
Mesenchymal Stem Cells and Exosomes: A Novel Therapeutic Approach for Corneal Diseases

 , ,
, , 
Abstract
1. Introduction
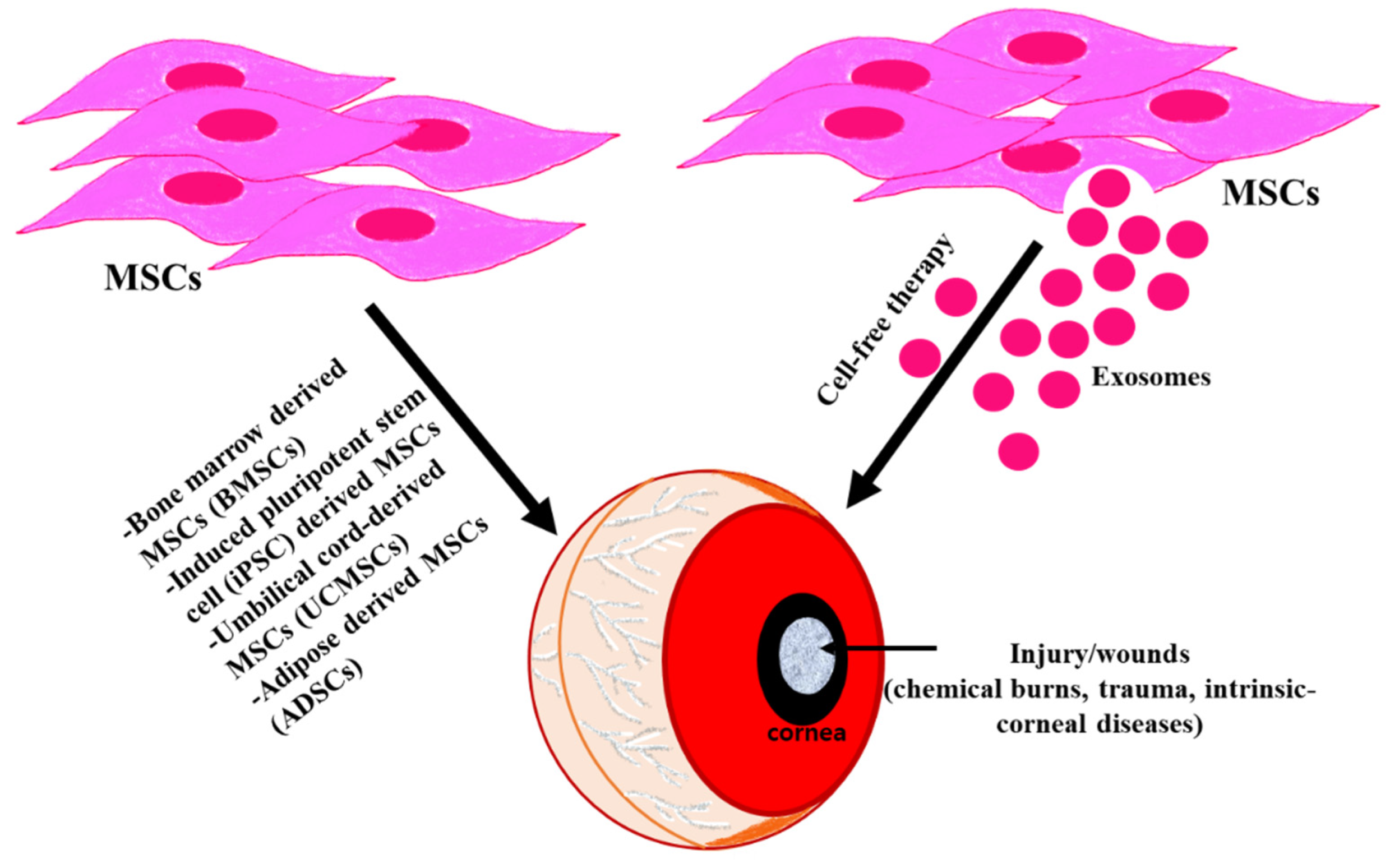
2. Mesenchymal Stem Cells (MSCs)
3. MSC Mobilization, Migration, and Homing in Corneal Changes
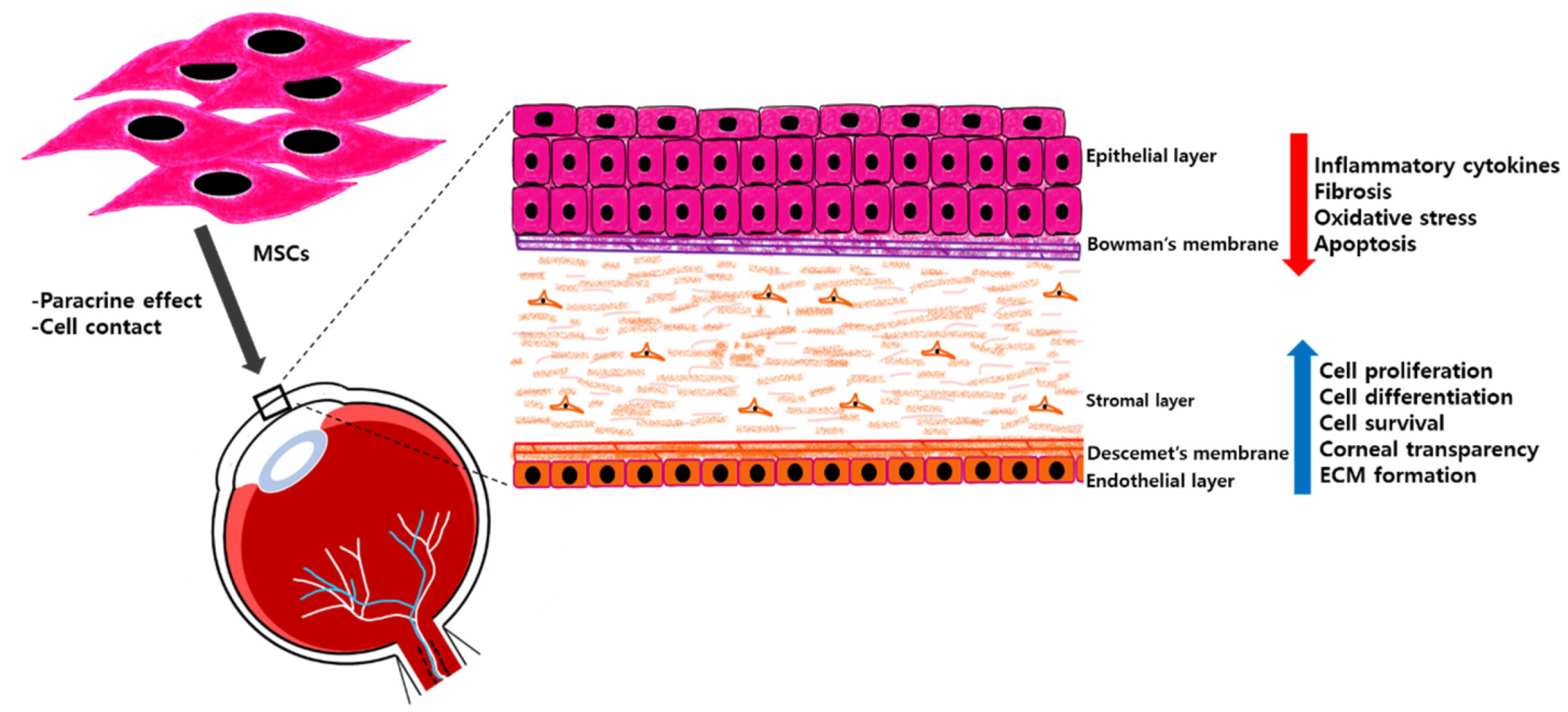
4. Corneal Regeneration with MSCs
4.1. Corneal Epithelial Regeneration with MSCs
4.2. Corneal Stromal Regeneration with MSCs
4.3. Corneal Endothelium Reconstruction with MSCs
5. Fate of MSCs in Corneal Inflammation and Angiogenesis
6. Corneal Transplantation with MSCs
7. Challenges of MSC Therapy
8. Paracrine Effect of MSCs
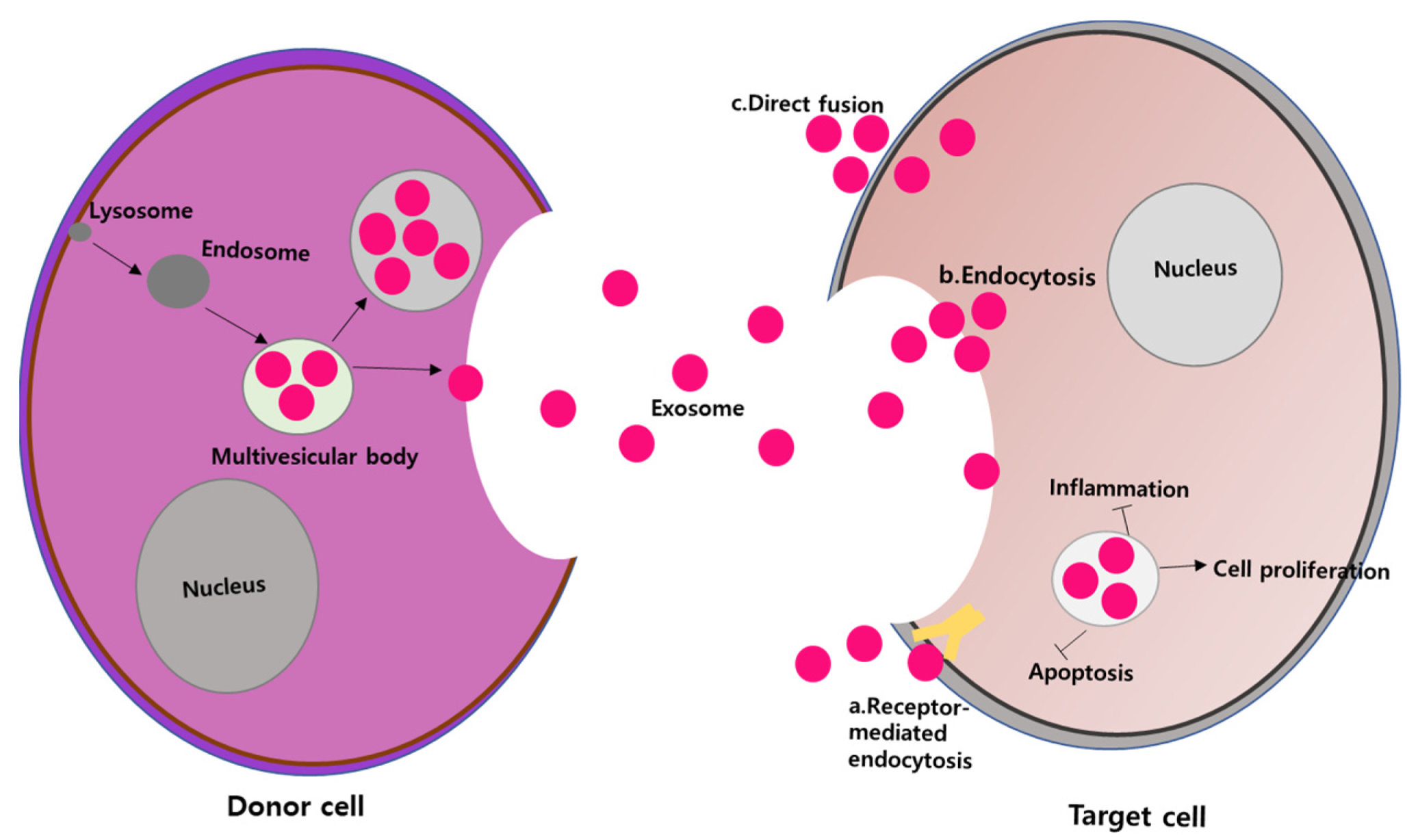
9. Mesenchymal Stem Cell-Derived Exosomes
9.1. Exosome Biogenesis
9.2. MSC-Derived Exosome Components
9.3. Isolation and Storage of Exosomes
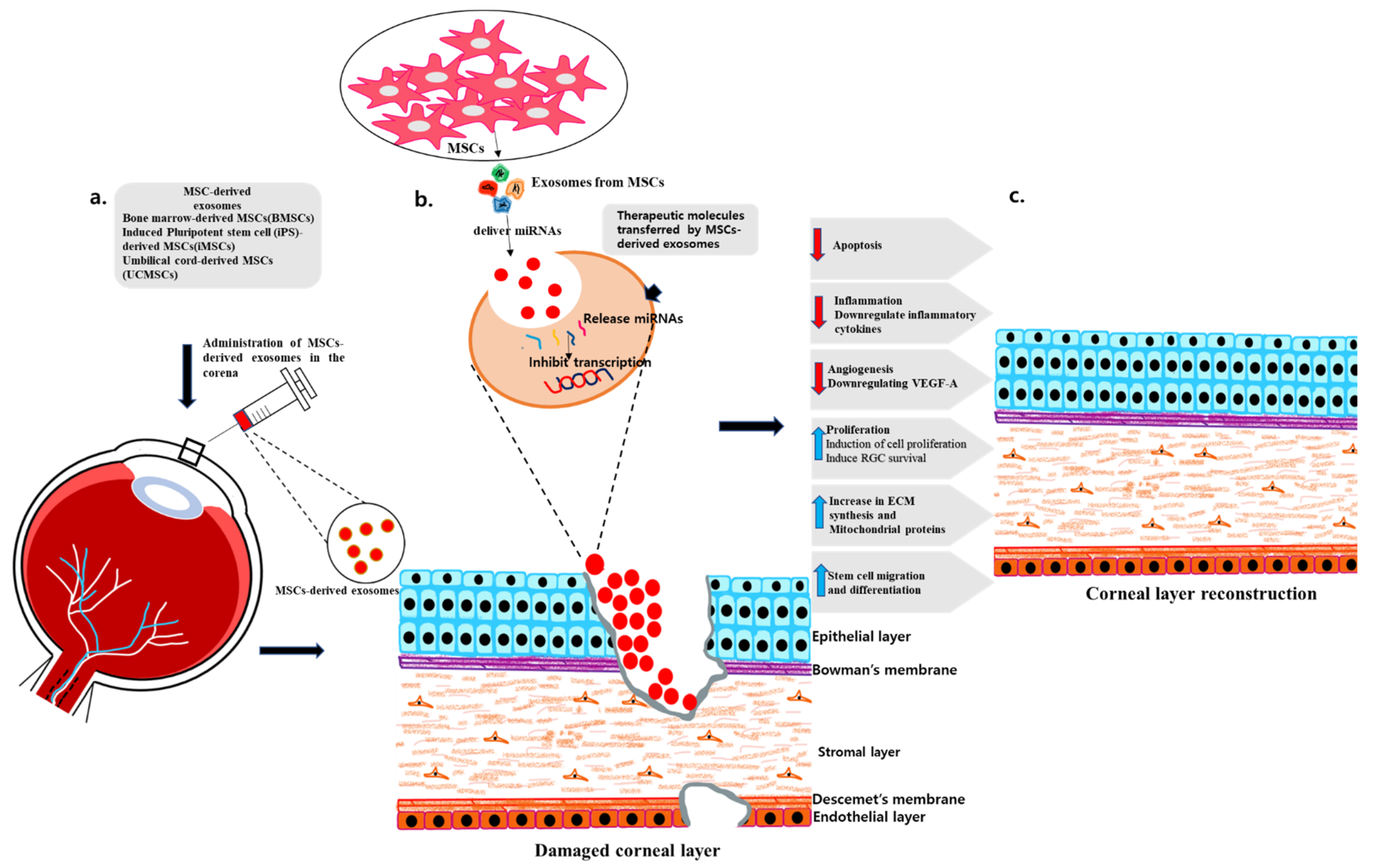
10. Therapeutic Promise of MSC-Derived Exosomes for Ocular Tissue
11. MSC-Derived Exosomes and the Cornea
12. Challenges and Future Perspectives
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MSCs | Mesenchymal stem cells |
| LSC | Limbal stem cell |
| iPSCs | Induced pluripotent stem cells |
| TSG-6 | Tumor necrosis factor-stimulated gene/protein-6 |
| ECM | Extracellular matrix |
| SPARS | Secreted protein acidic and rich in cysteine |
| PLGA | Polylactic-co-glycolic acid |
| NVs | Nanovesicles |
| TNF-α | Tumor necrosis factor-alpha |
| IL-1β | Interleukin 1 β |
| HMGB1 | High mobility group Box 1 |
| PDLSCs | Periodontal ligament stem cells |
| DPSCs | Dental pulp stem cells |
| SEC | Size exclusion chromatography |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MMP2 | Matrix metallopeptidase 2 |
| ADSCs | Adipose derived stem cells |
| TRAM2 | Translocation-associated membrane protein 2 |
| CK3 | Cytokeratin 3 |
| GSK | Glycogen synthase kinase 3 |
| TGF β | Transforming growth factor β |
| ROCK | Rho-associated protein kinase |
| MVBs | Multicellular bodies |
| ILVs | Intraluminal vesicles |
| ESCRT | Endosomal sorting complexes required for protein ESCRT |
| EAU | Experimental autoimmune uveitis |
| KCM | Keratocyte-conditioned medium |
| NVs | Nanovesicles |
| VEGF | Vascularly endothelial growth factor |
| PDGF | Platelet-derived growth factor |
| HGF | Hepatocyte growth factor |
| HCEC | Human corneal endothelial cell |
| DSAEK | Descemet’s stripping automated endothelial keratoplasty |
| DMEK | Descemet’s membrane endothelial keratoplasty |
References
- Aghamollaei, H.; Pirhadi, S.; Shafiee, S.; Sehri, M.; Goodarzi, V.; Jadidi, K. Application of polymethylmethacrylate, acrylic, and silicone in ophthalmology. In Materials for Biomedical Engineering; Elsevier: Berlin/Heidelberg, Germany, 2019; pp. 507–554. [Google Scholar]
- Eslani, M.; Baradaran-Rafii, A.; Movahedan, A.; Djalilian, A.R. The ocular surface chemical burns. J. Ophthalmol. 2014, 2014, 196827. [Google Scholar] [CrossRef] [PubMed]
- Reinhard, T.; Spelsberg, H.; Henke, L.; Kontopoulos, T.; Enczmann, J.; Wernet, P.; Berschick, P.; Sundmacher, R.; Böhringer, D. Long-term results of allogeneic penetrating limbo-keratoplasty in total limbal stem cell deficiency. Ophthalmology 2004, 111, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Eslani, M.; Haq, Z.; Movahedan, A.; Moss, A.; Baradaran-Rafii, A.; Mogilishetty, G.; Holland, E.J.; Djalilian, A.R. Late acute rejection after allograft limbal stem cell transplantation: Evidence for long-term donor survival. Cornea 2017, 36, 26. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.T.; Dart, J.K.; Holland, E.J.; Kinoshita, S. Corneal transplantation. Lancet 2012, 379, 1749–1761. [Google Scholar] [CrossRef]
- Flaxman, S.R.; Bourne, R.R.; Resnikoff, S.; Ackland, P.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H. Global causes of blindness and distance vision impairment 1990–2020: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e1221–e1234. [Google Scholar] [CrossRef]
- Avadhanam, V.S.; Liu, C.S. A brief review of Boston type-1 and osteo-odonto keratoprostheses. Br. J. Ophthalmol. 2015, 99, 878–887. [Google Scholar] [CrossRef]
- DelMonte, D.W.; Kim, T. Anatomy and physiology of the cornea. J. Cataract Refract. Surg. 2011, 37, 588–598. [Google Scholar] [CrossRef]
- Meek, K.M.; Knupp, C. Corneal structure and transparency. Prog. Retin. Eye Res. 2015, 49, 1–16. [Google Scholar] [CrossRef]
- Chong, E.-M.; Dana, M.R. Graft failure IV. Immunologic mechanisms of corneal transplant rejection. Int. Ophthalmol. 2008, 28, 209–222. [Google Scholar] [CrossRef]
- Gain, P.; Jullienne, R.; He, Z.; Aldossary, M.; Acquart, S.; Cognasse, F.; Thuret, G. Global survey of corneal transplantation and eye banking. JAMA Ophthalmol. 2016, 134, 167–173. [Google Scholar] [CrossRef]
- Fouladi, N.; Parker, M.; Kennedy, V.; Binley, K.; McCloskey, L.; Loader, J.; Kelleher, M.; Mitrophanous, K.A.; Stout, J.T.; Ellis, S. Safety and efficacy of OXB-202, a genetically engineered tissue therapy for the prevention of rejection in high-risk corneal transplant patients. Hum. Gene Ther. 2018, 29, 687–698. [Google Scholar] [CrossRef]
- Liu, S.; Wong, Y.L.; Walkden, A. Current perspectives on corneal transplantation. Clin. Ophthalmol. 2022, 16, 631–646. [Google Scholar] [CrossRef]
- Shafiq, M.A.; Gemeinhart, R.A.; Yue, B.Y.; Djalilian, A.R. Decellularized human cornea for reconstructing the corneal epithelium and anterior stroma. Tissue Eng. Part C Methods 2012, 18, 340–348. [Google Scholar] [CrossRef]
- Taylor, A.W. Ocular immune privilege and transplantation. Front. Immunol. 2016, 7, 37. [Google Scholar] [CrossRef]
- Sikora, B.; Skubis-Sikora, A.; Prusek, A.; Gola, J. Paracrine activity of adipose derived stem cells on limbal epithelial stem cells. Sci. Rep. 2021, 11, 19956. [Google Scholar] [CrossRef]
- Alvites, R.; Branquinho, M.; Sousa, A.C.; Lopes, B.; Sousa, P.; Maurício, A.C. Mesenchymal stem/stromal cells and their paracrine activity—Immunomodulation mechanisms and how to influence the therapeutic potential. Pharmaceutics 2022, 14, 381. [Google Scholar] [CrossRef]
- Bhujel, B.; Shin, H.-E.; Choi, D.-J.; Han, I. Mesenchymal stem cell-derived exosomes and intervertebral disc regeneration. Int. J. Mol. Sci. 2022, 23, 7306. [Google Scholar] [CrossRef]
- Hefley, B.S.; Deighan, C.; Vasini, B.; Khan, A.; Hjortdal, J.; Riaz, K.M.; Liu, Y.; Karamichos, D. Revealing the presence of tear extracellular vesicles in Keratoconus. Exp. Eye Res. 2022, 224, 109242. [Google Scholar] [CrossRef]
- Du, Y.; Funderburgh, M.L.; Mann, M.M.; SundarRaj, N.; Funderburgh, J.L. Multipotent stem cells in human corneal stroma. Stem Cells 2005, 23, 1266–1275. [Google Scholar] [CrossRef]
- Bajada, S.; Mazakova, I.; Richardson, J.B.; Ashammakhi, N. Updates on stem cells and their applications in regenerative medicine. J. Tissue Eng. Regen. Med. 2008, 2, 169–183. [Google Scholar] [CrossRef]
- Guillot, P.V.; Gotherstrom, C.; Chan, J.; Kurata, H.; Fisk, N.M. Human first-trimester fetal MSC express pluripotency markers and grow faster and have longer telomeres than adult MSC. Stem Cells 2007, 25, 646–654. [Google Scholar] [PubMed]
- Ho, P.-J.; Yen, M.-L.; Tang, B.-C.; Chen, C.-T.; Yen, B.L. H2O2 accumulation mediates differentiation capacity alteration, but not proliferative decline, in senescent human fetal mesenchymal stem cells. Antioxid. Redox Signal. 2013, 18, 1895–1905. [Google Scholar] [CrossRef] [PubMed]
- Yen, M.-L.; Hou, C.-H.; Peng, K.-Y.; Tseng, P.-C.; Jiang, S.-S.; Shun, C.-T.; Chen, Y.-C.; Kuo, M.-L. Efficient derivation and concise gene expression profiling of human embryonic stem cell-derived mesenchymal progenitors (EMPs). Cell Transplant. 2011, 20, 1529–1545. [Google Scholar] [CrossRef]
- Kang, S.K.; Shin, I.S.; Ko, M.S.; Jo, J.Y.; Ra, J.C. Journey of mesenchymal stem cells for homing: Strategies to enhance efficacy and safety of stem cell therapy. Stem Cells Int. 2012, 2012, 342968. [Google Scholar] [CrossRef]
- Wang, M.; Yuan, Q.; Xie, L. Mesenchymal stem cell-based immunomodulation: Properties and clinical application. Stem Cells Int. 2018, 2018, 3057624. [Google Scholar] [CrossRef] [PubMed]
- De Ugarte, D.A.; Alfonso, Z.; Zuk, P.A.; Elbarbary, A.; Zhu, M.; Ashjian, P.; Benhaim, P.; Hedrick, M.H.; Fraser, J.K. Differential expression of stem cell mobilization-associated molecules on multi-lineage cells from adipose tissue and bone marrow. Immunol. Lett. 2003, 89, 267–270. [Google Scholar] [CrossRef]
- Barlow, S.; Brooke, G.; Chatterjee, K.; Price, G.; Pelekanos, R.; Rossetti, T.; Doody, M.; Venter, D.; Pain, S.; Gilshenan, K. Comparison of human placenta-and bone marrow–derived multipotent mesenchymal stem cells. Stem Cells Dev. 2008, 17, 1095–1108. [Google Scholar] [CrossRef]
- Wang, L.; Ott, L.; Seshareddy, K.; Weiss, M.L.; Detamore, M.S. Musculoskeletal tissue engineering with human umbilical cord mesenchymal stromal cells. Regen. Med. 2011, 6, 95–109. [Google Scholar] [CrossRef]
- Skubis, A.; Gola, J.; Sikora, B.; Hybiak, J.; Paul-Samojedny, M.; Mazurek, U.; Łos, M.J. Impact of antibiotics on the proliferation and differentiation of human adipose-derived mesenchymal stem cells. Int. J. Mol. Sci. 2017, 18, 2522. [Google Scholar] [CrossRef]
- Van Harmelen, V.; Röhrig, K.; Hauner, H. Comparison of proliferation and differentiation capacity of human adipocyte precursor cells from the omental and subcutaneous adipose tissue depot of obese subjects. Metabolism 2004, 53, 632–637. [Google Scholar]
- Radtke, C.L.; Nino-Fong, R.; Gonzalez, B.P.E.; Stryhn, H.; McDuffee, L.A. Characterization and osteogenic potential of equine muscle tissue–and periosteal tissue–derived mesenchymal stem cells in comparison with bone marrow–and adipose tissue–derived mesenchymal stem cells. Am. J. Vet. Res. 2013, 74, 790–800. [Google Scholar] [CrossRef]
- Yu, B.; Zhang, X.; Li, X. Exosomes derived from mesenchymal stem cells. Int. J. Mol. Sci. 2014, 15, 4142–4157. [Google Scholar] [CrossRef]
- Baradaran-Rafii, A.; Eslani, M.; Haq, Z.; Shirzadeh, E.; Huvard, M.J.; Djalilian, A.R. Current and upcoming therapies for ocular surface chemical injuries. Ocul. Surf. 2017, 15, 48–64. [Google Scholar] [CrossRef]
- Wong, R.S. Mesenchymal stem cells: Angels or demons? J. Biomed. Biotechnol. 2011, 2011, 459510. [Google Scholar] [CrossRef]
- De Miguel, M.; Fuentes-Julián, S.; Blázquez-Martínez, A.; Pascual, C.; Aller, M.; Arias, J. Immunosuppressive properties of mesenchymal stem cells: Advances and 477 applications. Curr. Mol. Med. 2012, 12, 574–591. [Google Scholar] [CrossRef]
- Bassi, Ê.J.; de Almeida, D.C.; Moraes-Vieira, P.M.M.; Câmara, N.O.S. Exploring the role of soluble factors associated with immune regulatory properties of mesenchymal stem cells. Stem Cell Rev. Rep. 2012, 8, 329–342. [Google Scholar] [CrossRef]
- Fiorina, P.; Jurewicz, M.; Augello, A.; Vergani, A.; Dada, S.; La Rosa, S.; Selig, M.; Godwin, J.; Law, K.; Placidi, C. Immunomodulatory function of bone marrow-derived mesenchymal stem cells in experimental autoimmune type 1 diabetes. J. Immunol. 2009, 183, 993–1004. [Google Scholar] [CrossRef]
- English, K.; Ryan, J.; Tobin, L.; Murphy, M.; Barry, F.P.; Mahon, B.P. Cell contact, prostaglandin E2 and transforming growth factor beta 1 play non-redundant roles in human mesenchymal stem cell induction of CD4+ CD25Highforkhead box P3+ regulatory T cells. Clin. Exp. Immunol. 2009, 156, 149–160. [Google Scholar] [CrossRef]
- Bussche, L.; Harman, R.M.; Syracuse, B.A.; Plante, E.L.; Lu, Y.-C.; Curtis, T.M.; Ma, M.; Van de Walle, G.R. Microencapsulated equine mesenchymal stromal cells promote cutaneous wound healing in vitro. Stem Cell Res. Ther. 2015, 6, 66. [Google Scholar] [CrossRef]
- Xu, K.; Cantu, D.A.; Fu, Y.; Kim, J.; Zheng, X.; Hematti, P.; Kao, W.J. Thiol-ene Michael-type formation of gelatin/poly (ethylene glycol) biomatrices for three-dimensional mesenchymal stromal/stem cell administration to cutaneous wounds. Acta Biomater. 2013, 9, 8802–8814. [Google Scholar] [CrossRef]
- Sardesai, V.S.; Shafiee, A.; Fisk, N.M.; Pelekanos, R.A. Avoidance of maternal cell contamination and overgrowth in isolating fetal chorionic villi mesenchymal stem cells from human term placenta. Stem Cells Transl. Med. 2017, 6, 1070–1084. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.; Santos, S.G.; Gonçalves, R.M.; Teixeira, J.H.; Barbosa, M.A.; Almeida, M.I. Genetically engineered-MSC therapies for non-unions, delayed unions and critical-size bone defects. Int. J. Mol. Sci. 2019, 20, 3430. [Google Scholar] [CrossRef] [PubMed]
- Chikenji, T.S.; Saito, Y.; Konari, N.; Nakano, M.; Mizue, Y.; Otani, M.; Fujimiya, M. p16INK4A-expressing mesenchymal stromal cells restore the senescence–clearance–regeneration sequence that is impaired in chronic muscle inflammation. EBioMedicine 2019, 44, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Grafe, I.; Alexander, S.; Peterson, J.R.; Snider, T.N.; Levi, B.; Lee, B.; Mishina, Y. TGF-β family signaling in mesenchymal differentiation. Cold Spring Harb. Perspect. Biol. 2018, 10, a022202. [Google Scholar] [CrossRef]
- Willerth, S.M.; Sakiyama-Elbert, S.E. Combining stem cells and biomaterial scaffolds for constructing tissues and cell delivery. StemJournal 2019, 1, 1–25. [Google Scholar] [CrossRef]
- Lan, Y.; Kodati, S.; Lee, H.S.; Omoto, M.; Jin, Y.; Chauhan, S.K. Kinetics and function of mesenchymal stem cells in corneal injury. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3638–3644. [Google Scholar] [CrossRef]
- Ye, J.; Yao, K.; Kim, J. Mesenchymal stem cell transplantation in a rabbit corneal alkali burn model: Engraftment and involvement in wound healing. Eye 2006, 20, 482–490. [Google Scholar] [CrossRef]
- Karp, J.M.; Teo, G.S.L. Mesenchymal stem cell homing: The devil is in the details. Cell Stem Cell 2009, 4, 206–216. [Google Scholar] [CrossRef]
- Reinshagen, H.; Auw-Haedrich, C.; Sorg, R.V.; Boehringer, D.; Eberwein, P.; Schwartzkopff, J.; Sundmacher, R.; Reinhard, T. Corneal surface reconstruction using adult mesenchymal stem cells in experimental limbal stem cell deficiency in rabbits. Acta Ophthalmol. 2011, 89, 741–748. [Google Scholar] [CrossRef]
- Yao, L.; Li, Z.-R.; Su, W.-R.; Li, Y.-P.; Lin, M.-L.; Zhang, W.-X.; Liu, Y.; Wan, Q.; Liang, D. Role of mesenchymal stem cells on cornea wound healing induced by acute alkali burn. PLoS ONE 2012, 7, e30842. [Google Scholar] [CrossRef]
- Coulson-Thomas, V.J.; Caterson, B.; Kao, W.W.-Y. Transplantation of human umbilical mesenchymal stem cells cures the corneal defects of mucopolysaccharidosis VII mice. Stem Cells 2013, 31, 2116–2126. [Google Scholar]
- Jiang, T.-S.; Cai, L.; Ji, W.-Y.; Hui, Y.-N.; Wang, Y.-S.; Hu, D.; Zhu, J. Reconstruction of the corneal epithelium with induced marrow mesenchymal stem cells in rats. Mol. Vis. 2010, 16, 1304. [Google Scholar]
- Roddy, G.W.; Oh, J.Y.; Lee, R.H.; Bartosh, T.J.; Ylostalo, J.; Coble, K.; Rosa Jr, R.H.; Prockop, D.J. Action at a distance: Systemically administered adult stem/progenitor cells (MSCs) reduce inflammatory damage to the cornea without engraftment and primarily by secretion of TNF-α stimulated gene/protein 6. Stem Cells 2011, 29, 1572–1579. [Google Scholar]
- Lee, R.H.; Pulin, A.A.; Seo, M.J.; Kota, D.J.; Ylostalo, J.; Larson, B.L.; Semprun-Prieto, L.; Delafontaine, P.; Prockop, D.J. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell 2009, 5, 54–63. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, K.W.; Chun, Y.S.; Kim, J.C. Human mesenchymal stem cells differentiate into keratocyte-like cells in keratocyte-conditioned medium. Exp. Eye Res. 2012, 101, 16–26. [Google Scholar] [CrossRef]
- Ma, Y.; Xu, Y.; Xiao, Z.; Yang, W.; Zhang, C.; Song, E.; Du, Y.; Li, L. Reconstruction of chemically burned rat corneal surface by bone marrow–derived human mesenchymal stem cells. Stem Cells 2006, 24, 315–321. [Google Scholar] [CrossRef]
- Askenasy, N.; Zorina, T.; Farkas, D.L.; Shalit, I. Transplanted hematopoietic cells seed in clusters in recipient bone marrow in vivo. Stem Cells 2002, 20, 301–310. [Google Scholar] [CrossRef]
- Eslani, M.; Putra, I.; Shen, X.; Hamouie, J.; Tadepalli, A.; Anwar, K.N.; Kink, J.A.; Ghassemi, S.; Agnihotri, G.; Reshetylo, S. Cornea-derived mesenchymal stromal cells therapeutically modulate macrophage immunophenotype and angiogenic function. Stem Cells 2018, 36, 775–784. [Google Scholar] [CrossRef]
- Yao, L.; Bai, H. Mesenchymal stem cells and corneal reconstruction. Mol. Vis. 2013, 19, 2237. [Google Scholar]
- Liu, H.; Zhang, J.; Liu, C.-Y.; Wang, I.-J.; Sieber, M.; Chang, J.; Jester, J.V.; Kao, W.W. Cell therapy of congenital corneal diseases with umbilical mesenchymal stem cells: Lumican null mice. PLoS ONE 2010, 5, e10707. [Google Scholar] [CrossRef]
- Mittal, S.K.; Omoto, M.; Amouzegar, A.; Sahu, A.; Rezazadeh, A.; Katikireddy, K.R.; Shah, D.I.; Sahu, S.K.; Chauhan, S.K. Restoration of corneal transparency by mesenchymal stem cells. Stem Cell Rep. 2016, 7, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, J.; Liu, C.Y.; Hayashi, Y.; Kao, W.W.Y. Bone marrow mesenchymal stem cells can differentiate and assume corneal keratocyte phenotype. J. Cell. Mol. Med. 2012, 16, 1114–1124. [Google Scholar] [CrossRef] [PubMed]
- Galindo, S.; Herreras, J.M.; López-Paniagua, M.; Rey, E.; de la Mata, A.; Plata-Cordero, M.; Calonge, M.; Nieto-Miguel, T. Therapeutic effect of human adipose tissue-derived mesenchymal stem cells in experimental corneal failure due to limbal stem cell niche damage. Stem Cells 2017, 35, 2160–2174. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Miguel, T.; Galindo, S.; Reinoso, R.; Corell, A.; Martino, M.; Perez-Simon, J.A.; Calonge, M. In vitro simulation of corneal epithelium microenvironment induces a corneal epithelial-like cell phenotype from human adipose tissue mesenchymal stem cells. Curr. Eye Res. 2013, 38, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Setiawan, M.; Tan, X.-W.; Goh, T.-W.; Yam, G.H.-F.; Mehta, J.S. Inhibiting glycogen synthase kinase-3 and transforming growth factor-β signaling to promote epithelial transition of human adipose mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2017, 490, 1381–1388. [Google Scholar] [CrossRef]
- Gu, S.; Xing, C.; Han, J.; Tso, M.O.; Hong, J. Differentiation of rabbit bone marrow mesenchymal stem cells into corneal epithelial cells in vivo and ex vivo. Mol. Vis. 2009, 15, 99. [Google Scholar]
- Calonge, M.; Pérez, I.; Galindo, S.; Nieto-Miguel, T.; López-Paniagua, M.; Fernández, I.; Alberca, M.; García-Sancho, J.; Sánchez, A.; Herreras, J.M. A proof-of-concept clinical trial using mesenchymal stem cells for the treatment of corneal epithelial stem cell deficiency. Transl. Res. 2019, 206, 18–40. [Google Scholar] [CrossRef]
- Cejka, C.; Holan, V.; Trosan, P.; Zajicova, A.; Javorkova, E.; Cejkova, J. The favorable effect of mesenchymal stem cell treatment on the antioxidant protective mechanism in the corneal epithelium and renewal of corneal optical properties changed after alkali burns. Oxidative Med. Cell. Longev. 2016, 2016, 5843809. [Google Scholar] [CrossRef]
- Soleimanifar, F.; Mortazavi, Y.; Nadri, S.; Soleimani, M. Conjunctiva derived mesenchymal stem cell (CJMSCs) as a potential platform for differentiation into corneal epithelial cells on bioengineered electrospun scaffolds. J. Biomed. Mater. Res. Part A 2017, 105, 2703–2711. [Google Scholar] [CrossRef]
- Păunescu, V.; Deak, E.; Herman, D.; Siska, I.R.; Tănasie, G.; Bunu, C.; Anghel, S.; Tatu, C.A.; Oprea, T.I.; Henschler, R. In vitro differentiation of human mesenchymal stem cells to epithelial lineage. J. Cell. Mol. Med. 2007, 11, 502–508. [Google Scholar] [CrossRef]
- Syed-Picard, F.N.; Du, Y.; Lathrop, K.L.; Mann, M.M.; Funderburgh, M.L.; Funderburgh, J.L. Dental pulp stem cells: A new cellular resource for corneal stromal regeneration. Stem Cells Transl. Med. 2015, 4, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Roh, D.S.; Funderburgh, M.L.; Mann, M.M.; Marra, K.G.; Rubin, J.P.; Li, X.; Funderburgh, J.L. Adipose-derived stem cells differentiate to keratocytes in vitro. Mol. Vis. 2010, 16, 2680. [Google Scholar]
- Ma, X.-Y.; Bao, H.-J.; Cui, L.; Zou, J. The graft of autologous adipose-derived stem cells in the corneal stromal after mechanic damage. PLoS ONE 2013, 8, e76103. [Google Scholar] [CrossRef]
- Yam, G.H.F.; Teo, E.P.W.; Setiawan, M.; Lovatt, M.J.; Yusoff, N.Z.B.M.; Fuest, M.; Goh, B.T.; Mehta, J.S. Postnatal periodontal ligament as a novel adult stem cell source for regenerative corneal cell therapy. J. Cell. Mol. Med. 2018, 22, 3119–3132. [Google Scholar] [CrossRef]
- Yamashita, K.; Inagaki, E.; Hatou, S.; Higa, K.; Ogawa, A.; Miyashita, H.; Tsubota, K.; Shimmura, S. Corneal endothelial regeneration using mesenchymal stem cells derived from human umbilical cord. Stem Cells Dev. 2018, 27, 1097–1108. [Google Scholar] [CrossRef]
- Li, W.; Hayashida, Y.; Chen, Y.-T.; Tseng, S.C. Niche regulation of corneal epithelial stem cells at the limbus. Cell Res. 2007, 17, 26–36. [Google Scholar] [CrossRef]
- Farjo, A.; McDermott, M.; Soong, H.K. Corneal anatomy, physiology, and wound healing. Ophthalmology 2009, 44, 203–208. [Google Scholar]
- Movahedan, A.; Majdi, M.; Afsharkhamseh, N.; Sagha, H.M.; Saadat, N.S.; Shalileh, K.; Milani, B.Y.; Ying, H.; Djalilian, A.R. Notch inhibition during corneal epithelial wound healing promotes migration. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7476–7483. [Google Scholar] [CrossRef]
- Afsharkhamseh, N.; Movahedan, A.; Gidfar, S.; Huvard, M.; Wasielewski, L.; Milani, B.Y.; Eslani, M.; Djalilian, A.R. Stability of limbal stem cell deficiency after mechanical and thermal injuries in mice. Exp. Eye Res. 2016, 145, 88–92. [Google Scholar] [CrossRef]
- Dua, H.S.; Shanmuganathan, V.; Powell-Richards, A.; Tighe, P.; Joseph, A. Limbal epithelial crypts: A novel anatomical structure and a putative limbal stem cell niche. Br. J. Ophthalmol. 2005, 89, 529–532. [Google Scholar] [CrossRef]
- Phinney, D.G.; Prockop, D.J. Concise review: Mesenchymal stem/multipotent stromal cells: The state of transdifferentiation and modes of tissue repair—Current views. Stem Cells 2007, 25, 2896–2902. [Google Scholar] [CrossRef] [PubMed]
- Sikora, B.; Skubis-Sikora, A.; Kimsa-Furdzik, M.; Ciszek, W.; Kostrzewski, M.; Stojko, J.; Mazurek, U.; Gola, J. Adipose-derived stem cells undergo differentiation after co-culture with porcine limbal epithelial stem cells. Stem Cell Res. 2019, 41, 101609. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Nicolau, N.; Martínez-Conesa, E.M.; Fuentes-Julián, S.; Arnalich-Montiel, F.; García-Tuñón, I.; De Miguel, M.P.; Casaroli-Marano, R.P. Priming human adipose-derived mesenchymal stem cells for corneal surface regeneration. J. Cell. Mol. Med. 2021, 25, 5124–5137. [Google Scholar] [CrossRef] [PubMed]
- Teng, N.-Y.; Liu, Y.-S.; Wu, H.-H.; Liu, Y.-A.; Ho, J.H.; Lee, O.K.-S. Promotion of mesenchymal-to-epithelial transition by Rac1 inhibition with small molecules accelerates hepatic differentiation of mesenchymal stromal cells. Tissue Eng. Part A 2015, 21, 1444–1454. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Degyatoreva, N.; Kukielski, C.; Story, S.; Bhaduri, S.; Maiti, K.; Nahar, S.; Ray, A.; Arya, D.P.; Maiti, S. Targeting miRNA by tunable small molecule binders: Peptidic aminosugar mediated interference in miR-21 biogenesis reverts epithelial to mesenchymal transition. MedChemComm 2018, 9, 1147–1154. [Google Scholar] [CrossRef]
- Rama, P.; Ferrari, G.; Pellegrini, G. Cultivated limbal epithelial transplantation. Curr. Opin. Ophthalmol. 2017, 28, 387–389. [Google Scholar] [CrossRef]
- Ramachandran, C.; Basu, S.; Sangwan, V.S.; Balasubramanian, D. Concise review: The coming of age of stem cell treatment for corneal surface damage. Stem Cells Transl. Med. 2014, 3, 1160–1168. [Google Scholar] [CrossRef]
- Guo, T.; Wang, W.; Zhang, J.; Chen, X.; Li, B.; Li, L. Experimental study on repairing damage of corneal surface by mesenchymal stem cells transplantation. [Zhonghua Yan Ke Za Zhi] Chin. J. Ophthalmol. 2006, 42, 246–250. [Google Scholar]
- West-Mays, J.A.; Dwivedi, D.J. The keratocyte: Corneal stromal cell with variable repair phenotypes. Int. J. Biochem. Cell Biol. 2006, 38, 1625–1631. [Google Scholar] [CrossRef]
- Basu, S.; Hertsenberg, A.J.; Funderburgh, M.L.; Burrow, M.K.; Mann, M.M.; Du, Y.; Lathrop, K.L.; Syed-Picard, F.N.; Adams, S.M.; Birk, D.E. Human limbal biopsy–derived stromal stem cells prevent corneal scarring. Sci. Transl. Med. 2014, 6, 266ra172. [Google Scholar] [CrossRef]
- Kureshi, A.K.; Funderburgh, J.L.; Daniels, J.T. Human Corneal Stromal Stem Cells Exhibit Survival Capacity Following Isolation From Stored Organ–Culture Corneas. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7583–7588. [Google Scholar] [CrossRef]
- Du, Y.; Carlson, E.C.; Funderburgh, M.L.; Birk, D.E.; Pearlman, E.; Guo, N.; Kao, W.W.-Y.; Funderburgh, J.L. Stem cell therapy restores transparency to defective murine corneas. Stem Cells 2009, 27, 1635–1642. [Google Scholar] [CrossRef]
- Hertsenberg, A.J.; Shojaati, G.; Funderburgh, M.L.; Mann, M.M.; Du, Y.; Funderburgh, J.L. Corneal stromal stem cells reduce corneal scarring by mediating neutrophil infiltration after wounding. PLoS ONE 2017, 12, e0171712. [Google Scholar] [CrossRef] [PubMed]
- Funderburgh, J.L.; Funderburgh, M.L.; Du, Y. Stem cells in the limbal stroma. Ocul. Surf. 2016, 14, 113–120. [Google Scholar] [CrossRef]
- Wu, J.; Du, Y.; Watkins, S.C.; Funderburgh, J.L.; Wagner, W.R. The engineering of organized human corneal tissue through the spatial guidance of corneal stromal stem cells. Biomaterials 2012, 33, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, H.; Ong, H.S.; Riau, A.K.; Stanzel, T.P.; Mehta, J.S.; Yam, G.H.-F. Current trends and future perspective of mesenchymal stem cells and exosomes in corneal diseases. Int. J. Mol. Sci. 2019, 20, 2853. [Google Scholar] [CrossRef] [PubMed]
- Yam, G.H.-F.; Peh, G.S.-L.; Singhal, S.; Goh, B.-T.; Mehta, J.S. Dental stem cells: A future asset of ocular cell therapy. Expert Rev. Mol. Med. 2015, 17, e20. [Google Scholar] [CrossRef]
- Seo, B.-M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Trubiani, O.; Zalzal, S.F.; Paganelli, R.; Marchisio, M.; Giancola, R.; Pizzicannella, J.; Bühring, H.J.; Piattelli, M.; Caputi, S.; Nanci, A. Expression profile of the embryonic markers nanog, OCT-4, SSEA-1, SSEA-4, and frizzled-9 receptor in human periodontal ligament mesenchymal stem cells. J. Cell. Physiol. 2010, 225, 123–131. [Google Scholar] [CrossRef]
- Tomokiyo, A.; Maeda, H.; Fujii, S.; Monnouchi, S.; Wada, N.; Kono, K.; Yamamoto, N.; Koori, K.; Teramatsu, Y.; Akamine, A. A multipotent clonal human periodontal ligament cell line with neural crest cell phenotypes promotes neurocytic differentiation, migration, and survival. J. Cell. Physiol. 2012, 227, 2040–2050. [Google Scholar] [CrossRef]
- Kawanabe, N.; Murata, S.; Murakami, K.; Ishihara, Y.; Hayano, S.; Kurosaka, H.; Kamioka, H.; Takano-Yamamoto, T.; Yamashiro, T. Isolation of multipotent stem cells in human periodontal ligament using stage-specific embryonic antigen-4. Differentiation 2010, 79, 74–83. [Google Scholar] [CrossRef]
- Del Barrio, J.L.A.; El Zarif, M.; de Miguel, M.P.; Azaar, A.; Makdissy, N.; Harb, W.; El Achkar, I.; Arnalich-Montiel, F.; Alió, J.L. Cellular therapy with human autologous adipose-derived adult stem cells for advanced keratoconus. Cornea 2017, 36, 952–960. [Google Scholar] [CrossRef]
- Alió, J.L.; Del Barrio, J.L.A.; El Zarif, M.; Azaar, A.; Makdissy, N.; Khalil, C.; Harb, W.; El Achkar, I.; Jawad, Z.A.; De Miguel, M.P. Regenerative surgery of the corneal stroma for advanced keratoconus: 1-year outcomes. Am. J. Ophthalmol. 2019, 203, 53–68. [Google Scholar] [CrossRef]
- Arnalich-Montiel, F.; Pastor, S.; Blazquez-Martinez, A.; Fernandez-Delgado, J.; Nistal, M.; Alio, J.L.; De Miguel, M.P. Adipose-derived stem cells are a source for cell therapy of the corneal stroma. Stem Cells 2008, 26, 570–579. [Google Scholar] [CrossRef]
- Goldstein, A.S.; Janson, B.J.; Skeie, J.M.; Ling, J.J.; Greiner, M.A. The effects of diabetes mellitus on the corneal endothelium: A review. Surv. Ophthalmol. 2020, 65, 438–450. [Google Scholar] [CrossRef]
- Nakahara, M.; Okumura, N.; Kay, E.P.; Hagiya, M.; Imagawa, K.; Hosoda, Y.; Kinoshita, S.; Koizumi, N. Corneal endothelial expansion promoted by human bone marrow mesenchymal stem cell-derived conditioned medium. PLoS ONE 2013, 8, e69009. [Google Scholar] [CrossRef]
- Afshari, N.A.; Pittard, A.B.; Siddiqui, A.; Klintworth, G.K. Clinical study of Fuchs corneal endothelial dystrophy leading to penetrating keratoplasty: A 30-year experience. Arch. Ophthalmol. 2006, 124, 777–780. [Google Scholar] [CrossRef]
- Melles, G.R.; San Ong, T.; Ververs, B.; Van der Wees, J. Preliminary clinical results of Descemet membrane endothelial keratoplasty. Am. J. Ophthalmol. 2008, 145, 222–227 e221. [Google Scholar] [CrossRef]
- Okumura, N.; Koizumi, N.; Ueno, M.; Sakamoto, Y.; Takahashi, H.; Tsuchiya, H.; Hamuro, J.; Kinoshita, S. ROCK inhibitor converts corneal endothelial cells into a phenotype capable of regenerating in vivo endothelial tissue. Am. J. Pathol. 2012, 181, 268–277. [Google Scholar] [CrossRef]
- Kinoshita, S.; Koizumi, N.; Ueno, M.; Okumura, N.; Imai, K.; Tanaka, H.; Yamamoto, Y.; Nakamura, T.; Inatomi, T.; Bush, J. Injection of cultured cells with a ROCK inhibitor for bullous keratopathy. N. Engl. J. Med. 2018, 378, 995–1003. [Google Scholar] [CrossRef]
- Chen, S.; Zhu, Q.; Sun, H.; Zhang, Y.; Tighe, S.; Xu, L.; Zhu, Y. Advances in culture, expansion and mechanistic studies of corneal endothelial cells: A systematic review. J. Biomed. Sci. 2019, 26, 2. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, S.; Mimura, T.; Yokoo, S.; Takato, T.; Amano, S. Isolation of human corneal endothelial cell precursors and construction of cell sheets by precursors. Cornea 2006, 25, S90–S92. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-W.; Zhao, J.-L. Transplantation of autologous bone marrow mesenchymal stem cells for the treatment of corneal endothelium damages in rabbits. [Zhonghua Yan Ke Za Zhi] Chin. J. Ophthalmol. 2007, 43, 540–545. [Google Scholar]
- Shen, L.; Sun, P.; Zhang, C.; Yang, L.; Du, L.; Wu, X. Therapy of corneal endothelial dysfunction with corneal endothelial cell-like cells derived from skin-derived precursors. Sci. Rep. 2017, 7, 13400. [Google Scholar] [CrossRef]
- Gutermuth, A.; Maassen, J.; Harnisch, E.; Kuhlen, D.; Sauer-Budge, A.; Skazik-Voogt, C.; Engelmann, K. Descemet’s membrane biomimetic microtopography differentiates human mesenchymal stem cells into corneal endothelial-like cells. Cornea 2019, 38, 110. [Google Scholar] [CrossRef]
- Feiertag, E.; Maassen, J.; Mueller, A.; Harnisch, E.; Skazik-Voogt, C.; Engelmann, K.; Schrage, N.; Gutermuth, A. From Cord to Eye: Wharton Jelly-Derived Stem Cells Differentiate Into Corneal Endothelial–Like Cells. Cornea 2020, 39, 877–885. [Google Scholar] [CrossRef]
- Oh, J.Y.; Kim, M.K.; Shin, M.S.; Lee, H.J.; Ko, J.H.; Wee, W.R.; Lee, J.H. The anti-inflammatory and anti-angiogenic role of mesenchymal stem cells in corneal wound healing following chemical injury. Stem Cells 2008, 26, 1047–1055. [Google Scholar] [CrossRef]
- Pinnamaneni, N.; Funderburgh, J.L. Concise review: Stem cells in the corneal stroma. Stem Cells 2012, 30, 1059–1063. [Google Scholar] [CrossRef]
- Song, H.B.; Park, S.Y.; Ko, J.H.; Park, J.W.; Yoon, C.H.; Kim, D.H.; Kim, J.H.; Kim, M.K.; Lee, R.H.; Prockop, D.J. Mesenchymal stromal cells inhibit inflammatory lymphangiogenesis in the cornea by suppressing macrophage in a TSG-6-dependent manner. Mol. Ther. 2018, 26, 162–172. [Google Scholar] [CrossRef]
- Casiraghi, F.; Azzollini, N.; Todeschini, M.; Cavinato, R.; Cassis, P.; Solini, S.; Rota, C.; Morigi, M.; Introna, M.; Maranta, R. Localization of mesenchymal stromal cells dictates their immune or proinflammatory effects in kidney transplantation. Am. J. Transplant. 2012, 12, 2373–2383. [Google Scholar] [CrossRef]
- Oh, J.Y.; Lee, R.H.; Yu, J.M.; Ko, J.H.; Lee, H.J.; Ko, A.Y.; Roddy, G.W.; Prockop, D.J. Intravenous mesenchymal stem cells prevented rejection of allogeneic corneal transplants by aborting the early inflammatory response. Mol. Ther. 2012, 20, 2143–2152. [Google Scholar]
- Niederkorn, J.Y.; Stein-Streilein, J. History and physiology of immune privilege. Ocul. Immunol. Inflamm. 2010, 18, 19–23. [Google Scholar] [CrossRef]
- Casiraghi, F.; Remuzzi, G.; Perico, N. Mesenchymal stromal cells to promote kidney transplantation tolerance. Curr. Opin. Organ Transplant. 2014, 19, 47–53. [Google Scholar] [CrossRef]
- Coulson-Thomas, V.J.; Gesteira, T.F.; Hascall, V.; Kao, W. Umbilical cord mesenchymal stem cells suppress host rejection: The role of the glycocalyx. J. Biol. Chem. 2014, 289, 23465–23481. [Google Scholar] [CrossRef]
- Oh, J.Y.; Kim, M.K.; Ko, J.H.; Lee, H.J.; Lee, J.H.; Wee, W.R. Rat allogeneic mesenchymal stem cells did not prolong the survival of corneal xenograft in a pig-to-rat model. Vet. Ophthalmol. 2009, 12, 35–40. [Google Scholar] [CrossRef]
- Jia, Z.; Jiao, C.; Zhao, S.; Li, X.; Ren, X.; Zhang, L.; Han, Z.C.; Zhang, X. Immunomodulatory effects of mesenchymal stem cells in a rat corneal allograft rejection model. Exp. Eye Res. 2012, 102, 44–49. [Google Scholar] [CrossRef]
- Lukomska, B.; Stanaszek, L.; Zuba-Surma, E.; Legosz, P.; Sarzynska, S.; Drela, K. Challenges and controversies in human mesenchymal stem cell therapy. Stem Cells Int. 2019, 2019, 9628536. [Google Scholar] [CrossRef]
- Mo, M.; Wang, S.; Zhou, Y.; Li, H.; Wu, Y. Mesenchymal stem cell subpopulations: Phenotype, property and therapeutic potential. Cell. Mol. Life Sci. 2016, 73, 3311–3321. [Google Scholar] [CrossRef]
- Anitua, E.; Prado, R. Addressing reproducibility in stem cell and PRP therapies. Trends Biotechnol. 2019, 37, 340–344. [Google Scholar] [CrossRef]
- Dufrane, D. Impact of age on human adipose stem cells for bone tissue engineering. Cell Transplant. 2017, 26, 1496–1504. [Google Scholar] [CrossRef]
- Lohan, P.; Treacy, O.; Griffin, M.D.; Ritter, T.; Ryan, A.E. Anti-donor immune responses elicited by allogeneic mesenchymal stem cells and their extracellular vesicles: Are we still learning? Front. Immunol. 2017, 8, 1626. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-H.K. Aging of mesenchymal stem cells: Implication in regenerative medicine. Regen. Ther. 2018, 9, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Horn, P.; Castoldi, M.; Diehlmann, A.; Bork, S.; Saffrich, R.; Benes, V.; Blake, J.; Pfister, S.; Eckstein, V. Replicative senescence of mesenchymal stem cells: A continuous and organized process. PLoS ONE 2008, 3, e2213. [Google Scholar] [CrossRef] [PubMed]
- Pamies, D.; Bal-Price, A.; Simeonov, A.; Tagle, D.; Allen, D.; Gerhold, D.; Yin, D.; Pistollato, F.; Inutsuka, T.; Sullivan, K. Good cell culture practice for stem cells and stem-cell-derived models. Altern. Anim. Exp. ALTEX 2017, 34, 95–132. [Google Scholar] [CrossRef]
- King, N.M.; Perrin, J. Ethical issues in stem cell research and therapy. Stem Cell Res. Ther. 2014, 5, 85. [Google Scholar] [CrossRef]
- Lai, R.C.; Tan, S.S.; Teh, B.J.; Sze, S.K.; Arslan, F.; De Kleijn, D.P.; Choo, A.; Lim, S.K. Proteolytic potential of the MSC exosome proteome: Implications for an exosome-mediated delivery of therapeutic proteasome. Int. J. Proteom. 2012, 2012, 971907. [Google Scholar] [CrossRef]
- Chen, T.S.; Lai, R.C.; Lee, M.M.; Choo, A.B.H.; Lee, C.N.; Lim, S.K. Mesenchymal stem cell secretes microparticles enriched in pre-microRNAs. Nucleic Acids Res. 2010, 38, 215–224. [Google Scholar]
- Yin, K.; Wang, S.; Zhao, R.C. Exosomes from mesenchymal stem/stromal cells: A new therapeutic paradigm. Biomark. Res. 2019, 7, 8. [Google Scholar]
- Burrello, J.; Monticone, S.; Gai, C.; Gomez, Y.; Kholia, S.; Camussi, G. Stem cell-derived extracellular vesicles and immune-modulation. Front. Cell Dev. Biol. 2016, 4, 83. [Google Scholar] [CrossRef]
- Lin, K.-J.; Loi, M.-X.; Lien, G.-S.; Cheng, C.-F.; Pao, H.-Y.; Chang, Y.-C.; Ji, A.T.-Q.; Ho, J.H.-C. Topical administration of orbital fat-derived stem cells promotes corneal tissue regeneration. Stem Cell Res. Ther. 2013, 4, 72. [Google Scholar] [CrossRef]
- Trounson, A.; McDonald, C. Stem cell therapies in clinical trials: Progress and challenges. Cell Stem Cell 2015, 17, 11–22. [Google Scholar] [CrossRef]
- Phinney, D.G.; Pittenger, M.F. Concise review: MSC-derived exosomes for cell-free therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Pegtel, D.M.; Gould, S.J. Exosomes. Annu. Rev. Biochem. 2019, 88, 487–514. [Google Scholar] [CrossRef]
- Birtwistle, L.; Chen, X.-M.; Pollock, C. Mesenchymal stem cell-derived extracellular vesicles to the rescue of renal injury. Int. J. Mol. Sci. 2021, 22, 6596. [Google Scholar] [CrossRef]
- Sun, S.-J.; Wei, R.; Li, F.; Liao, S.-Y.; Tse, H.-F. Mesenchymal stromal cell-derived exosomes in cardiac regeneration and repair. Stem Cell Rep. 2021, 16, 1662–1673. [Google Scholar] [CrossRef]
- Tiwari, A.; Singh, A.; Verma, S.; Stephenson, S.; Bhowmick, T.; Sangwan, V.S. Mini review: Current trends and understanding of exosome therapeutic potential in corneal diseases. Front. Pharmacol. 2021, 12, 684712. [Google Scholar] [CrossRef]
- Xunian, Z.; Kalluri, R. Biology and therapeutic potential of mesenchymal stem cell-derived exosomes. Cancer Sci. 2020, 111, 3100–3110. [Google Scholar] [CrossRef]
- Nakamura, Y. Biomarkers for immune checkpoint inhibitor-mediated tumor response and adverse events. Front. Med. 2019, 6, 119. [Google Scholar] [CrossRef]
- Yu, L.-L.; Zhu, J.; Liu, J.-X.; Jiang, F.; Ni, W.-K.; Qu, L.-S.; Ni, R.-Z.; Lu, C.-H.; Xiao, M.-B. A comparison of traditional and novel methods for the separation of exosomes from human samples. BioMed Res. Int. 2018, 2018, 3634563. [Google Scholar] [CrossRef]
- Li, M.; Zhang, A.; Li, J.; Zhou, J.; Zheng, Y.; Zhang, C.; Xia, D.; Mao, H.; Zhao, J. Osteoblast/fibroblast coculture derived bioactive ECM with unique matrisome profile facilitates bone regeneration. Bioact. Mater. 2020, 5, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Chen, Z.; Liu, M.; Huang, W.; Zou, F.; Ma, X.; Tao, J.; Guo, J.; Xia, X.; Lyu, F. MSC-derived exosomes protect vertebral endplate chondrocytes against apoptosis and calcification via the miR-31-5p/ATF6 axis. Mol. Ther. -Nucleic Acids 2020, 22, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Ding, X.; Li, X.; Gao, S.; Yang, Q. 53 BP 1 suppresses epithelial–mesenchymal transition by downregulating ZEB 1 through micro RNA-200b/429 in breast cancer. Cancer Sci. 2015, 106, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Ailuno, G.; Baldassari, S.; Lai, F.; Florio, T.; Caviglioli, G. Exosomes and extracellular vesicles as emerging theranostic platforms in cancer research. Cells 2020, 9, 2569. [Google Scholar] [CrossRef] [PubMed]
- Sidhom, K.; Obi, P.O.; Saleem, A. A review of exosomal isolation methods: Is size exclusion chromatography the best option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Schiel, J.E.; Hage, D.S. Kinetic studies of biological interactions by affinity chromatography. J. Sep. Sci. 2009, 32, 1507–1522. [Google Scholar] [CrossRef]
- Wei, H.; Qian, X.; Xie, F.; Cui, D. Isolation of exosomes from serum of patients with lung cancer: A comparison of the ultra-high speed centrifugation and precipitation methods. Ann. Transl. Med. 2021, 9, 882. [Google Scholar] [CrossRef]
- Sokolova, V.; Ludwig, A.; Hornung, S.; Rotan, O.; Horn, P.; Epple, M.; Giebel, B. Colloids and surfaces B: Biointerfaces Characterisation of exosomes derived from human cells by nanoparticle tracking analysis and scanning electron microscopy. Colloids Surf. B Biointerfaces 2011, 87, 146–150. [Google Scholar] [CrossRef]
- Lorincz, Á.; Timár, C.; Marosvári, K.; Veres, D.; Otrokocsi, L.; Kittel, Á.; Ligeti, E. Effect of storage on physical and functional properties of extracellular vesicles derived from neutrophilic granulocytes. J. Extracell. Vesicles 2014, 3, 25465. [Google Scholar] [CrossRef]
- Zhou, H.; Yuen, P.S.; Pisitkun, T.; Gonzales, P.A.; Yasuda, H.; Dear, J.W.; Gross, P.; Knepper, M.A.; Star, R.A. Collection, storage, preservation, and normalization of human urinary exosomes for biomarker discovery. Kidney Int. 2006, 69, 1471–1476. [Google Scholar] [CrossRef]
- Bosch, S.; de Beaurepaire, L.; Allard, M.; Mosser, M.; Heichette, C.; Chrétien, D.; Jegou, D.; Bach, J.-M. Trehalose prevents aggregation of exosomes and cryodamage. Sci. Rep. 2016, 6, 36162. [Google Scholar] [CrossRef]
- Bai, L.; Shao, H.; Wang, H.; Zhang, Z.; Su, C.; Dong, L.; Yu, B.; Chen, X.; Li, X.; Zhang, X. Effects of mesenchymal stem cell-derived exosomes on experimental autoimmune uveitis. Sci. Rep. 2017, 7, 4323. [Google Scholar] [CrossRef]
- Yu, B.; Shao, H.; Su, C.; Jiang, Y.; Chen, X.; Bai, L.; Zhang, Y.; Li, Q.; Zhang, X.; Li, X. Exosomes derived from MSCs ameliorate retinal laser injury partially by inhibition of MCP-1. Sci. Rep. 2016, 6, 34562. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Y.; Kong, Y. Exosomes derived from mesenchymal stem cells modulate miR-126 to ameliorate hyperglycemia-induced retinal inflammation via targeting HMGB1. Investig. Ophthalmol. Vis. Sci. 2019, 60, 294–303. [Google Scholar] [CrossRef]
- Lee, J.-K.; Park, S.-R.; Jung, B.-K.; Jeon, Y.-K.; Lee, Y.-S.; Kim, M.-K.; Kim, Y.-G.; Jang, J.-Y.; Kim, C.-W. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS ONE 2013, 8, e84256. [Google Scholar] [CrossRef]
- He, C.; Zheng, S.; Luo, Y.; Wang, B. Exosome theranostics: Biology and translational medicine. Theranostics 2018, 8, 237. [Google Scholar] [CrossRef]
- Mead, B.; Tomarev, S. Bone marrow-derived mesenchymal stem cells-derived exosomes promote survival of retinal ganglion cells through miRNA-dependent mechanisms. Stem Cells Transl. Med. 2017, 6, 1273–1285. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, X.; Li, G.; Jiao, C.; Zhang, L.; Zhao, S.; Wang, J.; Han, Z.C.; Li, X. Mesenchymal stem cells ameliorate experimental autoimmune uveoretinitis by comprehensive modulation of systemic autoimmunity. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3143–3152. [Google Scholar] [CrossRef]
- Mead, B.; Berry, M.; Logan, A.; Scott, R.A.; Leadbeater, W.; Scheven, B.A. Stem cell treatment of degenerative eye disease. Stem Cell Res. 2015, 14, 243–257. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, J.; Yu, B.; Ma, F.; Ren, X.; Li, X. Effects of mesenchymal stem cells and their exosomes on the healing of large and refractory macular holes. Graefe’s Arch. Clin. Exp. Ophthalmol. 2018, 256, 2041–2052. [Google Scholar] [CrossRef]
- Shen, T.; Zheng, Q.-Q.; Shen, J.; Li, Q.-S.; Song, X.-H.; Luo, H.-B.; Hong, C.-Y.; Yao, K. Effects of adipose-derived mesenchymal stem cell exosomes on corneal stromal fibroblast viability and extracellular matrix synthesis. Chin. Med. J. 2018, 131, 704–712. [Google Scholar] [CrossRef]
- Samaeekia, R.; Rabiee, B.; Putra, I.; Shen, X.; Park, Y.J.; Hematti, P.; Eslani, M.; Djalilian, A.R. Effect of human corneal mesenchymal stromal cell-derived exosomes on corneal epithelial wound healing. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5194–5200. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Liu, G.; Meng, F.; Wang, W.; Hao, P.; Xiang, Y.; Wang, Y.; Han, R.; Li, F.; Wang, L. Paracrine effects of mesenchymal stem cells on the activation of keratocytes. Br. J. Ophthalmol. 2017, 101, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Song, W.; Teng, L.; Huang, Y.; Liu, J.; Peng, Y.; Lu, X.; Yuan, J.; Zhao, X.; Zhao, Q. MiRNA 24-3p-rich exosomes functionalized DEGMA-modified hyaluronic acid hydrogels for corneal epithelial healing. Bioact. Mater. 2022, 25, 640–656. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Lu, B.; He, J.; Chen, X.; Fu, Q.; Han, H.; Luo, C.; Yin, H.; Qin, Z.; Lyu, D. Exosomes-loaded thermosensitive hydrogels for corneal epithelium and stroma regeneration. Biomaterials 2022, 280, 121320. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Lv, Y.; Zhang, W.; Zhang, X.; Li, F.; Lu, X.; Zhao, S. Mesenchymal stem cell derived exosomes-based immunological signature in a rat model of corneal allograft rejection therapy. Front. Biosci.-Landmark 2022, 27, 86. [Google Scholar] [CrossRef]
- Liu, X.; Li, X.; Wu, G.; Qi, P.; Zhang, Y.; Liu, Z.; Li, X.; Yu, Y.; Ye, X.; Li, Y. Umbilical cord mesenchymal stem cell-derived small extracellular vesicles deliver miR-21 to promote corneal epithelial wound healing through PTEN/PI3K/Akt pathway. Stem Cells Int. 2022, 2022, 1252557. [Google Scholar] [CrossRef]
- Nuzzi, R.; Buono, L.; Scalabrin, S.; De Iuliis, M.; Bussolati, B. Effect of stem cell-derived extracellular vesicles on damaged human corneal endothelial cells. Stem Cells Int. 2021, 2021, 6644463. [Google Scholar] [CrossRef]
- Shen, T.; Zheng, Q.; Luo, H.; Li, X.; Chen, Z.; Song, Z.; Zhou, G.; Hong, C. Exosomal miR-19a from adipose-derived stem cells suppresses differentiation of corneal keratocytes into myofibroblasts. Aging 2020, 12, 4093. [Google Scholar] [CrossRef]
- Wang, S.; Hou, Y.; Li, X.; Song, Z.; Sun, B.; Li, X.; Zhang, H. Comparison of exosomes derived from induced pluripotent stem cells and mesenchymal stem cells as therapeutic nanoparticles for treatment of corneal epithelial defects. Aging 2020, 12, 19546. [Google Scholar] [CrossRef]
- Zhou, J.; Ding, Y.; Zhang, Y.; Zheng, D.; Yan, L.; Guo, M.; Mao, Y.; Yang, L. Exosomes from bone marrow-derived mesenchymal stem cells facilitate corneal wound healing via regulating the p44/42 MAPK pathway. Graefe’s Arch. Clin. Exp. Ophthalmol. 2023, 261, 723–734. [Google Scholar] [CrossRef]
- Saccu, G.; Menchise, V.; Gai, C.; Bertolin, M.; Ferrari, S.; Giordano, C.; Manco, M.; Dastrù, W.; Tolosano, E.; Bussolati, B. Bone Marrow Mesenchymal Stromal/Stem Cell-Derived Extracellular Vesicles Promote Corneal Wound Repair by Regulating Inflammation and Angiogenesis. Cells 2022, 11, 3892. [Google Scholar] [CrossRef]
- Ma, S.; Yin, J.; Hao, L.; Liu, X.; Shi, Q.; Diao, Y.; Yu, G.; Liu, L.; Chen, J.; Zhong, J. Exosomes from human umbilical cord mesenchymal stem cells treat corneal injury via autophagy activation. Front. Bioeng. Biotechnol. 2022, 10, 879192. [Google Scholar] [CrossRef]
- Tao, H.; Chen, X.; Cao, H.; Zheng, L.; Li, Q.; Zhang, K.; Han, Z.; Han, Z.-C.; Guo, Z.; Li, Z. Mesenchymal stem cell-derived extracellular vesicles for corneal wound repair. Stem Cells Int. 2019, 2019, 5738510. [Google Scholar] [CrossRef]
- Zhao, W.; He, X.; Liu, R.; Ruan, Q. Accelerating corneal wound healing using exosome-mediated targeting of NF-κB c-Rel. Inflamm. Regen. 2023, 43, 6. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, J.; Zhu, D.; Ma, S. Effect of PKH-26-labeled exosomes derived from bone marrow mesenchymal stem cells on corneal epithelium regeneration in diabetic mice. Ann. Transl. Med. 2023, 11, 167. [Google Scholar] [CrossRef]
- El-Mahalaway, A.M.; El-Azab, N.E.-E.; Abdrabbo, M.; Said, O.M.; Sabry, D.; El Sibaie, M.M. Comparative light and electron microscopic study on the therapeutic efficacy of adipose derived stem cells versus exosomes for experimentally induced acute corneal injuries in rats. Stem Cell Res. Ther. 2018, 8, 2. [Google Scholar] [CrossRef]
- Yu, Z.; Hao, R.; Du, J.; Wu, X.; Chen, X.; Zhang, Y.; Li, W.; Gu, Z.; Yang, H. A human cornea-on-a-chip for the study of epithelial wound healing by extracellular vesicles. Iscience 2022, 25, 104200. [Google Scholar] [CrossRef]
- Shang, Q.; Chu, Y.; Li, Y.; Han, Y.; Yu, D.; Liu, R.; Zheng, Z.; Song, L.; Fang, J.; Li, X. Adipose-derived mesenchymal stromal cells promote corneal wound healing by accelerating the clearance of neutrophils in cornea. Cell Death Dis. 2020, 11, 707. [Google Scholar] [CrossRef]
- Shojaati, G.; Khandaker, I.; Funderburgh, M.L.; Mann, M.M.; Basu, R.; Stolz, D.B.; Geary, M.L.; Dos Santos, A.; Deng, S.X.; Funderburgh, J.L. Mesenchymal stem cells reduce corneal fibrosis and inflammation via extracellular vesicle-mediated delivery of miRNA. Stem Cells Transl. Med. 2019, 8, 1192–1201. [Google Scholar] [CrossRef]
- Ong, H.S.; Riau, A.K.; Yam, G.H.-F.; Yusoff, N.Z.B.M.; Han, E.J.; Goh, T.-W.; Lai, R.C.; Lim, S.K.; Mehta, J.S. Mesenchymal Stem Cell Exosomes as Immunomodulatory Therapy for Corneal Scarring. Int. J. Mol. Sci. 2023, 24, 7456. [Google Scholar] [CrossRef] [PubMed]
- Dalmon, C.A.; Chandra, N.S.; Jeng, B.H. Use of autologous serum eyedrops for the treatment of ocular surface disease: First US experience in a large population as an insurance-covered benefit. Arch. Ophthalmol. 2012, 130, 1612–1613. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; He, C.; Lai, P.; Yang, Z.; Liu, Y.; Xu, H.; Lin, X.; Ni, B.; Ju, R.; Yi, W. miR-204–containing exosomes ameliorate GVHD-associated dry eye disease. Sci. Adv. 2022, 8, eabj9617. [Google Scholar] [CrossRef] [PubMed]
- Liao, F.-l.; Tan, L.; Liu, H.; Wang, J.-j.; Ma, X.-t.; Zhao, B.; Chen, Y.; Bihl, J.; Yang, Y.; Chen, R.-l. Hematopoietic stem cell-derived exosomes promote hematopoietic differentiation of mouse embryonic stem cells in vitro via inhibiting the miR126/Notch1 pathway. Acta Pharmacol. Sin. 2018, 39, 552–560. [Google Scholar] [CrossRef]
- Ko, K.-W.; Yoo, Y.-I.; Kim, J.Y.; Choi, B.; Park, S.-B.; Park, W.; Rhim, W.-K.; Han, D.K. Attenuation of tumor necrosis factor-α induced inflammation by umbilical cord-mesenchymal stem cell derived exosome-mimetic nanovesicles in endothelial cells. Tissue Eng. Regen. Med. 2020, 17, 155–163. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Corneal Tissues | MSC Source | Aim of Experiment | Human or Animal Model | Study Performed | In Vitro Study Results | In Vivo Study Results | References |
|---|---|---|---|---|---|---|---|
| Human adipose | -To investigate the impacts of inhibiting glycogen synthase kinase-3 (GSK3) and transforming growth factor β (TGF β) signaling on the epithelial differentiation of ADSCs. | Rat | In vitro In vivo | -Downregulation of the mesenchymal genes and upregulation of epithelial genes (E-cadherin, cytokeratins, and occudin). | -Demonstration of the human E-cadherin CK3, and 12 on rat corneal surface in rat model of total limbal stem cell deficiency. | [66] | |
| Human adipose | -To study whether extraocular human ADSCs exhibit some characteristics of corneal epithelial-like cells cultured in vitro. | - | In vitro | -Induction of corneal epithelial-like cells from human adipose-tissue-derived MSCs including the expression of CK3 and CK12 when cultured in corneal epithelium-conditioned media. | - | [65] | |
| Rabbit bone marrow | -To study the potential of bone-marrow-derived MSCs (BM-MSCs) to differentiate into corneal epithelial cells both in vitro and in vivo. | Rabbit | In vitro In vivo | -Differentiation of rabbit MSCs vigorously into cells with similar morphological and molecular characteristics to corneal epithelial-like cells. -Induction of CK3 expression with co-culture with rabbit limbal stem cells. | -Implantation of the cells incorporated with fibrin gel regenerated the corneal epithelium in alkali-induced corneal deficiency rabbit model and expressed CK3. | [67] | |
| Rabbit bone marrow | -To inspect the suitability of bone marrow MSCs trans-differentiating into corneal epithelial cells in a rat model with a deficit of LSCs. | Rat | In vitro In vivo | -Observation of CK12 expression and epithelial cell characteristics with co-culture on rat corneal stromal cells. | -Differentiation into epithelial-like cells expressing CK12, improving corneal opacity, and reconstructing the corneal surface in rats with transplantation on amnion in alkali injury rat corneal deficiency model. | [53] | |
| 1. Corneal epithelium | Human bone marrow | -To explore the potential of human MSCs to differentiate into corneal epithelial cells and to assess their ability to regenerate damaged corneal tissue. | Rat | In vivo | - | - Restoration of the injured or damaged surface of the cornea in rats. -Inhibition of corneal inflammation (CD45, IL-2, MMP-2) and angiogenesis in the presence of amnion in rat alkali-injured epithelial defect model. | [57] |
| Allogenic bone marrow | -To show that MSCs used in transplantation can be safe and effective and help in treating corneal pathology due to limbal stem cell deficiency (LSCD). | Human | In vivo | - | -Improvement of epithelial damage and demonstration of a more corneal epithelial-like phenotype in the central cornea with allogenic BM-MSCs. | [68] | |
| Rabbit bone marrow, adipose tissue | -To investigate whether MSCs or corneal limbal epithelial cells (LSCs) restore the corneal epithelium and optical properties in an alkali burn rabbit model. | Rabbit | In vivo | - | -Improvement in corneal optical properties, restoration of antioxidant protective mechanism and epithelial regeneration with MSCs. | [69] | |
| Conjunctiva-derived MSCs (CJMSCs) | -To identify the potential of hybrid polyurethane (PU) and silk nanofibrous scaffold with CJMSCs in treating corneal epithelium. | - | In vitro | -Improvement in the function of corneal epithelium with CJMSCs incorporated with silk fibers and PU fibers. -CJMSCs differentiate into corneal epithelial-like cells. | - | [70] | |
| Human bone marrow | -To demonstrate the aptitude of human MSCs derived from bone marrow to differentiate into functional cells with epithelial-like characteristics in vitro. | - | In vitro | -Differentiation of human MSCs derived from bone marrow into functional epithelial cells in the epithelial differentiation medium containing keratinocyte growth factor, epidermal growth factor, hepatocyte growth factor, and insulin-like growth factor. | - | [71] | |
| Human dental pulp | -To demonstrate the potential of adult dental pulp cells to differentiate into Keratocytes. | Mouse | In vitro In vivo | -Induction of Keratocan and Keratan sulfate proteoglycan (KSPG) in culture with keratocyte differentiation medium. | -Production of stromal components like human type-I collagen and Keratocan with intrastromal injection in mouse’s corneal stroma. | [72] | |
| Human adipose | -To determine whether keratocyte-specific phenotypic markers are expressed by ADSCs when cultured. | - | In vitro | -Induction of expression of stromal matrix components like KSPG, aldehyde-3-dehydrogenase-3A1 (ALDH3A1) in cell culture under reduced serum conditions supplemented with insulin and ascorbate. | - | [73] | |
| 2. Corneal stroma | Human bone marrow | -To investigate whether MSCs can differentiate into corneal keratocyte-like cells by using keratocyte-conditioned medium (KCM). | - | In vitro | -Expression of keratocyte markers such as aldehyde-1-dehydrogenase-1A1 (ALDH1A1), Lumican, and Kera in KCM. -Demonstrate MSCs could proliferate and differentiate into cells with similar characteristics to keratocytes when cultured in KCM. | - | [56] |
| Rabbit adipose | -To explore if the combination of autologous rabbit adipose-derived stem cells and polylactic-co-glycolic acid (PLGA) scaffold could be used to repair corneal stromal defects in a rabbit. | Rabbit | In vivo | - | -Differentiation of MSCs into functional keratocytes, detection of their presence up to 24 weeks following transplantation. -Differentiation to Kera and ALDH3A1 expressing cells in mechanically induced rabbit stromal defect model via transplantation of cells on a PLGA scaffold. | [74] | |
| Human periodontal ligament (PDL) | -To show the possibility of using PDL cells as a potential source for regenerative corneal cell therapy to treat corneal disorders. | - | In vivo | -Organ culture shows the presence of CD34, ALDH3A1, Kera and Lumican, collagen type 8 alpha 2 (COL8A2), CHST6 genes and their expressions. -Reduction in fibrosis, neurogenesis, and vaculogenesis gene expression. | - | [75] | |
| 3. Corneal endothelium | Human umbilical cord MSCs (HUC-MSCs) | -To study the significance of HUC-MSCs in treating corneal endothelial disease in a rabbit model with bullous keratopathy. | Rabbit | In vivo In vitro | -Stimulation of the expression of NA, K-ATPase in a medium containing GSK3β inhibitor. | -Progress in corneal thickness and transparency with cell injection in rabbit bullous keratopathy model. | [76] |
| Origin | Pros | Cons |
|---|---|---|
| MSCs | -Easy to isolate and obtain from accessible sources -High rate of proliferation, multilineal differentiation -Easily cultured in vitro -Low risk of immune-related problems -High stability in various pathological and physiological conditions | -Ethical issues -Risk of potentially transmitting genetic diseases and infections -Low number of cells -After transplantation, risk of teratoma formation is high |
| MSC-derived exosomes | -Capacity to cross natural barriers like blood–brain barrier -Perfect immune-compatibility and non-cytotoxic -Compared with cells, stable upon freezing and thawing -Ability of natural homing -Capacity for intracellular delivery of cargo by fusion of membranes | -No standard isolation protocol -No excellent mass production protocol -Less and immature research on exosome-based therapies -In vivo, after administration, fast clearance from the blood -Hard to isolate and purify the exosomes |
| Origin of Exosome | Aim of Experiment | Animal Model | Study Performed | In Vitro Study Conclusion | In Vivo Study Conclusion | References |
|---|---|---|---|---|---|---|
| ADSC exosomes | -To study how ADSCs exosomes may lead to phenotypic alterations in vitro in CSCs. | - | In vitro | -Inhibition of apoptosis, downregulation of MMPs, upregulation of ECM-related proteins (collagens and fibronectin) and significant proliferation of CSCs by ADSCs- exosomes. | - | [172] |
| Human corneal mesenchymal stromal cell-derived exosomes | -To investigate the impact of exosomes derived from human corneal MSCs on the healing of corneal epithelial wounds. | Mouse | In vitro In vivo | -Observation of corneal epithelial cell migration, proliferation, and adhesion. - Modulation of expression of genes related to cell signaling, inflammation, ECM remodeling. | -Improvement in corneal epithelial wound healing by augmenting cell proliferation. | [173] |
| BM-MSCs | -To examine how soluble factors derived from MSCs affect the functions of keratocytes (activation, migration, proliferation and synthesis of ECM). | Mouse | In vitro In vivo | -Enhancement of keratocye survival by inhibiting apoptosis, upregulation of ECM genes, increasing cell viability, and migration. | -Demonstration that various wound healing mediators like vascularly endothelial growth factor (VEGF), platelet-derived growth factor (PDGF), hepatocyte growth factor (HGF) accelerate corneal re-epithelization. | [174] |
| ADSCs exosomes | -To explore the potential of ocu-micro-RNA 24-3p to facilitate the migration and repair of rabbit corneal epithelial cells. | Rabbit | In vivo In vitro | -Increase in migration and proliferation of corneal epithelial cells. | -Promotion of rabbit corneal epithelial cell migration and repair by inhibiting fibrosis and keratitis, decreasing inflammatory reactions. | [175] |
| iPSC-MSCs exosomes | -To determine the effectiveness of exosomes obtained from iPSC-MSCs to repair damaged corneal epithelium and stromal layer, by decreasing the formation of scars and speeding up the healing process. | Rat | In vitro In vivo | -Downregulation of mRNA expression of COL1A, and COL5A2 in anterior lamellar stroma damage model in rats. -Reducing scar formation and regenerate corneal epithelium. | -Suppression of translocation-associated membrane protein 2 (TRAM2) by mi R-432-5p to prevent ECM decomposition. | [176] |
| MSC exosomes | -To study the effects of MSC-exosomes in corneal allograft rejection model. | Rat | In vivo | - | -Subconjunctival injection of 10 ug exosomes can effectively prolong the graft survival time. -Inhibit infiltration of CD4+ and CD25+T cells, downregulation of IFN-γ and CXCL11 levels in grafts. | [177] |
| HUC-MSCs-derived small extracellular vesicles (HUMSC-s EVs) | -To explore the mechanism through which HUMSCs-sEVs impact the healing process of corneal epithelial wounds. | Rat | In vitro In vivo | -Promotion of cell proliferation and migration via upregulating the P13k/Akt signaling pathway, achieved by restricting PTEN with transfer of miR-21. | -Corneal fluorescein staining and histological staining showed the healing of corneal wound in corneal mechanical wound rat model. | [178] |
| Stem cell-derived extracellular vesicles | -To explore the contribution of extracellular vesicles derived from stem cells in stem cell-induced regeneration, by reprogramming injured cells and triggering pro-degenerative pathways. | - | In vitro | -Decline in the quantity of apoptotic cells and faster wound repair in human corneal endothelial cells treatment with MSC-EVs. | - | [179] |
| ADSCs exosomes | -To examine the impact of exosomal miRNAs obtained from ADSCs on the differentiation process of rabbit corneal keratocytes. | - | In vitro | -Inhibition of HIPK2 expression suppresses the differentiation of corneal keratocytes into myofibroblasts. -Reduction in the expression of markers promoting pro-fibrosis and ECM components. | - | [180] |
| iPSCs exos and MSC exos | -To compare the efficacy of iPSCs-exos and MSC-exos in the treatment of corneal epithelial defects. | Rat | In vitro In vivo | -More promising result for iPSCs-exos than MSCs-exos. -Greater proliferation, migration, cell cycle progression, and inhibition of apoptosis in human corneal epithelial cells. | -Demonstration of stronger effects of iPSC-exos in healing corneal epithelial defect model than MSC-exos. | [181] |
| BM-MSCs exos | -To explore how BM-MSCs-exos promote corneal wound healing by activating the p44/42 MAPK signaling pathway. | Mouse | In vitro In vivo | -Enhancing the growth and migration of human corneal epithelial cells by BM-MSCs-exos. | -Downregulation of inflammation, fibrosis (α-SMA) fibrosis and vascularization (CD31) in corneal tissues of mice with alkali burn injury. | [182] |
| HUC-MSCs exos | -To explore the molecular mechanisms of HUC-MSCs-exos affect autophagy in vitro and corneal injury (CI) models in vivo. | Mouse | In vitro In vivo | -Combination of HUC- and MSCs-exos and an autophagy activator enhances cell proliferation, increases migration capacity, and boosts the expression of PCNA, Cyclin A, Cyclin E, and CDK2. | -Reduction in expression of apoptotic genes (Bax and Caspase 3), decrease in the inflammatory markers (TNF-α, IL-1β, IL-6 and CXCL-2) increment in BCL-2 in CI mice model. | [184] |
| HUC-MSCs | -To study whether the use of UMSC transplantation into corneal stroma has the potential to contribute in breakdown of glycosaminoglycans (GAGs), offering a viable approach for cell-based therapy for mucopolysaccharidoses (MPS) | Mouse | In vitro In vivo | -Release of neutral vesicles by HUC-MSCs which are taken by fibroblasts in a coculture assay | -Restoration of dendritic and hexagonal morphology of host keratocytes and endothelial cells, respectively, reduction in corneal haze. -Participation in breaking down extracellular GAGs and facilitating the host keratocytes to metabolize accumulated GAG products. | [52] |
| hMSCs exosomes | -To demonstrate the effectiveness of c-Rel-specific siRNA delivered through exosomes in accelerating corneal wound healing. | Mouse | In vivo | - | -Nano-polymers or exosomes containing c-Rel-specific siRNA on the corneal surface as a topical treatment help speed up the healing of corneal wounds in both regular and diabetic cases. | [186] |
| BM-MSCs-derived exosomes | -To examine how exosomes obtained from mouse BM-MSCs impact the regeneration of corneal epithelium in mice with diabetes. | Mouse | In vivo | - | -Restoration of corneal epithelial injury in diabetic mice by exosomes labelled with PKH-26 by downregulating the infiltration of cytokines and proliferation of corneal cells. -Detection of exosomes in the corneal stroma and nourishing it. | [187] |
| ADSCs and MSCs exosomes | -To assess and compare the effectiveness of ADSCs versus MSCs-exosome in treating corneal injuries induced by alkali in rats. | Rat | In vivo | - | -Improvement of corneal layers with decrease in inflammation and anti-angiogenic effects by MSCs-exosome-treated group in alkali burn injury model. | [188] |
| MSC exosomes | -To study the wound-healing and immunomodulatory effects of MSC exosomes in a rat corneal scarring model | Rat | In vitro In vivo | -Exerted immunomodulatory effect by modulating the expression and secretion of chemo-attractants. | -Improved the corneal epithelial wound healing by reducing corneal haze, supressing corneal neovascularization, downregulating inflamatory cytokines, and inhibiting angiogenesis in an excimer laser-induced rat corneal injury model. | [192] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhujel, B.; Oh, S.-H.; Kim, C.-M.; Yoon, Y.-J.; Kim, Y.-J.; Chung, H.-S.; Ye, E.-A.; Lee, H.; Kim, J.-Y. Mesenchymal Stem Cells and Exosomes: A Novel Therapeutic Approach for Corneal Diseases. Int. J. Mol. Sci. 2023, 24, 10917. https://doi.org/10.3390/ijms241310917
Bhujel B, Oh S-H, Kim C-M, Yoon Y-J, Kim Y-J, Chung H-S, Ye E-A, Lee H, Kim J-Y. Mesenchymal Stem Cells and Exosomes: A Novel Therapeutic Approach for Corneal Diseases. International Journal of Molecular Sciences. 2023; 24(13):10917. https://doi.org/10.3390/ijms241310917
Chicago/Turabian StyleBhujel, Basanta, Se-Heon Oh, Chang-Min Kim, Ye-Ji Yoon, Young-Jae Kim, Ho-Seok Chung, Eun-Ah Ye, Hun Lee, and Jae-Yong Kim. 2023. "Mesenchymal Stem Cells and Exosomes: A Novel Therapeutic Approach for Corneal Diseases" International Journal of Molecular Sciences 24, no. 13: 10917. https://doi.org/10.3390/ijms241310917
APA StyleBhujel, B., Oh, S.-H., Kim, C.-M., Yoon, Y.-J., Kim, Y.-J., Chung, H.-S., Ye, E.-A., Lee, H., & Kim, J.-Y. (2023). Mesenchymal Stem Cells and Exosomes: A Novel Therapeutic Approach for Corneal Diseases. International Journal of Molecular Sciences, 24(13), 10917. https://doi.org/10.3390/ijms241310917