
Comparative Transcriptome Analysis Reveals Novel Candidate Resistance Genes Involved in Defence against Phytophthora cactorum in Strawberry

 ,
,
Abstract
1. Introduction
2. Results
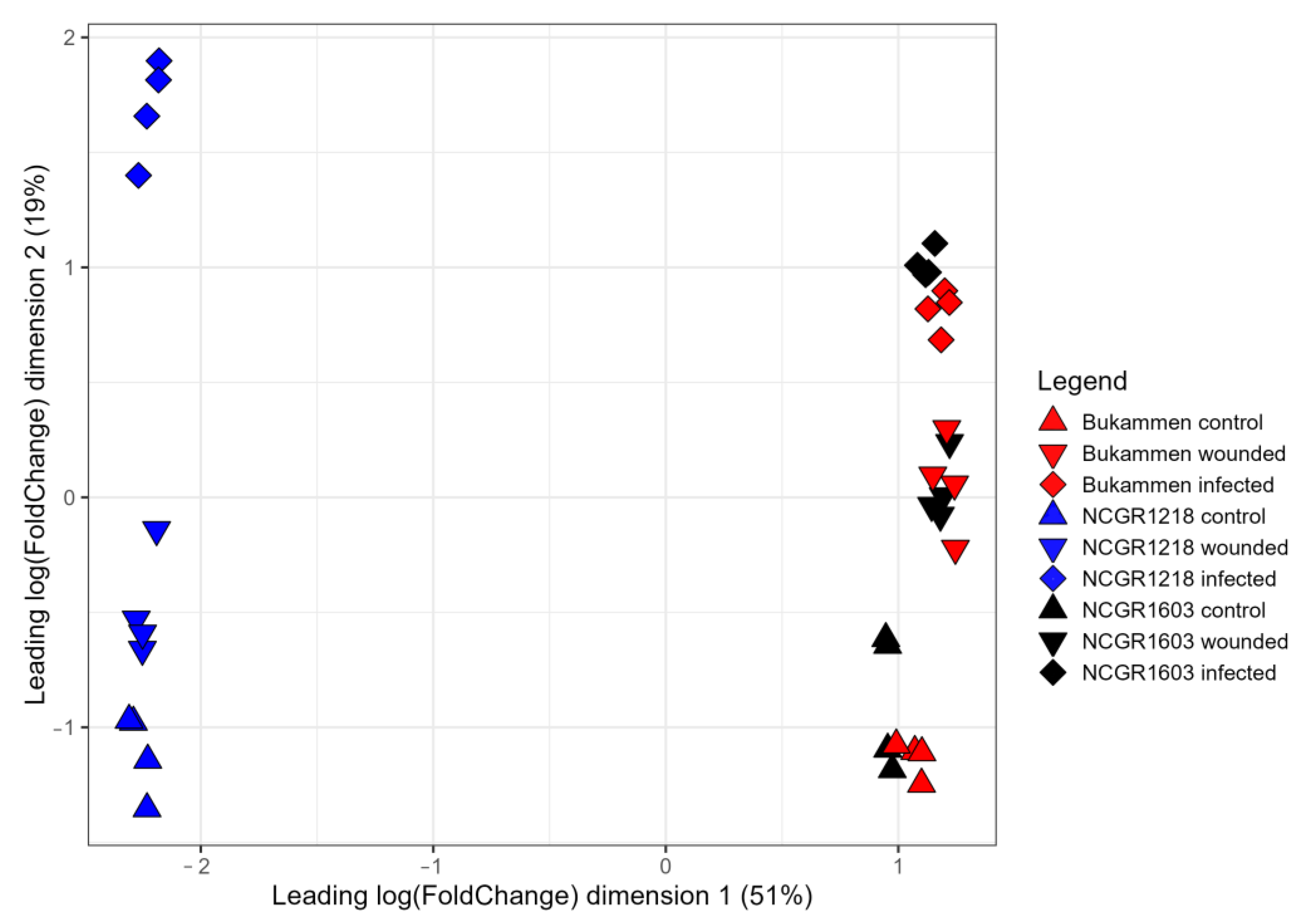
2.1. Transcriptome of Resistant and Susceptible Strawberry Genotypes Inoculated with P. cactorum
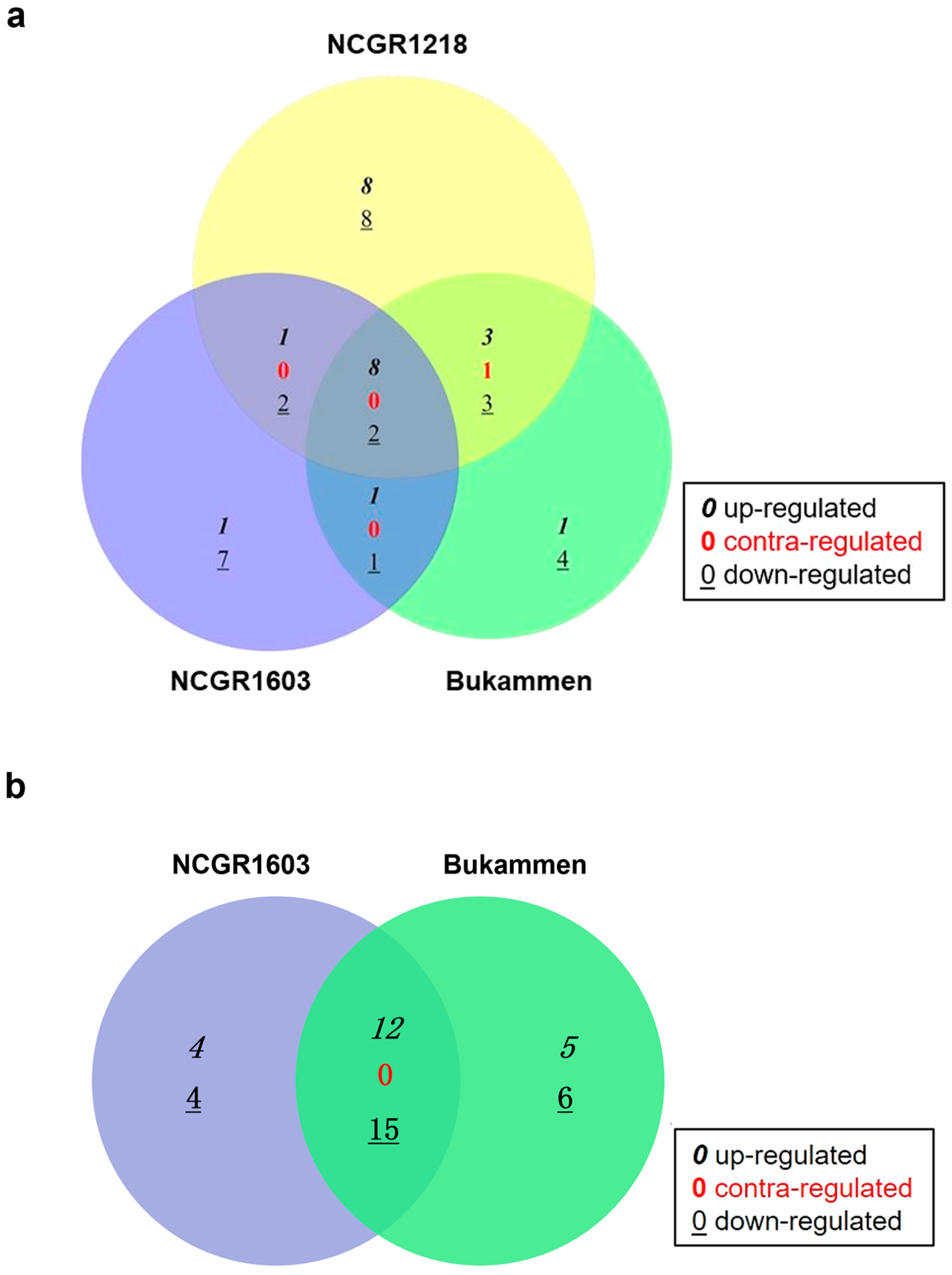
2.2. Analysis of Differentially Expressed Genes in F. vesca Genotypes
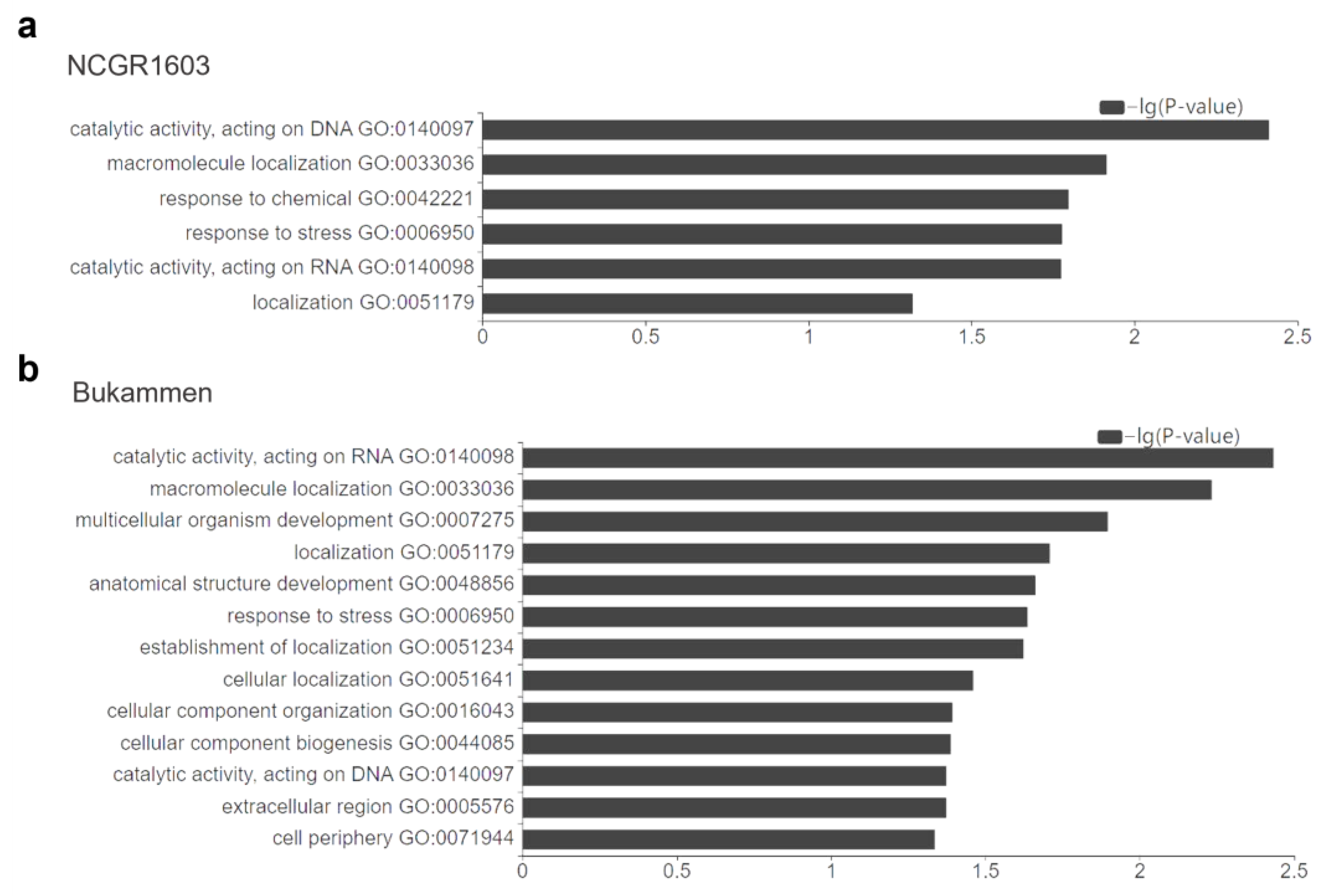
2.3. GO Term Enrichment Analysis of DEGs
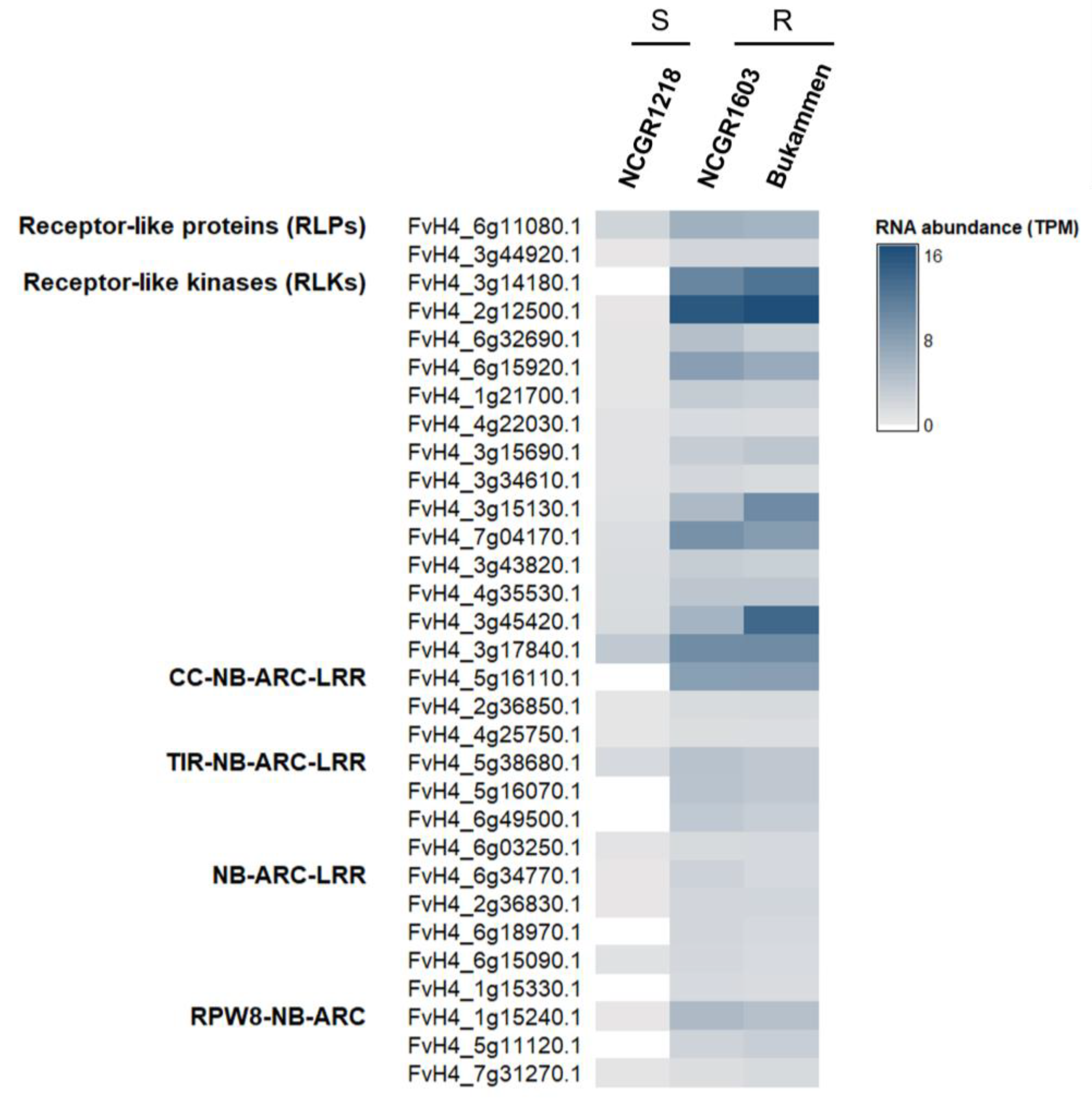
2.4. Genes Involved in Defence against P. cactorum
2.5. Expression of Defence-Related Genes in the RPc-1 Locus
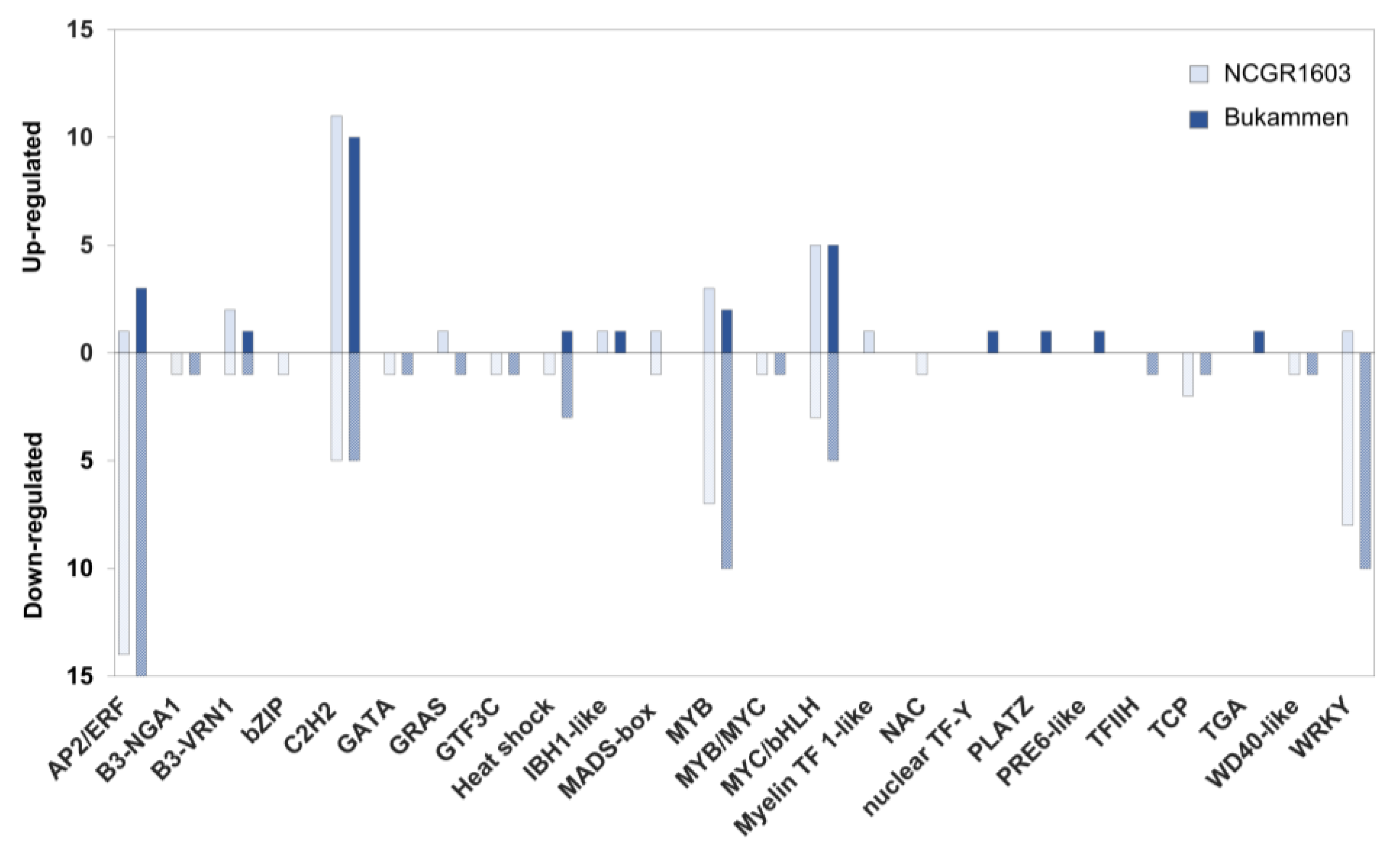
2.6. Differential Expression Analysis of Transcription Factor Genes
2.7. Flavonoid, Isoprenoid and Phytohormone Signalling Pathway Genes
2.8. Pathogenesis-Related Genes
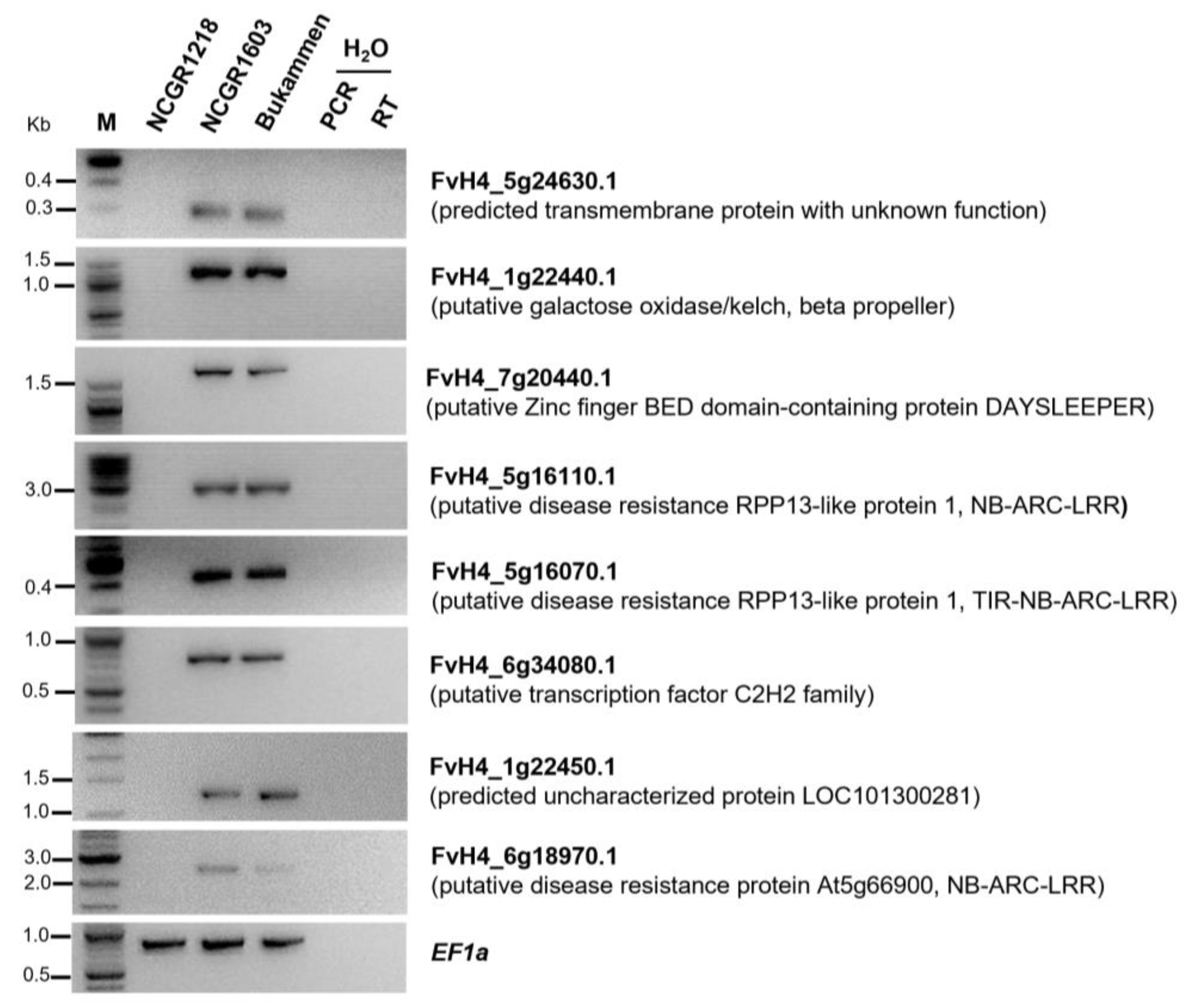
2.9. Validation of Genes Expressed Only in the Resistant Genotypes
2.10. Detection of Genes Distinct for the Resistant Genotypes
3. Discussion
4. Materials and Methods
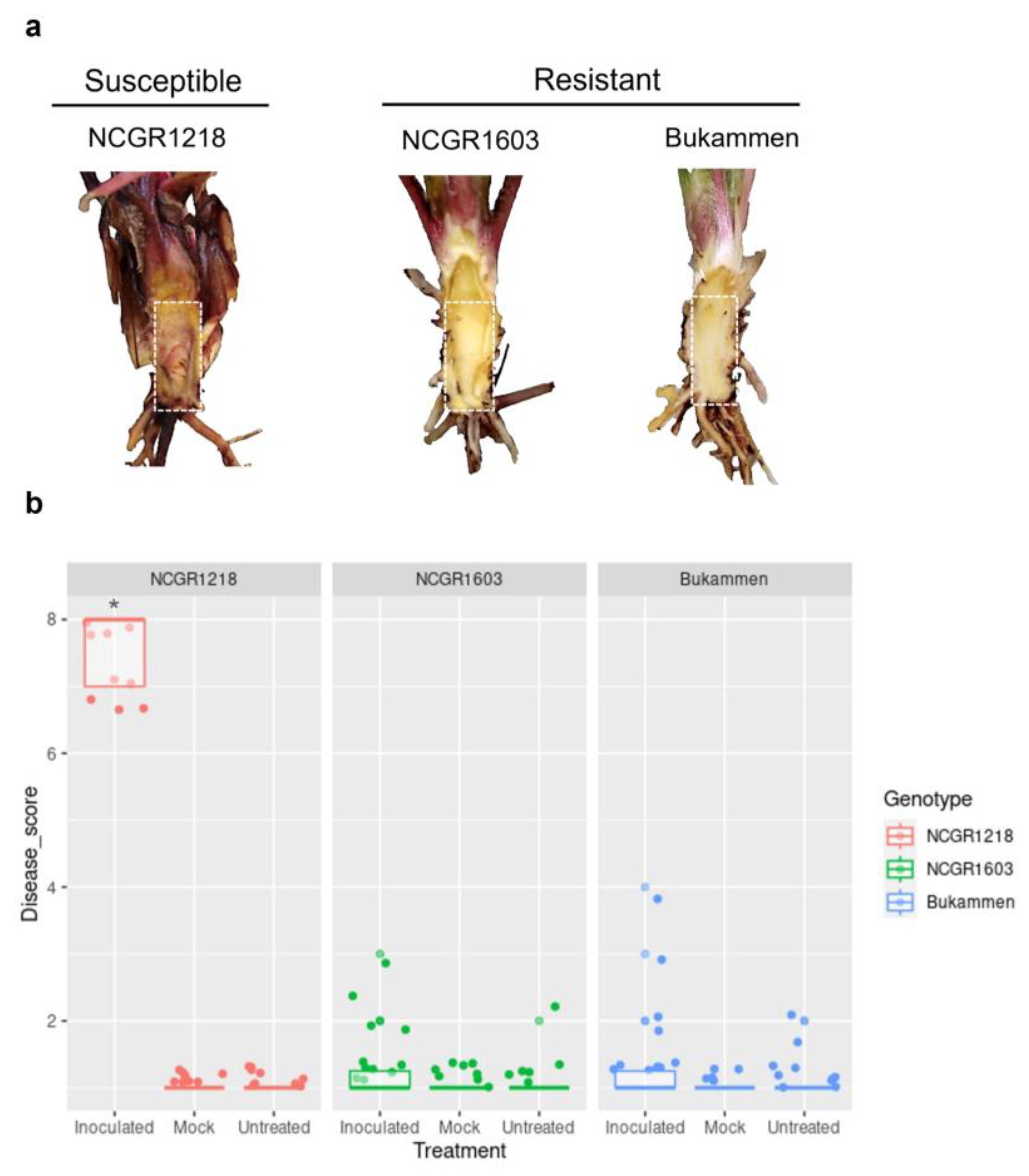
4.1. Plant Material and Pathogen Inoculation
4.2. Disease Scoring and Statistical Analysis
4.3. RNA Isolation, Library Preparation and Sequencing
4.4. De Novo Transcriptome Assembly and Data Analysis
4.5. RNA-Seq Data Validation by Semi-Quantitative Reverse Transcription-PCR (RT-PCR)
4.6. Isolation of Genomic DNA and PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, X.R.; Wen, K.; Zhou, X.; Zhu, M.Y.; Liu, Y.; Jin, J.H.; Nellist, C.F. The Devastating Oomycete Phytopathogen Phytophthora cactorum: Insights into Its Biology and Molecular Features. Mol. Plant Pathol. 2023; early view. [Google Scholar] [CrossRef]
- Golzar, H.; Phillips, D.; Mack, S. Occurrence of Strawberry Root and Crown Rot in Western Australia. Australas. Plant Dis. Notes 2007, 2, 145. [Google Scholar] [CrossRef]
- Stensvand, A.; Herrero, M.L.; Talgø, V. Crown Rot Caused by Phytophthora cactorum in Norwegian Strawberry Production. EPPO Bull. 1999, 29, 155–158. [Google Scholar] [CrossRef]
- van der Scheer, H.A.T. Isolation of Phytophthora cactorum from Soil in Orchards and Strawberry Fields and Differences in Pathogenicity to Apple. Neth. J. Plant Pathol. 1971, 77, 65–72. [Google Scholar] [CrossRef]
- Hantula, J.; Lilja, A.; Nuorteva, H.; Parikka, P.; Werres, S. Pathogenicity, Morphology and Genetic Variation of Phytophthora cactorum from Strawberry, Apple, Rhododendron, and Silver Birch. Mycol. Res. 2000, 104, 1062–1068. [Google Scholar] [CrossRef]
- Seemüller, E.; Schmidle, A. Einfluβ Der Herkunft von Phytophthora cactorum-isolaten Auf Ihre Virulenz an Apfelrinde, Erdbeerrhizomen Und Erdbeerfrüchten. J. Phytopathol. 1979, 94, 218–225. [Google Scholar] [CrossRef]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press: St. Paul, MN, USA, 1996; ISBN 9780890542125. [Google Scholar]
- Harris, D.C.; Stickels, J.E. Crown Rot (Phytophthora cactorum) in Glasshouse-Grown Strawberries at East Malling Research Station. Plant Pathol. 1981, 30, 205–212. [Google Scholar] [CrossRef]
- Deutschmann, V.F. Eine Wurzelfäule an Erdbeeren, Hervorgerufen Durch Phytophthora cactorum (Leb et Cohn) Schroet. Nachrichtenblatt Des Deutschen Pflanzen- Schutzdienst. Stuttgart 1954, 6, 7–9. [Google Scholar]
- Ellis, M.A.; Grove, G.G. Leather Rot in Ohio Strawberries. Plant Dis. 1998, 67, 549. [Google Scholar] [CrossRef]
- Ellis, M.A.; Wilcox, W.F.; Madden, L.V. Efficacy of Metalaxyl, Fosetyl-Aluminum, and Straw Mulch for Control of Strawberry Leather Rot Caused by Phytophthora cactorum. Plant Dis. 1998, 82, 329–332. [Google Scholar] [CrossRef]
- De Cal, A.; Martinez-Treceño, A.; Lopez-Aranda, J.M.; Melgarejo, P. Chemical Alternatives to Methyl Bromide in Spanish Strawberry Nurseries. Plant Dis. 2004, 88, 210–214. [Google Scholar] [CrossRef]
- Maas, J.L. Strawberry Diseases and Pests—Progress and Problems. Acta Hortic. 2014, 1049, 133–142. [Google Scholar] [CrossRef]
- Ali, A.; Kumar, R.; Mazákova, J.; Maňasová, M.; Zouhar, M.; Pánek, M. Evaluation of the Ability of Seven Active Ingredients of Fungicides to Suppress Phytophthora cactorum at Diverse Life Stages, and Variability in Resistance Found among Isolates. J. Fungi 2022, 8, 1039. [Google Scholar] [CrossRef] [PubMed]
- Hrelia, P.; Maffei, F.; Fimognari, C.; Vigagni, F.; Cantelli-Forti, G. Cytogenetic Effects of Metalaxyl on Human and Animal Chromosomes. Mutat. Res. -Genet. Toxicol. 1996, 369, 81–86. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority Conclusion on the Peer Review of the Pesticide Risk Assessment of the Active Substance Metalaxyl-M. EFSA J. 2015, 13, 3999. [CrossRef]
- Marin, M.V.; Seijo, T.E.; Zuchelli, E.; Peres, N.A. Resistance to Mefenoxam of Phytophthora cactorum and Phytophthora nicotianae Causing Crown and Leather Rot in Florida Strawberry. Plant Dis. 2021, 105, 3490–3495. [Google Scholar] [CrossRef]
- Liston, A.; Cronn, R.; Ashman, T.L. Fragaria: A Genus with Deep Historical Roots and Ripe for Evolutionary and Ecological Insights. Am. J. Bot. 2014, 101, 1686–1699. [Google Scholar] [CrossRef]
- Eikemo, H.; Brurberg, M.B.; Davik, J. Resistance to Phytophthora cactorum in Diploid Fragaria Species. HortScience 2010, 45, 193–197. [Google Scholar] [CrossRef]
- Edger, P.P.; Poorten, T.J.; VanBuren, R.; Hardigan, M.A.; Colle, M.; McKain, M.R.; Smith, R.D.; Teresi, S.J.; Nelson, A.D.L.; Wai, C.M.; et al. Origin and Evolution of the Octoploid Strawberry Genome. Nat. Genet. 2019, 51, 541–547. [Google Scholar] [CrossRef]
- Marin, M.V.; Seijo, T.E.; Baggio, J.S.; Whitaker, V.M.; Peres, N.A. Resistance of Strawberry Cultivars and the Effects of Plant Ontogenesis on Phytophthora cactorum and P. nicotianae Causing Crown Rot. Plant Dis. 2022, 107, 651–657. [Google Scholar] [CrossRef]
- Oosumi, T.; Gruszewski, H.A.; Blischak, L.A.; Baxter, A.J.; Wadl, P.A.; Shuman, J.L.; Veilleux, R.E.; Shulaev, V. High-Efficiency Transformation of the Diploid Strawberry (Fragaria vesca) for Functional Genomics. Planta 2006, 223, 1219–1230. [Google Scholar] [CrossRef]
- Folta, K.M.; Davis, T.M. Strawberry Genes and Genomics. CRC Crit. Rev. Plant Sci. 2006, 25, 399–415. [Google Scholar] [CrossRef]
- Edger, P.P.; VanBuren, R.; Colle, M.; Poorten, T.J.; Wai, C.M.; Niederhuth, C.E.; Alger, E.I.; Ou, S.; Acharya, C.B.; Wang, J.; et al. Single-Molecule Sequencing and Optical Mapping Yields an Improved Genome of Woodland Strawberry (Fragaria vesca) with Chromosome-Scale Contiguity. Gigascience 2018, 7, 1–7. [Google Scholar] [CrossRef]
- Li, Y.; Pi, M.; Gao, Q.; Liu, Z.; Kang, C. Updated Annotation of the Wild Strawberry Fragaria vesca V4 Genome. Hortic. Res. 2019, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Shulaev, V.; Sargent, D.J.; Crowhurst, R.N.; Mockler, T.C.; Folkerts, O.; Delcher, A.L.; Jaiswal, P.; Mockaitis, K.; Liston, A.; Mane, S.P.; et al. The Genome of Woodland Strawberry (Fragaria vesca). Nat. Genet. 2011, 43, 109–116. [Google Scholar] [CrossRef]
- Jones, J.; Jeffery, D. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Zipfel, C. Plant Pattern-Recognition Receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Boutrot, F.; Zipfel, C. Function, Discovery, and Exploitation of Plant Pattern Recognition Receptors for Broad-Spectrum Disease Resistance. Annu. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef] [PubMed]
- Van Ooijen, G.; Mayr, G.; Kasiem, M.M.A.; Albrecht, M.; Cornelissen, B.J.C.; Takken, F.L.W. Structure-Function Analysis of the NB-ARC Domain of Plant Disease Resistance Proteins. J. Exp. Bot. 2008, 59, 1383–1397. [Google Scholar] [CrossRef] [PubMed]
- McHale, L.; Tan, X.; Koehl, P.; Michelmore, R.W. Plant NBS-LRR Proteins: Adaptable Guards. Genome Biol. 2006, 7, 212. [Google Scholar] [CrossRef]
- Monteiro, F.; Nishimura, M.T. Structural, Functional, and Genomic Diversity of Plant NLR Proteins: An Evolved Resource for Rational Engineering of Plant Immunity. Annu. Rev. Phytopathol. 2018, 56, 243–267. [Google Scholar] [CrossRef]
- Van Wersch, S.; Tian, L.; Hoy, R.; Li, X. Plant NLRs: The Whistleblowers of Plant Immunity. Plant Commun. 2020, 1, 100016. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant Immunity: Towards an Integrated View of Plant–Pathogen Interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Ahn, H.K.; Ding, P.; Jones, J.D.G. Mutual Potentiation of Plant Immunity by Cell-Surface and Intracellular Receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.M.; He, S.Y.; Xin, X.F. Pattern-Recognition Receptors Are Required for NLR-Mediated Plant Immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Amil-Ruiz, F.; Blanco-Portales, R.; Muñoz-Blanco, J.; Caballero, J.L. The Strawberry Plant Defense Mechanism: A Molecular Review. Plant Cell Physiol. 2011, 52, 1873–1903. [Google Scholar] [CrossRef]
- Sharma, S.; Marin, M.V.; Lee, M.B.; Baggio, J.S.; Peres, N.A.; Lee, S. Genomic Approaches for Improving Resistance to Phytophthora Crown Rot Caused by P. cactorum in Strawberry (Fragaria × ananassa). Front. Agron. 2022, 4, 1–16. [Google Scholar] [CrossRef]
- Shaw, D.V.; Hansen, J.; Browne, G.T. Genotypic Variation for Resistance to Phytophthora cactorum in a California Strawberry Breeding Population. J. Am. Soc. Hortic. Sci. 2006, 131, 687–690. [Google Scholar] [CrossRef]
- Eikemo, H.; Stensvand, A. Resistance of Strawberry Genotypes to Leather Rot and Crown Rot Caused by Phytophthora cactorum. Eur. J. Plant Pathol. 2015, 143, 407–413. [Google Scholar] [CrossRef]
- Eikemo, H.; Stensvand, A.; Davik, J.; Tronsmo, A.M. Resistance to Crown Rot (Phytophthora cactorum) in Strawberry Cultivars and in Offspring from Crosses between Cultivars Differing in Susceptibility to the Disease. Ann. Appl. Biol. 2003, 142, 83–89. [Google Scholar] [CrossRef]
- Shaw, D.V.; Hansen, J.; Browne, G.T.; Shaw, S.M. Components of Genetic Variation for Resistance of Strawberry to Phytophthora cactorum Estimated Using Segregating Seedling Populations and Their Parent Genotypes. Plant Pathol. 2008, 57, 210–215. [Google Scholar] [CrossRef]
- Davik, J.; Eikemo, H.; Brurberg, M.B.; Sargent, D.J. Mapping of the RPc-1 Locus for Phytophthora cactorum Resistance in Fragaria vesca. Mol. Breed. 2015, 35, 1–11. [Google Scholar] [CrossRef]
- Nellist, C.F.; Vickerstaff, R.J.; Sobczyk, M.K.; Marina-Montes, C.; Wilson, F.M.; Simpson, D.W.; Whitehouse, A.B.; Harrison, R.J. Quantitative Trait Loci Controlling Phytophthora cactorum Resistance in the Cultivated Octoploid Strawberry (Fragaria × ananassa). Hortic. Res. 2019, 6, 60. [Google Scholar] [CrossRef]
- Mangandi, J.; Verma, S.; Osorio, L.; Peres, N.A.; van de Weg, E.; Whitaker, V.M. Pedigree-Based Analysis in a Multiparental Population of Octoploid Strawberry Reveals QTL Alleles Conferring Resistance to Phytophthora cactorum. G3 Genes Genomes Genet. 2017, 7, 1707–1719. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-R.; Brurberg, M.B.; Elameen, A.; Klemsdal, S.S.; Martinussen, I. Expression of Resistance Gene Analogs in Woodland Strawberry (Fragaria vesca) during Infection with Phytophthora cactorum. Mol. Genet. Genom. 2016, 291, 1967–1978. [Google Scholar] [CrossRef] [PubMed]
- Toljamo, A.; Blande, D.; Kärenlampi, S.; Kokko, H. Reprogramming of Strawberry (Fragaria vesca) Root Transcriptome in Response to Phytophthora cactorum. PLoS ONE 2016, 11, e0161078. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.A.; Simpson, D.W.; Harris, D.C. Development of a Method for Screening Strawberry Germplasm for Resistance to Phytophthora cactorum. Acta Hortic. 1997, 439, 175–179. [Google Scholar] [CrossRef]
- Xu, X.M.; Lin, H.; Maple, J.; Björkblom, B.; Alves, G.; Larsen, J.P.; Møller, S.G. The Arabidopsis DJ-1a Protein Confers Stress Protection through Cytosolic SOD Activation. J. Cell Sci. 2010, 123, 1644–1651. [Google Scholar] [CrossRef]
- Lin, J.; Nazarenus, T.J.; Frey, J.L.; Liang, X.; Wilson, M.A.; Stone, J.M. A Plant DJ-1 Homolog Is Essential for Arabidopsis thaliana Chloroplast Development. PLoS ONE 2011, 6, e23731. [Google Scholar] [CrossRef]
- Andrade, M.A.; González-Guzmán, M.; Serrano, R.; Rodríguez, P.L. A Combination of the F-Box Motif and Kelch Repeats Defines a Large Arabidopsis Family of F-Box Proteins. Plant Mol. Biol. 2001, 46, 603–614. [Google Scholar] [CrossRef]
- Baron, A.J.; Stevens, C.; Wilmot, C.; Seneviratne, K.D.; Blakeley, V.; Dooley, D.M.; Phillips, S.E.V.; Knowles, P.F.; McPherson, M.J. Structure and Mechanism of Galactose Oxidase. The Free Radical Site. J. Biol. Chem. 1994, 269, 25095–25105. [Google Scholar] [CrossRef]
- Akiyama, Y. Quality Control of Cytoplasmic Membrane Proteins in Escherichia coli. J. Biochem. 2009, 146, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Sakoh, M.; Ito, K.; Akiyama, Y. Proteolytic Activity of HtpX, a Membrane-Bound and Stress-Controlled Protease from Escherichia coli. J. Biol. Chem. 2005, 280, 33305–33310. [Google Scholar] [CrossRef] [PubMed]
- Marchal, C.; Zhang, J.; Zhang, P.; Fenwick, P.; Steuernagel, B.; Adamski, N.M.; Boyd, L.; McIntosh, R.; Wulff, B.B.H.; Berry, S.; et al. BED-Domain-Containing Immune Receptors Confer Diverse Resistance Spectra to Yellow Rust. Nat. Plants 2018, 4, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Bundock, P.; Hooykaas, P. An Arabidopsis HAT-like Transposase Is Essential for Plant Development. Nature 2005, 436, 282–284. [Google Scholar] [CrossRef]
- Rawat, N.; Pumphrey, M.O.; Liu, S.; Zhang, X.; Tiwari, V.K.; Ando, K.; Trick, H.N.; Bockus, W.W.; Akhunov, E.; Anderson, J.A.; et al. Wheat Fhb1 Encodes a Chimeric Lectin with Agglutinin Domains and a Pore-Forming Toxin-like Domain Conferring Resistance to Fusarium Head Blight. Nat. Genet. 2016, 48, 1576–1580. [Google Scholar] [CrossRef]
- Gish, L.A.; Clark, S.E. The RLK/Pelle Family of Kinases. Plant J. 2011, 66, 117–127. [Google Scholar] [CrossRef]
- Lehti-Shiu, M.D.; Zou, C.; Hanada, K.; Shiu, S.H. Evolutionary History and Stress Regulation of Plant Receptor-like Kinase/Pelle Genes. Plant Physiol. 2009, 150, 12–26. [Google Scholar] [CrossRef]
- Bittner-Eddy, P.D.; Crute, I.R.; Holub, E.B.; Beynon, J.L. RPP13 Is a Simple Locus in Arabidopsis thaliana for Alleles That Specify Downy Mildew Resistance to Different Avirulence Determinants in Peronospora parasitica. Plant J. 2000, 21, 177–188. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Z.; Xu, X.; Zhang, H.; Li, C. Genome-Wide Analysis of C2H2 Zinc-Finger Family Transcription Factors and Their Responses to Abiotic Stresses in Poplar (Populus trichocarpa). PLoS ONE 2015, 10, e0134753. [Google Scholar] [CrossRef]
- Gourcilleau, D.; Lenne, C.; Armenise, C.; Moulia, B.; Julien, J.L.; Bronner, G.; Leblanc-Fournier, N. Phylogenetic Study of Plant Q-Type C2H2 Zinc Finger Proteins and Expression Analysis of Poplar Genes in Response to Osmotic, Cold and Mechanical Stresses. DNA Res. 2011, 18, 77–92. [Google Scholar] [CrossRef]
- Grosse-Holz, F.M.; van der Hoorn, R.A.L. Juggling Jobs: Roles and Mechanisms of Multifunctional Protease Inhibitors in Plants. New Phytol. 2016, 210, 794–807. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.S.; Pearce, G.; Merryweather, J.; Titani, K.; Ericsson, L.H.; Ryan, C.A. Wound-Induced Proteinase Inhibitors from Tomato Leaves. II. The CDNA-Deduced Primary Structure of Pre-Inhibitor II. J. Biol. Chem. 1985, 260, 6561–6564. [Google Scholar] [CrossRef] [PubMed]
- Doxey, A.C.; Yaish, M.W.F.; Moffatt, B.A.; Griffith, M.; McConkey, B.J. Functional Divergence in the Arabidopsis β-1,3-Glucanase Gene Family Inferred by Phylogenetic Reconstruction of Expression States. Mol. Biol. Evol. 2007, 24, 1045–1055. [Google Scholar] [CrossRef] [PubMed]
- Meins, F.; Ahl, P. Induction of Chitinase and β-1,3-Glucanase in Tobacco Plants Infected with Pseudomonas tabaci and Phytophthora parasitica var. Nicotianae. Plant Sci. 1989, 61, 155–161. [Google Scholar] [CrossRef]
- Moeder, W.; Urquhart, W.; Ung, H.; Yoshioka, K. The Role of Cyclic Nucleotide-Gated Ion Channels in Plant Immunity. Mol. Plant 2011, 4, 442–452. [Google Scholar] [CrossRef]
- Saand, M.A.; Xu, Y.P.; Li, W.; Wang, J.P.; Cai, X.Z. Cyclic Nucleotide Gated Channel Gene Family in Tomato: Genome-Wide Identification and Functional Analyses in Disease Resistance. Front. Plant Sci. 2015, 6, 303. [Google Scholar] [CrossRef]
- Dinesh-Kumar, S.P.; Tham, W.H.; Baker, B.J. Structure-Function Analysis of the Tobacco Mosaic Virus Resistance Gene N. Proc. Natl. Acad. Sci. USA 2000, 97, 14789–14794. [Google Scholar] [CrossRef]
- Benedetti, M.; Verrascina, I.; Pontiggia, D.; Locci, F.; Mattei, B.; De Lorenzo, G.; Cervone, F. Four Arabidopsis Berberine Bridge Enzyme-like Proteins Are Specific Oxidases That Inactivate the Elicitor-Active Oligogalacturonides. Plant J. 2018, 94, 260–273. [Google Scholar] [CrossRef]
- Laluk, K.; Abuqamar, S.; Mengiste, T. The Arabidopsis Mitochondria-Localized Pentatricopeptide Repeat Protein PGN Functions in Defense against Necrotrophic Fungi and Abiotic Stress Tolerance. Plant Physiol. 2011, 156, 2053–2068. [Google Scholar] [CrossRef]
- Park, Y.J.; Lee, H.J.; Kwak, K.J.; Lee, K.; Hong, S.W.; Kang, H. MicroRNA400-Guided Cleavage of Pentatricopeptide Repeat Protein MRNAs Renders Arabidopsis thaliana More Susceptible to Pathogenic Bacteria and Fungi. Plant Cell Physiol. 2014, 55, 1660–1668. [Google Scholar] [CrossRef]
- Xiao, S.; Ellwood, S.; Calis, O.; Patrick, E.; Li, T.; Coleman, M.; Turner, J.G. Broad-Spectrum Mildew Resistance in Arabidopsis thaliana Mediated by RPW8. Science 2001, 291, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Cheong, Y.H.; Chang, H.; Gupta, R.; Wang, X.; Zhu, T.; Luan, S. Transcriptional Profiling Reveals Novel Interactions between Wounding, Pathogen, Abiotic Stress, and Hormonal Responses in Arabidopsis. Plant Physiol. 2002, 129, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Durrant, W.E.; Rowland, O.; Piedras, P.; Hammond-kosack, K.E.; Jones, J.D.G. CDNA-AFLP Reveals a Striking Overlap in Race-Specific Resistance and Wound Response Gene Expression Profiles. Plant Cell 2000, 12, 963–977. [Google Scholar] [CrossRef] [PubMed]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the Plant Tissue: The Defense of a Dangerous Passage. Front. Plant Sci. 2014, 5, 470. [Google Scholar] [CrossRef] [PubMed]
- Naveed, Z.A.; Ali, G.S. Comparative Transcriptome Analysis between a Resistant and a Susceptible Wild Tomato Accession in Response to Phytophthora parasitica. Int. J. Mol. Sci. 2018, 19, 3735. [Google Scholar] [CrossRef]
- Santos, S.A.; Vidigal, P.M.P.; Guimarães, L.M.S.; Mafia, R.G.; Templeton, M.D.; Alfenas, A.C. Transcriptome Analysis of Eucalyptus grandis Genotypes Reveals Constitutive Overexpression of Genes Related to Rust (Austropuccinia psidii) Resistance. Plant Mol. Biol. 2020, 104, 339–357. [Google Scholar] [CrossRef]
- Meng, H.; Sun, M.; Jiang, Z.; Liu, Y.; Sun, Y.; Liu, D.; Jiang, C.; Ren, M.; Yuan, G.; Yu, W.; et al. Comparative Transcriptome Analysis Reveals Resistant and Susceptible Genes in Tobacco Cultivars in Response to Infection by Phytophthora nicotianae. Sci. Rep. 2021, 11, 809. [Google Scholar] [CrossRef]
- Odilbekov, F.; He, X.; Armoniené, R.; Saripella, G.V.; Henriksson, T.; Singh, P.K.; Chawade, A. QTL Mapping and Transcriptome Analysis to Identify Differentially Expressed Genes Induced by Septoria Tritici Blotch Disease of Wheat. Agronomy 2019, 9, 510. [Google Scholar] [CrossRef]
- Zhao, Y.; Vlasselaer, L.; Ribeiro, B.; Terzoudis, K.; Van den Ende, W.; Hertog, M.; Nicolaï, B.; De Coninck, B. Constitutive Defense Mechanisms Have a Major Role in the Resistance of Woodland Strawberry Leaves against Botrytis cinerea. Front. Plant Sci. 2022, 13, 1–18. [Google Scholar] [CrossRef]
- Patkar, R.N.; Chattoo, B.B. Transgenic Indica Rice Expressing Ns-LTP-like Protein Shows Enhanced Resistance to Both Fungal and Bacterial Pathogens. Mol. Breed. 2006, 17, 159–171. [Google Scholar] [CrossRef]
- Gomès, E.; Sagot, E.; Gaillard, C.; Laquitaine, L.; Poinssot, B.; Sanejouand, Y.H.; Delrot, S.; Coutos-Thévenot, P. Nonspecific Lipid-Transfer Protein Genes Expression in Grape (Vitis sp.) Cells in Response to Fungal Elicitor Treatments. Mol. Plant-Microbe Interact. 2003, 16, 456–464. [Google Scholar] [CrossRef]
- Park, C.J.; Shin, R.; Park, J.M.; Lee, G.J.; You, J.S.; Paek, K.H. Induction of Pepper CDNA Encoding a Lipid Transfer Protein during the Resistance Response to Tobacco Mosaic Virus. Plant Mol. Biol. 2002, 48, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, A.M.; Doerner, P.; Dixonk, R.A.; Lamb, C.J.; Cameron, R.K. A Putative Lipid Transfer Protein Involved in Systemic Resistance Signalling in Arabidopsis. Nature 2002, 419, 399–403. [Google Scholar] [CrossRef]
- Wang, S.Y.; Wu, J.H.; Ng, T.B.; Ye, X.Y.; Rao, P.F. A Non-Specific Lipid Transfer Protein with Antifungal and Antibacterial Activities from the Mung Bean. Peptides 2004, 25, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.S.; Johnson, J.W.; Seo, Y.W. Differential Expression of TaLTP3 and TaCOMT1 Induced by Hessian Fly Larval Infestation in a Wheat Line Possessing H21 Resistance Gene. Plant Sci. 2005, 168, 1319–1326. [Google Scholar] [CrossRef]
- Kruijt, M.; De Kock, M.J.D.; De Wit, P.J.G.M. Receptor-like Proteins Involved in Plant Disease Resistance. Mol. Plant Pathol. 2005, 6, 85–97. [Google Scholar] [CrossRef]
- Brutus, A.; Sicilia, F.; Macone, A.; Cervone, F.; De Lorenzo, G. A Domain Swap Approach Reveals a Role of the Plant Wall-Associated Kinase 1 (WAK1) as a Receptor of Oligogalacturonides. Proc. Natl. Acad. Sci. USA 2010, 107, 9452–9457. [Google Scholar] [CrossRef]
- Li, H.; Zhou, S.Y.; Zhao, W.S.; Su, S.C.; Peng, Y.L. A Novel Wall-Associated Receptor-like Protein Kinase Gene, OsWAK1, Plays Important Roles in Rice Blast Disease Resistance. Plant Mol. Biol. 2009, 69, 337–346. [Google Scholar] [CrossRef]
- Zuo, W.; Chao, Q.; Zhang, N.; Ye, J.; Tan, G.; Li, B.; Xing, Y.; Zhang, B.; Liu, H.; Fengler, K.A.; et al. A Maize Wall-Associated Kinase Confers Quantitative Resistance to Head Smut. Nat. Genet. 2015, 47, 151–157. [Google Scholar] [CrossRef]
- Shen, D.; Chai, C.; Ma, L.; Zhang, M.; Dou, D. Comparative RNA-Seq Analysis of Nicotiana benthamiana in Response to Phytophthora parasitica Infection. Plant Growth Regul. 2016, 80, 59–67. [Google Scholar] [CrossRef]
- Le Berre, J.Y.; Gourgues, M.; Samans, B.; Keller, H.; Panabières, F.; Attard, A. Transcriptome Dynamic of Arabidopsis Roots Infected with Phytophthora parasitica Identifies VQ29, a Gene Induced during the Penetration and Involved in the Restriction of Infection. PLoS ONE 2017, 12, e0190341. [Google Scholar] [CrossRef] [PubMed]
- Kiełbowicz-Matuk, A. Involvement of Plant C2H2-Type Zinc Finger Transcription Factors in Stress Responses. Plant Sci. 2012, 185–186, 78–85. [Google Scholar] [CrossRef]
- Kim, S.H.; Hong, J.K.; Lee, S.C.; Sohn, K.H.; Jung, H.W.; Hwang, B.K. CAZFP1, Cys2/His2-Type Zinc-Finger Transcription Factor Gene Functions as a Pathogen-Induced Early-Defense Gene in Capsicum annuum. Plant Mol. Biol. 2004, 55, 883–904. [Google Scholar] [CrossRef]
- Lawrence, S.D.; Novak, N.G.; Perez, F.G.; Jones, R.W. Over Expression of the Q-Type ZFP StZFP2 in Potato Increases Resistance to Potato Late Blight (Phytophthora infestans) Infection. J. Plant Interact. 2019, 14, 129–136. [Google Scholar] [CrossRef]
- Ward, E.R.; Uknes, S.J.; Williams, S.C.; Dincher, S.S.; Wiederhold, D.L.; Alexander, D.C.; Ahl-Goy, P.; Métraux, J.P.; Ryals, J.A. Coordinate Gene Activity in Response to Agents That Induce Systemic Acquired Resistance. Plant Cell 1991, 3, 1085–1094. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Rakwal, R.; Jwa, N.S.; Agrawal, V.P. Signalling Molecules and Blast Pathogen Attack Activates Rice OsPR1a and OsPR1b Genes: A Model Illustrating Components Participating during Defence/Stress Response. Plant Physiol. Biochem. 2001, 39, 1095–1103. [Google Scholar] [CrossRef]
- Niderman, T.; Genetet, I.; Bruyère, T.; Gees, R.; Stintzi, A.; Legrand, M.; Fritig, B.; Mösinger, E. Pathogenesis-Related PR-1 Proteins Are Antifungal. Lsolation and Characterization of Three 14-Kilodalton Proteins of Tomato and of a Basic PR-1 of Tobacco with Inhibitory Activity against Phytophthora infestans. Plant Physiol. 1995, 108, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Gamir, J.; Darwiche, R.; van’t Hof, P.; Choudhary, V.; Stumpe, M.; Schneiter, R.; Mauch, F. The Sterol-Binding Activity of PATHOGENESIS-RELATED PROTEIN 1 Reveals the Mode of Action of an Antimicrobial Protein. Plant J. 2017, 89, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Cui, X.; Lin, S.; Gan, S.; Xing, H.; Dou, D. GmCYP82A3, a Soybean Cytochrome P450 Family Gene Involved in the Jasmonic Acid and Ethylene Signaling Pathway, Enhances Plant Resistance to Biotic and Abiotic Stresses. PLoS ONE 2016, 11, e0162253. [Google Scholar] [CrossRef]
- Halim, V.A.; Altmann, S.; Ellinger, D.; Eschen-Lippold, L.; Miersch, O.; Scheel, D.; Rosahl, S. PAMP-Induced Defense Responses in Potato Require Both Salicylic Acid and Jasmonic Acid. Plant J. 2009, 57, 230–242. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, X.; Fernando, W.G.D. Directing Trophic Divergence in Plant-Pathogen Interactions: Antagonistic Phytohormones with No Doubt? Front. Plant Sci. 2020, 11, e600063. [Google Scholar] [CrossRef] [PubMed]
- Eshraghi, L.; Anderson, J.P.; Aryamanesh, N.; McComb, J.A.; Shearer, B.; Hardy, G.S.J.E. Suppression of the Auxin Response Pathway Enhances Susceptibility to Phytophthora cinnamomi While Phosphite-Mediated Resistance Stimulates the Auxin Signalling Pathway. BMC Plant Biol. 2014, 14, 68. [Google Scholar] [CrossRef] [PubMed]
- Llorente, F.; Muskett, P.; Sánchez-Vallet, A.; López, G.; Ramos, B.; Sánchez-Rodríguez, C.; Jordá, L.; Parker, J.; Molina, A. Repression of the Auxin Response Pathway Increases Arabidopsis Susceptibility to Necrotrophic Fungi. Mol. Plant 2008, 1, 496–509. [Google Scholar] [CrossRef]
- Mutka, A.M.; Fawley, S.; Tsao, T.; Kunkel, B.N. Auxin Promotes Susceptibility to Pseudomonas syringae via a Mechanism Independent of Suppression of Salicylic Acid-Mediated Defenses. Plant J. 2013, 74, 746–754. [Google Scholar] [CrossRef]
- Armitage, A.D.; Lysøe, E.; Nellist, C.F.; Lewis, L.A.; Cano, L.M.; Harrison, R.J.; Brurberg, M.B. Bioinformatic Characterisation of the Effector Repertoire of the Strawberry Pathogen Phytophthora cactorum. PLoS ONE 2018, 13, e0202305. [Google Scholar] [CrossRef]
- Eikemo, H.; Stensvand, A.; Tronsmo, A.M. Evaluation of Methods of Screening Strawberry Cultivars for Resistance to Crown Rot Caused by Phytophthora cactorum. Ann. Appl. Biol. 2000, 137, 237–244. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-Optimal Probabilistic RNA-Seq Quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Wagner, G.P.; Kin, K.; Lynch, V.J. Measurement of MRNA Abundance Using RNA-Seq Data: RPKM Measure Is Inconsistent among Samples. Theory Biosci. 2012, 131, 281–285. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-Throughput Functional Annotation and Data Mining with the Blast2GO Suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Ye, J.; Zhang, Y.; Cui, H.; Liu, J.; Wu, Y.; Cheng, Y.; Xu, H.; Huang, X.; Li, S.; Zhou, A.; et al. WEGO 2.0: A Web Tool for Analyzing and Plotting GO Annotations, 2018 Update. Nucleic Acids Res. 2018, 46, W71–W75. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Chen, H.; Yi, T.; Daimon, C.M.; Boyle, J.P.; Peers, C.; Maudsley, S.; Martin, B. VennPlex-a Novel Venn Diagram Program for Comparing and Visualizing Datasets with Differentially Regulated Datapoints. PLoS ONE 2013, 8, e53388. [Google Scholar] [CrossRef] [PubMed]
- Clancy, M.A.; Rosli, H.G.; Chamala, S.; Barbazuk, W.B.; Civello, P.M.; Folta, K.M. Validation of Reference Transcripts in Strawberry (Fragaria spp.). Mol. Genet. Genom. 2013, 288, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.F.; Ferreira, J.L.; Fernandes, M.C.N.; Breves, S.d.S.; Generoso, A.L.; Soares, B.D.F.; Dias, M.S.C.; Pasqual, M.; Borem, A.; Cançado, G.M.d.A. An Improved Method for Genomic DNA Extraction from Strawberry Leaves. Ciência Rural 2011, 41, 1383–1389. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | RNA Abundance 48 h after Inoculation (Mean Transcripts per Million ± SE) | Annotation Based on NCBI Non-Redundant Database and InterProScan Domain Analysis | Putative Role in Defence | Reference | ||
|---|---|---|---|---|---|---|
| NCGR1218 | NCGR1603 | Bukammen | ||||
| FvH4_7g05680.1 | 0 ± 0.0 | 28 ± 1.0 | 33 ± 1.0 | Protein DJ-1 homolog D-like | Cellular defence response against oxidative stress, and chloroplast development | [49,50] |
| FvH4_1g22440.1 | 0 ± 0.0 | 28 ± 1.0 | 35 ± 3 | Putative galactose oxidase with kelch/beta-propeller | Production of H2O2 from galactose and O2; interaction with ASK1/MAP3K5 | [51,52] |
| FvH4_6g53350.1 | 0 ± 0.0 | 27 ± 2.0 | 32 ± 2.0 | Protease HtpX homolog, peptidase M48 | Stress-controlled protease in E. coli; proteolytic quality control of misfolded proteins | [53,54] |
| FvH4_7g20440.1 | 0 ± 0.0 | 16 ± 1.0 | 19 ± 1.0 | Zinc finger BED domain-containing protein DAYSLEEPER | Influence on global gene expression in Arabidopsis thaliana; BED-NLRs act as immune receptors in wheat (Triticum aestivum) | [55,56] |
| FvH4_5g03090.1 | 0 ± 0.0 | 11 ± 3.0 | 11 ± 3.0 | Putative Clostridium epsilon toxin ETX/mosquitocidal toxin MTX2; agglutinin domain | Resistance to Fusarium graminearum | [57] |
| FvH4_3g14180.1 | 0 ± 0.0 | 10 ± 1.0 | 12 ± 1.0 | Kinase RLK-Pelle-LRR-XI-1 family, protein kinase domain | Immune receptor against diverse pathogens; also involved in growth and development | [58,59] |
| FvH4_5g16110.1 | 0 ± 0.0 | 8 ± 0.5 | 8 ± 0.4 | Putative disease resistance RPP13-like protein 1, leucine-rich repeat | Resistance to biotrophic oomycete Peronospora parasitica | [60] |
| FvH4_6g34080.1 | 0 ± 0.0 | 7 ± 0.4 | 7 ± 0.4 | Putative transcription factor C2H2 family, zinc finger, RING-type | Response to abiotic stresses in plants | [61,62] |
| FvH4_5g32170.1 | 0 ± 0.0 | 6 ± 2.0 | 5 ± 1.0 | Proteinase inhibitor PSI-1.2, proteinase inhibitor I20 | Expression induced upon wounding and involved in plant defence against herbivory | [63,64] |
| FvH4_3g28530.1 | 0 ± 0.0 | 4 ± 0.5 | 3 ± 0.3 | Glucan endo-1,3-beta-glucosidase 11-like | Response to pathogen infection in plants | [65,66] |
| FvH4_6g00620.1 | 0 ± 0.0 | 4 ± 0.2 | 5 ± 0.5 | Putative transcription factor C2H2 family | Response to abiotic stresses in plants | [61,62] |
| FvH4_5g16070.1 | 0 ± 0.0 | 4 ± 0.2 | 3 ± 0.2 | Putative disease resistance RPP13-like protein 1 | Resistance to biotrophic oomycete Peronospora parasitica | [60] |
| FvH4_6g40960.1 | 0 ± 0.0 | 4 ± 0.2 | 3 ± 0.3 | Cyclic nucleotide-gated ion channel 1-like | Active role in plant immunity in several pathosystems | [67,68] |
| FvH4_6g49500.1 | 0 ± 0.0 | 3 ± 0.8 | 3 ± 0.1 | TMV resistance protein N-like, Toll/interleukin-1 receptor homology (TIR) domain | Resistance to Tobacco mosaic virus in Nicotiana benthamiana | [69] |
| FvH4_6g51870.1 | 0 ± 0.0 | 3 ± 0.4 | 2 ± 0.3 | Putative transcription factor C2H2 family, zinc finger, RING-type | Response to abiotic stresses in plants | [61,62] |
| FvH4_3g45070.1 | 0 ± 0.0 | 3 ± 0.1 | 7 ± 1.0 | Putative transcription factor C2H2 family, zinc finger, RING-type | Response to abiotic stresses in plants | [61,62] |
| FvH4_3g11010.1 | 0 ± 0.0 | 2 ± 0.2 | 4 ± 1.0 | Berberine bridge enzyme-like 8, FAD linked oxidase, N-terminal | Enhanced resistance to Botrytis cinerea | [70] |
| FvH4_5g35070.1 | 0 ± 0.0 | 2 ± 0.4 | 4 ± 0.3 | Putative pentatricopeptide repeat protein | Defence against biotic stress (Pseudomonas syringae pv. tomato and Botrytis cinerea) and abiotic stress | [71,72] |
| FvH4_5g11120.1 | 0 ± 0.0 | 2 ± 0.2 | 3 ± 0.1 | Putative powdery mildew resistance protein, RPW8 | Broad spectrum mildew resistance in Arabidopsis thaliana | [73] |
| FvH4_5g35080.1 | 0 ± 0.0 | 2 ± 0.5 | 4 ± 0.4 | Putative pentatricopeptide repeat protein | Defence against biotic stress (Pseudomonas syringae pv. tomato and Botrytis cinerea) | [71,72] |
| FvH4_6g18970.1 | 0 ± 0.0 | 2 ± 0.3 | 2 ± 0.1 | Probable disease resistance protein At5g66900 | Unknown | _ |
| FvH4_1g15330.1 | 0 ± 0.0 | 1 ± 0.2 | 1 ± 0.2 | Probable disease resistance protein At5g66910 | Unknown | _ |
| FvH4_5g31070.1 | 0 ± 0.0 | 1 ± 0.3 | 2 ± 0.1 | Putative pentatricopeptide repeat protein | Defence against biotic stress (Pseudomonas syringae pv. tomato and Botrytis cinerea) and abiotic stress | [71,72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gogoi, A.; Lysøe, E.; Eikemo, H.; Stensvand, A.; Davik, J.; Brurberg, M.B. Comparative Transcriptome Analysis Reveals Novel Candidate Resistance Genes Involved in Defence against Phytophthora cactorum in Strawberry. Int. J. Mol. Sci. 2023, 24, 10851. https://doi.org/10.3390/ijms241310851
Gogoi A, Lysøe E, Eikemo H, Stensvand A, Davik J, Brurberg MB. Comparative Transcriptome Analysis Reveals Novel Candidate Resistance Genes Involved in Defence against Phytophthora cactorum in Strawberry. International Journal of Molecular Sciences. 2023; 24(13):10851. https://doi.org/10.3390/ijms241310851
Chicago/Turabian StyleGogoi, Anupam, Erik Lysøe, Håvard Eikemo, Arne Stensvand, Jahn Davik, and May Bente Brurberg. 2023. "Comparative Transcriptome Analysis Reveals Novel Candidate Resistance Genes Involved in Defence against Phytophthora cactorum in Strawberry" International Journal of Molecular Sciences 24, no. 13: 10851. https://doi.org/10.3390/ijms241310851
APA StyleGogoi, A., Lysøe, E., Eikemo, H., Stensvand, A., Davik, J., & Brurberg, M. B. (2023). Comparative Transcriptome Analysis Reveals Novel Candidate Resistance Genes Involved in Defence against Phytophthora cactorum in Strawberry. International Journal of Molecular Sciences, 24(13), 10851. https://doi.org/10.3390/ijms241310851