
Comprehensive Review: Unveiling the Pro-Oncogenic Roles of IL-1ß and PD-1/PD-L1 in NSCLC Development and Targeting Their Pathways for Clinical Management

 , , ,
, , ,  ,
,
Abstract
1. Introduction
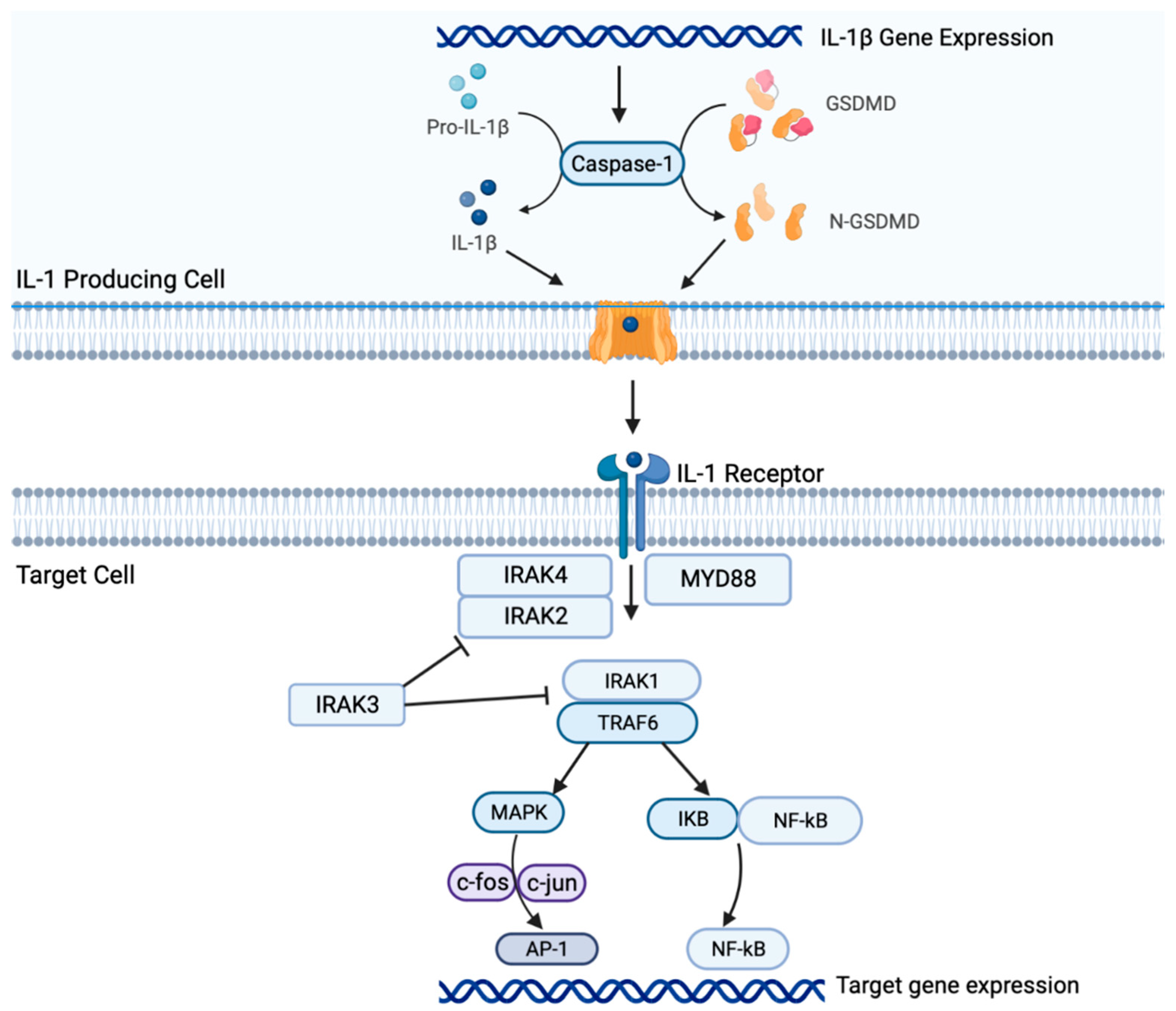
2. Il-1β Signaling in NSCLC
3. Interplay of PD-1/PD-L1 with TME and Their Role in Treatment Resistance
4. Complex Networking of IL-1ß and PD-1/PD-L1 Pathway and Implications for Targeted Therapies
5. IL-1β/PD-1/PD-L1 Pathways in Recent and Ongoing NSCLC Clinical Studies
6. The Unmet Need for Predictive Biomarkers of Response and Resistance in NSCLC
7. Future Perspective and Direction Targeting IL-1β and PD-1/PD-L1 Pathways in NSCLC
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| NSCLC | non-small cell lung cancer |
| ICI | immune-checkpoint inhibitor |
| PD-L1 or B7-H1 | programmed death ligand 1 |
| TME | tumor microenvironment |
| IL-1β | interleukin-1 beta |
| CANTOS | the Canakinumab Anti-inflammatory Thrombosis Outcome Study trial |
| PD-1 | programmed cell death protein 1 |
| TLR | toll-like receptor |
| TNF | tumor necrosis factor |
| ICE | IL-1β-converting enzyme |
| DC | dendritic cell |
| APC | antigen-presenting cell |
| IL-1R and IL-1R2 | IL-1 receptors |
| OS | overall survival |
| PFS | progression-free survival |
| MyD88 | myeloid differentiation primary response-88 |
| IRAK | IL-1R associated kinase |
| TRAF6 | tumor necrosis factor receptor-associated factor 6 |
| MAPK | mitogen-activated protein kinase |
| GSDMD | gasdermin D |
| IkB | inhibitor of nuclear factor kappa B |
| NF-kB | nuclear factor kappa B |
| AKT | protein kinase B |
| AP-1 | activator protein 1 |
| NK | natural killer |
| PIK3 | phosphoinositide 3-kinase |
| MDSC | myeloid-derived suppressor cell |
| Treg | regulatory T cell |
| TAM | tumor-associated macrophage |
| MCP-1 | monocyte chemoattractant protein |
| VEGF | vascular endothelial growth factor |
| bFGF | basic fibroblast growth factor |
| TGF-𝛼β | transforming growth factor |
| PDGF | platelet-derived endothelial growth factor |
| IL-8 or CXCL8 | interleukin-8 |
| FGF | fibroblast growth factor |
| MMP | matrix metalloproteinase |
| ECM | extracellular matrix |
| PTEN | phosphatase and tensin homolog |
| EMT | epithelial-mesenchymal transition |
| COX2 | cyclooxygenase 2 |
| iNOS | inducible nitric oxide synthase |
| V600E | BRAF |
| RNAis | RNA inhibitors |
| RCC) model | renal cell carcinoma |
| BRAFi | BRAF inhibitors |
| CAF | cancer-associated fibroblast |
| TIL | tumor infiltrating lymphocyte |
| EHD1 | EH domain-containing protein 1 |
| TKI | tyrosine kinase inhibitor |
| M-MDSC | monocytic myeloid-derived suppressor cell |
| LUAD | lung adenocarcinoma |
| CRP | C-reactive protein |
| OS | overall survival |
| PFS | progression-free survival |
| IFN | interferon |
| MSI | microsatellite instability |
| TMB | tumor mutational burden |
| RCC | renal cell carcinoma |
| MPR | major pathologic response |
| DLT | dose-limiting toxicity |
| DFS | disease-free survival |
| m | months |
| AE | adverse event |
| TEAE | treatment-emergent adverse event |
| SAE | serious adverse event |
| LUAD | lung adenocarcinoma |
| PFSR | progression free survival rate |
| ORR | overall response rate |
| OR | odds ratio |
| ctDNA | circulating tumor DNA |
| irAE | immune-related adverse event |
References
- de Alencar, V.T.L.; Figueiredo, A.B.; Corassa, M.; Gollob, K.J.; Cordeiro de Lima, V.C. Lung cancer in never smokers: Tumor immunology and challenges for immunotherapy. Front. Immunol. 2022, 13, 984349. [Google Scholar] [CrossRef] [PubMed]
- Gridelli, C.; Rossi, A.; Carbone, D.P.; Guarize, J.; Karachaliou, N.; Mok, T.; Petrella, F.; Spaggiari, L.; Rosell, R. Non-small-cell lung cancer. Nat. Rev. Dis. Primers 2015, 1, 15009. [Google Scholar] [CrossRef]
- Zhang, J.; Veeramachaneni, N. Targeting interleukin-1β and inflammation in lung cancer. Biomark. Res. 2022, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Tsuboi, M.; Kim, E.S.; Mok, T.S.; Garrido, P. Overcoming immunosuppression and pro-tumor inflammation in lung cancer with combined IL-1β and PD-1 inhibition. Future Oncol. 2022, 18, 3085–3100. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; MacFadyen, J.G.; Thuren, T.; Everett, B.M.; Libby, P.; Glynn, R.J. Effect of interleukin-1β inhibition with canakinumab on incident lung cancer in patients with atherosclerosis: Exploratory results from a randomised, double-blind, placebo-controlled trial. Lancet 2017, 390, 1833–1842. [Google Scholar] [CrossRef]
- Yi, M.; Niu, M.; Xu, L.; Luo, S.; Wu, K. Regulation of PD-L1 expression in the tumor microenvironment. J. Hematol. Oncol. 2021, 14, 10. [Google Scholar] [CrossRef]
- Budimir, N.; Thomas, G.D.; Dolina, J.S.; Salek-Ardakani, S. Reversing T-cell Exhaustion in Cancer: Lessons Learned from PD-1/PD-L1 Immune Checkpoint Blockade. Cancer Immunol. Res. 2022, 10, 146–153. [Google Scholar] [CrossRef]
- Wang, S.; Yuan, P.; Mao, B.; Li, N.; Ying, J.; Tao, X.; Tang, W.; Zhang, L.; Geng, X.; Zhang, F.; et al. Genomic features and tumor immune microenvironment alteration in NSCLC treated with neoadjuvant PD-1 blockade. NPJ Precis. Oncol. 2022, 6, 2. [Google Scholar] [CrossRef]
- Huang, J.; Lan, X.; Wang, T.; Lu, H.; Cao, M.; Yang, S.; Cui, Y.; Jia, D.; Cai, L.; Xing, Y. Targeting the IL-1β/EHD1/TUBB3 axis overcomes resistance to EGFR-TKI in NSCLC. Oncogene 2020, 39, 1739–1755. [Google Scholar] [CrossRef]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef]
- Yan, X.; Han, L.; Zhao, R.; Fatima, S.; Zhao, L.; Gao, F. Prognosis value of IL-6, IL-8, and IL-1β in serum of patients with lung cancer: A fresh look at interleukins as a biomarker. Heliyon 2022, 8, e09953. [Google Scholar] [CrossRef] [PubMed]
- Rébé, C.; Ghiringhelli, F. Interleukin-1β and Cancer. Cancers 2020, 12, 1791. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Li, T.; Li, S.; Yang, H.; Wu, C.; Zheng, C.; You, F.; Liu, Y. The tumor biochemical and biophysical microenvironments synergistically contribute to cancer cell malignancy. Cell. Mol. Immunol. 2020, 17, 1186–1187. [Google Scholar] [CrossRef] [PubMed]
- Litmanovich, A.; Khazim, K.; Cohen, I. The Role of Interleukin-1 in the Pathogenesis of Cancer and its Potential as a Therapeutic Target in Clinical Practice. Oncol. Ther. 2018, 6, 109–127. [Google Scholar] [CrossRef]
- Elkabets, M.; Ribeiro, V.S.; Dinarello, C.A.; Ostrand-Rosenberg, S.; Di Santo, J.P.; Apte, R.N.; Vosshenrich, C. IL-1β regulates a novel myeloid-derived suppressor cell subset that impairs NK cell development and function. Eur. J. Immunol. 2010, 40, 3347–3357. [Google Scholar] [CrossRef]
- Li, S.; Wang, W.; Zhang, N.; Ma, T.; Zhao, C. IL-1β mediates MCP-1 induction by Wnt5a in gastric cancer cells. BMC Cancer 2014, 14, 480. [Google Scholar] [CrossRef]
- Kolb, R.; Phan, L.; Borcherding, N.; Liu, Y.; Yuan, F.; Janowski, A.M.; Xie, Q.; Markan, K.; Li, W.; Potthoff, M.J.; et al. Obesity-associated NLRC4 inflammasome activation drives breast cancer progression. Nat. Commun. 2016, 7, 13007. [Google Scholar] [CrossRef]
- Carmi, Y.; Dotan, S.; Rider, P.; Kaplanov, I.; White, M.R.; Baron, R.; Abutbul, S.; Huszar, M.; Dinarello, C.; Apte, R.N.; et al. The role of IL-1β in the early tumor cell-induced angiogenic response. J. Immunol. 2013, 190, 3500–3509. [Google Scholar] [CrossRef]
- Fahey, E.; Doyle, S.L. IL-1 family cytokine regulation of vascular permeability and angiogenesis. Front. Immunol. 2019, 10, 1426. [Google Scholar] [CrossRef]
- Ping, P.H.; Bo, T.F.; Li, L.; Hui, Y.N.; Zhu, H. IL-1β/NF-kb signaling promotes colorectal cancer cell growth through miR-181a/PTEN axis. Arch. Biochem. Biophys. 2016, 604, 20–26. [Google Scholar] [CrossRef]
- Wu, C.; Xu, B.; Zhou, Y.; Ji, M.; Zhang, D.; Jiang, J.; Wu, C. Correlation between serum IL-1β and miR-144-3p as well as their prognostic values in LUAD and LUSC patients. Oncotarget. 2016, 7, 85876–85887. [Google Scholar] [CrossRef] [PubMed]
- Sanmamed, M.F.; Perez-Gracia, J.L.; Schalper, K.A.; Fusco, J.P.; Gonzalez, A.; Rodrigeuz-Ruiz, M.E.; Onate, C.; Perez, G.; Alfaro, C.; Martin-Algarra, S.; et al. Changes in serum interleukin-8 (IL-8) levels reflect and predict response to anti-PD-1 treatment in melanoma and non-small-cell lung cancer patients. Ann. Oncol. 2017, 28, 1988–1995. [Google Scholar] [CrossRef] [PubMed]
- Coffelt, S.B.; Kersten, K.; Doornebal, C.W.; Weiden, J.; Vrijland, K.; Hau, C.; Verstegen, N.J.M.; Ciampricotti, M.; Hawinkels, L.J.A.C.; Jonkers, J.; et al. IL-17-producing γδ T cells and neutrophils conspire to promote breast cancer metastasis. Nature 2015, 522, 345–348. [Google Scholar] [CrossRef]
- Jin, C.; Lagoudas, G.K.; Zhao, C.; Bullman, S.; Bhutkar, A.; Hu, B.; Ameh, S.; Sandel, D.; Liang, X.S.; Mazzilli, S.; et al. Commensal Microbiota Promote Lung Cancer Development via γδ T Cells. Cell 2019, 176, 998–1013.e16. [Google Scholar] [CrossRef]
- Syn, N.L.; Teng, M.W.L.; Mok, T.S.K.; Soo, R.A. De-novo and acquired resistance to immune checkpoint targeting. Lancet Oncol. 2017, 18, e731–e741. [Google Scholar] [CrossRef] [PubMed]
- Garon, E.B.; Chih-Hsin Yang, J.; Dubinett, S.M. The Role of Interleukin 1β in the Pathogenesis of Lung Cancer. JTO Clin. Res. Rep. 2020, 1, 100001. [Google Scholar] [CrossRef] [PubMed]
- Gelfo, V.; Romaniello, D.; Mazzeschi, M.; Sgarzi, M.; Grilli, G.; Morselli, A.; Manzan, B.; Rihawi, K.; Lauriola, M. Roles of IL-1 in Cancer: From Tumor Progression to Resistance to Targeted Therapies. Int. J. Mol. Sci. 2020, 21, 6009. [Google Scholar] [CrossRef]
- Yan, N.; Guo, S.; Zhang, H.; Zhang, Z.; Shen, S.; Li, X. BRAF-Mutated Non-Small Cell Lung Cancer: Current Treatment Status and Future Perspective. Front. Oncol. 2022, 12, 863043. [Google Scholar] [CrossRef]
- Hajek, E.; Krebs, F.; Bent, R.; Haas, K.; Bast, A.; Steinmetz, I.; Tuettenberg, A.; Grabbe, S.; Bros, M. BRAF inhibitors stimulate inflammasome activation and interleukin 1 beta production in dendritic cells. Oncotarget 2018, 9, 28294–28308. [Google Scholar] [CrossRef]
- Davies, A.M.; Lara PNJr Mack, P.C.; Gumerlock, P.H.; Bold, R.J.; Gandara, D.R. Bortezomib-based combinations in the treatment of non-small-cell lung cancer. Clin. Lung Cancer 2005, 7 (Suppl. 2), S59–S63. [Google Scholar] [CrossRef]
- McLoed, A.G.; Sherrill, T.P.; Cheng, D.S.; Han, W.; Saxon, J.; Gleaves, L.A.; Wu, P.; Polosukhin, V.; Karin, M.; Yull, F.E.; et al. Neutrophil-Derived IL-1β Impairs the Efficacy of NF-κB Inhibitors against Lung Cancer. Cell Rep. 2016, 16, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Clowers, M.J.; Velasco, W.V.; Peng, S.; Peng, Q.; Shi, Y.; Ramos-Castaneda, M.; Zarghooni, M.; Yang, S.; Babcock, R.L.; et al. Targeting IL-1β as an immunopreventive and therapeutic modality for K-ras-mutant lung cancer. JCI Insight 2022, 7, e157788. [Google Scholar] [CrossRef] [PubMed]
- Russano, M.; La Cava, G.; Cortellini, A.; Citarella, F.; Galletti, A.; Di Fazio, G.R.; Santo, V.; Brunetti, L.; Vendittelli, A.; Fioroni, I.; et al. Immunotherapy for Metastatic Non-Small Cell Lung Cancer: Therapeutic Advances and Biomarkers. Curr. Oncol. 2023, 30, 2366–2387. [Google Scholar] [CrossRef] [PubMed]
- Addeo, A.; Passaro, A.; Malapelle, U.; Banna, G.L.; Subbiah, V.; Friedlaender, A. Immunotherapy in non-small cell lung cancer harbouring driver mutations. Cancer Treat. Rev. 2021, 96, 102179. [Google Scholar] [CrossRef]
- Negrao, M.V.; Skoulidis, F.; Montesion, M.; Schulze, K.; Bara, I.; Shen, V.; Xu, H.; Hu, S.; Sui, D.; Elamin, Y.Y.; et al. Oncogene-specific differences in tumor mutational burden, PD-L1 expression, and outcomes from immunotherapy in non-small cell lung cancer. J. Immunother. Cancer 2021, 9, e002891. [Google Scholar] [CrossRef]
- Simon, S.; Labarriere, N. PD-1 expression on tumor-specific T cells: Friend or foe for immunotherapy? Oncoimmunology 2017, 7, e1364828. [Google Scholar] [CrossRef]
- Kumagai, S.; Togashi, Y.; Kamada, T.; Sugiyama, E.; Nishinakamura, H.; Takeuchi, Y.; Vitaly, K.; Itahashi, K.; Maeda, Y.; Shigeyuki, M.; et al. The PD-1 expression balance between effector and regulatory T cells predicts the clinical efficacy of PD-1 blockade therapies. Nat. Immunol. 2020, 21, 1346–1358. [Google Scholar] [CrossRef]
- Sari, M.I.; Ilyas, S. The Expression Levels and Concentrations of PD-1 and PD-L1 Proteins in Septic Patients: A Systematic Review. Diagnostics 2022, 12, 2004. [Google Scholar] [CrossRef]
- Xu, X.; Hou, B.; Fulzele, A.; Masubuchi, T.; Zhao, Y.; Wu, Z.; Hu, Y.; Jiang, Y.; Ma, Y.; Wang, H.; et al. PD-1 and BTLA regulate T cell signaling differentially and only partially through SHP1 and SHP2. J. Cell Biol. 2020, 219, e201905085. [Google Scholar] [CrossRef]
- Lei, Q.; Wang, D.; Sun, K.; Wang, L.; Zhang, Y. Resistance Mechanisms of Anti-PD1/PDL1 Therapy in Solid Tumors. Front. Cell Dev. Biol. 2020, 8, 672. [Google Scholar] [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar] [PubMed]
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csőszi, T.; Fülöp, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Pembrolizumab versus Chemotherapy for PD-L1-Positive Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2016, 375, 1823–1833. [Google Scholar] [CrossRef]
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csőszi, T.; Fülöp, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Five-Year Outcomes With Pembrolizumab Versus Chemotherapy for Metastatic Non-Small-Cell Lung Cancer With PD-L1 Tumor Proportion Score ≥ 50. J. Clin. Oncol. 2021, 39, 2339–2349. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S.; Garon, E.B.; Kim, D.W.; Cho, B.C.; Gervais, R.; Perez-Gracia, J.L.; Han, J.Y.; Majem, M.; Forster, M.D.; Monnet, I.; et al. Five Year Survival Update From KEYNOTE-010: Pembrolizumab Versus Docetaxel for Previously Treated, Programmed Death-Ligand 1-Positive Advanced NSCLC. J. Thorac. Oncol. 2021, 16, 1718–1732. [Google Scholar] [CrossRef]
- Horn, L.; Spigel, D.R.; Vokes, E.E.; Holgado, E.; Ready, N.; Steins, M.; Poddubskaya, E.; Borghaei, H.; Felip, E.; Paz-Ares, L.; et al. Nivolumab Versus Docetaxel in Previously Treated Patients With Advanced Non-Small-Cell Lung Cancer: Two-Year Outcomes From Two Randomized, Open-Label, Phase III Trials (CheckMate 017 and CheckMate 057). J. Clin. Oncol. 2017, 35, 3924–3933. [Google Scholar] [CrossRef]
- Sezer, A.; Kilickap, S.; Gümüş, M.; Bondarenko, I.; Özgüroğlu, M.; Gogishvili, M.; Turk, H.M.; Cicin, I.; Bentsion, D.; Gladkov, O.; et al. Cemiplimab monotherapy for first-line treatment of advanced non-small-cell lung cancer with PD-L1 of at least 50%: A multicentre, open-label, global, phase 3, randomized, controlled trial. Lancet 2021, 397, 592–604. [Google Scholar] [CrossRef]
- Antonia, S.J.; Villegas, A.; Daniel, D.; Vicente, D.; Murakami, S.; Hui, R.; Yokoi, T.; Chiappori, A.; Lee, K.H.; Wit, M.D.; et al. Durvalumab after Chemoradiotherapy in Stage III Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2017, 377, 1919–1929. [Google Scholar] [CrossRef]
- Rizvi, N.A.; Cho, B.C.; Reinmuth, N.; Lee, K.H.; Luft, A.; Ahn, M.J.; Van den Heuvel, M.M.; Cobo, M.; Vicente, D.; Smolin, A.; et al. Durvalumab With or Without Tremelimumab vs Standard Chemotherapy in First-line Treatment of Metastatic Non-Small Cell Lung Cancer: The MYSTIC Phase 3 Randomized Clinical Trial. JAMA Oncol. 2020, 6, 661–674, Correction in JAMA Oncol. 2020, 6, 1815. [Google Scholar] [CrossRef]
- Jotte, R.; Cappuzzo, F.; Vynnychenko, I.; Stroyakovskiy, D.; Rodríguez-Abreu, D.; Hussein, M.; Soo, R.; Conter, H.; Kozuki, T.; Huang, K.C.; et al. Atezolizumab in Combination With Carboplatin and Nab-Paclitaxel in Advanced Squamous NSCLC (IMpower131): Results From a Randomized Phase III Trial. J. Thorac. Oncol. 2020, 15, 1351–1360. [Google Scholar] [CrossRef]
- West, H.; McCleod, M.; Hussein, M.; Morabito, A.; Rittmeyer, A.; Conter, H.; Kopp, H.G.; Daniel, D.; McCune, S.; Mekhail, T.; et al. Atezolizumab in combination with carboplatin plus nab-paclitaxel chemotherapy compared with chemotherapy alone as first-line treatment for metastatic non-squamous non-small-cell lung cancer (IMpower130): A multicentre, randomised, open-label, phase 3 trial. Lancet Oncol. 2019, 20, 924–937. [Google Scholar] [CrossRef]
- Socinski, M.A.; Nishio, M.; Jotte, R.M.; Cappuzzo, F.; Orlandi, F.; Stroyakovskiy, D.; Nogami, N.; Rodríguez-Abreu, D.; Moro-Sibilot, D.; Thomas, C.A.; et al. IMpower150 Final Overall Survival Analyses for Atezolizumab Plus Bevacizumab and Chemotherapy in First-Line Metastatic Nonsquamous NSCLC. J. Thorac. Oncol. 2021, 16, 1909–1924. [Google Scholar] [CrossRef] [PubMed]
- Rittmeyer, A.; Barlesi, F.; Waterkamp, D.; Park, K.; Ciardiello, F.; Von Pawel, J.; Gadgeel, S.M.; Hida, T.; Kowalski, D.M.; Cobo Dols, M.; et al. Atezolizumab versus docetaxel in patients with previously treated non-small-cell lung cancer (OAK): A phase 3, open-label, multicentre randomised controlled trial. Lancet 2017, 389, 255–265, Correction in Lancet 2017, 389, e5. [Google Scholar] [CrossRef] [PubMed]
- Pathak, R.; Pharaon, R.R.; Mohanty, A.; Villaflor, V.M.; Salgia, R.; Massarelli, E. Acquired Resistance to PD-1/PD-L1 Blockade in Lung Cancer: Mechanisms and Patterns of Failure. Cancers 2020, 12, 3851. [Google Scholar] [CrossRef] [PubMed]
- Passaro, A.; Brahmer, J.; Antonia, S.; Mok, T.; Peters, S. Managing Resistance to Immune Checkpoint Inhibitors in Lung Cancer: Treatment and Novel Strategies. J. Clin. Oncol. 2022, 40, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Vesely, M.D.; Zhang, T.; Chen, L. Resistance Mechanisms to Anti-PD Cancer Immunotherapy. Annu. Rev. Immunol. 2022, 40, 45–74. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Wolchok, J.D.; Chen, L. PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: Mechanisms, response biomarkers, and combinations. Sci. Transl. Med. 2016, 8, 328rv4. [Google Scholar] [CrossRef]
- Cheng, C.; Zhuge, L.; Xiao, X.; Luan, S.; Yuan, Y. Overcoming resistance to PD-1/PD-L1 inhibitors in esophageal cancer. Front. Oncol. 2022, 12, 955163. [Google Scholar] [CrossRef]
- Walsh, R.J.; Soo, R.A. Resistance to immune checkpoint inhibitors in non-small cell lung cancer: Biomarkers and therapeutic strategies. Ther. Adv. Med. Oncol. 2020, 12, 1758835920937902. [Google Scholar] [CrossRef]
- Bagchi, S.; Yuan, R.; Engleman, E.G. Immune Checkpoint Inhibitors for the Treatment of Cancer: Clinical Impact and Mechanisms of Response and Resistance. Annu. Rev. Pathol. 2021, 16, 223–249. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, L. Classification of Advanced Human Cancers Based on Tumor Immunity in the MicroEnvironment (TIME) for Cancer Immunotherapy. JAMA Oncol. 2016, 2, 1403–1404, Correction in JAMA Oncol. 2016, 2, 1511. [Google Scholar] [CrossRef]
- Zhuang, Y.; Liu, C.; Liu, J.; Li, G. Resistance Mechanism of PD-1/PD-L1 Blockade in the Cancer-Immunity Cycle. Onco Targets Ther. 2020, 13, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Chen, J.Q.; Liu, C.; Malu, S.; Creasy, C.; Tetzlaff, M.T.; Xu, C.; McKenzie, J.A.; Zhang, C.; Liang, X.; et al. Loss of PTEN Promotes Resistance to T Cell-Mediated Immunotherapy. Cancer Discov. 2016, 6, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Ngiow, S.F.; Young, A.; Jacquelot, N.; Yamazaki, T.; Enot, D.; Zitvogel, L.; Smyth, M.J. A Threshold Level of Intratumor CD8+ T-cell PD1 Expression Dictates Therapeutic Response to Anti-PD1. Cancer Res. 2015, 75, 3800–3811. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yuan, X.; Wang, M.; He, Z.; Li, H.; Wang, J.; Li, Q. Gut microbiota influence immunotherapy responses: Mechanisms and therapeutic strategies. J. Hematol. Oncol. 2022, 15, 47. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef]
- Goodman, A.M.; Kato, S.; Bazhenova, L.; Patel, S.P.; Frampton, G.M.; Miller, V.; Stephens, P.J.; Daniels, G.A.; Kurzrock, R. Tumor Mutational Burden as an Independent Predictor of Response to Immunotherapy in Diverse Cancers. Mol. Cancer Ther. 2017, 16, 2598–2608. [Google Scholar] [CrossRef]
- Dong, Z.Y.; Zhang, J.T.; Liu, S.Y.; Su, J.; Zhang, C.; Xie, Z.; Zhou, Q.; Tu, H.Y.; Xu, C.R.; Yan, L.X.; et al. EGFR mutation correlates with uninflamed phenotype and weak immunogenicity, causing impaired response to PD-1 blockade in non-small cell lung cancer. Oncoimmunology 2017, 6, e1356145. [Google Scholar] [CrossRef]
- Gainor, J.F.; Shaw, A.T.; Sequist, L.V.; Fu, X.; Azzoli, C.G.; Piotrowska, Z.; Huynh, T.G.; Zhao, L.; Fulton, L.; Schultz, K.R.; et al. EGFR Mutations and ALK Rearrangements Are Associated with Low Response Rates to PD-1 Pathway Blockade in Non-Small Cell Lung Cancer: A Retrospective Analysis. Clin Cancer Res. 2016, 22, 4585–4593. [Google Scholar] [CrossRef]
- Sakai, T.; Udagawa, H.; Matsumoto, S.; Yoh, K.; Nosaki, K.; Ikeda, T.; Zenke, Y.; Kirita, K.; Niho, S.; Akimoto, T.; et al. Morphological, immune and genetic features in biopsy sample associated with the efficacy of pembrolizumab in patients with non-squamous non-small cell lung cancer. J. Cancer Res. Clin. Oncol. 2021, 147, 1227–1237. [Google Scholar] [CrossRef]
- Giustini, N.; Bazhenova, L. Recognizing Prognostic and Predictive Biomarkers in the Treatment of Non-Small Cell Lung Cancer (NSCLC) with Immune Checkpoint Inhibitors (ICIs). Lung Cancer 2021, 12, 21–34. [Google Scholar] [CrossRef]
- Mortezaee, K.; Majidpoor, J. Checkpoint inhibitor/interleukin-based combination therapy of cancer. Cancer Med. 2022, 11, 2934–2943. [Google Scholar] [CrossRef] [PubMed]
- Zong, Z.; Zou, J.; Mao, R.; Ma, C.; Li, N.; Wang, J.; Wang, X.; Zhou, H.; Zhang, L.; Shi, Y. M1 Macrophages Induce PD-L1 Expression in Hepatocellular Carcinoma Cells Through IL-1β Signaling. Front. Immunol. 2019, 10, 1643. [Google Scholar] [CrossRef] [PubMed]
- Perrichet, A.; Ghiringhelli, F.; Rébé, C. Understanding Inflammasomes and PD-1/PD-L1 Crosstalk to Improve Cancer Treatment Efficiency. Cancers 2020, 12, 3550. [Google Scholar] [CrossRef] [PubMed]
- Numata, Y.; Akutsu, N.; Ishigami, K.; Koide, H.; Wagatsuma, K.; Motoya, M.; Sasaki, S.; Nakase, H. Synergistic effect of IFN-γ and IL-1β on PD-L1 expression in hepatocellular carcinoma. Biochem. Biophys. Rep. 2022, 30, 101270. [Google Scholar] [CrossRef]
- Lu, X.; Li, Y.; Yang, W.; Tao, M.; Dai, Y.; Xu, J.; Xu, Q. Inhibition of NF-κB is required for oleanolic acid to downregulate PD-L1 by promoting DNA demethylation in gastric cancer cells. J. Biochem. Mol. Toxicol. 2021, 35, e22621. [Google Scholar] [CrossRef]
- Khalili, J.S.; Liu, S.; Rodríguez-Cruz, T.G.; Whittington, M.; Wardell, S.; Liu, C.; Zhang, M.; Cooper, Z.A.; Frederick, D.; Li, Y.; et al. Oncogenic BRAF(V600E) promotes stromal cell-mediated immunosuppression via induction of interleukin-1 in melanoma. Clin. Cancer Res. 2012, 18, 5329–5340. [Google Scholar] [CrossRef]
- Bar, N.; Costa, F.; Das, R.; Duffy, A.; Samur, M.; McCachren, S.; Gettinger, S.N.; Neparidze, N.; Parker, T.L.; Bailur, J.K.; et al. Differential effects of PD-L1 versus PD-1 blockade on myeloid inflammation in human cancer. JCI Insight 2020, 5, e129353. [Google Scholar] [CrossRef]
- Takahashi, R.; Macchini, M.; Sunagawa, M.; Jiang, Z.; Tanaka, T.; Valenti, G.; Renz, B.W.; White, R.A.; Hayakawa, Y.; Westphalen, C.B.; et al. Interleukin-1β-induced pancreatitis promotes pancreatic ductal adenocarcinoma via B lymphocyte-mediated immune suppression. Gut 2021, 70, 330–341. [Google Scholar] [CrossRef]
- Aggen, D.H.; Ager, C.R.; Obradovic, A.Z.; Chowdhury, N.; Ghasemzadeh, A.; Mao, W.; Chaimowitz, M.G.; Lopez-Bujanda, Z.A.; Spina, C.S.; Hawley, J.E.; et al. Blocking IL1 Beta Promotes Tumor Regression and Remodeling of the Myeloid Compartment in a Renal Cell Carcinoma Model: Multidimensional Analyses. Clin. Cancer Res. 2021, 27, 608–621. [Google Scholar] [CrossRef]
- Chen, J.; Del Valle, L.; Lin, H.Y.; Plaisance-Bonstaff, K.; Forrest, J.C.; Post, S.R.; Qin, Z. Expression of PD-1 and PD-Ls in Kaposi’s sarcoma and regulation by oncogenic herpesvirus lytic reactivation. Virology 2019, 536, 16–19. [Google Scholar] [CrossRef]
- Li, R.; Ong, S.L.; Tran, L.M.; Jing, Z.; Liu, B.; Park, S.J.; Huang, Z.L.; Walser, T.C.; Heinrich, E.L.; Lee, G.; et al. Chronic IL-1β-induced inflammation regulates epithelial-to-mesenchymal transition memory phenotypes via epigenetic modifications in non-small cell lung cancer. Sci Rep. 2020, 10, 377, Correction in Sci. Rep. 2020, 10, 4386. [Google Scholar] [CrossRef] [PubMed]
- Kaplanov, I.; Carmi, Y.; Kornetsky, R.; Shemesh, A.; Shurin, G.V.; Shurin, M.R.; Dinarello, C.A.; Voronov, E.; Apte, R.N. Blocking IL-1β reverses the immunosuppression in mouse breast cancer and synergizes with anti-PD-1 for tumor abrogation. Proc. Natl. Acad. Sci. USA 2019, 116, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, P.; Millholland, J.; O’Brien, N.; Wong, C.; Diwanji, R.; Wang, M.; Choi, E.; Linnartz, R.; Rose, K.; Rodrik-Outmezguine, V.; et al. Abstract C103: Targeting IL-1β pathway for cancer immunotherapy. Mol. Cancer Ther. 2019, 18 (Suppl. 12). [Google Scholar] [CrossRef]
- Loeuillard, E.; Yang, J.; Buckarma, E.; Wang, J.; Liu, Y.; Conboy, C.; Pavelko, K.D.; Li, Y.; O’Brien, D.; Wang, C.; et al. Targeting tumor-associated macrophages and granulocytic myeloid-derived suppressor cells augments PD-1 blockade in cholangiocarcinoma. J. Clin. Investig. 2020, 130, 5380–5396. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Chu, T.H.; Nienhüser, H.; Jiang, Z.; Del Portillo, A.; Remotti, H.E.; White, R.A.; Hayakawa, Y.; Tomita, H.; Fox, J.G.; et al. PD-1 Signaling Promotes Tumor-Infiltrating Myeloid-Derived Suppressor Cells and Gastric Tumorigenesis in Mice. Gastroenterology 2021, 160, 781–796. [Google Scholar] [CrossRef]
- Long, H.; Jia, Q.; Wang, L.; Fang, W.; Wang, Z.; Jiang, T.; Zhou, F.; Jin, Z.; Huang, J.; Zhou, L.; et al. Tumor-induced erythroid precursor-differentiated myeloid cells mediate immunosuppression and curtail anti-PD-1/PD-L1 treatment efficacy. Cancer Cell 2022, 40, 674–693.e7. [Google Scholar] [CrossRef]
- Tengesdal, I.W.; Dinarello, A.; Powers, N.E.; Burchill, M.A.; Joosten, L.A.B.; Marchetti, C.; Dinarello, C.A. Tumor NLRP3-Derived IL-1β Drives the IL-6/STAT3 Axis Resulting in Sustained MDSC-Mediated Immunosuppression. Front. Immunol. 2021, 12, 661323. [Google Scholar] [CrossRef]
- Ding, X.; Zhang, J.; Shi, M.; Liu, D.; Zhang, L.; Zhang, R.; Su, B.; Ai, K. High expression level of interleukin-1β is correlated with poor prognosis and PD-1 expression in patients with lung adenocarcinoma. Clin. Transl. Oncol. 2021, 23, 35–42. [Google Scholar] [CrossRef]
- Pretre, V.; Papadopoulos, D.; Regard, J.; Pelletier, M.; Woo, J. Interleukin-1 (IL-1) and the inflammasome in cancer. Cytokine 2022, 153, 155850. [Google Scholar] [CrossRef]
- Paz-Ares, L.; Goto, Y.; Lim, W.D.T.; Halmos, B.; Cho, B.C.; Dols, M.C.; Gonzalez-Larriba, J.L.; Zhou, C.; Demedts, I.; Atmaca, A.; et al. 1194MO Canakinumab (CAN) + docetaxel (DTX) for the second- or third-line (2/3L) treatment of advanced non-small cell lung cancer (NSCLC): CANOPY-2 phase III results. Ann. Oncol. 2021, 32, S953–S954. [Google Scholar] [CrossRef]
- AG NP. Novartis Provides Update on Phase III CANOPY-A Study Evaluating Canakinumab as Adjuvant Treatment in Non-Small Cell Lung Cancer. GlobeNewswire News Room. 2022. Available online: https://www.globenewswire.com/en/news-release/2022/08/15/2497914/0/en/Novartis-provides-update-on-Phase-III-CANOPY-A-study-evaluating-canakinumab-as-adjuvant-treatment-in-non-small-cell-lung-cancer.html (accessed on 28 April 2023).
- Wang, G.C.; Zhou, M.; Zhang, Y.; Cai, H.M.; Chiang, S.T.; Chen, Q.; Han, T.Z.; Li, R.X. Screening and identifying a novel M-MDSCs-related gene signature for predicting prognostic risk and immunotherapeutic responses in patients with lung adenocarcinoma. Front. Genet. 2023, 13, 989141. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T.; Katsuya, Y.; Sato, J.; Koyama, T.; Shimizu, T.; Yamamoto, N. Emerging PD-1/PD-L1 targeting immunotherapy in non-small cell lung cancer: Current status and future perspective in Japan, US, EU, and China. Front. Oncol. 2022, 12, 925938. [Google Scholar] [CrossRef] [PubMed]
- Keegan, A.; Ricciuti, B.; Garden, P.; Cohen, L.; Nishihara, R.; Adeni, A.; Paweletz, C.; Supplee, J.; Jänne, P.A.; Severgnini, M.; et al. Plasma IL-6 changes correlate to PD-1 inhibitor responses in NSCLC. J. Immunother. Cancer 2020, 8, e000678. [Google Scholar] [CrossRef] [PubMed]
- Calu, V.; Ionescu, A.; Stanca, L.; Geicu, O.I.; Iordache, F.; Pisoschi, A.M.; Serban, A.I.; Bilteanu, L. Key biomarkers within the colorectal cancer related inflammatory microenvironment. Sci. Rep. 2021, 11, 7940. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Shapiro, B.; Vucic, E.A.; Vogt, S.; Bar-Sagi, D. Tumor Cell-Derived IL1β Promotes Desmoplasia and Immune Suppression in Pancreatic Cancer. Cancer Res. 2020, 80, 1088–1101. [Google Scholar] [CrossRef]
- Chaturvedi, A.K.; Caporaso, N.E.; Katki, H.A.; Wong, H.L.; Chatterjee, N.; Pine, S.R.; Chanock, S.J.; Goedert, J.J.; Engels, E.A. C-reactive protein and risk of lung cancer. J. Clin. Oncol. 2010, 28, 2719–2726. [Google Scholar] [CrossRef]
- Kanoh, Y.; Abe, T.; Masuda, N.; Akahoshi, T. Progression of non-small cell lung cancer: Diagnostic and prognostic utility of matrix metalloproteinase-2, C-reactive protein and serum amyloid A. Oncol. Rep. 2013, 29, 469–473. [Google Scholar] [CrossRef]
- Riedl, J.M.; Barth, D.A.; Brueckl, W.M.; Zeitler, G.; Foris, V.; Mollnar, S.; Stotz, M.; Rossmann, C.H.; Terbuch, A.; Balic, M.; et al. C-Reactive Protein (CRP) Levels in Immune Checkpoint Inhibitor Response and Progression in Advanced Non-Small Cell Lung Cancer: A Bi-Center Study. Cancers 2020, 12, 2319. [Google Scholar] [CrossRef]
- Assaf, Z.J.F.; Zou, W.; Fine, A.D.; Socinski, M.A.; Young, A.; Lipson, D.; Freidin, J.; Kennedy, M.; Polisecki, E.; Nishio, M.; et al. A longitudinal circulating tumor DNA-based model associated with survival in metastatic non-small-cell lung cancer. Nat. Med. 2023, 29, 859–868. [Google Scholar] [CrossRef]
- Wong, C.C.; Baum, J.; Silvestro, A.; Beste, M.T.; Bharani-Dharan, B.; Xu, S.; Wang, Y.A.; Wang, X.; Prescott, M.F.; Krajkovich, L.; et al. Inhibition of IL1β by Canakinumab May Be Effective against Diverse Molecular Subtypes of Lung Cancer: An Exploratory Analysis of the CANTOS Trial. Cancer Res. 2020, 80, 5597–5605. [Google Scholar] [CrossRef]
- Xiao, K.; Liu, S.; Xiao, Y.; Wang, Y.; Zhu, Z.; Wang, Y.; Tong, D.; Jiang, J. Bioinformatics prediction of differential miRNAs in non-small cell lung cancer. PLoS ONE 2021, 16, e0254854. [Google Scholar] [CrossRef] [PubMed]
- Funkhouser, A.T.; Strigenz, A.M.; Blair, B.B.; Miller, A.P.; Shealy, J.C.; Ewing, J.A.; Martin, J.C.; Funk, C.R.; Edenfield, W.J.; Blenda, A.V. KIT Mutations Correlate with Higher Galectin Levels and Brain Metastasis in Breast and Non-Small Cell Lung Cancer. Cancers 2022, 14, 2781. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Chaudhary, N.; Garg, M.; Floudas, C.S.; Soni, P.; Chandra, A.B. Current Diagnosis and Management of Immune-Related Adverse Events (irAEs) Induced by Immune Checkpoint Inhibitor Therapy. Front. Pharmacol. 2017, 8, 49, Correction in Front. Pharmacol. 2017, 8, 311. [Google Scholar] [CrossRef] [PubMed]
- Gogishvili, M.; Melkadze, T.; Makharadze, T.; Giorgadze, D.; Dvorkin, M.; Penkov, K.; Laktionov, K.; Nemsadze, G.; Nechaeva, M.; Rozhkova, I.; et al. Cemiplimab plus chemotherapy versus chemotherapy alone in non-small cell lung cancer: A randomized, controlled, double-blind phase 3 trial. Nat. Med. 2022, 28, 2374–2380. [Google Scholar] [CrossRef]
- Kelly, K.; Infante, J.R.; Taylor, M.H.; Patel, M.R.; Wong, D.J.; Iannotti, N.; Mehnert, J.M.; Loos, A.H.; Koch, H.; Speit, I.; et al. Safety profile of avelumab in patients with advanced solid tumors: A pooled analysis of data from the phase 1 JAVELIN solid tumor and phase 2 JAVELIN Merkel 200 clinical trials. Cancer 2018, 124, 2010–2017. [Google Scholar] [CrossRef]
- Song, P.; Zhang, D.; Cui, X.; Zhang, L. Meta-analysis of immune-related adverse events of immune checkpoint inhibitor therapy in cancer patients. Thorac. Cancer. 2020, 11, 2406–2430. [Google Scholar] [CrossRef]
- Gross, N.D.; Miller, D.M.; Khushalani, N.I.; Divi, V.; Ruiz, E.S.; Lipson, E.J.; Meier, F.; Su, Y.B.; Swiecicki, P.; Atlas, J.; et al. Neoadjuvant Cemiplimab for Stage II to IV Cutaneous Squamous-Cell Carcinoma. N. Engl. J. Med. 2022, 387, 1557–1568. [Google Scholar] [CrossRef]
- Vaddepally, R.; Doddamani, R.; Sodavarapu, S.; Madam, N.R.; Katkar, R.; Kutadi, A.P.; Mathew, N.; Garje, R.; Chandra, A.B. Review of Immune-Related Adverse Events (irAEs) in Non-Small-Cell Lung Cancer (NSCLC)-Their Incidence, Management, Multiorgan irAEs, and Rechallenge. Biomedicines 2022, 10, 790. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| IL-1β Inhibitors | Type | Target | Trials | Phase, Progress | Experimental Arms | Primary Outcome | Results of the Primary Outcome |
|---|---|---|---|---|---|---|---|
| Canakinumab | human IgGκ mAb | IL-1β | NCT03968419 (CANOPY-N) | Phase II, completed | canakinumab +/− pembrolizumab; pembrolizumab monotherapy | MPR | MPR: 2.9% (CAN)/17.1% (CAN + PEM)/11.1% (PEM), does not meet the primary endpoint |
| NCT03626545 (CANOPY-2) | Phase III, completed | docetaxel +/− canakinumab | OS and incidence of DLTs | Median OS: 10.5 m (CAN) /11.3 m (Placebo) (HR 1.06 95% CI, 0.76–1.48); does not meet the primary endpoint | |||
| NCT03631199 (CANOPY-1) | Phase III, active, not recruiting | pembrolizumab + platinum-based doublet chemotherapy +/− canakinumab | OS, PFS, and incidence DLTs | Median OS: 20.8 m (CAN) /20.2 m (Placebo) (HR 0.87, 95% CI, 0.70–1.10; one-sided p = 0.123); median PFS: 6.8 m for both treatment arms (hazard ratio (HR), 0.85; 95% CI, 0.67–1.09; p = 0.1); does not meet the primary endpoint | |||
| NCT03447769 (CANOPY-A) | Phase III, terminated | Canakinumab (Canakinumab versus Placebo | DFS | Median DFS: 35 m (CAN)/ 29.7 m (Placebo) (HR 0.94; 95% CI 0.78–1.14; one-sided p = 0.258); does not meet the primary endpoint | |||
| NCT04905316 (CHORUS) | Phase II, recruiting | Canakinumab + Chemoradiation + Durvalumab (Single-arm, prospective, phase I/II study) | PFS | Recruiting, no result yet |
| IL-1 Inhibitors | Type | Target | Trials | Phase, Progress | Experimental Arms | Primary Outcome | Results of the Primary Outcome |
|---|---|---|---|---|---|---|---|
| Anakinra | IL-1 receptor mAb | IL-1 receptor | NCT01624766 | Phase I, completed | Everolimus (mTOR Inhibitor) + Anakinra/Denosumab | Incidence of AEs, MTD of everolimus | No result published yet |
| IL-1R inhibitors | Type | Target | Trials | Phase, Progress | Experimental Arms | Primary Outcome | Results of the Primary Outcome |
| Nadunolimab (CAN04) | First-in-class fully humanized and ADCC enhanced mAb | IL1RAP | NCT03267316 (CANFOUR) | Phase I/II, Recruiting | CAN04 +/− standard of care treatment | Incidence of Treatment-Emergent AE (Safety and Tolerability) | Infusion-related reactions (41%), fatigue (32%), constipation (27%), diarrhea (27%), decreased appetite (23%), nausea (23%), and vomiting (23%); ORR 53% |
| NCT05116891 (CESTAFOUR) | Phase I/II, active, not recruiting | CAN04 + chemotherapy (mFOLFOX or DTX or G/C) | ORR, frequency, duration, and severity of AEs | No result yet | |||
| NCT04452214 (CIRIFOUR) | Phase I, active, not recruiting | CAN04 + pembrolizumab +/− carboplatin and pemetrexed | TEAEs, DLTs, SAEs | No result yet |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillo, D.R.; Jeon, W.J.; Park, D.; Pham, B.; Yang, C.; Joung, B.; Moon, J.H.; Lee, J.; Chong, E.G.; Park, K.; et al. Comprehensive Review: Unveiling the Pro-Oncogenic Roles of IL-1ß and PD-1/PD-L1 in NSCLC Development and Targeting Their Pathways for Clinical Management. Int. J. Mol. Sci. 2023, 24, 11547. https://doi.org/10.3390/ijms241411547
Castillo DR, Jeon WJ, Park D, Pham B, Yang C, Joung B, Moon JH, Lee J, Chong EG, Park K, et al. Comprehensive Review: Unveiling the Pro-Oncogenic Roles of IL-1ß and PD-1/PD-L1 in NSCLC Development and Targeting Their Pathways for Clinical Management. International Journal of Molecular Sciences. 2023; 24(14):11547. https://doi.org/10.3390/ijms241411547
Chicago/Turabian StyleCastillo, Dani Ran, Won Jin Jeon, Daniel Park, Bryan Pham, Chieh Yang, Bowon Joung, Jin Hyun Moon, Jae Lee, Esther G. Chong, Kiwon Park, and et al. 2023. "Comprehensive Review: Unveiling the Pro-Oncogenic Roles of IL-1ß and PD-1/PD-L1 in NSCLC Development and Targeting Their Pathways for Clinical Management" International Journal of Molecular Sciences 24, no. 14: 11547. https://doi.org/10.3390/ijms241411547
APA StyleCastillo, D. R., Jeon, W. J., Park, D., Pham, B., Yang, C., Joung, B., Moon, J. H., Lee, J., Chong, E. G., Park, K., Reeves, M. E., Duerksen-Hughes, P., Mirshahidi, H. R., & Mirshahidi, S. (2023). Comprehensive Review: Unveiling the Pro-Oncogenic Roles of IL-1ß and PD-1/PD-L1 in NSCLC Development and Targeting Their Pathways for Clinical Management. International Journal of Molecular Sciences, 24(14), 11547. https://doi.org/10.3390/ijms241411547