
Beneficial Effects of Astragalus membranaceus (Fisch.) Bunge Extract in Controlling Inflammatory Response and Preventing Asthma Features

 ,
,  , ,
, ,  ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Results
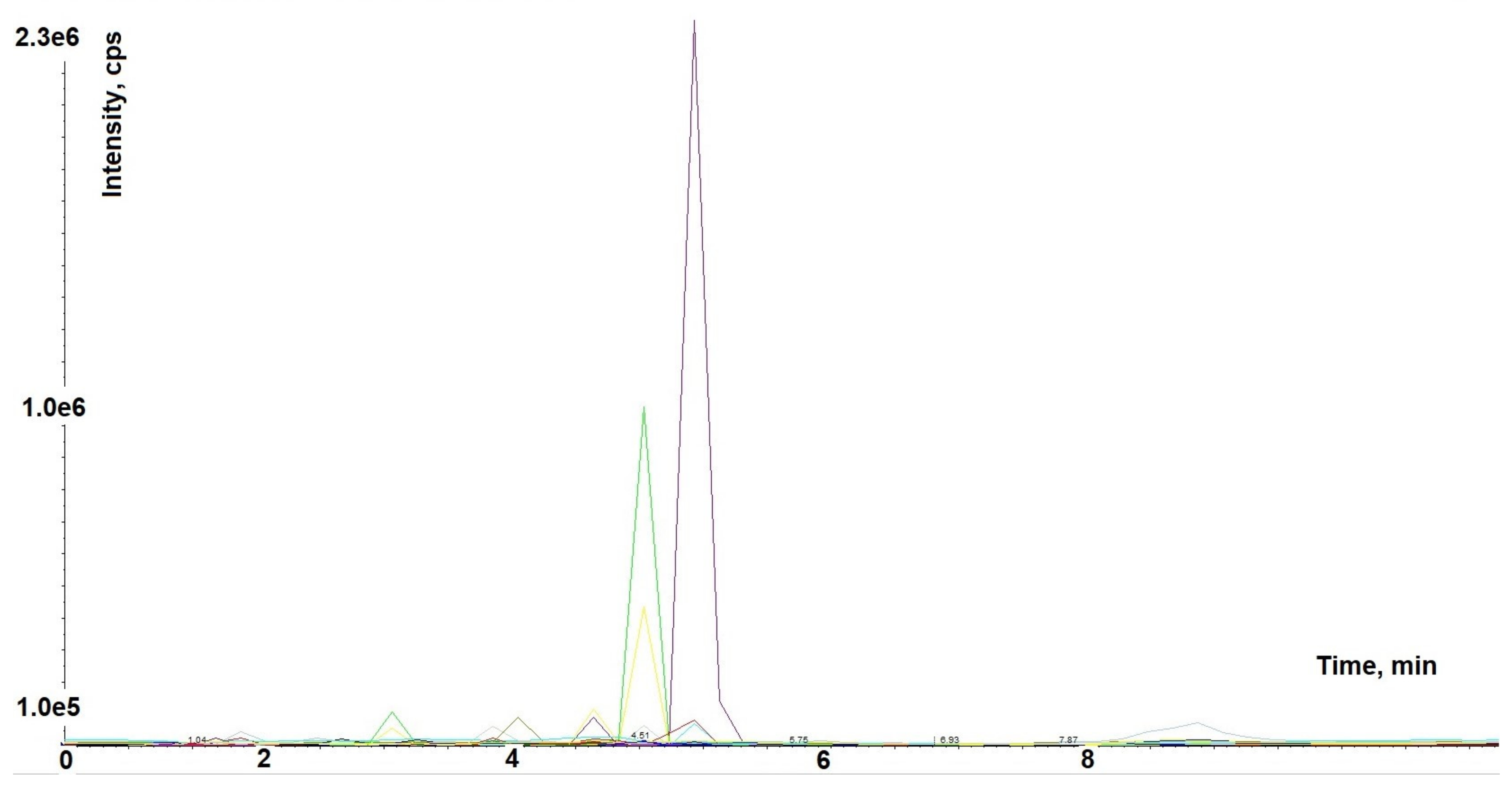
2.1. Astragalus membranaceus Extract Chromatographic Analysis
2.2. Astragalus membranaceus Extract Inhibits Macrophage Activation
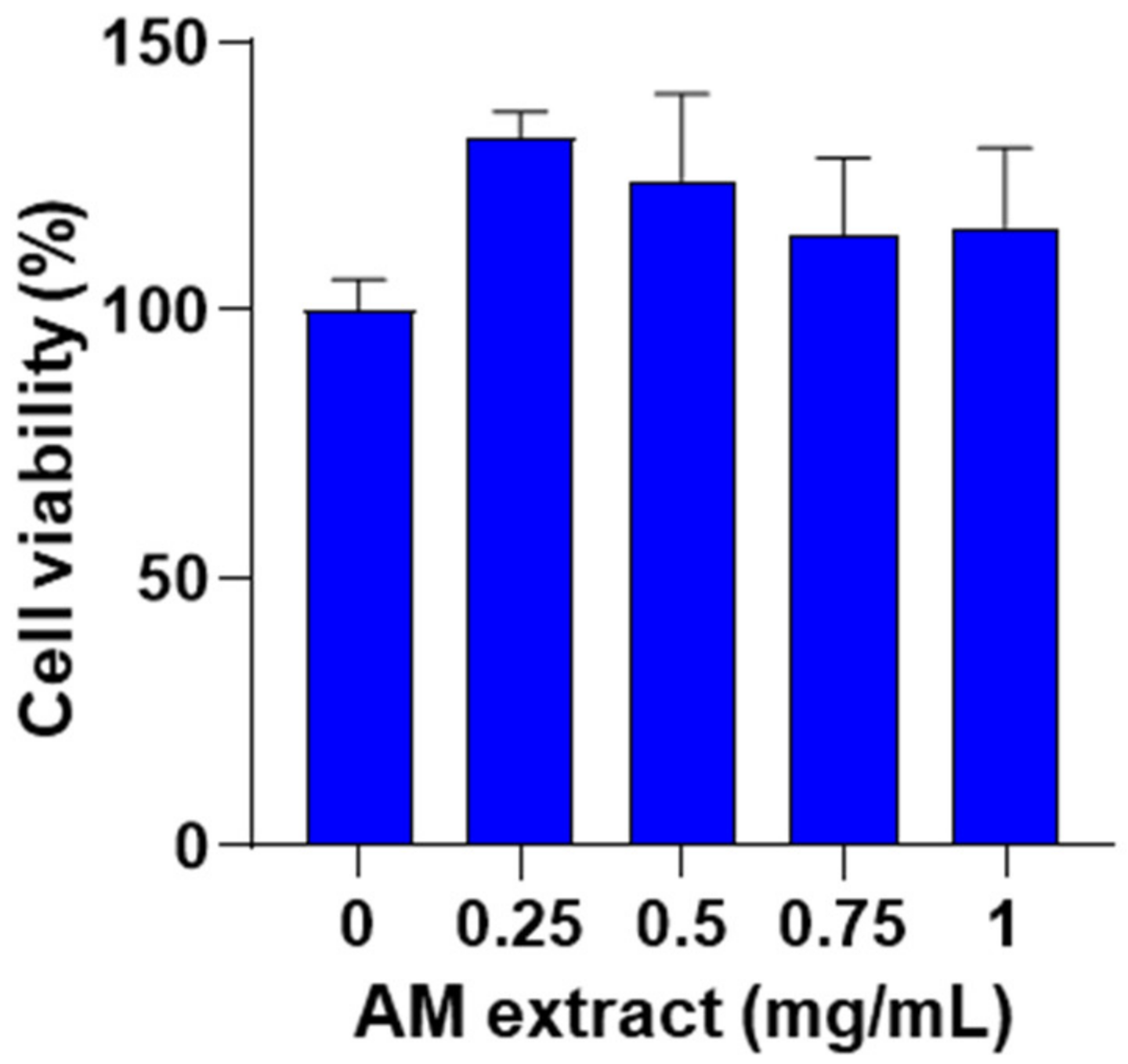
2.2.1. Effect of Astragalus membranaceus Extract on Cell Viability
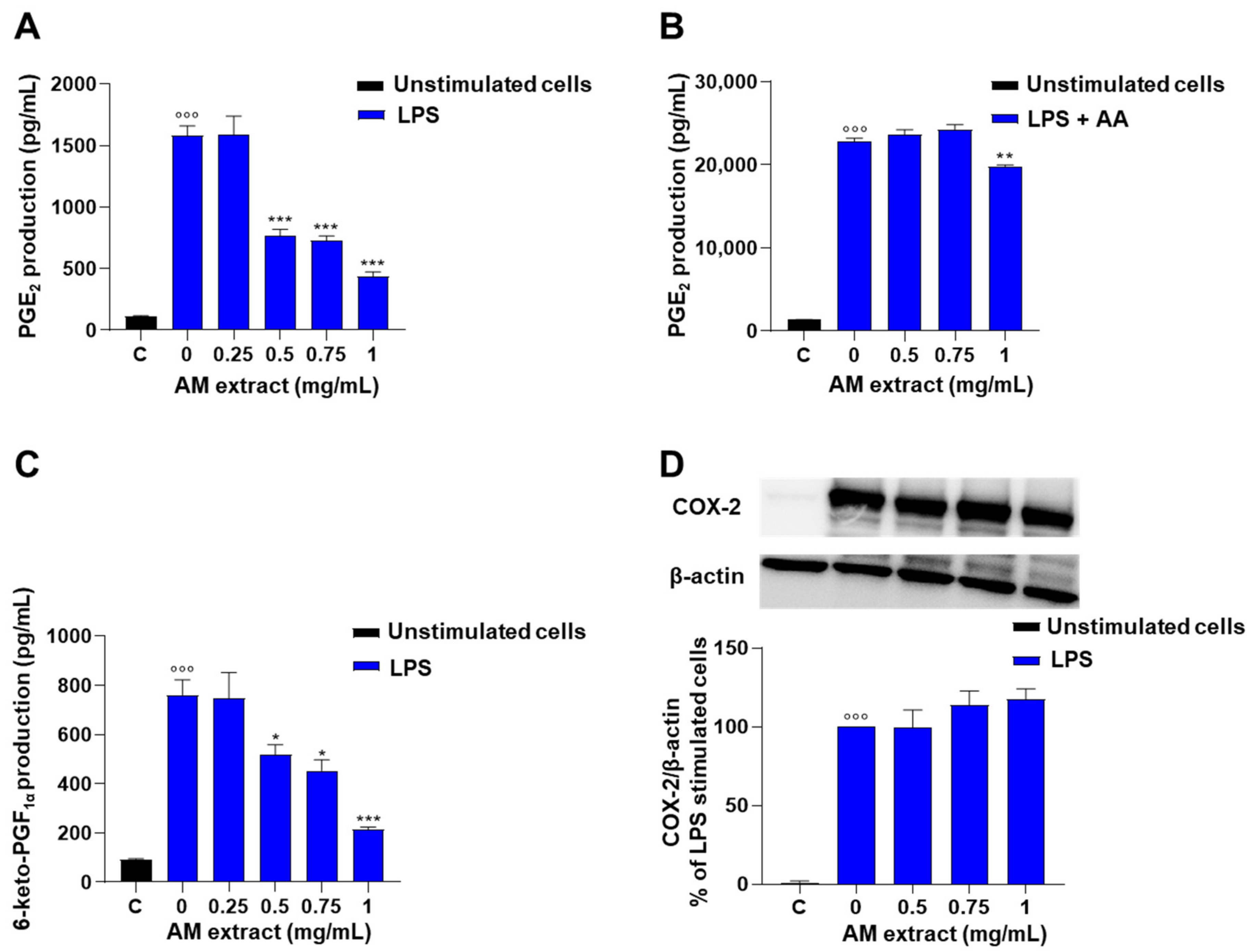
2.2.2. Astragalus membranaceus Extract Inhibits LPS-Induced PG Production in J774 Macrophages
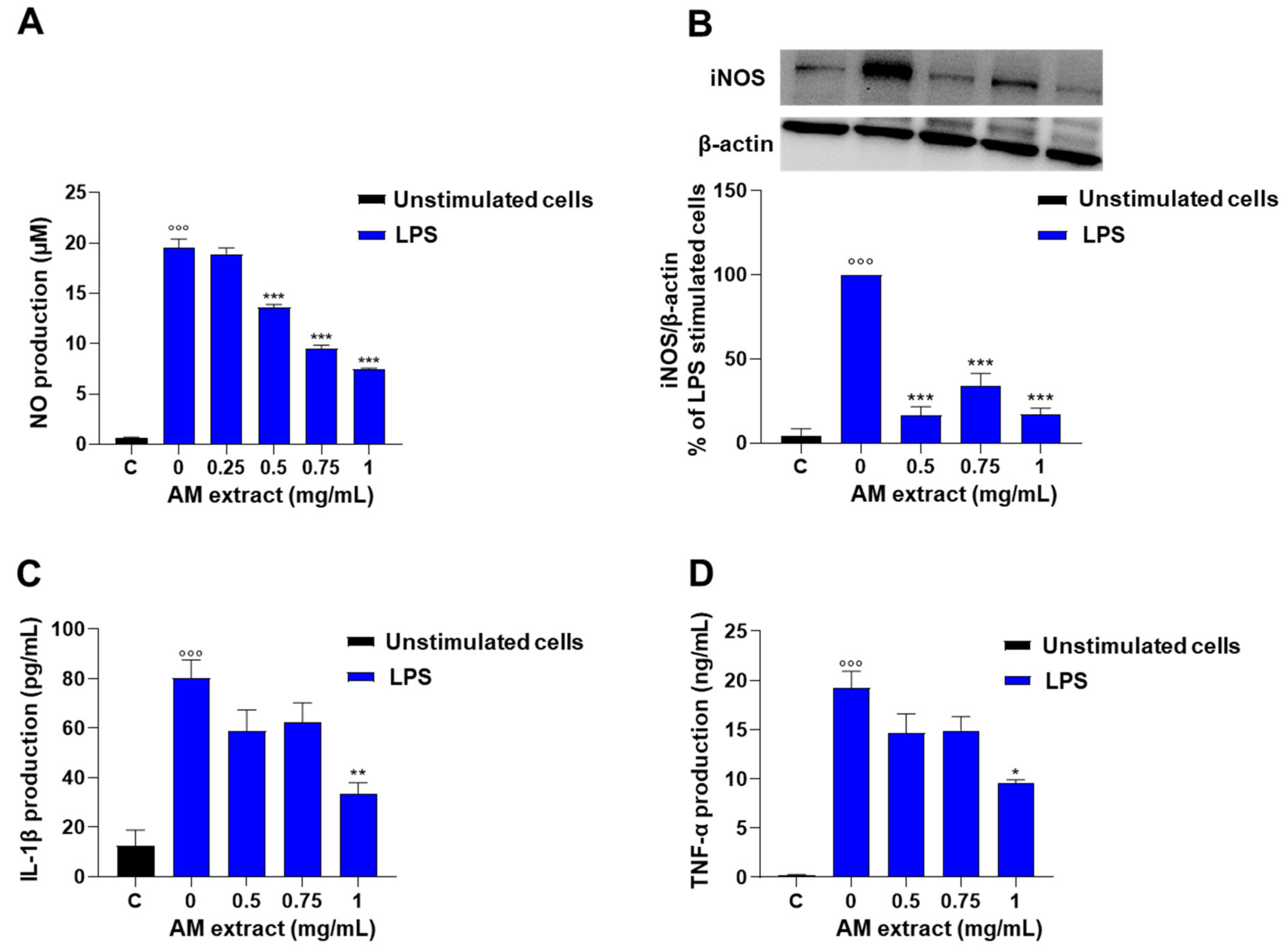
2.2.3. Astragalus membranaceus Extract Inhibits LPS-Induced Nitrite Production in J774 Macrophages
2.2.4. Astragalus membranaceus Extract Reduces LPS-Induced IL-1β and TNF-α Production in J774 Macrophages
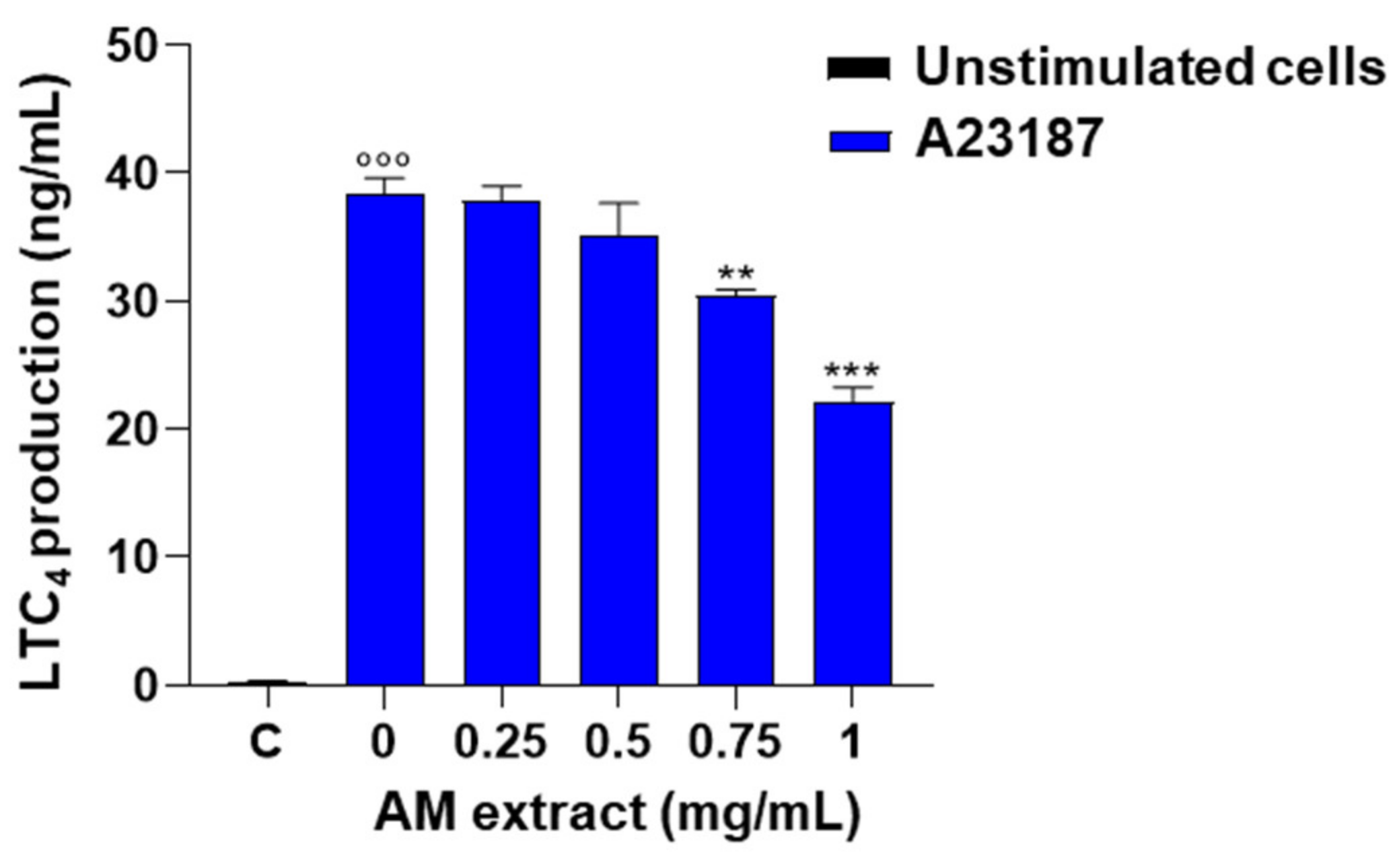
2.2.5. Astragalus membranaceus Extract Reduces LTC4 Production in A23187-Activated Peritoneal Macrophages
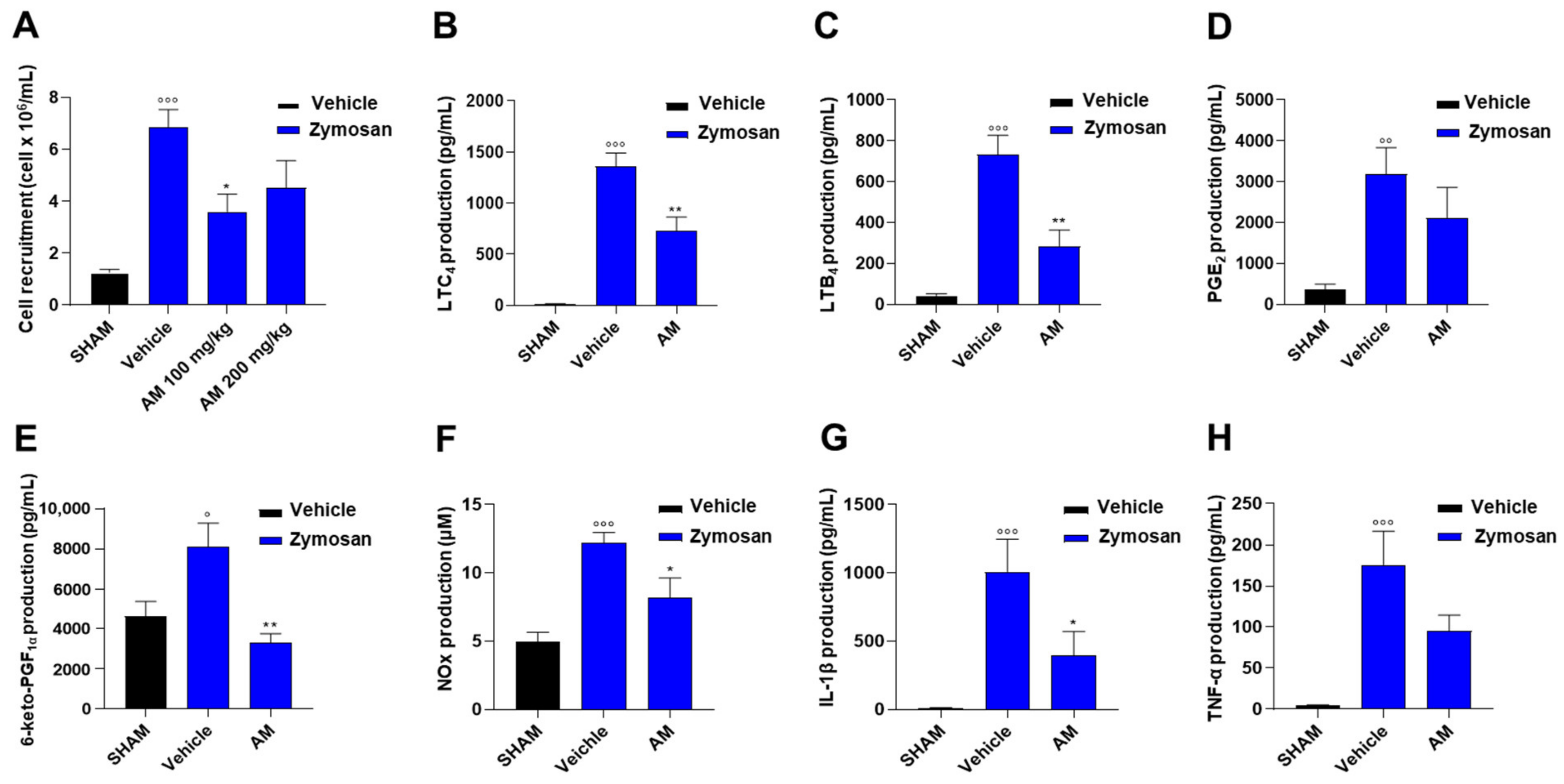
2.3. Astragalus membranaceus Extract Attenuates Inflammation in Murine Peritonitis
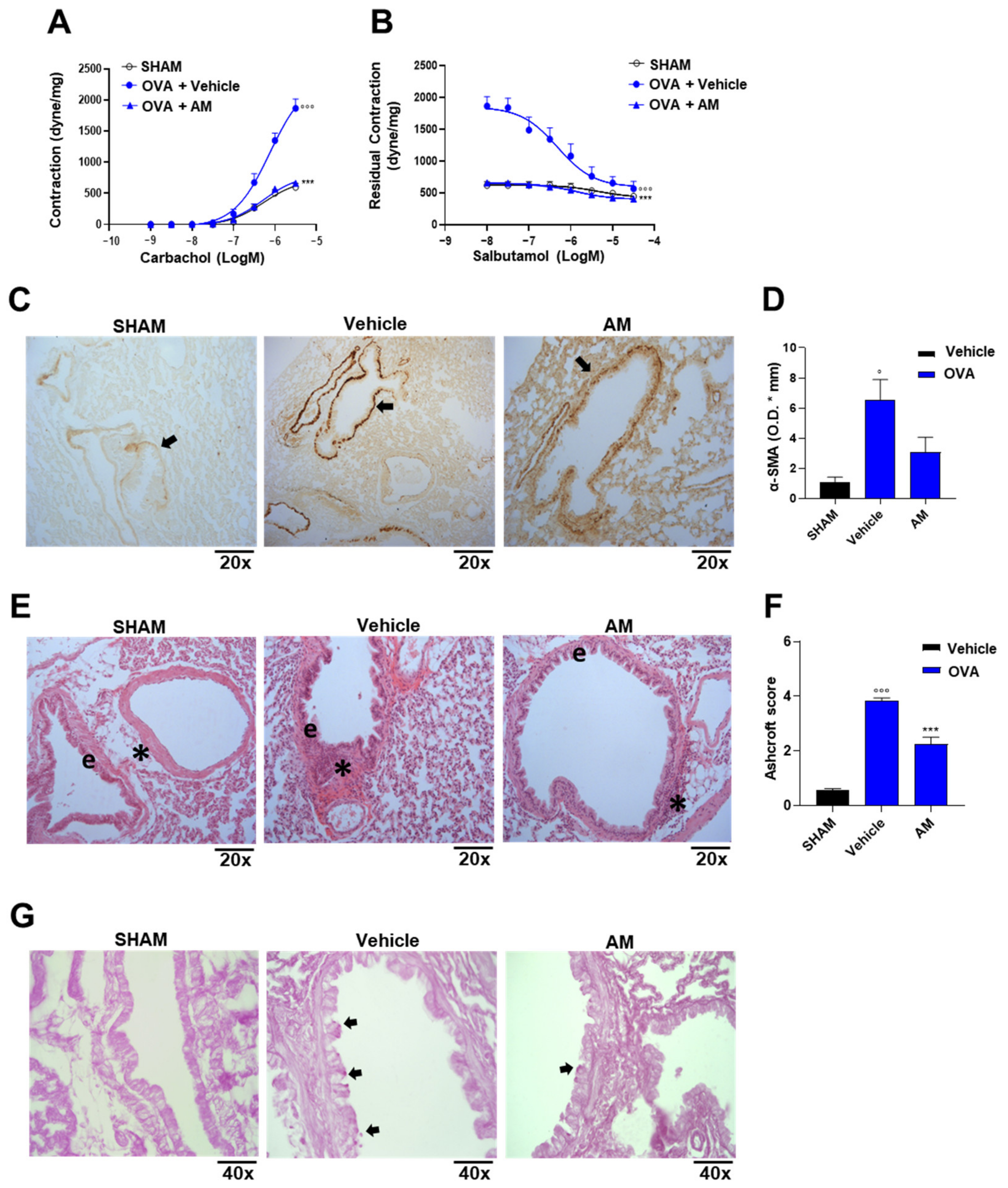
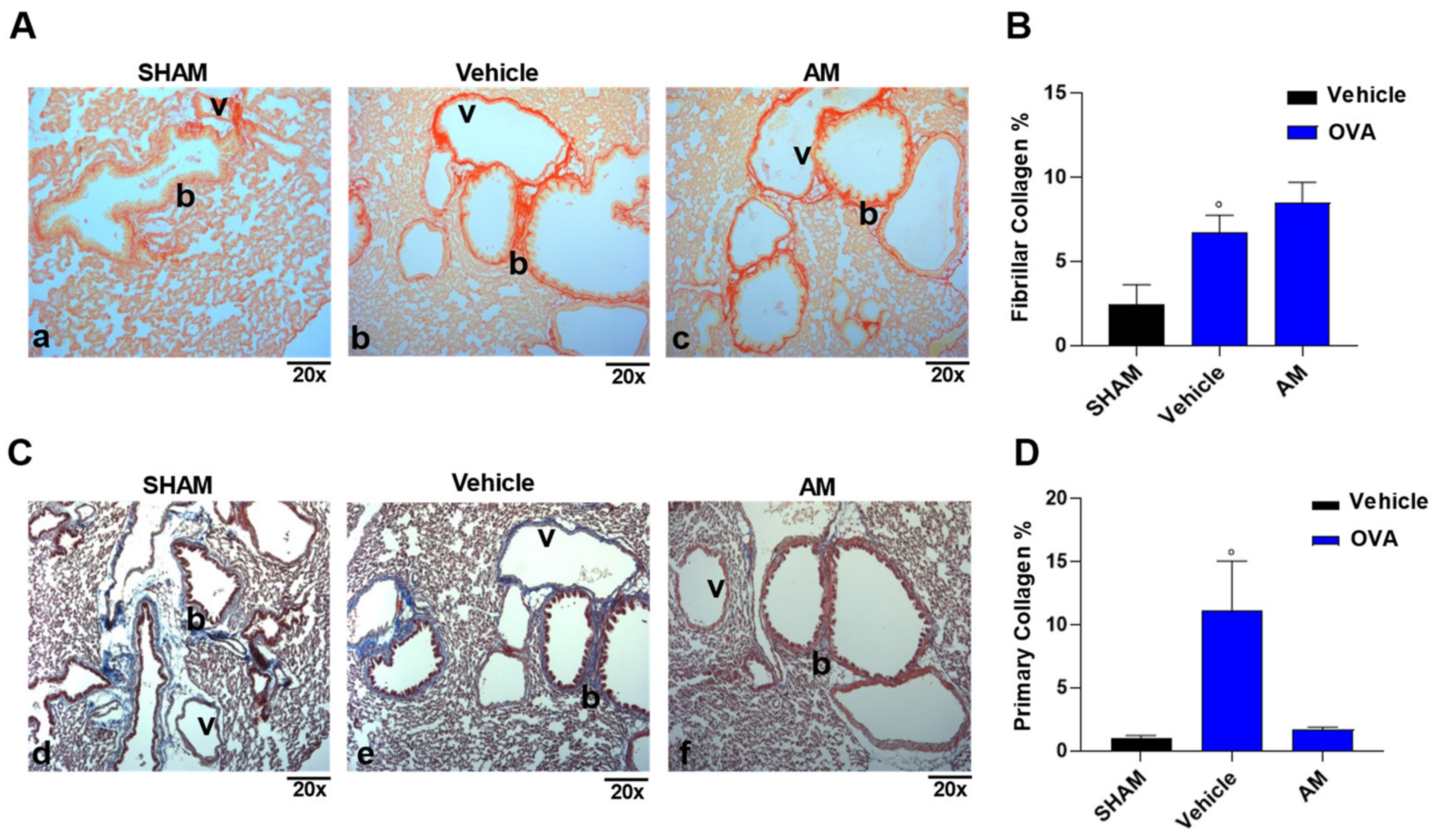
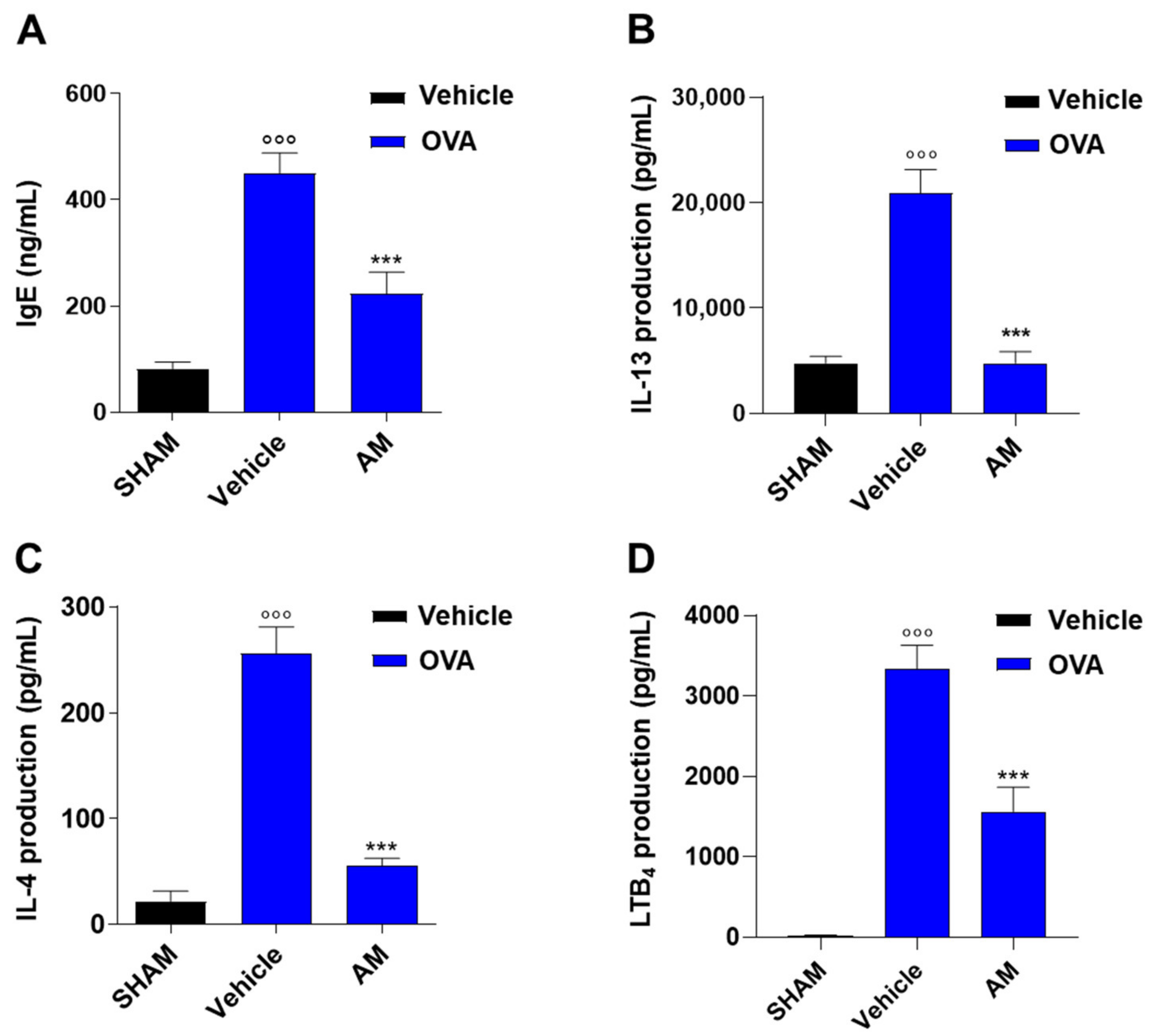
2.4. Astragalus membranaceus Extract Exhibits Beneficial Effect on Asthma Features
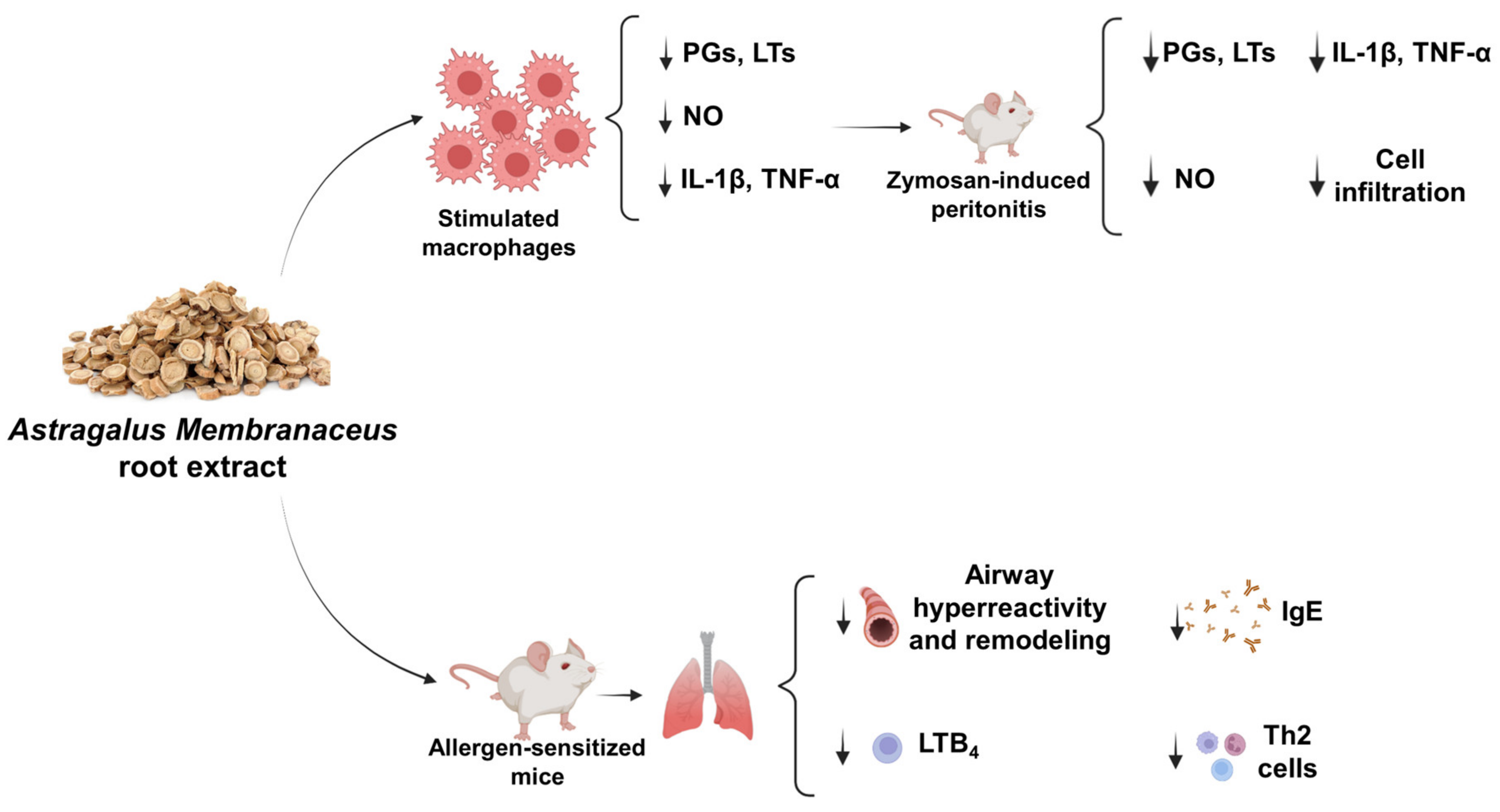
3. Discussion
4. Materials and Methods
4.1. Astragalus membranaceus Extract
4.2. Astragalus membranaceus Extract Analysis by HPLC-MS/MS in Multiple Reaction Monitoring
4.3. J774 Cell Culture
4.3.1. Cell Viability
4.3.2. Western Blot Analysis
4.4. Animals
4.5. Resident Peritoneal Macrophages
4.6. Zymosan-Induced Peritonitis
4.7. OVA Sensitization
4.7.1. Bronchial Reactivity
4.7.2. Lung Histology
4.7.3. Immunohistochemistry
4.7.4. Measurement of Plasma IgE Levels
4.7.5. IL-13, IL-4 and LTB4 Levels
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fu, J.; Wang, Z.; Huang, L.; Zheng, S.; Wang, D.; Chen, S.; Zhang, H.; Yang, S. Review of the botanical characteristics, phytochemistry, and pharmacology of Astragalus membranaceus (Huangqi). Phytother. Res. 2014, 28, 1275–1283. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press (CFDA): Beijing, China, 2015. [Google Scholar]
- The United States Pharmacopeial Convention. U.S. Pharmacopeia/National Formulary; The United States Pharmacopeial Convention: New York, NY, USA, 2017. [Google Scholar]
- European Directorate for the Quality of Medicines & HealthCare. European Pharmacopoeia 8.0; European Directorate for the Quality of Medicines & HealthCare: Strasbourg, France, 2017. [Google Scholar]
- Japanese Pharmacopoeia Commission. The Japanese Pharmacopoeia; Ministry of Health and Welfare: Sejong, Republic of Korea, 2016.
- Korea Food and Drug Administration. Korean Pharmacopoeia; Korea Food and Drug Administration: Seoul, Republic of Korea, 2016.
- Guo, Z.; Lou, Y.; Kong, M.; Luo, Q.; Liu, Z.; Wu, J. A Systematic Review of Phytochemistry, Pharmacology and Pharmacokinetics on Astragali Radix: Implications for Astragali Radix as a Personalized Medicine. Int. J. Mol. Sci. 2019, 20, 1463. [Google Scholar] [CrossRef]
- Lee, K.Y.; Jeon, Y.J. Macrophage activation by polysaccharide isolated from Astragalus membranaceus. Int. Immunopharmacol. 2005, 5, 1225–1233. [Google Scholar] [CrossRef]
- Lee, Y.S.; Han, O.K.; Park, C.W.; Yang, C.H.; Jeon, T.W.; Yoo, W.K.; Kim, S.H.; Kim, H.J. Pro-inflammatory cytokine gene expression and nitric oxide regulation of aqueous extracted Astragali radix in RAW 264.7 macrophage cells. J. Ethnopharmacol. 2005, 100, 289–294. [Google Scholar] [CrossRef]
- Luo, Y.; Jin, M.; Lou, L.; Yang, S.; Li, C.; Li, X.; Zhou, M.; Cai, C. Role of arachidonic acid lipoxygenase pathway in Asthma. Prostaglandins Other Lipid Mediat. 2022, 158, 106609. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Q.; Li, Z.; Gao, Y.; Pang, Z.; Wu, Y.; Li, G.; Lu, D.; Zhang, L.; Li, D. Astragalus polysaccharides attenuate ovalbumin-induced allergic rhinitis in rats by inhibiting NLRP3 inflammasome activation and NOD2-mediated NF-κB activation. J. Med. Food. 2021, 24, 1–9. [Google Scholar] [CrossRef]
- Rossi, A.; Pergola, C.; Pace, S.; Rådmark, O.; Werz, O.; Sautebin, L. In vivo sex differences in leukotriene biosynthesis in zymosan-induced peritonitis. Pharmacol. Res. 2014, 87, 1–7. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef]
- Hamza, A.A.; Lashin, F.M.; Gamel, M.; Hassanin, S.O.; Abdalla, Y.; Amin, A. Hawthorn Herbal Preparation from Crataegus oxyacantha Attenuates In Vivo Carbon Tetrachloride -Induced Hepatic Fibrosis via Modulating Oxidative Stress and Inflammation. Antioxidants 2020, 9, 1173. [Google Scholar] [CrossRef]
- Juaid, N.; Amin, A.; Abdalla, A.; Reese, K.; Alamri, Z.; Moulay, M.; Abdu, S.; Miled, N. Anti-Hepatocellular Carcinoma Biomolecules: Molecular Targets Insights. Int. J. Mol. Sci. 2021, 22, 10774. [Google Scholar] [CrossRef]
- Cha, R.J.; Zeng, D.W.; Chang, Q.S. Non-surgical treatment of small cell lung cancer with chemo-radio-immunotherapy and traditional Chinese medicine. Zhonghua Nei Ke Za Zhi 1994, 33, 462–466. [Google Scholar]
- Kusum, M.; Klinbuayaem, V.; Bunjob, M.; Sangkitporn, S. Preliminary efficacy and safety of oral suspension SH, combination of five chinese medicinal herbs, in people living with HIV/AIDS; the phase I/II study. J. Med. Assoc. Thail. 2004, 87, 1065–1067. [Google Scholar]
- Tolleson, W.H.; Doerge, D.R.; Churchwell, M.I.; Marques, M.M.; Roberts, D.W. Metabolism of Biochanin A and Formononetin by Human Liver Microsomes in Vitro. J. Agric. Food Chem. 2002, 50, 4783–4790. [Google Scholar] [CrossRef]
- Govindasami, S.; Uddandrao, V.V.S.; Raveendran, N.; Sasikumar, V. Therapeutic Potential of Biochanin-A Against Isoproterenol-Induced Myocardial Infarction in Rats. Cardiovasc. Hematol. Agents Med. Chem. 2020, 18, 31–36. [Google Scholar] [CrossRef]
- Hsu, Y.L.; Kuo, P.L. Diosmetin induces human osteoblastic differentiation through the protein kinase C/p38 and extracellular signal-regulated kinase 1/2 pathway. J. Bone Miner. Res. 2008, 23, 949–960. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Tejada, S.; Setzer, W.N.; Gortzi, O.; Sureda, A.; Braidy, N.; Daglia, M.; Manayi, A.; Nabavi, S.M. Chlorogenic Acid and Mental Diseases: From Chemistry to Medicine. Curr. Neuropharmacol. 2017, 15, 471–479. [Google Scholar] [CrossRef]
- Mahmoudi, M.; Abdellaoui, R.; Boughalleb, F.; Yahia, B.; Mabrouk, M.; Nasri, N. Characterization of lipids, proteins, and bioactive compounds in the seeds of three Astragalus species. Food Chem. 2021, 339, 127824. [Google Scholar] [CrossRef]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef]
- Hara, S.; Kamei, D.; Sasaki, Y.; Tanemoto, A.; Nakatani, Y.; Murakami, M. Prostaglandin E synthases: Understanding their pathophysiological roles through mouse genetic models. Biochimie 2010, 92, 651–659. [Google Scholar] [CrossRef]
- Ryu, M.; Kim, E.H.; Chun, M.; Kang, S.; Shim, B.; Yu, Y.-B.; Jeong, G.; Lee, J.-S. Astragali Radix elicits anti-inflammation via activation of MKP-1, concomitant with attenuation of p38 and Erk. J. Ethnopharmacol. 2008, 115, 184–193. [Google Scholar] [CrossRef]
- Lai, P.K.; Chan, J.Y.; Wu, S.B.; Cheng, L.; Ho, G.K.; Lau, C.P.; Kennelly, E.J.; Leung, P.C.; Fung, K.P.; Lau, C.B. Anti-inflammatory activities of an active fraction isolated from the root of Astragalus membranaceus in RAW 264.7 macrophages. Phytother. Res. 2014, 28, 395–404. [Google Scholar] [CrossRef]
- Jun, M.; Hong, J.; Jeong, W.-S.; Ho, C.-T. Suppression of arachidonic acid metabolism and nitric oxide formation by kudzu isoflavones in murine macrophages. Mol. Nutr. Food Res. 2005, 49, 1154–1159. [Google Scholar] [CrossRef]
- Rasheed, A.; Rayner, K.J. Macrophage Responses to Environmental Stimuli During Homeostasis and Disease. Endocr. Rev. 2021, 42, 407–435. [Google Scholar] [CrossRef]
- Tay, K.-C.; Tan, L.T.-H.; Chan, C.K.; Hong, S.L.; Chan, K.-G.; Yap, W.H.; Pusparajah, P.; Lee, L.-H.; Goh, B.-H. Formononetin: A Review of Its Anticancer Potentials and Mechanisms. Front. Pharmacol. 2019, 10, 820. [Google Scholar] [CrossRef]
- Singh, L.; Kaur, N.; Bhatti, R. Neuroprotective potential of biochanin-A and review of the molecular mechanisms involved. Mol. Biol. Rep. 2023, 50, 5369–5378. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, G.; Chen, J.; Liu, X.; Lu, X.; Chen, H.; Tian, Y. Integrated Metabolome and Transcriptome Analysis Unveils Novel Pathway Involved in the Formation of Yellow Peel in Cucumber. Int. J. Mol. Sci. 2021, 22, 1494. [Google Scholar] [CrossRef]
- Matkovic, Z.; Zivkovic, V.; Korica, M.; Plavec, D.; Pecanic, S.; Tudoric, N. Efficacy and safety of Astragalus membranaceus in the treatment of patients with seasonal allergic rhinitis. Phytother. Res. 2010, 24, 175–181. [Google Scholar] [CrossRef]
- Xu, S.; Tian, B.P.; Zhang, L.H.; Hua, W.; Xia, L.X.; Chen, Z.H.; Li, W.; Shen, H.H. Prevention of allergic airway hyperresponsiveness and remodeling in mice by Astragali Radix antiasthmatic decoction. BMC Complement. Altern. Med. 2013, 13, 369. [Google Scholar] [CrossRef]
- Cerqua, I.; Musella, S.; Peltner, L.K.; D’avino, D.; Di Sarno, V.; Granato, E.; Vestuto, V.; Di Matteo, R.; Pace, S.; Ciaglia, T.; et al. Discovery and Optimization of Indoline-Based Compounds as Dual 5-LOX/sEH Inhibitors: In Vitro and In Vivo Anti-Inflammatory Characterization. J. Med. Chem. 2022, 65, 14456–14480. [Google Scholar] [CrossRef]
- Ullah, H.; Sommella, E.; Santarcangelo, C.; D’avino, D.; Rossi, A.; Dacrema, M.; Di Minno, A.; Di Matteo, G.; Mannina, L.; Campiglia, P.; et al. Hydroethanolic Extract of Prunus domestica L.: Metabolite Profiling and In Vitro Modulation of Molecular Mechanisms Associated to Cardiometabolic Diseases. Nutrients 2022, 14, 340. [Google Scholar] [CrossRef]
- Rossi, A.; Pace, S.; Tedesco, F.; Pagano, E.; Guerra, G.; Troisi, F.; Werner, M.; Roviezzo, F.; Zjawiony, J.K.; Werz, O.; et al. The hallucinogenic diterpene salvinorin A inhibits leukotriene synthesis in experimental models of inflammation. Pharmacol. Res. 2016, 106, 64–71. [Google Scholar] [CrossRef]
- Rossi, A.; Caiazzo, E.; Bilancia, R.; Riemma, M.A.; Pagano, E.; Cicala, C.; Ialenti, A.; Zjawiony, J.K.; Izzo, A.A.; Capasso, R.; et al. Salvinorin A Inhibits Airway Hyperreactivity Induced by Ovalbumin Sensitization. Front. Pharmacol. 2017, 7, 525. [Google Scholar] [CrossRef]
- Roviezzo, F.; Rossi, A.; Caiazzo, E.; Orlando, P.; Riemma, M.A.; Iacono, V.M.; Guarino, A.; Ialenti, A.; Cicala, C.; Peritore, A.; et al. Palmitoylethanolamide Supplementation during Sensitization Prevents Airway Allergic Symptoms in the Mouse. Front. Pharmacol. 2017, 8, 857. [Google Scholar] [CrossRef]
- Cerqua, I.; Terlizzi, M.; Bilancia, R.; Riemma, M.A.; Citi, V.; Martelli, A.; Pace, S.; Spaziano, G.; D’agostino, B.; Werz, O.; et al. 5α-dihydrotestosterone abrogates sex bias in asthma like features in the mouse. Pharmacol. Res. 2020, 158, 104905. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | µg/g | SD (±) |
|---|---|---|
| Formononetin | 140.35 | 3.28 |
| Biochanin A | 63.17 | 3.19 |
| Glycitein | 30.67 | 1.81 |
| Diosmetin | 11.85 | 0.30 |
| Hyperoside | 9.58 | 0.38 |
| Chlorogenic acid | 7.62 | 0.36 |
| Apigenin-8-C-glucoside (Vitexin) | 5.65 | 0.40 |
| Luteolin | 4.96 | 0.08 |
| Kaempferol | 4.77 | 0.40 |
| Caffeoylquinic acid derivative | 4.47 | 0.11 |
| Ferulic acid | 4.29 | 0.20 |
| Naringenin | 4.15 | 0.22 |
| Pelargonidin | 4.01 | 0.03 |
| Cyanidin | 3.56 | 0.08 |
| Astragalin | 3.42 | 0.12 |
| Delphinidin | 3.33 | 0.15 |
| Naringenin-7-O-glucoside | 3.27 | 0.09 |
| Caffeoyliquinic acid lactone | 2.91 | 0.07 |
| Luteolin-6-glucoside (Isoorientin) | 2.77 | 0.11 |
| Quercetin-3-O-galactoside | 2.75 | 0.20 |
| Kaempeferol-3-O-rhamnoside | 2.68 | 0.13 |
| Eriodictyol | 2.67 | 0.09 |
| Rutin | 2.64 | 0.03 |
| Quercetin-O-hexoside | 2.63 | 0.09 |
| Piceatannol | 2.57 | 0.12 |
| Caffeine | 2.52 | 0.07 |
| Myricetin | 2.48 | 0.12 |
| Haesperetin | 2.48 | 0.08 |
| Resveratrol | 2.47 | 0.16 |
| Quercetin | 2.46 | 0.12 |
| Kaempferol-3-O-rutinoside | 2.46 | 0.11 |
| Quercetin-O-rhamnoside | 2.42 | 0.06 |
| Naringin | 2.40 | 0.14 |
| Linarin | 2.32 | 0.10 |
| Isorhamnetin | 2.30 | 0.09 |
| Quercetin acetylhexoside | 2.30 | 0.05 |
| EGC gallate glucoside | 2.30 | 0.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Avino, D.; Cerqua, I.; Ullah, H.; Spinelli, M.; Di Matteo, R.; Granato, E.; Capasso, R.; Maruccio, L.; Ialenti, A.; Daglia, M.; et al. Beneficial Effects of Astragalus membranaceus (Fisch.) Bunge Extract in Controlling Inflammatory Response and Preventing Asthma Features. Int. J. Mol. Sci. 2023, 24, 10954. https://doi.org/10.3390/ijms241310954
D’Avino D, Cerqua I, Ullah H, Spinelli M, Di Matteo R, Granato E, Capasso R, Maruccio L, Ialenti A, Daglia M, et al. Beneficial Effects of Astragalus membranaceus (Fisch.) Bunge Extract in Controlling Inflammatory Response and Preventing Asthma Features. International Journal of Molecular Sciences. 2023; 24(13):10954. https://doi.org/10.3390/ijms241310954
Chicago/Turabian StyleD’Avino, Danilo, Ida Cerqua, Hammad Ullah, Michele Spinelli, Rita Di Matteo, Elisabetta Granato, Raffaele Capasso, Lucianna Maruccio, Armando Ialenti, Maria Daglia, and et al. 2023. "Beneficial Effects of Astragalus membranaceus (Fisch.) Bunge Extract in Controlling Inflammatory Response and Preventing Asthma Features" International Journal of Molecular Sciences 24, no. 13: 10954. https://doi.org/10.3390/ijms241310954
APA StyleD’Avino, D., Cerqua, I., Ullah, H., Spinelli, M., Di Matteo, R., Granato, E., Capasso, R., Maruccio, L., Ialenti, A., Daglia, M., Roviezzo, F., & Rossi, A. (2023). Beneficial Effects of Astragalus membranaceus (Fisch.) Bunge Extract in Controlling Inflammatory Response and Preventing Asthma Features. International Journal of Molecular Sciences, 24(13), 10954. https://doi.org/10.3390/ijms241310954