
Yeast Lipid Produced through Glycerol Conversions and Its Use for Enzymatic Synthesis of Amino Acid-Based Biosurfactants

 ,
, 
 and
and
Abstract
1. Introduction
2. Results and Discussion
2.1. Production and Characterization of Microbial Lipids
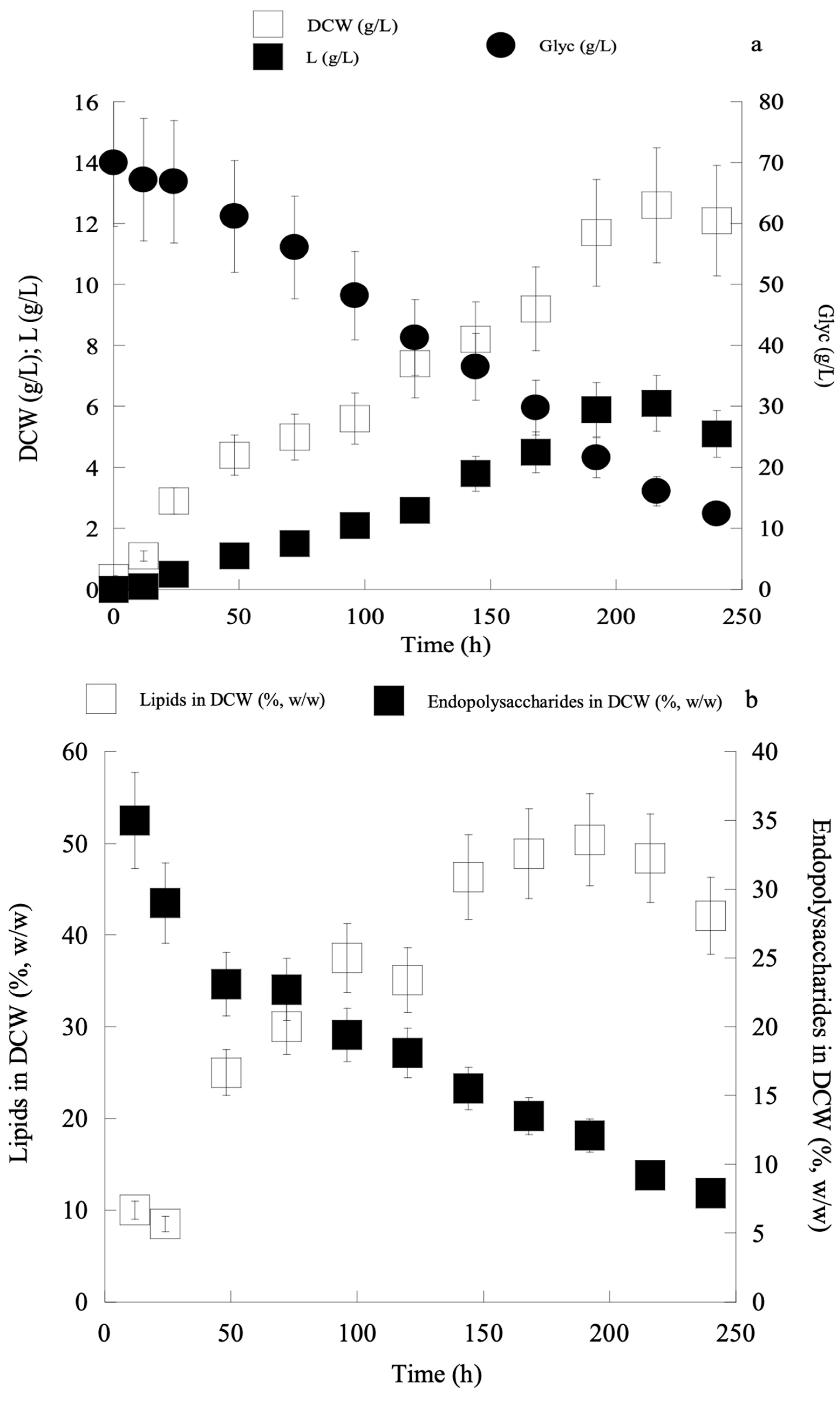
2.1.1. Lipid Production by Yeast Cultures Growing on Biodiesel-Derived Glycerol
2.1.2. Yeast Lipid Analysis
2.2. N-Acylation Activity and Regioselectivity of CALB and Aminoacylases
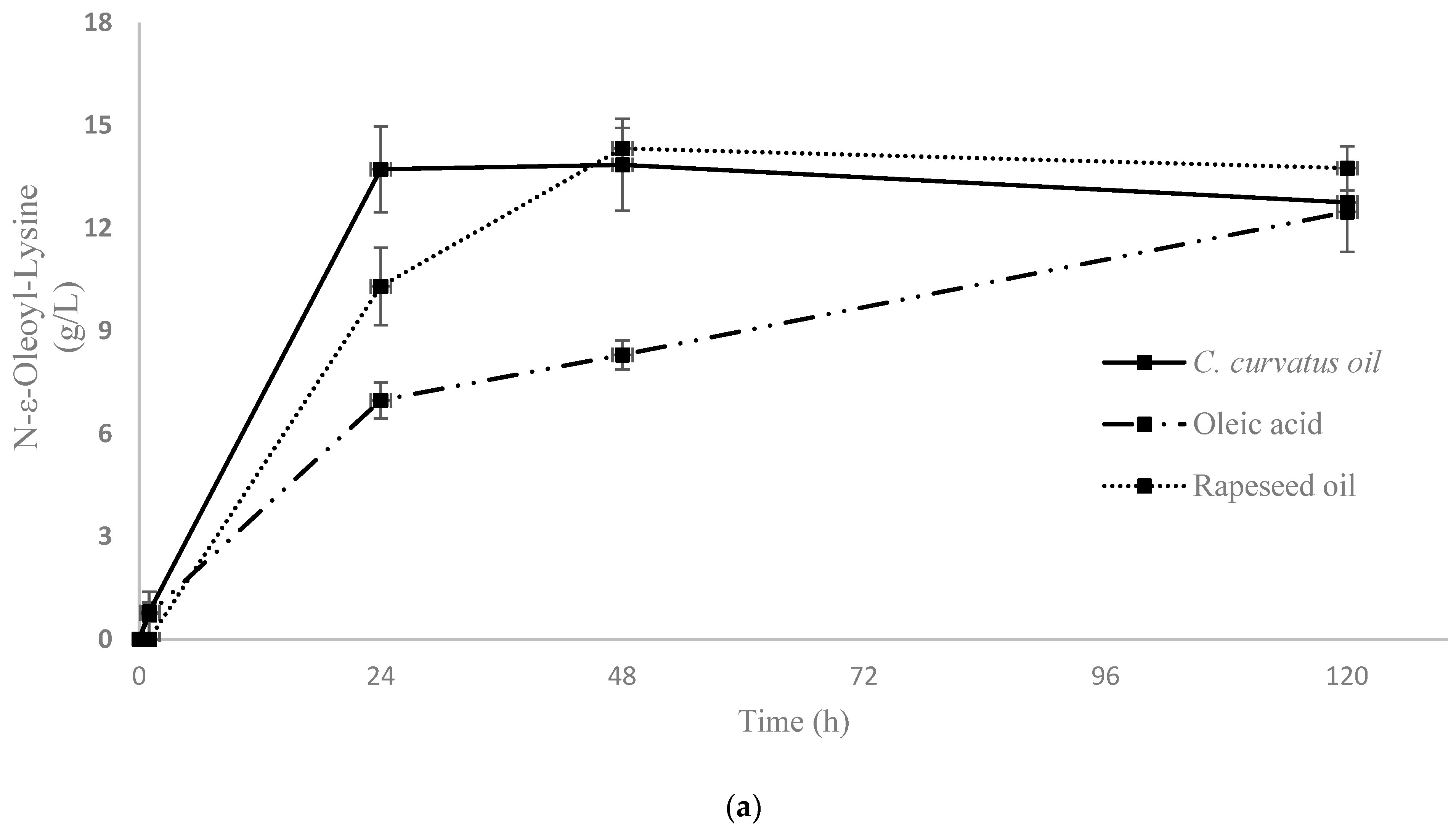
2.2.1. N-Acylation of Lysine Using Free and Oil-Derived AD by CALB in Non-Aqueous Medium
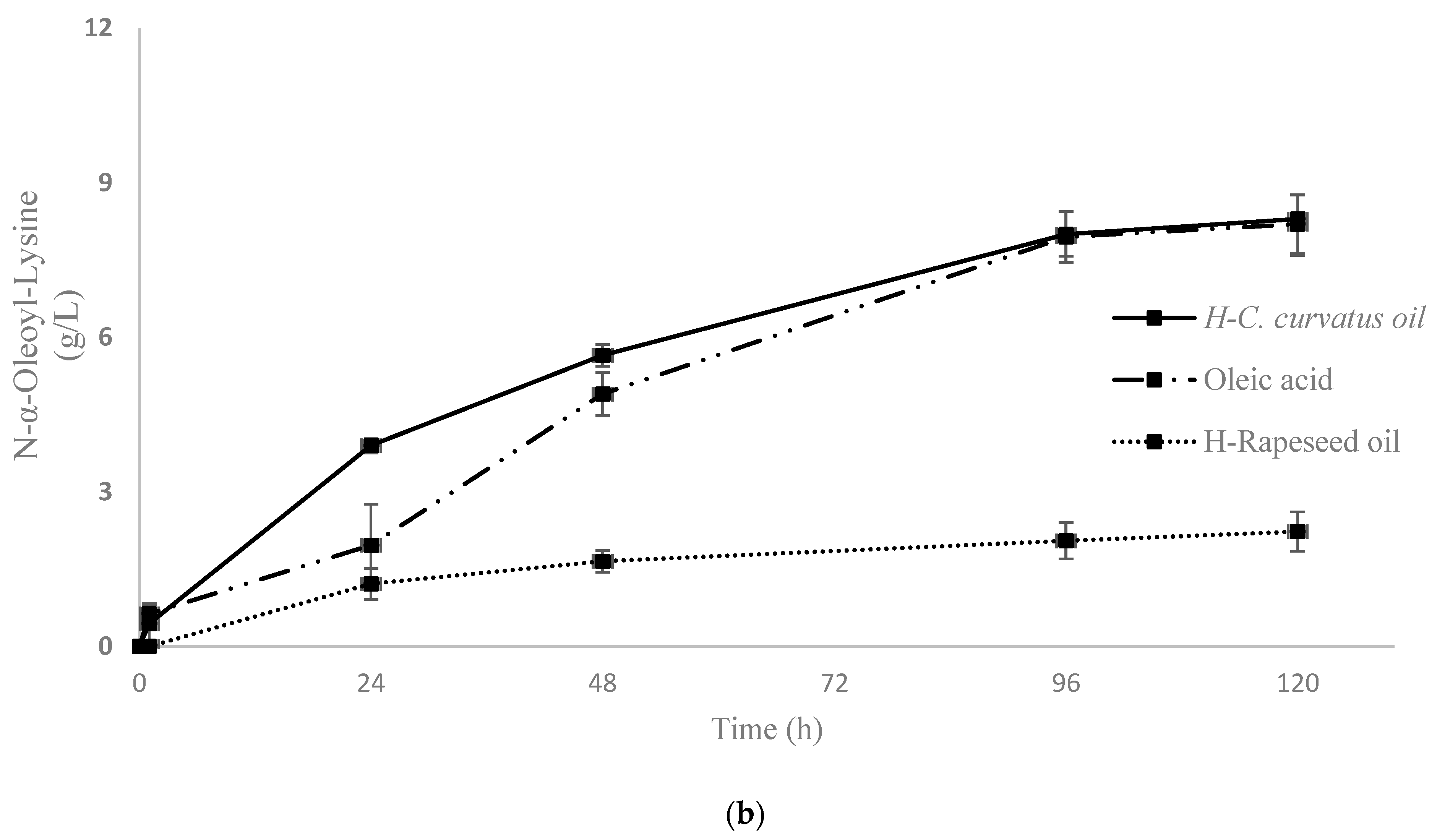
2.2.2. N-Acylation of Lysine Using Free and Oil-Derived AD by Aminoacylases, in Aqueous Medium
2.3. Substrate Specificity of CALB and Aminoacylases towards FAs from Oils or Mixtures
3. Materials and Methods
3.1. Microorganisms, Enzymes and Chemicals
3.2. Microbial Lipid Production
3.3. Production of Aminoacylases from Streptomyces ambofaciens
3.4. Acylation Reaction
3.4.1. N-Acylation Reaction Catalysed by CALB
3.4.2. N-Acylation Reaction Catalysed by Aminoacylases
3.5. Analytical Methods
3.5.1. Microbial Lipid Production
3.5.2. Protein Concentration Measurement
3.5.3. Quantitative Analysis
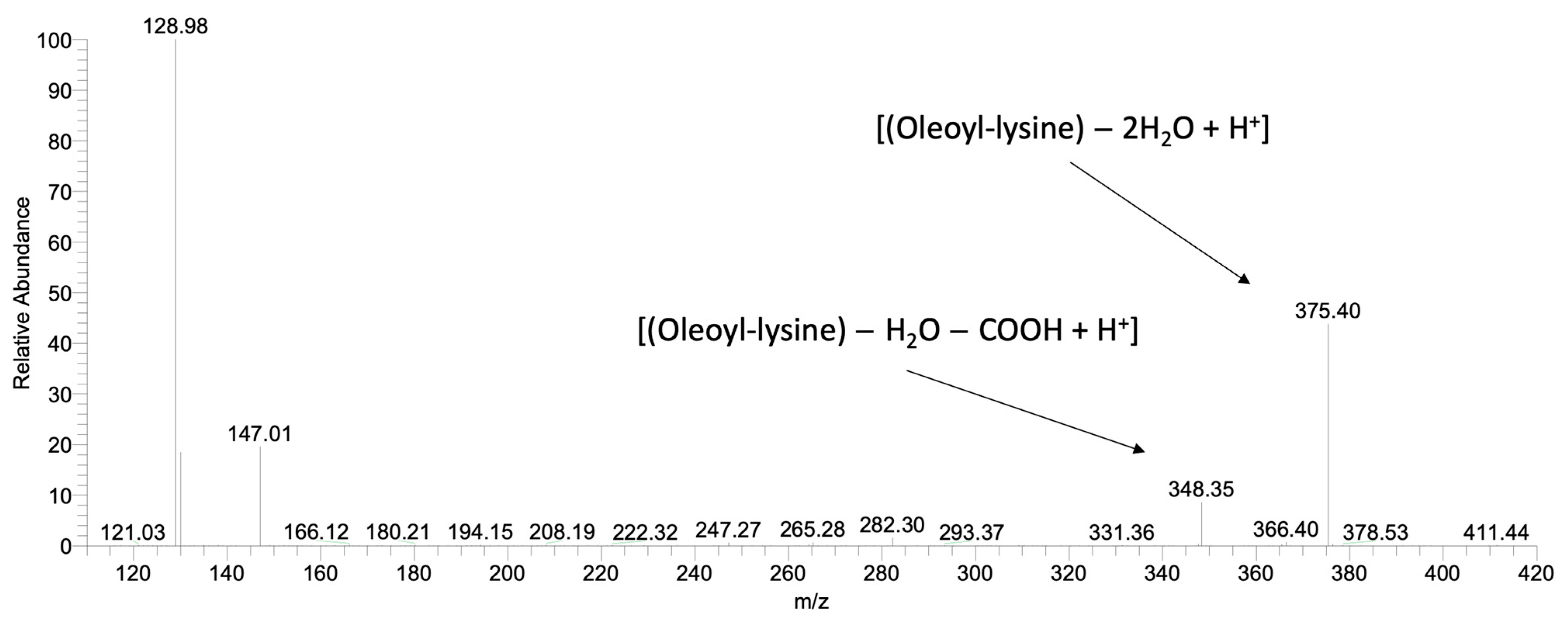
3.5.4. Qualitative and Semi-Quantitative Analysis by Mass Spectrometry
3.6. Specificity Factor (Sf) Formula
3.7. Tailor-Made FA mixtures
3.8. Abbreviations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ivanković, T.; Hrenović, J. Surfactants in the Environment. Arch. Hig. Rada. Toksikol. 2010, 61, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Infante, M.R.; Perez, L.; Pinazo, A.; Clapes, P.; Moran, M.C.; Angelet, M.; Garcia, M.T.; Vinardell, M.P. Amino acid-based surfactants. C. R. Chim. 2004, 7, 583–592. [Google Scholar] [CrossRef]
- Fortune Business Insights. Biosurfactants Market Size, Share & COVID-19 Impact Analysis, by Type (Glycolipids, Lipopeptides, Phospholipids, Polymeric, and Others) by Application (Household Cleaners, Industrial and Institutional Cleaners, Food Processing, Oil & Petroleum, Personal Care, Agrochemicals, and Others), and Regional Forecast, (2021–2028); Fortune Business Insights: Pune, India, 2021; Available online: https://www.fortunebusinessinsights.com/biosurfactants-market-102761 (accessed on 15 February 2022).
- Soo, E.L.; Salleh, A.B.; Basri, M.; Rahman, R.N.Z.A.; Kamaruddin, K. Response surface methodological study on lipase-catalyzed synthesis of amino acid surfactants. Process Biochem. 2004, 39, 1511–1518. [Google Scholar] [CrossRef]
- George, A.; Modi, J.; Jain, N.; Bahadur, P. A comparative study on the surface activity and micellar behavior of some N-acylamino acid based surfactants. Ind. J. Chem. 1998, 37A, 985–992. [Google Scholar]
- Rondel, C.; Alric, I.; Mouloungui, Z.; Blanco, J.F.; Silvestre, F. Synthesis and properties of lipoamino acid–fatty acid mixtures: Influence of the amphiphilic structure. J. Surf. Det. 2009, 12, 269–275. [Google Scholar] [CrossRef]
- Clapés, P.; Infante, M.R. Amino acid-based surfactants: Enzymatic synthesis, properties and potential applications. Biocatal. Biotransform. 2002, 20, 215–233. [Google Scholar] [CrossRef]
- Chandra, N.; Tyagi, V.K. Synthesis, properties, and applications of amino acids based surfactants: A review. J. Dispers. Sci. Technol. 2013, 34, 800–808. [Google Scholar] [CrossRef]
- Babizhayev, M.A. Biological activities of the natural imidazole-containing peptidomimetics n-acetylcarnosine, carcinine and L-carnosine in ophthalmic and skin care products. Life Sci. 2006, 78, 2343–2357. [Google Scholar] [CrossRef]
- Khandelia, H.; Ipsen, J.H.; Mouritsen, O.G. The impact of peptides on lipid membranes. Biochim. Biophys. Acta Biomembr. 2008, 1778, 1528–1536. [Google Scholar] [CrossRef]
- Hanuš, L.; Shohami, E.; Bab, I.; Mechoulam, R. N-Acyl amino acids and their impact on biological processes. Biofactors 2014, 40, 381–388. [Google Scholar] [CrossRef]
- Asada, H.; Douen, T.; Mizokoshi, Y.; Fujita, T.; Murakami, M.; Yamamoto, A.; Muranishi, S. Stability of acyl derivatives of insulin in the small intestine: Relative importance of insulin association characteristics in aqueous solution. Pharm. Res. 1994, 11, 1115–1120. [Google Scholar] [CrossRef]
- Furutani, T.; Furui, M.; Ooshima, H.; Kato, J. N-acylation of [beta]-amino alcohol by acyl migration following enzyme-catalyzed esterification. Enzyme Microb. Technol. 1996, 19, 578–584. [Google Scholar] [CrossRef]
- Zhang, X.; Adachi, S.; Watanabe, Y.; Matsuno, R. Lipase-catalyzed synthesis of O- lauroyl L-serinamide and O-lauroyl L-threoninamide. Food Res. Int. 2005, 38, 297–300. [Google Scholar] [CrossRef]
- Husson, E.; Humeau, C.; Blanchard, F.; Framboisier, X.; Marc, I.; Chevalot, I. Chemo- selectivity of the N,O-Enzymatic acylation in organic media and in ionic liquids. J. Mol. Catal. B Enzym. 2008, 55, 110–117. [Google Scholar] [CrossRef]
- Ratledge, C.; Cohen, Z. Microbial and algal oils: Do they have a future for biodiesel or as commodity oils? Lipid Technol. 2008, 20, 155–160. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Sources of microbial oils with emphasis to Mortierella (Umbelopsis) isabellina fungus. World J. Microbiol. Biotechnol. 2019, 35, 63. [Google Scholar] [CrossRef]
- Shah, A.M.; Yang, W.; Mohamed, H.; Zhang, Y.; Song, Y. Microbes: A hidden treasure of polyunsaturated fatty acids. Front. Nutr. 2022, 9, 827837. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Li, B.; Huang, B.C.; Wang, F.B.; Zhang, Y.Q.; Zhao, S.G.; Li, M.; Wang, H.Y.; Yu, X.J.; Liu, X.Y.; et al. Production, biosynthesis, and commercial applications of fatty acids from oleaginous fungi. Front. Nutr. 2022, 9, 873657. [Google Scholar] [CrossRef]
- Papadaki, A.; Kopsahelis, N.; Mallouchos, A.; Mandala, I.; Koutinas, A.A. Bioprocess development for the production of novel oleogels from soybean and microbial oils. Food Res. Int. 2019, 126, 108684. [Google Scholar] [CrossRef]
- Woodley, J.M.; Breuer, M.; Mink, D. A future perspective on the role of bioprocesses for chemical production. ChERD 2013, 91, 2029–2036. [Google Scholar]
- Schmid, A.; Dordick, J.S.; Hauer, B.; Kiener, A.; Wubbolts, M.; Witholt, B. Industrial biocatalysis today and tomorrow. Nature 2011, 409, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A.; Woodley, J.M. Role of biocatalysis in sustainable chemistry. Chem. Rev. 2018, 118, 801–838. [Google Scholar] [CrossRef] [PubMed]
- Wender, P.A.; Handy, S.T.; Wright, D.L. Towards the ideal synthesis. Chem. Ind. 1997, 19, 765–769. [Google Scholar]
- Zaks, A.; Klibanov, A. Enzyme-catalyzed processes in organic-solvents. Proc. Natl. Acad. Sci. USA 1985, 82, 3192–3196. [Google Scholar] [CrossRef]
- Wang, X.; Ohlin, C.A.; Lu, Q.; Fei, Z.; Hu, J.; Dyson, P.J. Cytotoxicity of ionic liquids and precursor compounds towards human cell line HeLa. Green Chem. 2007, 9, 1191–1197. [Google Scholar] [CrossRef]
- Husson, E.; Humeau, C.; Paris, C.; Vanderesse, R.; Framboisier, X.; Marc, I.; Chevalot, I. Enzymatic acylation of polar dipeptides: Influence of reaction media and molecular environment of functional groups. Process Biochem. 2008, 44, 428–434. [Google Scholar] [CrossRef]
- Koreishi, M.; Zhang, D.; Imanaka, H.; Imamura, K.; Adachi, S.; Matsuno, R.; Nakanishi, K. A novel acylase from Streptomyces mobaraensis that efficiently catalyzes hydrolysis/synthesis of capsaicins as well as N-Acyl-l-amino acids and N- Acyl-peptides. J. Agric. Food Chem. 2006, 54, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Koreishi, M.; Kawasaki, R.; Imanaka, J.; Imamura, K.; Takakura, Y.; Nakanishi, K. Efficient Nɛ-lauroyl-l-lysine production by recombinant ɛ-lysine acylase from Streptomyces mobaraensis. J. Biotechnol. 2009, 141, 160–165. [Google Scholar] [CrossRef]
- Dettori, L.; Ferrari, F.; Framboisier, X.; Paris, C.; Guiavarc’h, Y.; Hôtel, L.; Aymes, A.; Leblond, P.; Humeau, C.; Kapel, R.; et al. An aminoacylase activity from Streptomyces ambofaciens catalyzes the acylation of lysine on α-position and peptides on N-terminal position. Eng. Life Sci. 2017, 18, 589–599. [Google Scholar] [CrossRef]
- Bourkaib, M.C.; Delaunay, S.; Framboisier, X.; Hôtel, L.; Aigle, B.; Humeau, C.; Guiavarc’h, Y.; Chevalot, I. N-acylation of L-amino acids in aqueous media: Evaluation of the catalytic performances of Streptomyces ambofaciens aminoacylases. Enzyme Microb. Technol. 2020, 137, 109536. [Google Scholar] [CrossRef] [PubMed]
- Pérez, L.; Pinazo, A.; García, M.T.; Lozano, M.; Manresa, A.; Angelet, M.; Vinardell, M.P.; Mitjans, M.; Pons, R.; Infante, M.R. Cationic surfactants from lysine: Synthesis, micellization and biological evaluation. Eur. J. Med. Chem. 2009, 44, 1884–1892. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Fakas, S.; Fick, M.; Chevalot, I.; Galiotou-Panayotou, M.; Komaitis, M.; Marc, I.; Aggelis, G. Biotechnological valorization of raw glycerol discharged after bio-diesel (fatty acid methyl esters) manufacturing process: Production of 1,3-propanediol, citric acid and single cell oil. Biomass Bioenergy 2008, 32, 60–71. [Google Scholar] [CrossRef]
- Sarantou, S.; Stoforos, N.G.; Kalantzi, O.; Papanikolaou, S. Biotechnological valorization of biodiesel-derived glycerol: Trials with the non-conventional yeasts Yarrowia lipolytica and Rhodosporidium sp. Carbon Resour. Convers. 2021, 4, 61–75. [Google Scholar] [CrossRef]
- Filippousi, R.; Tsouko, E.; Mordini, K.; Ladakis, D.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Sustainable arabitol production by a newly isolated Debaryomyces prosopidis strain cultivated on biodiesel-derived glycerol. Carbon Resour. Convers. 2022, 5, 92–99. [Google Scholar] [CrossRef]
- Rzechonek, D.A.; Dobrowolski, A.; Rymowicz, W.; Mirończuk, A.M. Recent advances in biological production of erythritol. Crit. Rev. Biotechnol. 2018, 38, 620–633. [Google Scholar] [CrossRef]
- Erian, A.M.; Sauer, M. Utilizing yeasts for the conversion of renewable feedstocks to sugar alcohols—A review. Biores. Technol. 2022, 346, 126296. [Google Scholar] [CrossRef]
- Tchakouteu, S.S.; Chatzifragkou, A.; Kalantzi, O.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Oleaginous yeast Cryptococcus curvatus exhibits interplay between biosynthesis of intracellular sugars and lipids. Eur. J. Lipid Sci. Technol. 2015, 117, 657–672. [Google Scholar] [CrossRef]
- Diamantopoulou, P.; Filippousi, R.; Antoniou, D.; Varfi, E.; Xenopoulos, E.; Sarris, D.; Papanikolaou, S. Production of added-value microbial metabolites during growth of yeast strains on media composed of biodiesel-derived crude glycerol and glycerol/xylose blends. FEMS Microbiol. Lett. 2020, 367, fnaa063. [Google Scholar] [CrossRef]
- Evans, C.T.; Ratledge, C. Influence of nitrogen metabolism on lipid accumulation by Rhodosporidium toruloides CBS 14. J. Gen. Microbiol. 1984, 130, 1705–1710. [Google Scholar] [CrossRef][Green Version]
- Vasilakis, G.; Karayannis, D.; Massouras, T.; Politis, I.; Papanikolaou, S. Biotechnological conversions of Mizithra second cheese whey by wild-type non-conventional yeast strains: Production of yeast cell biomass, single-cell oil and polysaccharides. Appl. Sci. 2022, 12, 11471. [Google Scholar] [CrossRef]
- Meesters, P.A.E.P.; Huijiberts, G.N.M.; Eggink, G. Hight-cell-density cultivation of the lipid accumulating yeast Cryptococcus curvatus using glycerol as a carbon source. Appl. Microbiol. Biotechnol. 1996, 45, 575–579. [Google Scholar] [CrossRef]
- Bellou, S.; Baeshen, M.N.; Elazzazy, A.M.; Aggeli, D.; Sayegh, F.; Aggelis, G. Microalgal lipids biochemistry and biotechnological perspectives. Biotechnol. Adv. 2014, 32, 476–1493. [Google Scholar] [CrossRef] [PubMed]
- Bellou, S.; Triantaphyllidou, I.E.; Aggeli, D.; Elazzazy, A.M.; Baeshen, M.N.; Aggelis, G. Microbial oils as food additives: Recent approaches for improving microbial oil production and its polyunsaturated fatty acid content. Curr. Opin. Biotechnol. 2016, 37, 24–35. [Google Scholar] [CrossRef]
- Dettori, L.; Jelsch, C.; Guiavarc’h, Y.; Delaunay, S.; Framboisier, X.; Chevalot, I.; Humeau, C. Molecular rules for selectivity in lipase-catalysed acylation of lysine. Process Biochem. 2018, 74, 50–60. [Google Scholar] [CrossRef]
- Τryfinopoulou, P.; Tsakalidou, E.; Nychas, G.J.E. Characterization of Pseudomonas spp. associated with spoilage of gilt-head sea-bream stored under various conditions. Appl. Environ. Microbiol. 2002, 68, 65–72. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Yarrowia lipolytica as a potential producer of citric acid from raw glycerol. J. Appl. Microbiol. 2002, 92, 737–744. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Time (h) | Glyccons (g/L) | X (g/L) | L (g/L) | YL/X (g/g) | YIPS/X (g/g) | |
|---|---|---|---|---|---|---|---|
| Metschnikowia | 2 | 146 | 35.5 | 11.0 | 0.61 | 0.06 | 0.18 |
| pulcherrima FMCC Y2 | 1, 3, 4 | 288 | 47.9 | 16.5 | 0.85 | 0.05 | 0.43 |
| Cryptococcus | 4 | 162 | 36.2 | 15.5 | 0.26 | 0.02 | 0.25 |
| curvatus | 1 | 186 | 40.7 | 17.6 | 0.36 | 0.02 | 0.19 |
| NRRL Y-1511 | 2, 3 | 211 | 45.5 | 16.6 | 0.95 | 0.06 | 0.14 |
| Cryptococcus | 4 | 47 | 14.8 | 7.9 | 0.91 | 0.11 | 0.41 |
| curvatus | 1 | 119 | 30.5 | 15.9 | 3.78 | 0.24 | 0.32 |
| ATCC 20509 | 2, 3 | 201 | 47.9 | 15.1 | 4.98 | 0.33 | 0.19 |
| Rhodosporidium | 4 | 48 | 11.9 | 7.2 | 0.74 | 0.10 | 0.31 |
| toruloides | 1 | 191 | 43.2 | 16.4 | 3.18 | 0.19 | 0.17 |
| NRRL Y-6984 | 2, 3 | 215 | 43.2 | 16.0 | 3.23 | 0.20 | 0.16 |
| Rhodotorula sp. | 2, 3 | 210 | 31.9 | 15.8 | 2.60 | 0.16 | 0.36 |
| FMCC Y76 | 1, 4 | 255 | 36.9 | 17.2 | 1.91 | 0.11 | 0.39 |
| Nitrogen Source | Time (h) | Glyccons (g/L) | X (g/L) | L (g/L) | YL/X (g/g) | YIPS/X (g/g) | |
|---|---|---|---|---|---|---|---|
| Ammonium | 4 | 71 | 14.8 | 13.6 | 1.69 | 0.12 | 0.27 |
| sulfate | 2, 3 | 144 | 30.5 | 18.5 | 3.35 | 0.18 | 0.21 |
| (1.34 g/L) | 1 | 192 | 44.8 | 20.0 | 2.90 | 0.15 | 0.11 |
| Peptone | 4 | 47 | 14.8 | 7.9 | 0.91 | 0.11 | 0.41 |
| (2.00 g/L) | 1 | 119 | 30.5 | 15.9 | 3.78 | 0.24 | 0.32 |
| 2, 3 | 201 | 47.9 | 15.1 | 4.98 | 0.33 | 0.19 | |
| Potassium | 4 | 47 | 8.3 | 6.0 | 0.79 | 0.13 | 0.40 |
| nitrate | 2, 3 | 192 | 29.1 | 11.6 | 4.52 | 0.39 | 0.09 |
| (2.04 g/L) | 1 | 206 | 29.7 | 12.0 | 3.92 | 0.33 | 0.08 |
| Urea | 3 | 46 | 8.8 | 6.8 | 1.07 | 0.16 | 0.26 |
| (0.61 g/L) | 4 | 96 | 18.3 | 11.4 | 1.01 | 0.09 | 0.44 |
| 1, 2 | 170 | 24.7 | 14.8 | 1.90 | 0.12 | 0.36 | |
| Yeast | 4 | 72 | 12.6 | 14.0 | 2.17 | 0.16 | 0.36 |
| extract | 2, 3 | 170 | 29.4 | 21.2 | 3.16 | 0.25 | 0.20 |
| (2.54 g/L) | 1 | 216 | 31.1 | 21.4 | 3.08 | 0.24 | 0.18 |
| Glyc0 (g/L) | Initial C/N (moles/moles) | Time (h) | Glycons (g/L) | X (g/L) | L (g/L) | YL/X (g/g) | YIPS/X (g/g) | |
|---|---|---|---|---|---|---|---|---|
| 4 | 45 | 13.9 | 5.1 | 0.88 | 0.17 | 0.39 | ||
| ≈30 g/L | ≈34 | 2, 3 | 120 | 29.5 | 14.0 | 2.71 | 0.19 | 0.31 |
| 1 | 155 | 34.0 | 16.1 | 1.95 | 0.12 | 0.27 | ||
| 4 | 47 | 14.8 | 7.9 | 0.91 | 0.11 | 0.41 | ||
| ≈50 g/L | ≈57 | 1 | 119 | 30.5 | 15.9 | 3.78 | 0.24 | 0.32 |
| 2, 3 | 201 | 47.9 | 15.1 | 4.98 | 0.33 | 0.19 | ||
| 4 | 12 | 2.9 | 1.1 | 0.11 | 0.10 | 0.35 | ||
| ≈70 g/L | ≈80 | 2 | 192 | 48.4 | 11.7 | 5.90 | 0.50 | 0.12 |
| 1, 3 | 216 | 57.4 | 12.6 | 6.10 | 0.48 | 0.09 |
| Fatty Acid Composition of Yeast Lipids (%, w/w) | ||||||
|---|---|---|---|---|---|---|
| Yeast Strain | C16:0 | C16:1 | C18:0 | C18:1 | C18:2 | C18:3 |
| Metschnikowia pulcherrima FMCC Y2 | 17.7 | 7.7 | Tr. | 59.4 | 14.5 | Tr. |
| Cryptococcus curvatus NRRL Y-1511 | 33.8 | 5.6 | 0.7 | 45.2 | 11.7 | 3.0 |
| Cryptococcus curvatus ATCC 20509 | 26.0 | 1.2 | 3.9 | 57.2 | 11.2 | Tr. |
| Rhodosporidium toruloides NRRL Y-6984 | 33.9 | 0.5 | 1.2 | 56.7 | 7.8 | Τr. |
| Rhodotorula sp. FMCC Y76 | 21.0 | 0.5 | 7.5 | 51.0 | 12.1 | 1.0 |
| Fatty Acid Composition of Yeast Lipids (%, w/w) | ||||||
|---|---|---|---|---|---|---|
| Nitrogen Source | C16:0 | C16:1 | C18:0 | C18:1 | C18:2 | C18:3 |
| Ammonium sulfate | 37.2 | Tr. | Tr. | 49.4 | 13.0 | Tr. |
| Potassium nitrate | 35.6 | 6.8 | Tr. | 54.5 | 3.2 | Tr. |
| Peptone | 26.0 | 1.2 | 3.9 | 57.2 | 11.2 | Tr. |
| Urea | 34.3 | 3.6 | Tr. | 47.4 | 8.4 | Τr. |
| Yeast extract | 39.2 | 4.2 | Tr. | 54.9 | 8.8 | 0.5 |
| Fatty Acid Composition of Yeast Lipids (%, w/w) | ||||||
|---|---|---|---|---|---|---|
| Glyc0 (g/L) | C16:0 | C16:1 | C18:0 | C18:1 | C18:2 | C18:3 |
| ≈30 g/L | 30.2 | 3.5 | 4.2 | 48.7 | 12.7 | Tr. |
| ≈50 g/L | 26.0 | 1.2 | 3.9 | 57.2 | 11.2 | Tr. |
| ≈70 g/L | 27.0 | 1.5 | 3.0 | 45.0 | 9.5 | 2.0 |
| Major Fatty Acid Composition (%, w/w) | |||
|---|---|---|---|
| C16:0 | C18:1 | C18:2 | |
| C. curvatus oil | 27.0 | 45.0 | 9.5 |
| Rapeseed oil | 4.8 | 60.0 | 20.0 |
| Iso-mix | 33.3 | 33.3 | 33.3 |
| ε-Palmitoyl-Lysine | ε-Oleoyl-Lysine | ε-Linoleoyl-Lysine | ||||
|---|---|---|---|---|---|---|
| Product (Mass Area) | Product (%) | Product (Mass Area) | Product (%) | Product (Mass Area) | Product (%) | |
| C. curvatus oil | 2.8 | 6.8 | 27.5 | 66.3 | 11.2 | 27 |
| Rapeseed oil | 1 | 2.8 | 18.8 | 51.2 | 16.9 | 46 |
| Iso-mix | 6 | 20.3 | 10.1 | 34.4 | 13.3 | 45.3 |
| α-Palmitoyl-Lysine | α-Oleoyl-Lysine | α-Linoleoyl-Lysine | ||||
|---|---|---|---|---|---|---|
| Product (Mass Area) | Product (%) | Product (Mass Area) | Product (%) | Product (Mass Area) | Product (%) | |
| H-C. curvatus oil | 26.3 | 59.2 | 14.9 | 33.6 | 3.17 | 7.1 |
| TM-C. curvatus | 27.1 | 53.9 | 19.2 | 38.2 | 4 | 7.9 |
| H-Rapeseed oil | 3.4 | 14.7 | 16.8 | 73.1 | 2.8 | 12.2 |
| TM-Rapeseed | 3.9 | 13.1 | 21.6 | 72.2 | 4.4 | 14.6 |
| Iso-mix | 28.9 | 65.8 | 10.3 | 23.5 | 4.7 | 10.7 |
| C16:0 (Sf) | C18:1 (Sf) | C18:2 (Sf) | |
|---|---|---|---|
| CALB | |||
| C. curvatus oil | 0.25 | 1.47 | 2.84 |
| Rapeseed oil | 0.57 | 0.85 | 2.30 |
| Iso-mix | 0.61 | 1.03 | 1.36 |
| Aminoacylases | |||
| H-C. curvatus oil | 2.19 | 0.75 | 0.75 |
| TM-C. curvatus | 2.00 | 0.85 | 0.84 |
| H-Rapeseed oil | 3.06 | 1.22 | 0.61 |
| TM-Rapeseed | 2.74 | 1.20 | 0.73 |
| Iso-mix | 1.98 | 0.71 | 0.32 |
| Abbreviation | Meaning |
|---|---|
| Cu | Oil derived from Cryptococcus curvatus ATCC 20509 culture |
| H-Cu or H-C. curvatus oil | Hydrolysed Cryptococcus curvatus oil |
| TMCu or TM-C. curvatus | Tailor-made solution of the major fatty acids of Cryptococcus curvatus oil |
| Rap | Rapeseed oil |
| H-Rap or H-Rapeseed oil | Hydrolysed rapeseed oil |
| TMRap or TM-Rapeseed | Tailor-made mixture of the major fatty acids of rapeseed oil |
| Iso-mix | Tailor-made solution containing 1/3 palmitic acid, 1/3 oleic acid and 1/3 palmitic acid |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karayannis, D.; Papanikolaou, S.; Vatistas, C.; Paris, C.; Chevalot, I. Yeast Lipid Produced through Glycerol Conversions and Its Use for Enzymatic Synthesis of Amino Acid-Based Biosurfactants. Int. J. Mol. Sci. 2023, 24, 714. https://doi.org/10.3390/ijms24010714
Karayannis D, Papanikolaou S, Vatistas C, Paris C, Chevalot I. Yeast Lipid Produced through Glycerol Conversions and Its Use for Enzymatic Synthesis of Amino Acid-Based Biosurfactants. International Journal of Molecular Sciences. 2023; 24(1):714. https://doi.org/10.3390/ijms24010714
Chicago/Turabian StyleKarayannis, Dimitris, Seraphim Papanikolaou, Christos Vatistas, Cédric Paris, and Isabelle Chevalot. 2023. "Yeast Lipid Produced through Glycerol Conversions and Its Use for Enzymatic Synthesis of Amino Acid-Based Biosurfactants" International Journal of Molecular Sciences 24, no. 1: 714. https://doi.org/10.3390/ijms24010714
APA StyleKarayannis, D., Papanikolaou, S., Vatistas, C., Paris, C., & Chevalot, I. (2023). Yeast Lipid Produced through Glycerol Conversions and Its Use for Enzymatic Synthesis of Amino Acid-Based Biosurfactants. International Journal of Molecular Sciences, 24(1), 714. https://doi.org/10.3390/ijms24010714