Abstract
Cucumber (Cucumis sativus L.) is an important vegetable worldwide, but its yield is affected by a wide range of pathogens and pests. As the major subunit of the exocyst complex, the roles of Exo70 members have been shown in Arabidopsis and rice, but their function are unknown in cucumber. Here, we identified 18 CsExo70 members in cucumber, which were divided into three groups (Exo70.1–Exo70.3) and nine subgroups (Exo70A–Exo70I) based on the phylogenetic tree. Subsequently, systematical analyses were performed, including collinearity, gene structure, cis-acting elements, conserved motifs, expression patterns, and subcellular localization. Our results showed that CsExo70 genes were generally expressed in all tissues, and CsExo70C1 and CsExo70C2 were highly expressed in the stamen. Moreover, the expression levels of most CsExo70 genes were induced by Pseudomonas syringae pv. lachrymans (Psl) and Fusarium oxysporum f. sp. cucumerinum Owen (Foc), especially CsExo70E2 and CsExo70H3. In addition, these CsExo70s displayed similar location patterns with discrete and punctate signals in the cytoplasm. Together, our results indicate that CsExo70 members may be involved in plant development and resistance, and provide a reference for future in-depth studies of Exo70 genes in cucumber.
1. Introduction
Secretion is a vital cellular process and is responsible for the transport of newly synthesized materials, ending with the exocytosis event at the plasma membrane (PM) in eukaryotes [1]. De novo cargoes of a vast array of proteins, including PM proteins, signaling peptides, or small RNAs, are transported to the PM or extracellular space by vesicles [2,3,4]. Prior to soluble N-ethylmaleimide-sensitive factor (NSF) attachment protein receptor (SNARE)-mediated membrane fusion, the first attachment of secretory vesicles to the target PM is mediated by the exocyst complex [1,5], which was firstly identified in budding yeast (Saccharomyces cerevisiae). The exocyst complex consists of eight subunits: SEC3, SEC5, SEC6, SEC8, SEC10, SEC15, Exo70, and Exo84 [6]. Of note, it has been suggested that SEC3 and Exo70 are activated by Rho GTPases and create the initial contact points between vesicle and PM as the major tethers [7,8,9].
Although only one copy of the Exo70 exists in Saccharomyces, Drosophila, and Caenorhabditis, there are multiple copies of Exo70 in plants [10], such as 13 paralogs in moss (Physcomitrella patens), 23 paralogs in Arabidopsis thaliana, 41 paralogs in rice (Oryza sativa), and 23 paralogs in poplar (Populus trichocarpa). These Exo70 members are divided into three clades (Exo70.1 to Exo70.3) and are broadly involved in multiple biological processes in plants [11,12,13]. For instance, Arabidopsis Exo70A1 is required for polar auxin transport in a PIN1/2-dependent pathway. The mutation of Exo70A1 leads to altered auxin distribution, loss of apical dominance, and shorter root hairs [14,15]. Additionally, Exo70A1/2 and Exo70C1/2 function as regulators in sexual reproduction; the exo70a1 mutant exhibits fertility defects, and Exo70A2 mutation results in impaired pollen germination and pollen tube growth [14,16], while Exo70C1/2 are involved in the regulation of optimal tip growth of pollen tubes [17]. Furthermore, the Arabidopsis Exo70H4 is involved in trichome cell wall maturation, and its paralog is also highly expressed in cucumber fruit trichomes, suggesting a conserved function of Exo70H in trichome development [18,19]. The Exo70 members also participate in plant immunity. For instance, the expression of Exo70H1 and Exo70B2 was up-regulated after pathogen incubation in Arabidopsis [20]. Exo70B2 and its homolog Exo70B1 are both required for pattern-triggered immunity (PTI). Accordingly, exo70B1-3 and exo70B2-1 mutants both display enhanced susceptibility to Pseudomonas syringae pv tomato (Pst) DC3000 [21,22,23]. Interestingly, the loss function of Exo70B1 also leads to an activation of TIR-NBS2 (TN2), thus enhancing resistance to multiple pathogens in TN2-dependent autoimmunity [24]. Recent studies showed that Exo70B1/2 regulated FLAGELLIN SENSING 2 (FLS2) accumulation at the PM, which is required for the outcome of immune responses in Arabidopsis [25]. Moreover, OsExo70B1 interacted with Chitin Elicitor Receptor Kinase 1 (CERK1) and was involved in resistance to rice blast fungus Magnaporthe oryzae (M. oryzae) [26], but the mechanism is still largely unknown.
Cucumber (Cucumis sativus L.) is an important vegetable crop of high economic and biological value [27,28]. Along its evolution, the cucumber has diverse sex patterns and has become a model plant for studying plant sex determination [29,30]. Moreover, cucumber usually bears fruits of various sizes, shapes, and colors [31,32,33,34], covered with tubercules, trichomes, and a thick cuticle [35,36]. However, its production is substantially affected by the frequent incidence of multiple diseases. For instance, cucumber bacterial angular leaf spot disease (ALS) is caused by Pseudomonas syringae pv. lachrymans (Psl), which mainly infects the leaves and results in limited vein and necrotic tissues [37,38]. In addition, cucumber fusarium wilt, one of the major fungal diseases, is a soil-borne disease caused by Fusarium oxysporum f. sp. cucumerinum Owen (Foc), which usually leads to a decrease in photosynthesis capacity and loss of yield [39,40]. The Exo70s have been studied in multiple plant species [41,42,43,44], but not cucumber.
At present, genome-wide identification and gene expression analysis are effective approaches to study the classification and potential functions of gene family members, which provides a foundation for further gene function identification [43]. In this study, we aim to identify the CsExo70 members in cucumber. A genome-wide analysis of CsExo70 members was performed, including the system evolution, collinearity, gene structure, cis-acting elements, conserved motifs, subcellular localization, expression patterns in different tissues, and treatment with pathogens. This study will provide a reference for further investigating the functions of the CsExo70 gene family in the future.
2. Results
2.1. Identification and Characterization of CsExo70 Family in Cucumber
In this study, 18 CsExo70 members were identified, which have conserved Exo70 domains (Figure S1). To explore the evolutionary relationships of Exo70s among different species, the full-length sequences of Exo70s of cucumber, Arabidopsis, rice, and moss were used to generate a phylogenetic tree. As shown in Figure 1, all CsExo70 members were divided into three groups (Exo70.1 to Exo70.3) and nine subgroups (Exo70A to Exo70I), and named according to the phylogenetic tree. Except for CsExo70B, CsExo70D, and CsExo70I, other members have several homologs in cucumber.
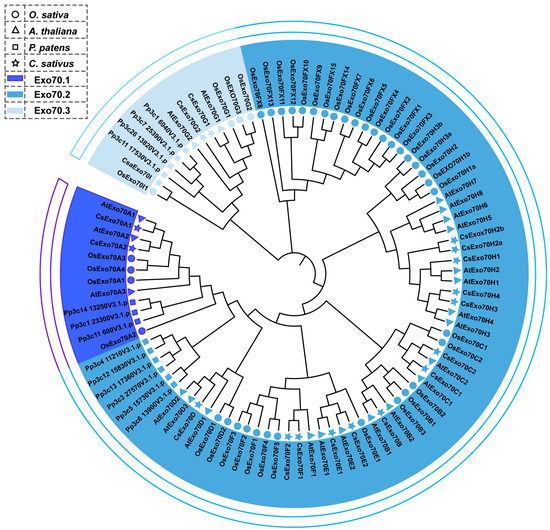
Figure 1.
Phylogenetic analysis of Exo70 family. The circle, triangle, square, and star represent Exo70s in Oryza sativa (O. sativa), Arabidopsis thaliana (A. thaliana), Physcomitrella patens (P. patens), and Cucumis sativus (C. sativus), respectively.
Additionally, the physical and chemical characteristics of CsExo70s were predicted in Table 1, including the length of the coding sequence (CDS) and amino acid (AA) sequence, molecular weight (MW), and isoelectric point (PI) values. In detail, the protein length ranged from 582 (CsExo70H4) to 704 (CsExo70C2) amino acids, MW varied from 66,156.59 Da (CsExo70H4) to 81346.62 Da (CsExo70C2), and the PI value varied from 4.94 (CsExo70E1) to 8.87 (CsExo70G1).

Table 1.
Information of the Exo70 family in cucumber.
2.2. Chromosomal Localization and Collinearity Analysis of CsExo70 Genes in Cucumber
According to the physical location of all CsExo70 genes in the C. sativus genome, the gene chromosomal distribution was drawn. As shown in Figure 2A, 18 CsExo70 genes were distributed over the seven cucumber chromosomes. There were five genes located on chromosome 1, three genes on chromosomes 4 and 5, two genes on chromosomes 2, 3, and 6, and only one gene on chromosome 7. Moreover, some CsExo70 genes had adjacent locations, such as CsExo70E2 and CsExo70F2, CsExo70B and CsExo70H2b, and CsExo70G1 and CsExo70I, but no tandem-duplicated CsExo70 genes were found.
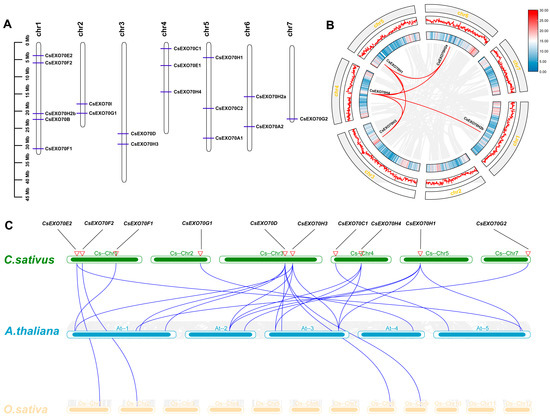
Figure 2.
Chromosomal location and collinearity analysis of Exo70s in cucumber. (A) Locations of CsExo70 genes in seven chromosomes of cucumber. (B) Genome-wide synteny analysis of Exo70s in C. sativus genome. Red lines indicate the paralogous genes. (C) Genome-wide synteny analysis of Exo70s between C. sativus and A. thaliana, and C. sativus and O. sativa genomes. Blue lines represent the orthologous genes, the red triangles represent different gene pairs.
We further explored the segmental duplication events of CsExo70 genes via collinearity analysis in the C. sativus genome. As shown in Figure 2B, six pairs of segmentally duplicated genes were identified in the C. sativus genome: CsExo70H1/CsExo70H4, CsExo70H1/CsExo70H2a, CsExo70H1/CsExo70H3, CsExo70H2a/CsExo70H4, CsExo70H3/CsExo70H4, and CsExo70H4/CsExo70H2b, indicating that CsExo70H members were amplified via segmental duplication events. Furthermore, to better understand the gene amplification pattern during evolution, collinearity analysis among cucumber, Arabidopsis, and rice was performed (Figure 2C). In total, 23 gene pairs were identified. There were 19 pairs between C. sativus and A. thaliana, and 4 pairs between C. sativus and O. sativa, suggesting the Exo70 genes of C. sativus had higher synteny with A. thaliana than O. sativa during the evolution.
2.3. Gene Structure and Conserved Motif Analysis of CsExo70 Genes
The gene structure and sequence characteristics of Exo70 genes were further investigated in cucumber. Gene structure analysis showed that most genes had 5′Untranslated Region (UTR) or 3′UTR. Among them, genes in Exo70.2 and Exo70.3 clades displayed a similar structure with 1–2 exons and 0–2 introns, while the genes in the Exo70.1 clade consisted of 11–12 exons and 11 introns (Figure 3B). These results suggest that genes in the same clade have a similar structure and that functional differentiation may exist between different clades.
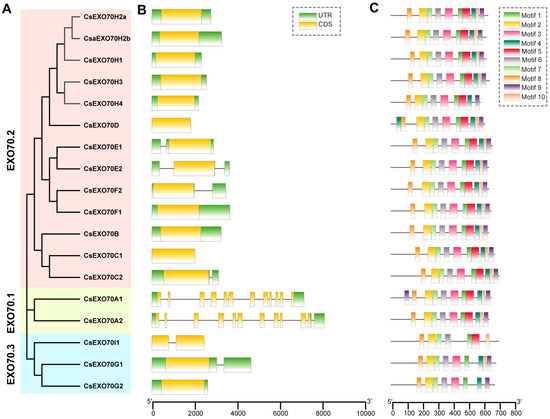
Figure 3.
Gene structure and conserved motifs of Exo70 family in cucumber. (A) The phylogenetic tree of Exo70 members in cucumber. (B) Gene structures of CsExo70s. The yellow and green boxes represented coding sequence (CDS) and untranslated region (UTR), respectively. The black lines indicated introns. (C) Conserved motifs of CsExo70 proteins; the ten motifs are displayed in different colors.
Moreover, the conserved motifs in CsExo70s were analyzed via MEME. A total of 10 conserved motifs were identified in all CsExo70 members. Except for Exo70H4, Exo70D, and Exo70A1, the members in Exo70.1 and Exo70.2 clades had all of these motifs, while Exo70I, Exo70G1, and Exo70G2 in the Exo70.3 clade lacked three, one, and two motifs, respectively. Although the functions of motifs 1–10 have not been revealed, these motifs were mainly located on the C-terminal, which contains the conserved Exo70 domain that determines the function of these proteins. Thus, it speculated that these members of the Exo70.3 clade may have different functions from that in other clades.
2.4. Analysis of Cis-Acting Elements on CsExo70 Promotors
To study the CsExo70 gene expression regulation, the promoter sequences (2000 bp upstream of the start codon) of 18 CsExo70s were analyzed. The major cis-acting elements were identified (Figure 4). The most cis-acting elements were relevant to phytohormones, including related to responses of methyl jasmonate (TGACG motif/CGTCA motif), abscisic acid (ABRE motif), gibberellin (P-box/GARE motif/TATC box), auxin (AuxRE/AuxRR/ TGA motif), and salicylic acid (TCA motif). Among these, the abscisic acid-response element (ABRE) could be found on all CsExo70 promoters, except CsExo70G1. Moreover, these CsExo70 promoters contain stress-responsive elements, including anaerobic-induction elements (ARE), defense-responsive elements (TC-rich repeats), low-temperature responsive (LTR) elements, drought-induction elements (MBS), and wound-responsive elements (WUN motif). Cis-acting elements relevant to growth and development were also found, including circadian control, meristem expression (CAT box), endosperm-specific expression (AACA motif/GCN4 motif), zein metabolism regulation (O2 site), mesophyll cell differentiation (HD-Zip), and cell cycle regulation (MSA-like). These results suggest that CsExo70 genes may be involved in multiple biological processes.
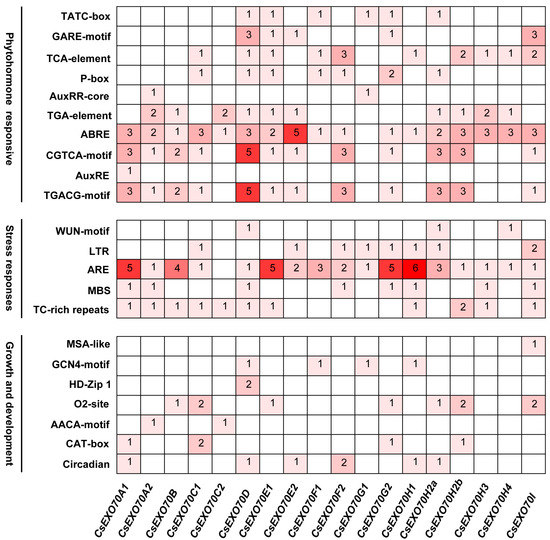
Figure 4.
Cis-acting elements in the promotors of Exo70 genes in cucumber. The colors indicate the different cis-elements numbers. Values indicate the statistical number of cis-elements.
2.5. Expression Patterns of CsExo70 Genes in Cucumber
To investigate tissue-specific expression, the expression patterns of all 18 CsExo70 genes were examined by quantitative RT-PCR (qRT-PCR) in different cucumber tissues during the reproductive growth stage, including tendril, root, stem, leaf, male-flower petal, stamen, female-flower petal, stigma, and ovary (Figure 5). Among 18 CsExo70s, seven genes (CsExo70A1, CsExo70A2, CsExo70G2, CsExo70H1, CsExo70H2a, CsExo70H2b, and CsExo70H4) have lower transcript levels and were almost not detected in all tissues. Alternatively, these genes were expressed in other development stages. Interestingly, most of the remaining 11 genes displayed a similar tissue expression pattern with the highest transcripts in the stamen, especially for CsExo70C1 and CsExo70C2 (Figure 5B,C), consistent with Exo70C homologous in Arabidopsis, rice, and cotton [13,17,41], indicating that Exo70C members may have a conserved function in the regulation of stamen fertility. Moreover, CsExo70F1, CsExo70G1, and CsExo70H3 were highly expressed in stigma (Figure 5G,I,J), the transcript of CsExo70E2 was higher in root and stem, and CsExo70H3 was highly expressed in the leaf (Figure 5F,J). These results suggest that CsExo70 members may regulate the growth and development of multiple tissues.
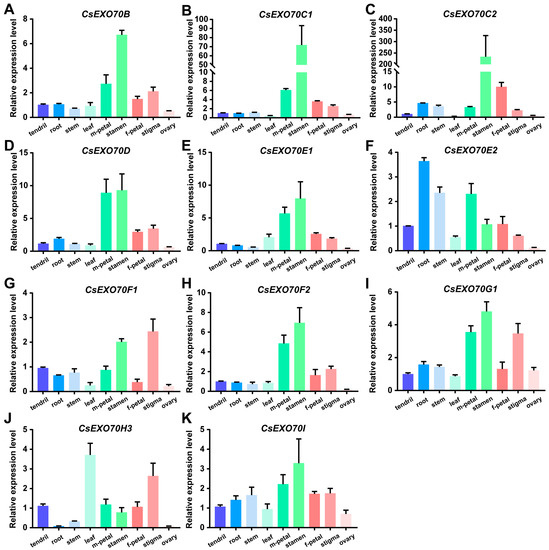
Figure 5.
Expression patterns of CsExo70B (A), CsExo70C1 (B), CsExo70C2 (C), CsExo70D (D), CsExo70E1 (E), CsExo70E2 (F), CsExo70F1 (G), CsExo70F2 (H), CsExo70G1 (I), CsExo70H3 (J), and CsExo70I (K) in cucumber tissues. The cucumber UBQ gene was used as an internal standard. m, male; f, female.
2.6. CsExo70 Gene Expression in Response to Different Pathogens
Previous studies have demonstrated that the transcripts of Exo70B2 and Exo70H1 were induced by bacterial elongation factor-TU epitope elf18 in Arabidopsis [20]. In rice, the transcript of OsExo70B1 was increased after the pathogen-associated molecular pattern (PAMP) and M. oryzae treatment [26]. To investigate pathogen-triggered CsExo70 expression in cucumber, the expression levels of these genes at different time points after Psl treatment were detected. We observed that the expression of CsExo70E2 was about 40-fold higher at 2 days post inoculation (dpi) than that at 0 dpi (Figure 6F). The expression of CsExo70H3 was also significantly increased to 7–8-fold at 2–3 dpi (Figure 6J), while the expression of CsExo70B, CsExo70C2, and CsExo70F2 was slightly up-regulated at 2 dpi (Figure 6A,C,H). In contrast, CsExo70C1, CsExo70G1, and CsExo70I were down-regulated after Psl infection (Figure 6B,I,K), and the expression levels of CsExo70D, CsExo70E1, and CsExo70F1 were unaffected after Psl treatment (Figure 6D,E,G). These results indicate that some CsExo70 genes may play essential roles in resistance to Psl in cucumber.
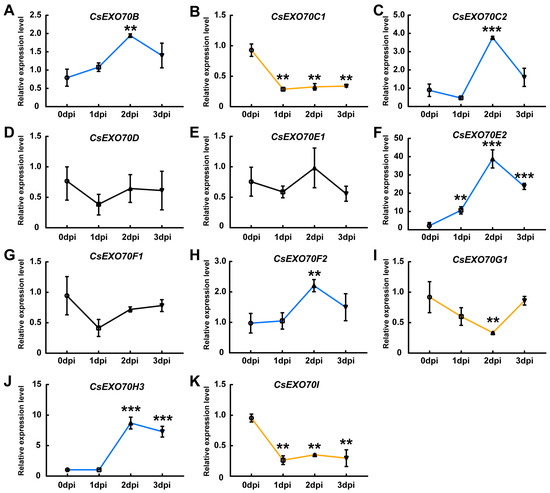
Figure 6.
Pseudomonas syringae pv. lachrymans (Psl)-triggered CsExo70 expression in cucumber. (A–K) The transcript levels of CsExo70B (A), CsExo70C1 (B), CsExo70C2 (C), CsExo70D (D), CsExo70E1 (E), CsExo70E2 (F), CsExo70F1 (G), CsExo70F2 (H), CsExo70G1 (I), CsExo70H3 (J), and CsExo70I (K) were detected after Psl infection. Leaves of four-week-old cucumber seedlings were treated with Psl, and harvested at 0, 1, 2, 3 days after inoculation (dpi) for expression analysis. CsExo70 transcripts were quantified by qRT-PCR using UBQ as the internal standard. Significant differences are indicated by asterisks (** p < 0.01, *** p < 0.001, Student’s t test). Blue and yellow lines represent increased or decreased genes, respectively; the black line indicates genes without a significant change.
To verify whether CsExo70 genes are also responsive to fungal pathogen, Foc-triggered CsExo70 expression was examined. Similar to Psl treatment, the expressions of CsExo70E2 and CsExo70H3 were also significantly increased in response to Foc (Figure 7F,J). The same results were also observed for CsExo70B, CsExo70C2, and CsExo70F2 expression levels at 2 dpi or 4 dpi after Foc treatment (Figure 7A,C,H). Unlike that after Psl inoculation, the expression of CsExo70C1 and CsExo70I was up-regulated at 6 and 2 dpi, respectively (Figure 7B,K), while the expression of CsExo70G1 was unchanged in response to Foc (Figure 7I). Furthermore, the expression levels of CsExo70D and CsExo70F1 were instantaneously increased at 2 dpi, then decreased to normal levels at 4 dpi, and increased at 6 dpi (Figure 7D,G). CsExo70E1 was weakly up-regulated after Foc infection, and CsExo70G1 expression was unchanged by Foc treatment (Figure 7E,I). These results indicate that the expression of most CsExo70 genes was induced by pathogen treatment, especially CsExo70E2 and CsExo70H3, which may play important roles in plant defense against both fungal and bacterial diseases in cucumber.
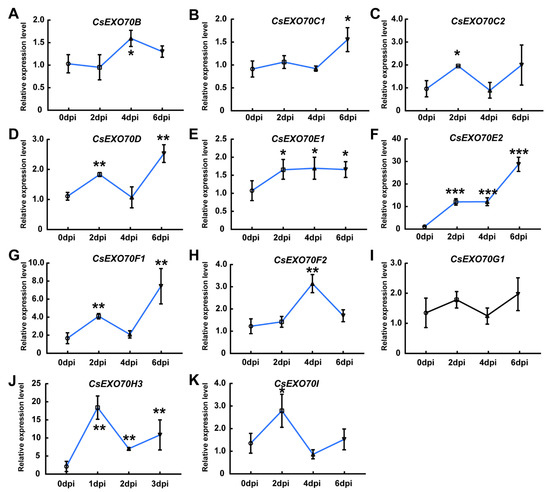
Figure 7.
Expression analysis of CsExo70B (A), CsExo70C1 (B), CsExo70C2 (C), CsExo70D (D), CsExo70E1 (E), CsExo70E2 (F), CsExo70F1 (G), CsExo70F2 (H), CsExo70G1 (I), CsExo70H3 (J), and CsExo70I (K) after Fusarium oxysporum f. sp. cucumerinum Owen (Foc) treatment. One-week-old cucumber seedlings were treated with Foc and roots were collected at 0, 2, 4, and 6 days after inoculation (dpi). Values are means ± sd of three biological replicates. Significant differences between 0 dpi and other time points are indicated by asterisks (* p < 0.05, ** p < 0.01, *** p < 0.001, Student’s t test). The blue lines represent up-regulated genes, and the black line indicates genes without a significant change at any time point.
2.7. Subcellular Localization Analysis
To examine the subcellular location of CsExo70 members, we randomly selected one member from each clade for analysis. CsExo70A1, CsExo70B, and CsExo70G1 were fused with the GFP (green fluorescent protein) and transiently expressed in Nicotiana benthamiana (N. benthamiana) leaves. CsExo70A1 and CsExo70G1 were mainly localized in the cytoplasm and nucleus, and CsExo70B was widely distributed in PM, cytoplasm, and nucleus. Furthermore, the fluorescence signals in the cytoplasm of CsExo70A1, CsExo70B, and CsExo70G1 were discrete and punctate (Figure 8), consistent with the localization of Exo70B1 and Exo70H3 in Arabidopsis and rice [26,45,46,47], indicating a similar localization of Exo70s in different species.
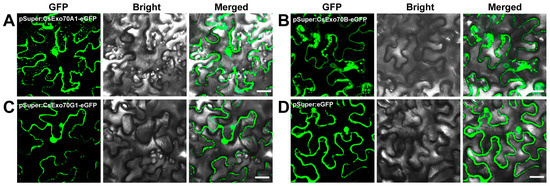
Figure 8.
Subcellular localizations of CsExo70A1 (A), CsExo70B (B), and CsExo70G1 (C) in Nicotiana benthamiana (N. benthamiana). (D) The empty vector was used as a control. These indicated structures were transiently expressed in N. benthamiana leaves. Bar, 40 μm.
3. Discussion
Previous studies have shown that the Exo70 subunit has expanded to a larger family in plants compared to that in yeast and animals [13,14,41], indicating the amplification of the Exo70 gene in plants. In this study, 18 cucumber Exo70 members were identified by sequence BLASTp search and phylogenetic analysis (Figure 1), and divided into three clades (Exo70.1 to Exo70.3) and nine subclades (Exo70A to Exo70I) based on the different gene structures (Figure 3), which was consistent with previous studies in other species [13,42,43,44], suggesting the conservation and similarity of Exo70 gene family in the evolution among different species. Moreover, previous studies have shown that the Exo70I subclade is commonly found in monocotyledonous plants (rice and wheat) but not dicotyledonous plants (Arabidopsis, cotton, and grape) [13,42,43,44]. However, CsExo70I was also identified in this study, more similar to rice but not Arabidopsis, suggesting that the Exo70I branch also exists in dicotyledonous plants.
Tandem and segmental duplication are the main types of gene replication events [48]. Through collinear analysis, we found six segmental duplication events in the CsExo70H subclade (Figure 2B), which contained the largest numbers among the CsExo70 subclades, meaning segmental duplication was the dominant driver of duplication of CsExo70H genes. On the other hand, CsExo70 genes in Exo70.1 clade contained 11–12 exons, while only one or two exons were found in Exo70.2 and Exo70.3 clade genes (Figure 3B). Moreover, it is noteworthy to observe that all ten conserved motifs were generally distributed members in Exo70.1 and Exo70.2 clades, but not in Exo70.3 clade (Figure 3C). As mentioned above, the differences in gene structures and conserved motifs may result in function differentiation.
Moreover, the cis-acting elements on the CsExo70 promoters are related to phytohormones, stress, and growth and development (Figure 4). Consistent with a previous report that Exo70A1 is involved in auxin transport and PIN recycling in Arabidopsis [15], more auxin-related elements (AuxRE, AuxRR, TGA elements) were observed on the promoters of CsExo70A1 and CsExo70A2. In addition, when plants are subjected to stresses, some transcription factors will bind the cis-elements to promote the expression of related genes [49,50,51]. Notably, methyl jasmonate (MeJA) is one of the vital regulatory factors in plant resistance signal transduction pathways, and the elements of MeJA-responsive (CGTCA-motif and TGACG-motif) were mainly presented in CsExo70D promotor; abscisic acid (ABA) is broadly involved in drought stress and salt stress [52,53]. ABRE as the major element regulated ABA-responsive gene expression [53], was distributed on almost every promoter of CsExo70s, with a maximum of CsExo70E2, suggesting the significant roles of CsExo70D and CsExo70E2 in MeJA and ABA signaling pathways, respectively. Furthermore, a total of 43 AREs were found on the promotors of CsExo70s, implying the potential function of CsExo70s in oxidative responsive, especially for CsExo70A1, CsExo70B, CsExo70E1, CsExo70G2, and CsExo70H2. It is speculated that CsExo70s may play essential roles in plant development and stress responses.
A previous study has reported that Exo70C1/2 is involved in the regulation of optimal tip growth of pollen tubes [17]; the expression pattern analysis in cucumber showed that CsExo70C1 and CsExo70C2 were highly expressed in the stamen, which further provided support for the involvement of Exo70C1/2 in sexual reproduction [13,17,41,54], suggesting the conservative function of Exo70 members in different plant species. On the other hand, pathogens affect cucumber growth and development and cause a variety of diseases, resulting in a decrease in cucumber yield [55]. qRT-PCR results showed that most CsExo70 genes were induced by pathogens (Figure 6 and Figure 7). Consistent with the exo70H1 mutant being susceptible to Pseudomonas syringae pv. maculicola (Psm) [20], Psl-induced expression of CsExo70H3 was about 10-fold higher at 2 dpi than that at 0 dpi, indicating that CsExo70H3 may be also involved in Psl resistance in cucumber. Cucumber fusarium wilt is a soil-borne vascular disease, which usually infects cucumber roots [56]. Our data showed that the expression of CsExo70E2 was about 30-fold higher at 6 dpi than 0 dpi after Foc treatment. These results suggested that CsExo70E2 may play an essential role in resistance to Foc in cucumber.
The feature of subcellular compartments is related to their specialized biological functions [57]. In this study, CsExo70A1, CsExo70B, and CsExo70G1 all showed fluorescent signals in the cytoplasm and nucleus, and the signals were discrete and punctate, similar to previous studies [26,45,46,47]. Moreover, previous studies showed that Exo70B interacts with PM-localization receptor kinases (RKs) to regulate plant immunity [25,26]. This can explain why CsExo70B also displayed the PM signals. Given that the transcripts of CsExo70B are also induced by Psl and Foc infection, it is worth exploring whether Exo70B interacts with immune-related RKs to regulate pathogen resistance in cucumber.
4. Material and Methods
4.1. Plant Materials
Cucumber (Cucumis sativus L.) inbred line XTMC was used in this study. The cucumber seedlings at the two-true leaf stage were transplanted to a greenhouse under standard management at the China Agricultural University, Beijing. The N. bethamiana plants used for CsExo70 subcellular localization analysis were grown in a chamber at 24 °C with 16 h light/8 h dark.
4.2. Identification and Phylogenetic Tree Construction of Exo70 Family
To identify the predicted Exo70 genes in the C. sativus genome, the amino acid sequences of 23 AtExo70 members were downloaded from the TAIR database (https://www.arabidopsis.org, accessed on 3 March 2023), and used as queries to BLASTp against the cucumber version 3 genome database (http://www.cucurbitgenomics.org/organism/20, accessed on 4 March 2023). All predicted CsExo70s were further verified by the Exo70 conserved domain by SMART (http://smart.emblheidelberg.de, accessed on 4 March 2023) and CDD-research (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi, accessed on 4 March 2023). The characteristics of CsExo70s, including the protein molecular weight (MW) and isoelectric point (pI), were analyzed via ExPaSy (http://web.expasy.org/protparam/, accessed on 5 March 2023).
The Exo70 protein sequences of O. sativa and P. patens were obtained from Phytozome13 website (https://phytozome-next.jgi.doe.gov/, accessed on 6 March 2023). Multiple sequence alignment analysis of Exo70s in cucumber, Arabidopsis, rice, and moss were performed in MEGA V7.0 software according to the Clustalw algorithm. Subsequently, the alignment was used to generate a phylogenetic tree using the neighbor-joining (NJ) method (bootstrap = 1000 repetitions).
4.3. Chromosomal Location and Synteny Analysis
The chromosomal locations of CsExo70 genes were obtained through the Cucurbit Genomics Database. The CsExo70 genes were mapped on different chromosomes and ultimately plotted using the TBtools [58] according to their physical positions. The syntenic maps of Exo70 genes were generated and analyzed by TBtools [58] and displayed by Advanced Circos and Multiple Synteny Plot.
4.4. Gene Structures, Conserved Motifs, and Cis-Element Analysis
Gene structure analysis was performed by TBtools [58], and the conserved motifs of CsExo70 proteins were analyzed using Multiple Expectation Maximization for Motif Elicitation (MEME) (https://meme-suite.org/meme/tools/meme, accessed on 12 March 2023). The promoter sequences (2000 bp upstream of the start codon) were extracted using NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 18 March 2023), and the transcriptional response cis-elements in promoters were predicted using the PlantCARE database (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 20 March 2023) and drawn by TBtools [58].
4.5. RNA Extraction and Gene Expression Analysis
To determine the expression patterns of these CsExo70 genes in different tissues, samples of roots, stems, leaves, male flowers, female flowers, stigmas, stamens, ovaries, and tendrils were collected during the reproductive growth stage for RNA extraction.
The total RNA was extracted using an Eastep® Super Total RNA Extraction Kit (Promega, Madison, WI, USA) according to the manufacturer’s instructions, and then reverse-transcribed to complementary DNA (cDNA) using a FastKing gDNA Dispelling RT SuperMix Kit (Tiangen, Beijing, China). Subsequently, qRT-PCR was performed with Taq Pro Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China) in a CFX384 Real-Time PCR System (BIO-RAD, Hercules, CA, USA). Three biological replicates and three technical replicates were performed for every CsExo70 gene and UBQ gene (CsaV3_5G031430, an internal standard). The data were analyzed using the 2−∆∆Ct method [59]. All primers used for qRT-PCR are listed in Table S1.
4.6. Plant Pathogen Treatment
For analysis of Psl-inducible gene expression, a pathogen infection assay was performed as described previously in cucumber [60]. Briefly, the second true leaves of four-week-old cucumber seedlings were sprayed with 106 cfu/mL of Psl, samples were collected at pre-inoculation (0 day) as control, and 1, 2, 3 dpi for expression analysis.
For analysis of Foc-inducible gene expression, the roots of one-week-old cucumber seedlings were dipped in Foc spore suspension (1 × 106 spores/mL) as described previously [56]. Similarly, root samples were harvested on 0 (as control), and 2, 4, and 6 dpi for further expression analysis.
4.7. Subcellular Localization Analysis
The full-length coding sequences without stop codon of CsExo70A1, CsExo70B, and CsExo70G1 were amplified and cloned into pSuper-1300-eGFP vector. Then the resultant vectors were transformed into agrobacterium strain GV3101 and infiltrated into N. benthamiana leaves as previously described [61]. After 72 h infiltration, the fluorescence signals of GFP were observed using a confocal microscope (Zeiss LSM880, Jena, Germany) at an excitation wavelength of 488 nm and an emission wavelength of 510 nm. All primers used for subcellular localization analysis are listed in Table S1.
5. Conclusions
A total of 18 CsExo70 members were identified in this study. Tissue expression analyses showed that most CsExo70 genes (except for CsExo70E2 and CsExo70H3) exhibited a similar expression pattern, and CsExo70C1 and CsExo70C2 were highly expressed in the stamen. In addition, the expression levels of most CsExo70 genes were induced by pathogens, especially CsExo70E2 and CsExo70H3. Together, our results provide the basis for studying the functions of Exo70 members in cucumber.
Supplementary Materials
The supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms241310929/s1.
Author Contributions
X.Z., Z.Z. and L.L. designed experiments. L.L performed the major experiments, analyzed the data, and wrote this manuscript. C.G. performed the qRT-PCR assays and checked this manuscript. J.Z. and J.G. constructed the vectors for subcellular localization and provided experimental assistances. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32025033, 31930097); the 2115 Talent Development Program of China Agricultural University; Sanya Institute of China Agricultural University (SYND-2022-30).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within this article and Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Saeed, B.; Brillada, C.; Trujillo, M. Dissecting the plant exocyst. Curr. Opin. Plant Biol. 2019, 52, 69–76. [Google Scholar] [CrossRef]
- Stegmann, M.; Monaghan, J.; Smakowska-Luzan, E.; Rovenich, H.; Lehner, A.; Holton, N.; Belkhadir, Y.; Zipfel, C. The receptor kinase FER is a RALF-regulated scaffold controlling plant immune signaling. Science 2017, 355, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.-M.; Palmquist, J.; Huang, S.-D.; Jin, H. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef] [PubMed]
- Baldrich, P.; Rutter, B.D.; Karimi, H.Z.; Podicheti, R.; Meyers, B.C.; Innes, R.W. Plant extracellular vesicles contain diverse small RNA species and are enriched in 10- to 17-nucleotide “Tiny” RNAs. Plant Cell 2019, 31, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Guo, W. The Exocyst at a Glance. J. Cell Sci. 2015, 128, 2957–2964. [Google Scholar] [CrossRef]
- TerBush, D.R.; Maurice, T.; Roth, D.; Novick, P. The Exocyst is a multiprotein complex required for exocytosis in Saccharomyces cerevisiae. EMBO J. 1996, 15, 6483–6494. [Google Scholar] [CrossRef]
- Pleskot, R.; Cwiklik, L.; Jungwirth, P.; Žárský, V.; Potocký, M. Membrane targeting of the yeast exocyst complex. Biochim. Biophys. Acta 2015, 1848, 1481–1489. [Google Scholar] [CrossRef]
- He, B.; Xi, F.; Zhang, X.; Zhang, J.; Guo, W. Exo70 interacts with phospholipids and mediates the targeting of the exocyst to the plasma membrane. EMBO J. 2007, 26, 4053–4065. [Google Scholar] [CrossRef]
- Boyd, C.; Hughes, T.; Pypaert, M.; Novick, P. Vesicles carry most exocyst subunits to exocytic sites marked by the remaining two subunits, Sec3p and Exo70p. J. Cell Biol. 2004, 167, 889–901. [Google Scholar] [CrossRef]
- Elias, M.; Drdova, E.; Ziak, D.; Bavlnka, B.; Hala, M.; Cvrckova, F.; Soukupova, H.; Zarsky, V. The exocyst complex in plants. Cell Biol. Int. 2003, 27, 199–201. [Google Scholar] [CrossRef]
- Žárský, V.; Sekereš, J.; Kubátová, Z.; Pečenková, T.; Cvrčková, F. Three subfamilies of exocyst EXO70 family subunits in land plants: Early divergence and ongoing functional specialization. J. Exp. Bot. 2020, 71, 49–62. [Google Scholar] [CrossRef]
- Cvrčková, F.; Grunt, M.; Bezvoda, R.; Hála, M.; Kulich, I.; Rawat, A.; Žárský, V. Evolution of the land plant exocyst complexes. Front. Plant Sci. 2012, 3, 159. [Google Scholar] [CrossRef]
- Chong, Y.T.; Gidda, S.K.; Sanford, C.; Parkinson, J.; Mullen, R.T.; Goring, D.R. Characterization of the Arabidopsis thaliana exocyst complex gene families by phylogenetic, expression profiling, and subcellular localization studies. New Phytol. 2010, 185, 401–419. [Google Scholar] [CrossRef]
- Synek, L.; Schlager, N.; Eliás, M.; Quentin, M.; Hauser, M.-T.; Žárský, V. AtEXO70A1, a member of a family of putative exocyst subunits specifically expanded in land plants, is important for polar growth and plant development. Plant J. 2006, 48, 54–72. [Google Scholar] [CrossRef] [PubMed]
- Drdová, E.J.; Synek, L.; Pečenková, T.; Hála, M.; Kulich, I.; Fowler, J.E.; Murphy, A.S.; Žárský, V. The exocyst complex contributes to PIN auxin efflux carrier recycling and polar auxin transport in Arabidopsis. Plant J. 2013, 73, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Marković, V.; Cvrčková, F.; Potocký, M.; Kulich, I.; Pejchar, P.; Kollárová, E.; Synek, L.; Žárský, V. EXO70A2 is critical for exocyst complex function in pollen development. Plant Physiol. 2020, 184, 1823–1839. [Google Scholar] [CrossRef] [PubMed]
- Synek, L.; Vukašinović, N.; Kulich, I.; Hála, M.; Aldorfová, K.; Fendrych, M.; Žárský, V. EXO70C2 is a key regulatory factor for optimal tip growth of pollen. Plant Physiol. 2017, 174, 223–240. [Google Scholar] [CrossRef]
- Kulich, I.; Vojtíková, Z.; Glanc, M.; Ortmannová, J.; Rasmann, S.; Žárský, V. Cell Wall Maturation of Arabidopsis trichomes is dependent on exocyst subunit EXO70H4 and involves callose deposition. Plant Physiol. 2015, 168, 120–131. [Google Scholar] [CrossRef]
- Chen, C.; Liu, M.; Jiang, L.; Liu, X.; Zhao, J.; Yan, S.; Yang, S.; Ren, H.; Liu, R.; Zhang, X. Transcriptome profiling reveals roles of meristem regulators and polarity genes during fruit trichome development in cucumber (Cucumis sativus L.). J. Exp. Bot. 2014, 65, 4943–4958. [Google Scholar] [CrossRef]
- Pecenkova, T.; Hala, M.; Kulich, I.; Kocourkova, D.; Drdova, E.; Fendrych, M.; Toupalova, H.; Zarsky, V. The role for the exocyst complex subunits Exo70B2 and Exo70H1 in the plant-pathogen interaction. J. Exp. Bot. 2011, 62, 2107–2116. [Google Scholar] [CrossRef]
- Wang, W.; Liu, N.; Gao, C.; Rui, L.; Tang, D. The Pseudomonas syringae Effector AvrPtoB associates with and ubiquitinates Arabidopsis exocyst subunit EXO70B1. Front. Plant Sci. 2019, 10, 1027. [Google Scholar] [CrossRef]
- Stegmann, M.; Anderson, R.G.; Westphal, L.; Rosahl, S.; McDowell, J.M.; Trujillo, M. The exocyst subunit Exo70B1 is involved in the immune response of Arabidopsis thaliana to different pathogens and cell death. Plant Signal. Behav. 2013, 8, e27421. [Google Scholar] [CrossRef] [PubMed]
- Stegmann, M.; Anderson, R.G.; Ichimura, K.; Pecenkova, T.; Reuter, P.; Žársky, V.; McDowell, J.M.; Shirasu, K.; Trujillo, M. The ubiquitin ligase PUB22 targets a subunit of the exocyst complex required for PAMP-triggered responses in Arabidopsis. Plant Cell 2012, 24, 4703–4716. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Rui, L.; Li, J.; Nishimura, M.T.; Vogel, J.P.; Liu, N.; Liu, S.; Zhao, Y.; Dangl, J.L.; Tang, D. A truncated NLR protein, TIR-NBS2, is required for activated defense responses in the exo70B1 Mutant. PLoS Genet. 2015, 11, e1004945. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, N.; Gao, C.; Cai, H.; Romeis, T.; Tang, D. The Arabidopsis exocyst subunits EXO70B1 and EXO70B2 regulate FLS2 homeostasis at the plasma membrane. New Phytol. 2020, 227, 529–544. [Google Scholar] [CrossRef]
- Hou, H.; Fang, J.; Liang, J.; Diao, Z.; Wang, W.; Yang, D.; Li, S.; Tang, D. OsExo70B1 positively regulates disease resistance to Magnaporthe oryzae in rice. Int. J. Mol. Sci. 2020, 21, 7049. [Google Scholar] [CrossRef]
- Sharif, R.; Su, L.; Chen, X.; Qi, X. Involvement of auxin in growth and stress response of cucumber. Veg. Res. 2022, 2, 13. [Google Scholar] [CrossRef]
- Robinson, R.W.; Decker-Walters, D.S. Cucurbits; CAB International: Oxford, UK; New York, NY, USA, 1997. [Google Scholar]
- Rodriguez-Granados, N.Y.; Lemhemdi, A.; Choucha, F.A.; Latrasse, D.; Benhamed, M.; Boualem, A.; Bendahmane, A. Sex determination in cucumis. In Genetics and Genomics of Cucurbitaceae; Springer International Publishing: Cham, Switzerland, 2017; pp. 307–319. [Google Scholar]
- Foucart, C.; Boualem, A.; Lasseur, B.; Eleblu, J.; Fahraj, I.; Bendahmane, A. Sex determination in cucurbits. Biol. Aujourd'hui 2012, 206, 57–62. [Google Scholar] [CrossRef]
- Yang, L.; Koo, D.-H.; Li, Y.; Zhang, X.; Luan, F.; Havey, M.J.; Jiang, J.; Weng, Y. Chromosome rearrangements during domestication of cucumber as revealed by high-density genetic mapping and draft genome assembly. Plant J. 2012, 71, 895–906. [Google Scholar] [CrossRef]
- Sebastian, P.; Schaefer, H.; Telford, I.R.H.; Renner, S.S. Cucumber (Cucumis sativus) and melon (C. melo) have numerous wild relatives in Asia and Australia, and the sister species of melon is from Australia. Proc. Natl. Acad. Sci. USA 2010, 107, 14269–14273. [Google Scholar] [CrossRef]
- Naegele, R.P.; Wehner, T.C. Genetic resources of cucumber. In Genetics and Genomics of Cucurbitaceae; Springer International Publishing: Cham, Switzerland, 2017; pp. 61–86. [Google Scholar]
- Cui, Y.; Li, S.; Dong, Y.; Wu, H.; Gao, Y.; Feng, Z.; Zhao, X.; Shan, L.; Zhang, Z.; Liu, Z.; et al. Genetic regulation and molecular mechanism of immature cucumber peel color: A review. Veg. Res. 2023, 3, 9. [Google Scholar] [CrossRef]
- Miao, H.; Zhang, S.; Wang, X.; Zhang, Z.; Li, M.; Mu, S.; Cheng, Z.; Zhang, R.; Huang, S.; Xie, B.; et al. A linkage map of cultivated cucumber (Cucumis sativus L.) with 248 microsatellite marker loci and seven genes for horticulturally important traits. Euphytica 2011, 182, 167–176. [Google Scholar] [CrossRef]
- Grumet, R.; Lin, Y.C.; Rett-Cadman, S.; Malik, A. Morphological and genetic diversity of Cucumber (Cucumis sativus L.) fruit development. Plants 2022, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- Slomnicka, R.; Olczak-Woltman, H.; Sobczak, M.; Bartoszewski, G. Transcriptome profiling of Cucumber (Cucumis sativus L.) early response to Pseudomonas syringae pv. lachrymans. Int. J. Mol. Sci. 2021, 22, 4192. [Google Scholar] [CrossRef]
- Bhat, N.A.; Bhat, K.A.; Zargar, M.Y.; Teli, M.A.; Nazir, M.; Zargar, S.M. Current status of angular leaf spot (Pseudomonas syringae pv. lachrymans) of cucumber: A review. Int. J. Curr. Res. 2010, 8, 1–11. [Google Scholar]
- Raza, W.; Ling, N.; Zhang, R.; Huang, Q.; Xu, Y.; Shen, Q. Success evaluation of the biological control of Fusarium wilts of cucumber, banana, and tomato since 2000 and future research strategies. Crit. Rev. Biotechnol. 2017, 37, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Mao, Q.; Yan, Y.; Wu, M.; Wang, Y.; Ren, J.; Guo, P.; Liu, A.; Chen, S. Role of melatonin in arbuscular mycorrhizal fungi-induced resistance to Fusarium wilt in cucumber. Phytopathology 2020, 110, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-Q.; Qiu, L.; Liu, L.-L.; Luo, L.; Han, X.-P.; Zhai, Y.-H.; Wang, W.-J.; Ren, M.-Z.; Xing, Y.-D. Identification and comprehensive structural and functional analyses of the EXO70 gene family in cotton. Genes 2021, 12, 1594. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Wan, W.; Zhang, H.; Liu, J.; Li, M.; Wang, H.; Xiao, J.; Wang, X. Identification and characterization of the EXO70 gene family in polyploid wheat and related species. Int. J. Mol. Sci. 2018, 20, 60. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ma, Z.H.; Mao, J.; Chen, B.H. Genome-wide identification and expression analysis of the EXO70 gene family in grape (Vitis vinifera L). PeerJ 2021, 9, e11176. [Google Scholar] [CrossRef]
- Marković, V.; Kulich, I.; Žárský, V. Functional specialization within the EXO70 gene family in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 7595. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Guo, J.; Zhang, Q.; Shi, S.; Guan, W.; Zhou, C.; Chen, R.; Du, B.; Zhu, L.; He, G. Necessity of rice resistance to planthoppers for OsEXO70H3 regulating SAMSL excretion and lignin deposition in cell walls. New Phytol. 2022, 234, 1031–1046. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.H.; Ahn, M.Y.; Park, K.Y.; Kim, E.Y.; Kim, W.T. The N-Terminal UND motif of the Arabidopsis U-Box E3 Ligase PUB18 is critical for the negative regulation of ABA-mediated stomatal movement and determines its ubiquitination specificity for exocyst subunit Exo70B1. Plant Cell 2016, 28, 2952–2973. [Google Scholar] [CrossRef] [PubMed]
- Kulich, I.; Pecenkova, T.; Sekeres, J.; Smetana, O.; Fendrych, M.; Foissner, I.; Hoeftberger, M.; Zarsky, V. Arabidopsis exocyst subcomplex containing subunit EXO70B1 is involved in autophagy-related transport to the vacuole. Traffic 2013, 14, 1155–1165. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Teixeira, F.K.; Menezes-Benavente, L.; Margis, R.; Margis-Pinheiro, M. Analysis of the molecular evolutionary history of the ascorbate peroxidase gene family: Inferences from the rice genome. J. Mol. Evol. 2004, 59, 761–770. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene expression and signal transduction in water-stress response. Plant Physiol. 1997, 115, 327–334. [Google Scholar] [CrossRef]
- Hadiarto, T.; Tran, L.S. Progress studies of drought-responsive genes in rice. Plant Cell Rep. 2011, 30, 297–310. [Google Scholar] [CrossRef]
- Pan, W.; Zheng, P.; Zhang, C.; Wang, W.; Li, Y.; Fan, T.; Liu, Y.; Cao, S. The effect of ABRE BINDING FACTOR 4-mediated FYVE1 on salt stress tolerance in Arabidopsis. Plant Sci. 2020, 296, 110489. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Rep. 2013, 32, 959–970. [Google Scholar] [CrossRef]
- Grobei, M.A.; Qeli, E.; Brunner, E.; Rehrauer, H.; Zhang, R.; Roschitzki, B.; Basler, K.; Ahrens, C.H.; Grossniklaus, U. Deterministic protein inference for shotgun proteomics data provides new insights into Arabidopsis pollen development and function. Genome Res. 2009, 19, 1786–1800. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Wehner, T.C. Cucumber Gene Catalog 2017. In Cucurbit Genetic Cooperative; USDA, U.S. Vegetable Laboratory: Charleston, SC, USA, 2017; pp. 17–29. [Google Scholar]
- Bartholomew, E.S.; Xu, S.; Zhang, Y.; Yin, S.; Feng, Z.; Chen, S.; Sun, L.; Yang, S.; Wang, Y.; Liu, P.; et al. A chitinase CsChi23 promoter polymorphism underlies cucumber resistance against Fusarium oxysporum f. sp. cucumerinum. New Phytol. 2022, 236, 1471–1486. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Catanzariti, A.M.; DeBoer, K.; Petre, B.; Gardiner, D.M.; Singh, K.B.; Dodds, P.N.; Taylor, J.M. LOCALIZER: Subcellular localization prediction of both plant and effector proteins in the plant cell. Sci. Rep. 2017, 7, 44598. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-Time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, J.; Li, W.; Xiang, T.; Liu, Z.; Laluk, K.; Ding, X.; Zou, Y.; Gao, M.; Zhang, X.; Chen, S.; et al. Receptor-like cytoplasmic kinases integrate signaling from multiple plant immune receptors and are targeted by a Pseudomonas syringae effector. Cell Host Microbe 2010, 7, 290–301. [Google Scholar] [CrossRef]
- Kokkirala, V.R.; Yonggang, P.; Abbagani, S.; Zhu, Z.; Umate, P. Subcellular localization of proteins of Oryza sativa L. in the model tobacco and tomato plants. Plant Signal. Behav. 2010, 5, 1336–1341. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).