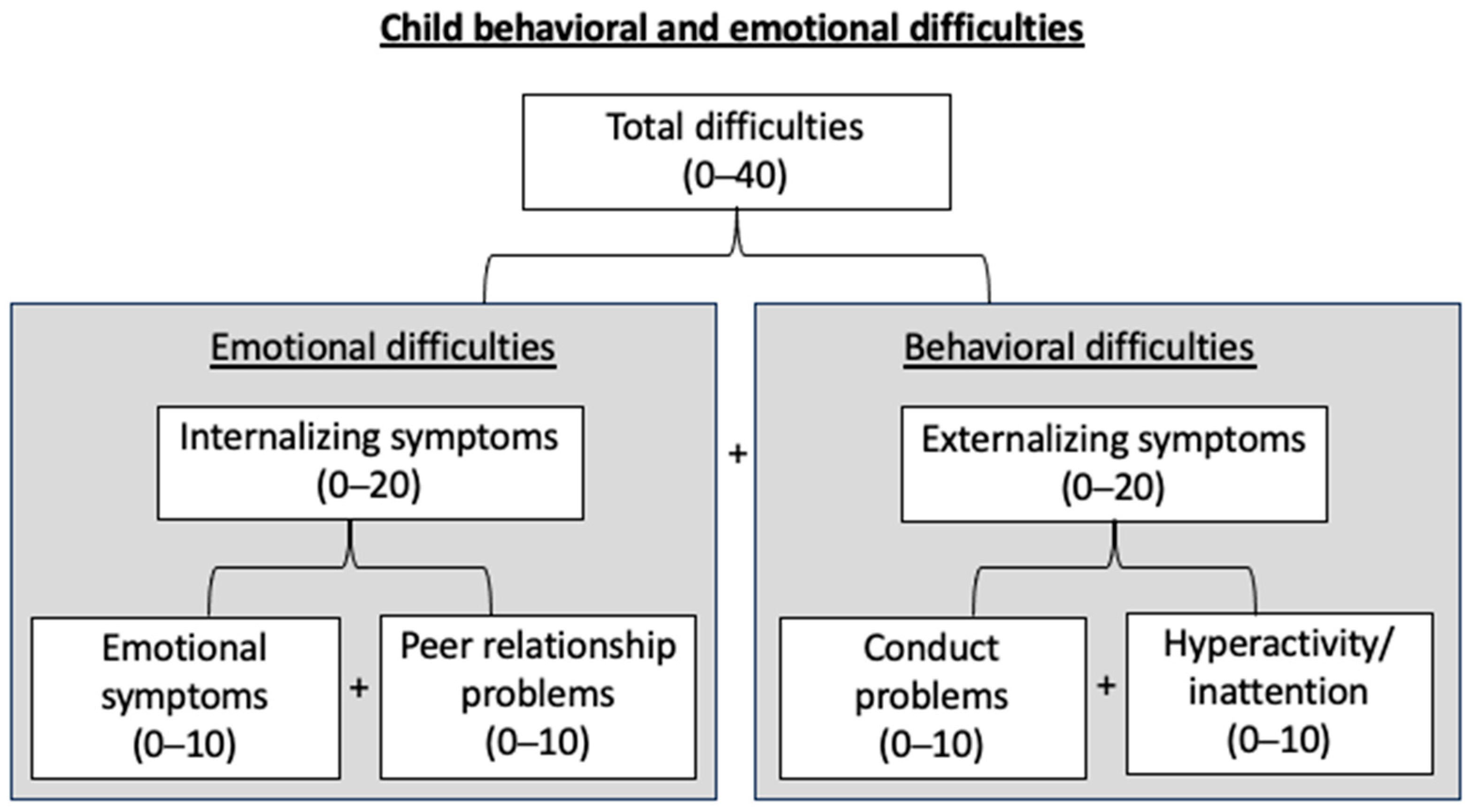
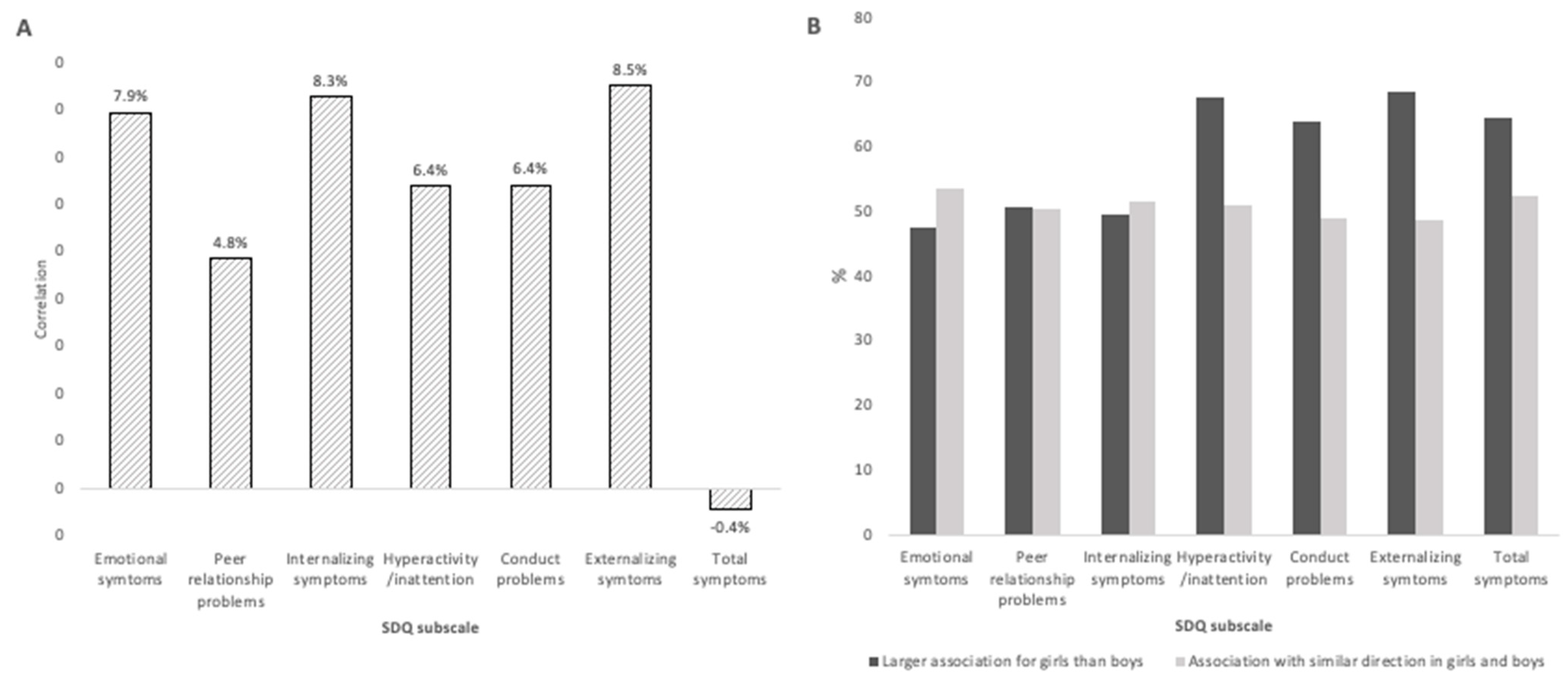
2.2. Emotional and Behavioral Difficulties at 3 years of age
2.3. Hypothesis-Driven Analyses
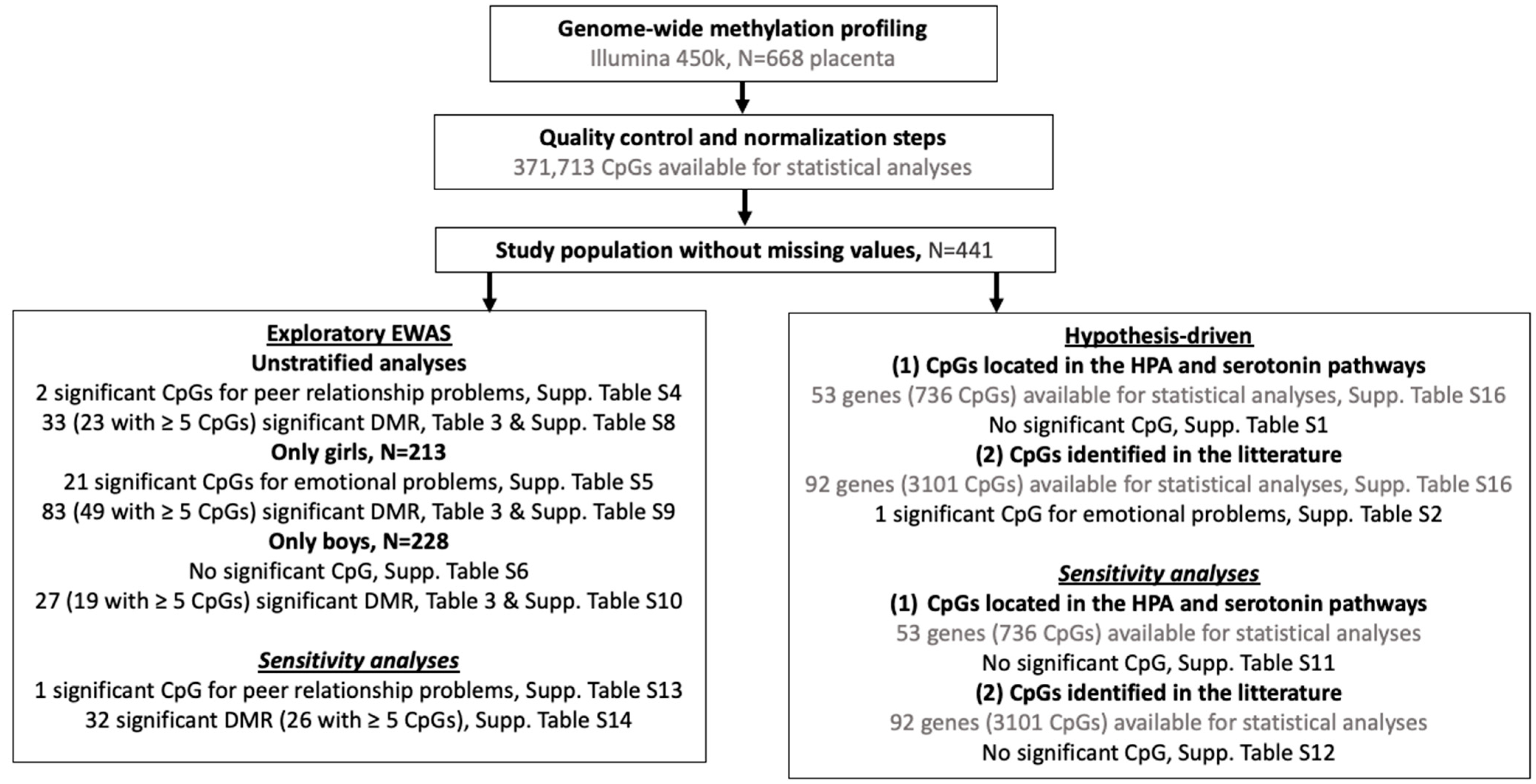
2.4. Epigenome-Wide Association Study Analyses (EWAS)
2.5. Differentially Methylated Regional (DMR) Analyses
2.6. Sensitivity Analyses
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Child’s Emotional and Behavioral Difficulties at 3 Years of Age
4.3. Placental DNA Methylation Levels
4.4. Covariates
4.5. Cellular Heterogeneity of Placenta Samples
4.6. Statistical Analyses
4.7. Sensitivity Analyses
4.8. Softwares
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bagner, D.M.; Rodríguez, G.M.; Blake, C.A.; Linares, D.; Carter, A.S. Assessment of Behavioral and Emotional Problems in Infancy: A Systematic Review. Clin. Child Fam. Psychol. Rev. 2012, 15, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Ogundele, M.O. Behavioural and Emotional Disorders in Childhood: A Brief Overview for Paediatricians. World J. Clin. Pediatr. 2018, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Barican, J.L.; Yung, D.; Schwartz, C.; Zheng, Y.; Georgiades, K.; Waddell, C. Prevalence of Childhood Mental Disorders in High-Income Countries: A Systematic Review and Meta-Analysis to Inform Policymaking. Evid.-Based Ment. Health 2022, 25, 36–44. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Data and Statistics on Children’s Mental Health. Cent. Dis. Control Prev. 2019, 13, 2019. [Google Scholar]
- Barker, D.J. The Origins of the Developmental Origins Theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Scher, M.S. “The First Thousand Days” Define a Fetal/Neonatal Neurology Program. Front. Pediatr. 2021, 9, 683138. [Google Scholar] [CrossRef]
- Selevan, S.G.; Kimmel, C.A.; Mendola, P. Identifying Critical Windows of Exposure for Children’s Health. Environ. Health Perspect. 2000, 108, 451–455. [Google Scholar]
- Victora, C.G.; De Onis, M.; Hallal, P.C.; Blössner, M.; Shrimpton, R. Worldwide Timing of Growth Faltering: Revisiting Implications for Interventions. Pediatrics 2010, 125, e473–e480. [Google Scholar] [CrossRef]
- Guilbert, A.; Rolland, M.; Pin, I.; Thomsen, C.; Sakhi, A.K.; Sabaredzovic, A.; Slama, R.; Guichardet, K.; Philippat, C. Associations between a Mixture of Phenols and Phthalates and Child Behaviour in a French Mother–Child Cohort with Repeated Assessment of Exposure. Environ. Int. 2021, 156, 106697. [Google Scholar] [CrossRef]
- Jedynak, P.; Maitre, L.; Guxens, M.; Gützkow, K.B.; Julvez, J.; López-Vicente, M.; Sunyer, J.; Casas, M.; Chatzi, L.; Gražulevičienė, R. Prenatal Exposure to a Wide Range of Environmental Chemicals and Child Behaviour between 3 and 7 Years of Age–An Exposome-Based Approach in 5 European Cohorts. Sci. Total Environ. 2021, 763, 144115. [Google Scholar] [CrossRef]
- Maitre, L.; Julvez, J.; López-Vicente, M.; Warembourg, C.; Tamayo-Uria, I.; Philippat, C.; Gützkow, K.B.; Guxens, M.; Andrusaityte, S.; Basagaña, X. Early-Life Environmental Exposure Determinants of Child Behavior in Europe: A Longitudinal, Population-Based Study. Environ. Int. 2021, 153, 106523. [Google Scholar] [CrossRef] [PubMed]
- Fekom, M.; Bonello, K.; Gomajee, R.; Ibanez, G.; Martin, S.; Keyes, K.; Nakamura, A.; Lepeule, J.; Strandberg-Larsen, K.; Melchior, M. Smoking during Pregnancy and Children’s Emotional and Behavioural Trajectories: Mathilde Fekom. Eur. J. Public Health 2022, 32, ckac129-669. [Google Scholar] [CrossRef]
- Tsang, T.W.; Lucas, B.R.; Carmichael Olson, H.; Pinto, R.Z.; Elliott, E.J. Prenatal Alcohol Exposure, FASD, and Child Behavior: A Meta-Analysis. Pediatrics 2016, 137, e20152542. [Google Scholar] [CrossRef]
- Van Der Waerden, J.; Galéra, C.; Larroque, B.; Saurel-Cubizolles, M.-J.; Sutter-Dallay, A.-L.; Melchior, M.; EDEN Mother–Child Cohort Study Group. Maternal Depression Trajectories and Children’s Behavior at Age 5 Years. J. Pediatr. 2015, 166, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Turco, M.Y.; Moffett, A. Development of the Human Placenta. Development 2019, 146, dev163428. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.; Rousseaux, S.; Agier, L.; Giorgis-Allemand, L.; Tost, J.; Galineau, J.; Hulin, A.; Siroux, V.; Vaiman, D.; Charles, M.-A. Pregnancy Exposure to Atmospheric Pollution and Meteorological Conditions and Placental DNA Methylation. Environ. Int. 2018, 118, 334–347. [Google Scholar] [CrossRef]
- Broséus, L.; Vaiman, D.; Tost, J.; Martin, C.R.S.; Jacobi, M.; Schwartz, J.D.; Béranger, R.; Slama, R.; Heude, B.; Lepeule, J. Maternal Blood Pressure Associates with Placental DNA Methylation Both Directly and through Alterations in Cell-Type Composition. BMC Med. 2022, 20, 397. [Google Scholar] [CrossRef]
- Heijmans, B.T.; Tobi, E.W.; Lumey, L.; Slagboom, P.E. The Epigenome: Archive of the Prenatal Environment. Epigenetics 2009, 4, 526–531. [Google Scholar] [CrossRef]
- Jedynak, P.; Tost, J.; Calafat, A.M.; Bourova-Flin, E.; Busato, F.; Forhan, A.; Heude, B.; Jakobi, M.; Rousseaux, S.; Schwartz, J. Pregnancy Exposure to Synthetic Phenols and Placental DNA Methylation-An Epigenome-Wide Association Study in Male Infants from the EDEN Cohort. Environ. Pollut. 2021, 290, 118024. [Google Scholar] [CrossRef]
- Nakamura, A.; François, O.; Lepeule, J. Epigenetic Alterations of Maternal Tobacco Smoking during Pregnancy: A Narrative Review. Int. J. Environ. Res. Public Health 2021, 18, 5083. [Google Scholar] [CrossRef]
- Rousseaux, S.; Seyve, E.; Chuffart, F.; Bourova-Flin, E.; Benmerad, M.; Charles, M.-A.; Forhan, A.; Heude, B.; Siroux, V.; Slama, R. Immediate and Durable Effects of Maternal Tobacco Consumption Alter Placental DNA Methylation in Enhancer and Imprinted Gene-Containing Regions. BMC Med. 2020, 18, 306. [Google Scholar] [CrossRef] [PubMed]
- Lesseur, C.; Armstrong, D.A.; Murphy, M.A.; Appleton, A.A.; Koestler, D.C.; Paquette, A.G.; Lester, B.M.; Marsit, C.J. Sex-Specific Associations between Placental Leptin Promoter DNA Methylation and Infant Neurobehavior. Psychoneuroendocrinology 2014, 40, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Marsit, C.J.; Maccani, M.A.; Padbury, J.F.; Lester, B.M. Placental 11-Beta Hydroxysteroid Dehydrogenase Methylation Is Associated with Newborn Growth and a Measure of Neurobehavioral Outcome. PLoS ONE 2012, 7, e33794. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S. The Placenta-brain-axis. J. Neurosci. Res. 2021, 99, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Zeltser, L.M.; Leibel, R.L. Roles of the Placenta in Fetal Brain Development. Proc. Natl. Acad. Sci. USA 2011, 108, 15667–15668. [Google Scholar] [CrossRef]
- Jensen Peña, C.L.; Champagne, F.A. Epigenetic and Neurodevelopmental Perspectives on Variation in Parenting Behavior. Parenting 2012, 12, 202–211. [Google Scholar] [CrossRef]
- Bonnin, A.; Levitt, P. Fetal, Maternal, and Placental Sources of Serotonin and New Implications for Developmental Programming of the Brain. Neuroscience 2011, 197, 1–7. [Google Scholar] [CrossRef]
- Novakovic, B.; Ryan, J.; Pereira, N.; Boughton, B.; Craig, J.M.; Saffery, R. Postnatal Stability, Tissue, and Time Specific Effects of AHRR Methylation Change in Response to Maternal Smoking in Pregnancy. Epigenetics 2014, 9, 377–386. [Google Scholar] [CrossRef]
- Appleton, A.A.; Lester, B.M.; Armstrong, D.A.; Lesseur, C.; Marsit, C.J. Examining the Joint Contribution of Placental NR3C1 and HSD11B2 Methylation for Infant Neurobehavior. Psychoneuroendocrinology 2015, 52, 32–42. [Google Scholar] [CrossRef]
- Breton, E.; Gagné-Ouellet, V.; Thibeault, K.; Guérin, R.; Van Lieshout, R.; Perron, P.; Hivert, M.; Bouchard, L. Placental NEGR1 DNA Methylation Is Associated with BMI and Neurodevelopment in Preschool-Age Children. Epigenetics 2020, 15, 323–335. [Google Scholar] [CrossRef]
- Bromer, C.; Marsit, C.J.; Armstrong, D.A.; Padbury, J.F.; Lester, B. Genetic and Epigenetic Variation of the Glucocorticoid Receptor (NR3C1) in Placenta and Infant Neurobehavior. Dev. Psychobiol. 2013, 55, 673–683. [Google Scholar] [CrossRef]
- Paquette, A.G.; Lester, B.M.; Lesseur, C.; Armstrong, D.A.; Guerin, D.J.; Appleton, A.A.; Marsit, C.J. Placental Epigenetic Patterning of Glucocorticoid Response Genes Is Associated with Infant Neurodevelopment. Epigenomics 2015, 7, 767–779. [Google Scholar] [CrossRef]
- Paquette, A.G.; Houseman, E.A.; Green, B.B.; Lesseur, C.; Armstrong, D.A.; Lester, B.; Marsit, C.J. Regions of Variable DNA Methylation in Human Placenta Associated with Newborn Neurobehavior. Epigenetics 2016, 11, 603–613. [Google Scholar] [CrossRef]
- Stroud, L.R.; Papandonatos, G.D.; Salisbury, A.L.; Phipps, M.G.; Huestis, M.A.; Niaura, R.; Padbury, J.F.; Marsit, C.J.; Lester, B.M. Epigenetic Regulation of Placental NR3C1: Mechanism Underlying Prenatal Programming of Infant Neurobehavior by Maternal Smoking? Child Dev. 2016, 87, 49–60. [Google Scholar] [CrossRef]
- Zhu, Y.; Mordaunt, C.E.; Yasui, D.H.; Marathe, R.; Coulson, R.L.; Dunaway, K.W.; Jianu, J.M.; Walker, C.K.; Ozonoff, S.; Hertz-Picciotto, I. Placental DNA Methylation Levels at CYP2E1 and IRS2 Are Associated with Child Outcome in a Prospective Autism Study. Hum. Mol. Genet. 2019, 28, 2659–2674. [Google Scholar] [CrossRef]
- Holmes, M.C.; Abrahamsen, C.T.; French, K.L.; Paterson, J.M.; Mullins, J.J.; Seckl, J.R. The Mother or the Fetus? 11β-Hydroxysteroid Dehydrogenase Type 2 Null Mice Provide Evidence for Direct Fetal Programming of Behavior by Endogenous Glucocorticoids. J. Neurosci. 2006, 26, 3840–3844. [Google Scholar]
- Wyrwoll, C.S.; Holmes, M.C. Prenatal Excess Glucocorticoid Exposure and Adult Affective Disorders: A Role for Serotonergic and Catecholamine Pathways. Neuroendocrinology 2012, 95, 47–55. [Google Scholar] [CrossRef]
- Andrews, S.V.; Yang, I.J.; Froehlich, K.; Oskotsky, T.; Sirota, M. Large-Scale Placenta DNA Methylation Integrated Analysis Reveals Fetal Sex-Specific Differentially Methylated CpG Sites and Regions. Sci. Rep. 2022, 12, 9396. [Google Scholar] [CrossRef]
- Kaczkurkin, A.N.; Raznahan, A.; Satterthwaite, T.D. Sex Differences in the Developing Brain: Insights from Multimodal Neuroimaging. Neuropsychopharmacology 2019, 44, 71–85. [Google Scholar] [CrossRef]
- Polanczyk, G.; De Lima, M.S.; Horta, B.L.; Biederman, J.; Rohde, L.A. The Worldwide Prevalence of ADHD: A Systematic Review and Metaregression Analysis. Am. J. Psychiatry 2007, 164, 942–948. [Google Scholar] [CrossRef]
- Caramaschi, D.; Neumann, A.; Cardenas, A.; Tindula, G.; Alemany, S.; Zillich, L.; Pesce, G.; Lahti, J.M.; Havdahl, A.; Mulder, R. Meta-Analysis of Epigenome-Wide Associations between DNA Methylation at Birth and Childhood Cognitive Skills. Mol. Psychiatry 2022, 27, 2126–2135. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, A.; Rifas-Shiman, S.L.; Agha, G.; Hivert, M.-F.; Litonjua, A.A.; DeMeo, D.L.; Lin, X.; Amarasiriwardena, C.J.; Oken, E.; Gillman, M.W. Persistent DNA Methylation Changes Associated with Prenatal Mercury Exposure and Cognitive Performance during Childhood. Sci. Rep. 2017, 7, 288. [Google Scholar] [CrossRef] [PubMed]
- Cecil, C.A.; Walton, E.; Jaffee, S.R.; O’Connor, T.; Maughan, B.; Relton, C.L.; Smith, R.G.; McArdle, W.; Gaunt, T.R.; Ouellet-Morin, I. Neonatal DNA Methylation and Early-Onset Conduct Problems: A Genome-Wide, Prospective Study. Dev. Psychopathol. 2018, 30, 383–397. [Google Scholar] [CrossRef]
- Hodyl, N.A.; Roberts, C.T.; Bianco-Miotto, T. Cord Blood DNA Methylation Biomarkers for Predicting Neurodevelopmental Outcomes. Genes 2016, 7, 117. [Google Scholar] [CrossRef]
- Rijlaarsdam, J.; Barker, E.D.; Caserini, C.; Koopman-Verhoeff, M.E.; Mulder, R.H.; Felix, J.F.; Cecil, C.A. Genome-Wide DNA Methylation Patterns Associated with General Psychopathology in Children. J. Psychiatr. Res. 2021, 140, 214–220. [Google Scholar] [CrossRef]
- Everson, T.M.; Vives-Usano, M.; Seyve, E.; Cardenas, A.; Lacasaña, M.; Craig, J.M.; Lesseur, C.; Baker, E.R.; Fernandez-Jimenez, N.; Heude, B. Placental DNA Methylation Signatures of Maternal Smoking during Pregnancy and Potential Impacts on Fetal Growth. Nat. Commun. 2021, 12, 5095. [Google Scholar] [CrossRef]
- Joubert, B.R.; Håberg, S.E.; Nilsen, R.M.; Wang, X.; Vollset, S.E.; Murphy, S.K.; Huang, Z.; Hoyo, C.; Midttun, Ø.; Cupul-Uicab, L.A. 450K Epigenome-Wide Scan Identifies Differential DNA Methylation in Newborns Related to Maternal Smoking during Pregnancy. Environ. Health Perspect. 2012, 120, 1425–1431. [Google Scholar] [CrossRef]
- Joubert, B.R.; Felix, J.F.; Yousefi, P.; Bakulski, K.M.; Just, A.C.; Breton, C.; Reese, S.E.; Markunas, C.A.; Richmond, R.C.; Xu, C.-J. DNA Methylation in Newborns and Maternal Smoking in Pregnancy: Genome-Wide Consortium Meta-Analysis. Am. J. Hum. Genet. 2016, 98, 680–696. [Google Scholar] [CrossRef] [PubMed]
- Knopik, V.S.; Marceau, K.; Bidwell, L.C.; Rolan, E. Prenatal Substance Exposure and Offspring Development: Does DNA Methylation Play a Role? Neurotoxicol. Teratol. 2019, 71, 50–63. [Google Scholar] [CrossRef]
- Katrinli, S.; Maihofer, A.X.; Wani, A.H.; Pfeiffer, J.R.; Ketema, E.; Ratanatharathorn, A.; Baker, D.G.; Boks, M.P.; Geuze, E.; Kessler, R.C. Epigenome-Wide Meta-Analysis of PTSD Symptom Severity in Three Military Cohorts Implicates DNA Methylation Changes in Genes Involved in Immune System and Oxidative Stress. Mol. Psychiatry 2022, 27, 1720–1728. [Google Scholar] [CrossRef]
- Smith, A.K.; Ratanatharathorn, A.; Maihofer, A.X.; Naviaux, R.K.; Aiello, A.E.; Amstadter, A.B.; Ashley-Koch, A.E.; Baker, D.G.; Beckham, J.C.; Boks, M.P. Epigenome-Wide Meta-Analysis of PTSD across 10 Military and Civilian Cohorts Identifies Methylation Changes in AHRR. Nat. Commun. 2020, 11, 5965. [Google Scholar] [CrossRef]
- Brew, O.; Sullivan, M.H.; Woodman, A. Comparison of Normal and Pre-Eclamptic Placental Gene Expression: A Systematic Review with Meta-Analysis. PLoS ONE 2016, 11, e0161504. [Google Scholar] [CrossRef] [PubMed]
- Eckmann, M.; Sheng, Q.; Baldwin, S.; Lister, R.L. Maternal Hyperglycemia Induces Changes in Gene Expression and Morphology in Mouse Placentas. Gynecol. Reprod. Health 2020, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Clarkson-Townsend, D.A.; Everson, T.M.; Deyssenroth, M.A.; Burt, A.A.; Hermetz, K.E.; Hao, K.; Chen, J.; Marsit, C.J. Maternal Circadian Disruption Is Associated with Variation in Placental DNA Methylation. PLoS ONE 2019, 14, e0215745. [Google Scholar] [CrossRef] [PubMed]
- Alliance of Genome Resources Consortium. The Alliance of Genome Resources: Building a Modern Data Ecosystem for Model Organism Databases. Genetics 2019, 213, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Safran, M.; Dalah, I.; Alexander, J.; Rosen, N.; Iny Stein, T.; Shmoish, M.; Nativ, N.; Bahir, I.; Doniger, T.; Krug, H. GeneCards Version 3: The Human Gene Integrator. Database 2010, 2010, baq020. [Google Scholar] [CrossRef]
- Hu, Y.; O’Boyle, K.; Auer, J.; Raju, S.; You, F.; Wang, P.; Fikrig, E.; Sutton, R.E. Multiple UBXN Family Members Inhibit Retrovirus and Lentivirus Production and Canonical NFκΒ Signaling by Stabilizing IκBα. PLoS Pathog. 2017, 13, e1006187. [Google Scholar] [CrossRef]
- Tekola-Ayele, F.; Zeng, X.; Ouidir, M.; Workalemahu, T.; Zhang, C.; Delahaye, F.; Wapner, R. DNA Methylation Loci in Placenta Associated with Birthweight and Expression of Genes Relevant for Early Development and Adult Diseases. Clin. Epigenet. 2020, 12, 78. [Google Scholar] [CrossRef]
- Chaplin, T.M.; Aldao, A. Gender Differences in Emotion Expression in Children: A Meta-Analytic Review. Psychol. Bull. 2013, 139, 735. [Google Scholar] [CrossRef]
- Bozack, A.K.; Colicino, E.; Just, A.C.; Wright, R.O.; Baccarelli, A.A.; Wright, R.J.; Lee, A.G. Associations between Infant Sex and DNA Methylation across Umbilical Cord Blood, Artery, and Placenta Samples. Epigenetics 2021, 17, 1080–1097. [Google Scholar] [CrossRef]
- Inkster, A.M.; Yuan, V.; Konwar, C.; Matthews, A.M.; Brown, C.J.; Robinson, W.P. A Cross-Cohort Analysis of Autosomal DNA Methylation Sex Differences in the Term Placenta. Biol. Sex Differ. 2021, 12, 38. [Google Scholar] [CrossRef]
- Martin, E.; Smeester, L.; Bommarito, P.A.; Grace, M.R.; Boggess, K.; Kuban, K.; Karagas, M.R.; Marsit, C.J.; O’Shea, T.M.; Fry, R.C. Sexual Epigenetic Dimorphism in the Human Placenta: Implications for Susceptibility during the Prenatal Period. Epigenomics 2017, 9, 267–278. [Google Scholar] [CrossRef]
- López-Ojeda, W.; Hurley, R.A. Sexual Dimorphism in Brain Development: Influence on Affective Disorders. J. Neuropsychiatry Clin. Neurosci. 2021, 33, A4-85. [Google Scholar] [CrossRef]
- Pallayova, M.; Brandeburova, A.; Tokarova, D. Update on Sexual Dimorphism in Brain Structure–Function Interrelationships: A Literature Review. Appl. Psychophysiol. Biofeedback 2019, 44, 271–284. [Google Scholar] [CrossRef]
- Gabory, A.; Attig, L.; Junien, C. Developmental Programming and Epigenetics. Am. J. Clin. Nutr. 2011, 94, 1943S–1952S. [Google Scholar] [CrossRef]
- Rosenfeld, C.S.; Trainor, B.C. Environmental Health Factors and Sexually Dimorphic Differences in Behavioral Disruptions. Curr. Environ. Health Rep. 2014, 1, 287–301. [Google Scholar] [CrossRef]
- Bronson, S.L.; Bale, T.L. The Placenta as a Mediator of Stress Effects on Neurodevelopmental Reprogramming. Neuropsychopharmacology 2016, 41, 207–218. [Google Scholar] [CrossRef]
- Burton, G.J.; Fowden, A.L.; Thornburg, K.L. Placental Origins of Chronic Disease. Physiol. Rev. 2016, 96, 1509–1565. [Google Scholar] [CrossRef]
- Everson, T.; Vives-Usano, M.; Craig, J.; Hivert, M.; Karagas, M.; Lacasaña, M.; Lepeule, J.; Marsit, C.; Bustamante, M. Placental Methylation Signatures from Maternal Smoking during Pregnancy and Potential Impacts on Fetal Growth: Meta-Analyses from 7 Cohorts. Environ. Epidemiol. 2019, 3, 115. [Google Scholar]
- Niculescu, A.; Levey, D.; Phalen, P.; Le-Niculescu, H.; Dainton, H.; Jain, N.; Belanger, E.; James, A.; George, S.; Weber, H. Understanding and Predicting Suicidality Using a Combined Genomic and Clinical Risk Assessment Approach. Mol. Psychiatry 2015, 20, 1266–1285. [Google Scholar] [CrossRef]
- Lionel, A.C.; Crosbie, J.; Barbosa, N.; Goodale, T.; Thiruvahindrapuram, B.; Rickaby, J.; Gazzellone, M.; Carson, A.R.; Howe, J.L.; Wang, Z. Rare Copy Number Variation Discovery and Cross-Disorder Comparisons Identify Risk Genes for ADHD. Sci. Transl. Med. 2011, 3, 95ra75. [Google Scholar] [CrossRef]
- Sikdar, S.; Joehanes, R.; Joubert, B.R.; Xu, C.-J.; Vives-Usano, M.; Rezwan, F.I.; Felix, J.F.; Ward, J.M.; Guan, W.; Richmond, R.C. Comparison of Smoking-Related DNA Methylation between Newborns from Prenatal Exposure and Adults from Personal Smoking. Epigenomics 2019, 11, 1487–1500. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Wan, E.; Morrow, J.; Cho, M.H.; Crapo, J.D.; Silverman, E.K.; DeMeo, D.L. The Impact of Genetic Variation and Cigarette Smoke on DNA Methylation in Current and Former Smokers from the COPDGene Study. Epigenetics 2015, 10, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, J.D.; Huang, R.-C.; Barton, S.J.; Saffery, R.; Lillycrop, K.A. Is Cellular Heterogeneity Merely a Confounder to Be Removed from Epigenome-Wide Association Studies? Epigenomics 2017, 9, 1143–1150. [Google Scholar] [CrossRef]
- Konwar, C.; Del Gobbo, G.F.; Terry, J.; Robinson, W.P. Association of a Placental Interleukin-6 Genetic Variant (Rs1800796) with DNA Methylation, Gene Expression and Risk of Acute Chorioamnionitis. BMC Med. Genet. 2019, 20, 36. [Google Scholar] [CrossRef] [PubMed]
- Houseman, E.A.; Kile, M.L.; Christiani, D.C.; Ince, T.A.; Kelsey, K.T.; Marsit, C.J. Reference-Free Deconvolution of DNA Methylation Data and Mediation by Cell Composition Effects. BMC Bioinform. 2016, 17, 259. [Google Scholar] [CrossRef] [PubMed]
- Yuan, V.; Hui, D.; Yin, Y.; Peñaherrera, M.S.; Beristain, A.G.; Robinson, W.P. Cell-Specific Characterization of the Placental Methylome. BMC Genom. 2021, 22, 6. [Google Scholar] [CrossRef]
- Dieckmann, L.; Cruceanu, C.; Lahti-Pulkkinen, M.; Lahti, J.; Kvist, T.; Laivuori, H.; Sammallahti, S.; Villa, P.M.; Suomalainen-König, S.; Rancourt, R.C. Reliability of a Novel Approach for Reference-Based Cell Type Estimation in Human Placental DNA Methylation Studies. Cell. Mol. Life Sci. 2022, 79, 115. [Google Scholar] [CrossRef]
- Yousefi, P.; Huen, K.; Davé, V.; Barcellos, L.; Eskenazi, B.; Holland, N. Sex Differences in DNA Methylation Assessed by 450 K BeadChip in Newborns. BMC Genom. 2015, 16, 911. [Google Scholar] [CrossRef]
- Shojaei, T.; Wazana, A.; Pitrou, I.; Kovess, V. The Strengths and Difficulties Questionnaire: Validation Study in French School-Aged Children and Cross-Cultural Comparisons. Soc. Psychiatry Psychiatr. Epidemiol. 2009, 44, 740–747. [Google Scholar] [CrossRef]
- Gustafsson, B.M.; Gustafsson, P.A.; Proczkowska-Björklund, M. The Strengths and Difficulties Questionnaire (SDQ) for Preschool Children—A Swedish Validation. Nord. J. Psychiatry 2016, 70, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Heude, B.; Forhan, A.; Slama, R.; Douhaud, L.; Bedel, S.; Saurel-Cubizolles, M.-J.; Hankard, R.; Thiebaugeorges, O.; De Agostini, M.; Annesi-Maesano, I. Cohort Profile: The EDEN Mother-Child Cohort on the Prenatal and Early Postnatal Determinants of Child Health and Development. Int. J. Epidemiol. 2016, 45, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R. The Strengths and Difficulties Questionnaire: A Research Note. J. Child Psychol. Psychiatry 1997, 38, 581–586. [Google Scholar] [CrossRef]
- Goodman, A.; Lamping, D.L.; Ploubidis, G.B. When to Use Broader Internalising and Externalising Subscales Instead of the Hypothesised Five Subscales on the Strengths and Difficulties Questionnaire (SDQ): Data from British Parents, Teachers and Children. J. Abnorm. Child Psychol. 2010, 38, 1179–1191. [Google Scholar] [CrossRef]
- Gomajee, R.; El-Khoury, F.; Côté, S.; van der Waerden, J.; Pryor, L.; Melchior, M. Early Childcare Type Predicts Children’s Emotional and Behavioural Trajectories into Middle Childhood. Data from the EDEN Mother–Child Cohort Study. J. Epidemiol. Community Health 2018, 72, 1033–1043. [Google Scholar] [CrossRef]
- Teschendorff, A.E.; Marabita, F.; Lechner, M.; Bartlett, T.; Tegner, J.; Gomez-Cabrero, D.; Beck, S. A Beta-Mixture Quantile Normalization Method for Correcting Probe Design Bias in Illumina Infinium 450 k DNA Methylation Data. Bioinformatics 2013, 29, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Yuan, V.; Price, E.M.; Del Gobbo, G.; Mostafavi, S.; Cox, B.; Binder, A.M.; Michels, K.B.; Marsit, C.; Robinson, W.P. Accurate Ethnicity Prediction from Placental DNA Methylation Data. Epigenet. Chromatin 2019, 12, 51. [Google Scholar] [CrossRef]
- Radloff, L.S. The CES-D Scale: A Self-Report Depression Scale for Research in the General Population. Appl. Psychol. Meas. 1977, 1, 385–401. [Google Scholar] [CrossRef]
- Vilagut, G.; Forero, C.G.; Barbaglia, G.; Alonso, J. Screening for Depression in the General Population with the Center for Epidemiologic Studies Depression (CES-D): A Systematic Review with Meta-Analysis. PLoS ONE 2016, 11, e0155431. [Google Scholar] [CrossRef]
- Grigoriu, A.; Ferreira, J.C.; Choufani, S.; Baczyk, D.; Kingdom, J.; Weksberg, R. Cell Specific Patterns of Methylation in the Human Placenta. Epigenetics 2011, 6, 368–379. [Google Scholar] [CrossRef]
- Teschendorff, A.E.; Breeze, C.E.; Zheng, S.C.; Beck, S. A Comparison of Reference-Based Algorithms for Correcting Cell-Type Heterogeneity in Epigenome-Wide Association Studies. BMC Bioinform. 2017, 18, 105. [Google Scholar] [CrossRef]
- Pappa, I.; St Pourcain, B.; Benke, K.; Cavadino, A.; Hakulinen, C.; Nivard, M.G.; Nolte, I.M.; Tiesler, C.M.; Bakermans-Kranenburg, M.J.; Davies, G.E. A Genome-wide Approach to Children’s Aggressive Behavior: The EAGLE Consortium. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2016, 171, 562–572. [Google Scholar] [CrossRef]
- Gerritsen, L.; Milaneschi, Y.; Vinkers, C.H.; Van Hemert, A.M.; Van Velzen, L.; Schmaal, L.; Penninx, B.W. HPA Axis Genes, and Their Interaction with Childhood Maltreatment, Are Related to Cortisol Levels and Stress-Related Phenotypes. Neuropsychopharmacology 2017, 42, 2446–2455. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, J.; Gong, P.; Wang, J.; Fang, W.; Yan, H.; Zhu, L.; Zhou, X. Identifying New Susceptibility Genes on Dopaminergic and Serotonergic Pathways for the Framing Effect in Decision-Making. Soc. Cogn. Affect. Neurosci. 2017, 12, 1534–1544. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Pedersen, B.S.; Schwartz, D.A.; Yang, I.V.; Kechris, K.J. Comb-p: Software for Combining, Analyzing, Grouping and Correcting Spatially Correlated p-Values. Bioinformatics 2012, 28, 2986–2988. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Kahraman, A.; Law, C.W.; Lindsay, H.; Nowicka, M.; Weber, L.M.; Zhou, X. Statistical Methods for Detecting Differentially Methylated Loci and Regions. Front. Genet. 2014, 5, 324. [Google Scholar] [CrossRef] [PubMed]
- Kechris, K.J.; Biehs, B.; Kornberg, T.B. Generalizing Moving Averages for Tiling Arrays Using Combined p-Value Statistics. Stat. Appl. Genet. Mol. Biol. 2010, 9, 29. [Google Scholar] [CrossRef]
- Šidák, Z. Rectangular Confidence Regions for the Means of Multivariate Normal Distributions. J. Am. Stat. Assoc. 1967, 62, 626–633. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N = 441 | N or Mean | % or sd |
|---|---|---|
| Emotional and behavioural difficulties | ||
| Emotional symptoms | 1.8 | 1.7 |
| Girls | 2.0 | 1.8 |
| Boys | 1.6 | 1.7 |
| Peer relationship problems | 1.4 | 1.5 |
| Girls | 1.3 | 1.3 |
| Boys | 1.6 | 1.6 |
| Internalizing symptoms | 3.2 | 2.6 |
| Girls | 3.3 | 2.4 |
| Boys | 3.2 | 2.8 |
| Conduct problems | 3.2 | 2.0 |
| Girls | 2.9 | 1.9 |
| Boys | 3.4 | 2.0 |
| Inattention/hyperactivity | 3.4 | 2.2 |
| Girls | 3.1 | 2.1 |
| Boys | 3.6 | 2.3 |
| Externalizing symptoms | 6.5 | 3.6 |
| Girls | 6.1 | 3.4 |
| Boys | 6.9 | 3.7 |
| Total symptoms | 9.8 | 4.9 |
| Girls | 9.4 | 4.6 |
| Boys | 10.1 | 5.1 |
| Sociodemographic characteristics | ||
| Mother’s age at conception (years) | 29.6 | 4.8 |
| Parity | ||
| No other child | 206 | 0.47 |
| At least one other child | 235 | 0.53 |
| Mother’s ethnicity | ||
| Caucasian | 415 | 0.94 |
| Other | 26 | 0.06 |
| Mother’s educational attainment | ||
| Low | 139 | 0.32 |
| High | 302 | 0.68 |
| Pregnancy characteristics | ||
| Depressive symptoms during pregnancy | ||
| No (CES-D < 16) | 343 | 0.78 |
| Yes (CES-D >=16) | 98 | 0.22 |
| Adverse events during pregnancy | ||
| None | 270 | 0.61 |
| At least one | 171 | 0.39 |
| Maternal tobacco smoking exposure during pregnancy | ||
| Non-smoker or former smoker and no secondhand | 251 | 0.57 |
| Non-smoker or former smoker and secondhand | 74 | 0.17 |
| Occasional or smoker during all pregnancy | 116 | 0.26 |
| Infant characteristics | ||
| Sex | ||
| Boy | 228 | 0.52 |
| Girl | 213 | 0.48 |
| Estimated cellular composition | ||
| Endothelial | 0.10 | 0.03 |
| Hofbauer | 0.02 | 0.01 |
| nRBC | 0.02 | 0.01 |
| Stromal | 0.11 | 0.03 |
| Syncytiotrophoblast | 0.64 | 0.08 |
| Trophoblasts | 0.13 | 0.06 |
| Emotional Difficulties | Behavioural Difficulties | ||||||
|---|---|---|---|---|---|---|---|
| Approach/Subscale | Emotional Symptoms | Peer Relationship Problems | Internalizing symptoms | Inattention/ Hyperactivity | Conduct Problems | Externalizing Symptoms | Total |
| Hypothesis driven | |||||||
| HPA/Serotonin pathways (53 genes—736 CpGs) | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Literature review (92 genes—3 101 CpGs) | 1+ | 0 | 0 | 0 | 0 | 0 | 0 |
| EWAS | 0 | 2− | 0 | 0 | 0 | 0 | 0 |
| Girls | 17+/4− | 0 | 1+ | 0 | 0 | 0 | 0 |
| Boys | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| DMR | 11 | 5 | 12 | 5 | 4 | 4 | 12 |
| Girls | 48 | 12 | 17 | 7 | 3 | 0 | 11 |
| Boys | 4 | 4 | 9 | 5 | 4 | 0 | 9 |
| EWAS without CC adjustment | 0 | 1− | 0 | 0 | 0 | 0 | 0 |
| DMR without CC adjustment | 13 | 4 | 7 | 6 | 5 | 5 | 12 |
| All Children (N = 441) | Only Boys (N = 228) | Only Girls (N = 213) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DMR | Emotional Symptoms | Peer relationship Problems | Internalizing Symptoms | Hyperactivity/Inattention | Conduct Problems | Externalizing Symptoms | Total Symptoms | Emotional Symptoms | Peer relationship Problems | Internalizing Symptoms | Hyperactivity/Inattention | Conduct Problems | Externalizing Symptoms | Total Symptoms | Emotional Symptoms | Peer relationship Problems | Internalizing Symptoms | Hyperactivity/Inattention | Conduct Problems | Externalizing Symptoms | Total Symptoms |
| (A) DMRs only found in unstratified analyses (N = 9) | |||||||||||||||||||||
| ANKS1B * | 6 | ||||||||||||||||||||
| BIN2 * | 7 | ||||||||||||||||||||
| CALCB * | 6 | ||||||||||||||||||||
| EVX1 * | 6 | 6 | 6 | ||||||||||||||||||
| NPY * | 6 | ||||||||||||||||||||
| ProSAPiP1 † | 9 | ||||||||||||||||||||
| THSD7A * | 7 | 7 | 8 | ||||||||||||||||||
| VWCE * | 8 | 11 | |||||||||||||||||||
| ZBBX *† | 11 | 9 | |||||||||||||||||||
| (B) DMRs found both in unstratified and stratified analyses by child sex (N = 14) | |||||||||||||||||||||
| BSCL2 * | 9 | 9 | |||||||||||||||||||
| C3orf26 * | 6 | 6 | 5 | ||||||||||||||||||
| C5orf13 * | 13 | 12 | 13 | 13 | 7 | ||||||||||||||||
| C6orf47 *† | 18 | 18 | 22 | 22 | 27 | 18 | 28 | 25 | |||||||||||||
| CCDC62 | 9 | 8 | 8 | ||||||||||||||||||
| CLIP4 * | 8 | 8 | 9 | ||||||||||||||||||
| GGT1 * | 9 | 9 | 10 | 9 | |||||||||||||||||
| GPR75 † | 9 | 9 | |||||||||||||||||||
| FAM3B † | 8 | 8 | 7 | 7 | |||||||||||||||||
| MIR564 † | 12 | 13 | |||||||||||||||||||
| RUFY2 † | 8 | 7 | |||||||||||||||||||
| THBD † | 6 | 7 | |||||||||||||||||||
| UBXN11* | 5 | 12 | 11 | ||||||||||||||||||
| ZNF844 † | 6 | 6 | 6 | 7 | |||||||||||||||||
| (C) Sex-specific DMRs (N = 54) (** No significant DMR) | |||||||||||||||||||||
| ACP 5* | 5 | ||||||||||||||||||||
| ADAMTS17 * | 11 | 6 | |||||||||||||||||||
| B4GALNT2 * | 7 | 5 | |||||||||||||||||||
| C6orf52 * | 5 | ||||||||||||||||||||
| C10orf25 * | 10 | ||||||||||||||||||||
| C16orf67 * | 5 | ||||||||||||||||||||
| C17orf46 * | 7 | 6 | |||||||||||||||||||
| CCNA1 * | 5 | ||||||||||||||||||||
| CROT * | 12 | ||||||||||||||||||||
| CWH43 * | 8 | ||||||||||||||||||||
| EN1 | 5 | ||||||||||||||||||||
| FLJ42289 * | 6 | ||||||||||||||||||||
| FLOT1 * | 11 | 8 | |||||||||||||||||||
| GNAS * | 10 | ||||||||||||||||||||
| HECW1 * | 6 | ||||||||||||||||||||
| HFE * | 6 | ||||||||||||||||||||
| HLA-F * | 10 | ||||||||||||||||||||
| HORMAD 2* | 9 | ||||||||||||||||||||
| IGFBP3 * | 8 | 7 | |||||||||||||||||||
| KIAA0101 | 8 | ||||||||||||||||||||
| LOC148824 * | 5 | ||||||||||||||||||||
| LOC641518 * | 5 | ||||||||||||||||||||
| MECOM * | 5 | ||||||||||||||||||||
| MOCS1 | 8 | ||||||||||||||||||||
| NA (cg04373548) † | 8 | ||||||||||||||||||||
| NA (cg05931423) * | 5 | ||||||||||||||||||||
| NA (cg18081456) * | 5 | ||||||||||||||||||||
| NA (cg18356974) | 5 | ||||||||||||||||||||
| NA (cg21334513) † | 12 | ||||||||||||||||||||
| NA (cg27405554) | 5 | ||||||||||||||||||||
| NR2E1 * | 9 | ||||||||||||||||||||
| PAX6 * | 10 | ||||||||||||||||||||
| PRRT1 * | 14 | ||||||||||||||||||||
| RIN2 | 5 | ||||||||||||||||||||
| RPS6KA2 † | 9 | ||||||||||||||||||||
| SEC14L4 * | 7 | ||||||||||||||||||||
| SERPING1 * | 6 | ||||||||||||||||||||
| SOX11 * | 5 | 5 | 5 | ||||||||||||||||||
| SP9 † | 6 | ||||||||||||||||||||
| STAM † | 6 | 6 | |||||||||||||||||||
| TBX15 * | 6 | ||||||||||||||||||||
| TEKT1 | 6 | ||||||||||||||||||||
| TM6SF1 * | 5 | ||||||||||||||||||||
| TMEM176B * | 6 | ||||||||||||||||||||
| TRIM26 * | 10 | ||||||||||||||||||||
| TRIM72 * | 14 | ||||||||||||||||||||
| TSPYL3 * | 13 | ||||||||||||||||||||
| TSTD1 * | 10 | ||||||||||||||||||||
| WSCD2 * | 5 | 5 | 5 | ||||||||||||||||||
| ZFP42 * | 9 | ||||||||||||||||||||
| ZMYND10 † | 11 | ||||||||||||||||||||
| ZNF655 | 10 | ||||||||||||||||||||
| ZNF780B * | 5 | ||||||||||||||||||||
| ZSCAN16 * | 8 | ||||||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakamura, A.; Broséus, L.; Tost, J.; Vaiman, D.; Martins, S.; Keyes, K.; Bonello, K.; Fekom, M.; Strandberg-Larsen, K.; Sutter-Dallay, A.-L.; et al. Epigenome-Wide Associations of Placental DNA Methylation and Behavioral and Emotional Difficulties in Children at 3 Years of Age. Int. J. Mol. Sci. 2023, 24, 11772. https://doi.org/10.3390/ijms241411772
Nakamura A, Broséus L, Tost J, Vaiman D, Martins S, Keyes K, Bonello K, Fekom M, Strandberg-Larsen K, Sutter-Dallay A-L, et al. Epigenome-Wide Associations of Placental DNA Methylation and Behavioral and Emotional Difficulties in Children at 3 Years of Age. International Journal of Molecular Sciences. 2023; 24(14):11772. https://doi.org/10.3390/ijms241411772
Chicago/Turabian StyleNakamura, Aurélie, Lucile Broséus, Jörg Tost, Daniel Vaiman, Silvia Martins, Katherine Keyes, Kim Bonello, Mathilde Fekom, Katrine Strandberg-Larsen, Anne-Laure Sutter-Dallay, and et al. 2023. "Epigenome-Wide Associations of Placental DNA Methylation and Behavioral and Emotional Difficulties in Children at 3 Years of Age" International Journal of Molecular Sciences 24, no. 14: 11772. https://doi.org/10.3390/ijms241411772
APA StyleNakamura, A., Broséus, L., Tost, J., Vaiman, D., Martins, S., Keyes, K., Bonello, K., Fekom, M., Strandberg-Larsen, K., Sutter-Dallay, A.-L., Heude, B., Melchior, M., & Lepeule, J. (2023). Epigenome-Wide Associations of Placental DNA Methylation and Behavioral and Emotional Difficulties in Children at 3 Years of Age. International Journal of Molecular Sciences, 24(14), 11772. https://doi.org/10.3390/ijms241411772