
Alpha-Lipoic Acid and Its Enantiomers Prevent Methemoglobin Formation and DNA Damage Induced by Dapsone Hydroxylamine: Molecular Mechanism and Antioxidant Action

 ,
,  , , and
, , and
Abstract

1. Introduction
2. Results
2.1. Effect of Racemic Alpha-Lipoic Acid (ALA) on Methemoglobin Formation Induced by DDS-NOH
2.2. Comparative Effect of Pre and Post Treatment with ALA or Methylene Blue (MB) on Methemoglobin Formation Induced by DDS-NHOH
2.3. Effect of Pretreatment with ALA on Reactive Oxygen Species (ROS) Generation Induced by DDS-NOH
2.4. Effect of Pretreatment with ALA on CAT and SOD Activities and GSH Production on In Vitro DDS-NOH Model
2.5. Effect of ALA on DNA Damage Induced by DDS-NOH
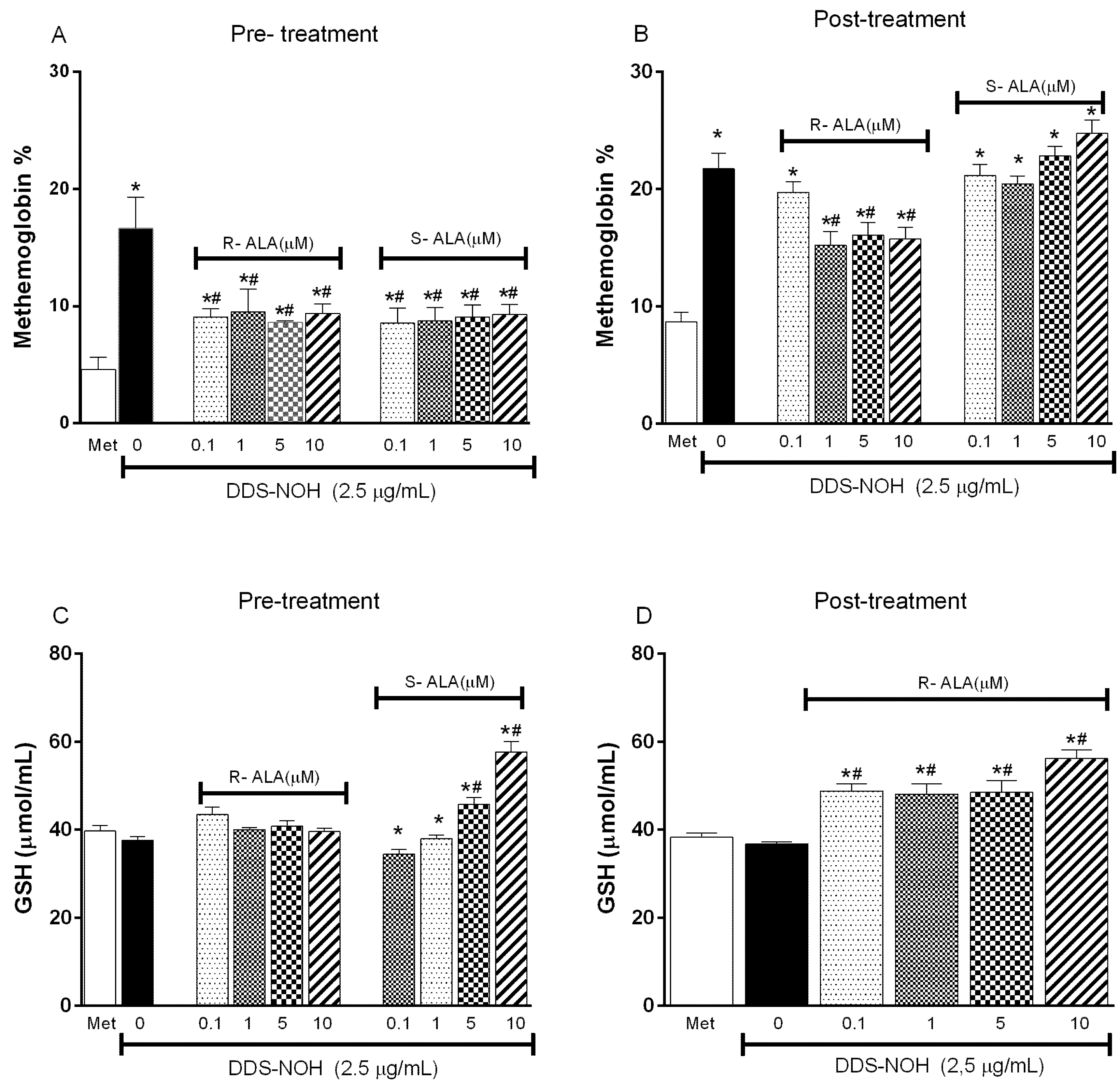
2.6. Effect of the Pre- or Post-Treatment with Enantiomers (R-ALA or S-ALA) on Methemoglobin Formation and GSH Levels Induced by DDS-NOH
2.7. Effect of Pretreatment with R-ALA and S-ALA on Oxidative Parameters Induced by DDS-NOH
2.8. Effect of Pretreatment with R-ALA and S-ALA on DNA Damage Induced by DDS-NOH
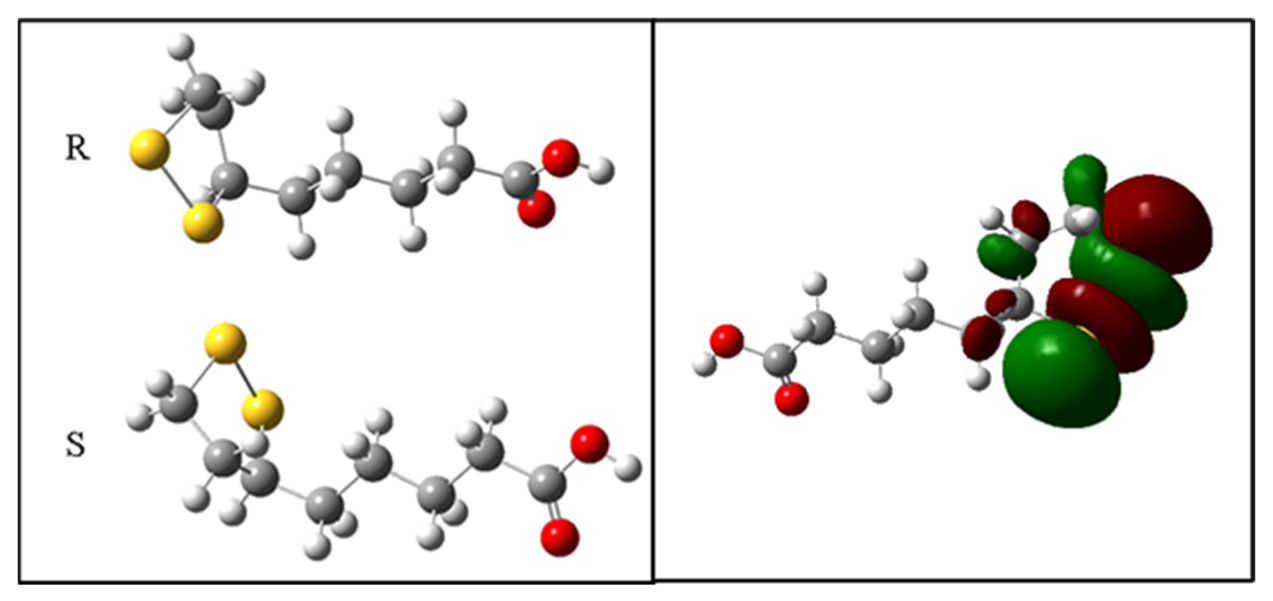
2.9. Theoretical Antioxidant Mechanism of R-ALA and S-ALA Isomers
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of ALA, ALA Enantiomers and DDS-NOH Solutions
4.3. Ethics Statement
4.4. Preparation of Erythrocyte Suspensions
4.5. Pretreatment of Erythrocytes with ALA, ALA Enantiomers, or MB and Treatment with DDS-NOH
4.6. Post-Treatment of Erythrocytes with ALA, ALA Enantiomers, or MB and Treatment with DDS-NOH
4.7. Determination of Methemoglobin Content
4.8. Cell Culture and Sample Preparations to Comet Assay
4.9. DNA Damage Using the Comet Assay
4.10. Measurement of Intracellular Reactive Oxygen Species (ROS)
4.11. Reduced Glutathione Activity
4.12. Superoxide Dismutase (SOD) Activity
4.13. Trolox Equivalent Antioxidant Capacity
4.14. Determination of Lipid Peroxidation (MDA)
4.15. Data Analysis
4.16. Molecular Modeling
4.16.1. Preparation of the Ligands and the Receptor
4.16.2. Molecular Docking
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coleman, M.D.; Jacobus, D.P. Reduction of Dapsone Hydroxylamine to Dapsone during Methaemoglobin Formation in Human Erythrocytes in Vitro—II. Biochem. Pharmacol. 1993, 46, 1363–1368. [Google Scholar] [CrossRef]
- Thuong-Nguyen, V.; Kadunce, D.P.; Hendrix, J.D.; Ray Gammon, W.; Zone, J.J. Inhibition of Neutrophil Adherence to Antibody by Dapsome: A Possible Therapeutic Mechanism of Dapsone in the Treatment of IgA Dermatoses. J. Investig. Dermatol. 1993, 100, 349–355. [Google Scholar] [CrossRef]
- Amukoye, E.; Winstanley, P.A.; Watkins, W.M.; Snow, R.W.; Hatcher, J.; Mosobo, M.; Ngumbao, E.; Lowe, B.; Ton, M.; Minyiri, G.; et al. Chlorproguanil-Dapsone: Effective Treatment for Uncomplicated Falciparum Malaria. Antimicrob. Agents Chemother. 1997, 41, 2261–2264. [Google Scholar] [CrossRef]
- Wolf, R.; Tuzun, B.; Tuzun, Y. Dapsone: Unapproved Uses or Indications. Clin. Dermatol. 2000, 18, 37–53. [Google Scholar] [CrossRef]
- Sangiolo, D.; Storer, B.; Nash, R.; Corey, L.; Davis, C.; Flowers, M.; Hackman, R.C.; Boeckh, M. Toxicity and Efficacy of Daily Dapsone as Pneumocystis Jiroveci Prophylaxis after Hematopoietic Stem Cell Transplantation: A Case-Control Study. Biol. Blood Marrow Transplant. 2005, 11, 521–529. [Google Scholar] [CrossRef]
- Wolverton, S.E. Monitoring for Adverse Effects from Systemic Drugs Used in Dermatology. J. Am. Acad. Dermatol. 1992, 26, 661–679. [Google Scholar] [CrossRef]
- Coleman, M.D. Dapsone: Modes of Action, Toxicity and Possible Strategies for Increasing Patient Tolerance. Br. J. Dermatol. 1993, 129, 507–513. [Google Scholar] [CrossRef]
- Farhi, D.; Bégon, E.; Wolkenstein, P.; Chosidow, O. Dapsone in Dermatology. EMC-Dermatol. -Cosmetol. 2005, 2, 103–117. [Google Scholar] [CrossRef]
- Goncalves, H.D.S.; Pontes, M.A.d.A.; Bührer-Sékula, S.; Cruz, R.; Almeida, P.C.; de Moraes, M.E.A.; Penna, G.O. Brazilian Clinical Trial of Uniform Multidrug Therapy for Leprosy Patients-The Correlation between Clinical Disease Types and Adverse Effects. Mem. Inst. Oswaldo Cruz 2012, 107, 74–78. [Google Scholar] [CrossRef]
- Winter, H.R.; Yi, W.; Unadkat, J.D. CYP2C8/9 Mediate Dapsone N-Hydroxylation at Clinical Concentrations of Dapsone. Drug Metab. Dispos. 2000, 28, 865–868. [Google Scholar]
- Albuquerque, R.V.; Malcher, N.S.; Amado, L.L.; Coleman, M.D.; Dos Santos, D.C.; Borges, R.S.; Valente, S.A.S.; Valente, V.C.; Monteiro, M.C. In Vitro Protective Effect and Antioxidant Mechanism of Resveratrol Induced by Dapsone Hydroxylamine in Human Cells. PLoS ONE 2015, 10, e0134768. [Google Scholar] [CrossRef] [PubMed]
- Sohal, R.S.; Ku, H.H.; Agarwal, S.; Forster, M.J.; Lal, H. Oxidative Damage, Mitochondrial Oxidant Generation and Antioxidant Defenses during Aging and in Response to Food Restriction in the Mouse. Mech. Ageing Dev. 1994, 74, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Gualandri, W.; Gualandri, L.; Demartini, G.; Esposti, R.; Marthyn, P.; Volontè, S.; Stangoni, L.; Borgonovo, M.; Fraschini, F. Redox Balance in Patients with Down’s Syndrome before and after Dietary Supplementation with α-Lipoic Acid and L-Cysteine. Int. J. Clin. Pharmacol. Res. 2003, 23, 23–30. [Google Scholar] [PubMed]
- Golbidi, S.; Badran, M.; Laher, I. Diabetes and Alpha Lipoic Acid. Front. Pharmacol. 2011, 2, 69. [Google Scholar] [CrossRef]
- Gomes, M.B.; Negrato, C.A. Alpha-Lipoic Acid as a Pleiotropic Compound with Potential Therapeutic Use in Diabetes and Other Chronic Diseases. Diabetol. Metab. Syndr. 2014, 6, 80. [Google Scholar] [CrossRef]
- El Midaoui, A.; Fantus, I.G.; Boughrous, A.A.; Couture, R. Beneficial Effects of Alpha-Lipoic Acid on Hypertension, Visceral Obesity, UCP-1 Expression and Oxidative Stress in Zucker Diabetic Fatty Rats. Antioxidants 2019, 8, 23–30. [Google Scholar] [CrossRef]
- Palaniappan, A.R.; Dai, A. Mitochondrial Ageing and the Beneficial Role of α-Lipoic Acid. Neurochem. Res. 2007, 32, 1552–1558. [Google Scholar] [CrossRef]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-Lipoic Acid as a Dietary Supplement: Molecular Mechanisms and Therapeutic Potential. Biochim. Biophys. Acta-Gen. Subj. 2009, 1790, 1149–1160. [Google Scholar] [CrossRef]
- Bustamante, J.; Lodge, J.K.; Marcocci, L.; Tritschler, H.J.; Packer, L.; Rihn, B.H. α-Lipoic Acid in Liver Metabolism and Disease. Free Radic. Biol. Med. 1998, 24, 1023–1039. [Google Scholar] [CrossRef]
- Reed, L.J. From Lipoic Acid to Multi-Enzyme Complexes. Protein Sci. 1998, 7, 220–224. [Google Scholar] [CrossRef]
- Moini, H.; Packer, L.; Saris, N.E.L. Antioxidant and Prooxidant Activities of α-Lipoic Acid and Dihydrolipoic Acid. Toxicol. Appl. Pharmacol. 2002, 182, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.D.; Taylor, C.T. Effects of Dihydrolipoic Acid (DHLA), α-Lipoic Acid. N-Acetyl Cysteine and Ascorbate on Xenobiotic-Mediated Methaemoglobin Formation in Human Erythrocytes in Vitro. Environ. Toxicol. Pharmacol. 2003, 14, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Lexis, L.A.; Fassett, R.G.; Coombes, J.S. α-Tocopherol and α-Lipoic Acid Enhance the Erythrocyte Antioxidant Defence in Cyclosporine A-Treated Rats. Basic Clin. Pharmacol. Toxicol. 2006, 98, 68–73. [Google Scholar] [CrossRef]
- Coleman, M.D.; Williams, C.; Haenen, G.R.M.M. Effects of Lipoic Acid and Dihydrolipoic Acid on 4-Aminophenol-Mediated Erythrocytic Toxicity in Vitro. Basic Clin. Pharmacol. Toxicol. 2006, 99, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.D. Improvement of Patient Tolerance to Dapsone: Current and Future Developments. Dermatol. Online J. 2007, 13, 18. [Google Scholar] [CrossRef]
- Ibrahim, I.H.; Sallam, S.M.; Omar, H.; Rizk, M. Oxidative Hemolysis of Erythrocytes Induced by Various Vitamins. Int. J. Biomed. Sci. 2006, 2, 295–298. [Google Scholar]
- Oliveira, F.R.; Pessoa, M.C.; Albuquerque, R.F.V.; Schalcher, T.R.; Monteiro, M.C. Clinical Applications and Methemoglobinemia Induced by Dapsone. J. Braz. Chem. Soc. 2014, 25, 1770–1779. [Google Scholar] [CrossRef]
- Halim, N.K.D.; Ogbeide, E. Haematological Alterations in Leprosy Patients Treated with Dapsone. East Afr. Med. J. 2002, 79, 100–102. [Google Scholar] [CrossRef][Green Version]
- Schalcher, T.R.; Borges, R.S.; Coleman, M.D.; Júnior, J.B.; Salgado, C.G.; Vieira, J.L.F.; Romão, P.R.T.; Oliveira, F.R.; Monteiro, M.C. Clinical Oxidative Stress during Leprosy Multidrug Therapy: Impact of Dapsone Oxidation. PLoS ONE 2014, 9, e85712. [Google Scholar] [CrossRef]
- Leonard, J.N.; Fry, L. Treatment and Management of Dermatitis Herpetiformis. Clin. Dermatol. 1991, 9, 403–408. [Google Scholar] [CrossRef]
- Vage, C.; Saab, N.; Woster, P.M.; Svensson, C.K. Dapsone-Induced Hematologic Toxicity: Comparison of the Methemoglobin-Forming Ability of Hydroxylamine Metabolites of Dapsone in Rat and Human Blood. Toxicol. Appl. Pharmacol. 1994, 129, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Reilly, T.P.; Woster, P.M.; Svensson, C.K. Methemoglobin Formation by Hydroxylamine Metabolites of Sulfamethoxazole and Dapsone: Implications for Differences in Adverse Drug Reactions. J. Pharmacol. Exp. Ther. 1999, 288, 951–959. [Google Scholar] [PubMed]
- Winterbourn, C.C. Oxidative Reactions of Hemoglobin. Methods Enzymol. 1990, 186, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Hatherill, J.R.; Till, G.O.; Ward, P.A. Mechanisms of Oxidant-Induced Changes in Erythrocytes. Agents Actions 1991, 32, 351–358. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Coleman, M.D.; Simpson, J.; Jacobus, D.P. Reduction of Dapsone Hydroxylamine to Dapsone during Methaemoglobin Formation in Human Erythrocytes in Vitro IV: Implications for the Development of Agranulocytosis. Biochem. Pharmacol. 1994, 48, 1349–1354. [Google Scholar] [CrossRef]
- Don, G.; Ragazzi, E.; Clari, G.; Bordi, L. Hemolysis and Anemia Induced by Dapsone Hydroxylamine; INTECH Open Access Publisher: London, UK, 2012. [Google Scholar]
- Chikezie, P.C.; Ekechukwu, C.U. Acute Patho-Toxicological Indicators of Methaemoglobinemia. J. Acute Dis. 2016, 5, 179–184. [Google Scholar] [CrossRef][Green Version]
- Veggi, L.M.; Pretto, L.; Ochoa, E.J.; Catania, V.A.; Luquita, M.G.; Taborda, D.R.; Sánchez Pozzi, E.J.; Ikushiro, S.; Coleman, M.D.; Roma, M.G.; et al. Dapsone Induces Oxidative Stress and Impairs Antioxidant Defenses in Rat Liver. Life Sci. 2008, 83, 155–163. [Google Scholar] [CrossRef]
- Maddux, B.A.; See, W.; Lawrence, J.C.; Goldfine, A.L.; Goldfine, I.D.; Evans, J.L. Protection against Oxidative Stress-Induced Insulin Resistance in Rat L6 Muscle Cells by Micromolar Concentrations of α-Lipoic Acid. Diabetes 2001, 50, 404–410. [Google Scholar] [CrossRef]
- Morikawa, T.; Yasuno, R.; Wada, H. Do Mammalian Cells Synthesize Lipoic Acid? Identification of a Mouse CDNA Encoding a Lipoic Acid Synthase Located in Mitochondria. FEBS Lett. 2001, 498, 16–21. [Google Scholar] [CrossRef]
- Salehi, B.; Berkay Yılmaz, Y.; Antika, G.; Boyunegmez Tumer, T.; Fawzi Mahomoodally, M.; Lobine, D.; Akram, M.; Riaz, M.; Capanoglu, E.; Sharopov, F.; et al. Insights on the Use of α-Lipoic Acid for Therapeutic Purposes. Biomolecules 2019, 9, 356. [Google Scholar] [CrossRef]
- Uchida, R.; Okamoto, H.; Ikuta, N.; Terao, K.; Hirota, T. Enantioselective Pharmacokinetics of α-Lipoic Acid in Rats. Int. J. Mol. Sci. 2015, 16, 22781–22794. [Google Scholar] [CrossRef] [PubMed]
- Klaus Krämer, L.P. R-α-Lipoic Acid. In Nutraceuticals in Health and Disease Prevention; CRC Press: Boca Raton, FL, USA, 2001; p. 34. [Google Scholar]
- Carlson, D.A.; Smith, A.R.; Fischer, S.J.; Young, K.L.; Packer, L. The Plasma Pharmacokinetics of R-(+)-Lipoic Acid Administered as Sodium R-(+)-Lipoate to Healthy Human Subjects. Altern. Med. Rev. 2007, 12, 343–351. [Google Scholar] [PubMed]
- Jibril, A.T.; Jayedi, A.; Shab-Bidar, S. Efficacy and Safety of Oral Alpha-Lipoic Acid Supplementation for Type 2 Diabetes Management: A Systematic Review and Dose–Response Meta-Analysis of Randomized Trials. Endocr. Connect. 2022, 11, e220322. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Moon, S.J.; Lee, K.O.; Yoon, S.H.; Jang, I.J.; Yu, K.S.; Lee, S.H. Comparison of R(+)-α-Lipoic Acid Exposure after R(+)-α-Lipoic Acid 200 Mg and 300 Mg and Thioctic Acid 600 Mg in Healthy Korean Male Subjects. Transl. Clin. Pharmacol. 2016, 24, 137–142. [Google Scholar] [CrossRef][Green Version]
- Seifar, F.; Khalili, M.; Khaledyan, H.; Amiri Moghadam, S.; Izadi, A.; Azimi, A.; Shakouri, S.K. α-Lipoic Acid, Functional Fatty Acid, as a Novel Therapeutic Alternative for Central Nervous System Diseases: A Review. Nutr. Neurosci. 2019, 22, 306–316. [Google Scholar] [CrossRef]
- Haramaki, N.; Han, D.; Handelman, G.J.; Tritschler, H.J.; Packer, L. Cytosolic and Mitochondrial Systems for NADH- and NADPH-Dependent Reduction of α-Lipoic Acid. Free Radic. Biol. Med. 1997, 22, 535–542. [Google Scholar] [CrossRef]
- Solmonson, A.; DeBerardinis, R.J. Lipoic Acid Metabolism and Mitochondrial Redox Regulation. J. Biol. Chem. 2018, 293, 7522–7530. [Google Scholar] [CrossRef]
- Ikuta, N.; Sugiyama, H.; Shimosegawa, H.; Nakane, R.; Ishida, Y.; Uekaji, Y.; Nakata, D.; Pallauf, K.; Rimbach, G.; Terao, K.; et al. Analysis of the Enhanced Stability of R(+)-Alpha Lipoic Acid by the Complex Formation with Cyclodextrins. Int. J. Mol. Sci. 2013, 14, 3639–3655. [Google Scholar] [CrossRef]
- Dos Santos, S.M.; Romeiro, C.F.R.; Rodrigues, C.A.; Cerqueira, A.R.L.; Monteiro, M.C. Mitochondrial Dysfunction and Alpha-Lipoic Acid: Beneficial or Harmful in Alzheimer’s Disease? Oxid. Med. Cell. Longev. 2019, 2019, 8409329. [Google Scholar] [CrossRef]
- Hagen, T.M.; Ingersoll, R.T.; Lykkesfeldt, J.; Liu, J.; Wehr, C.M.; Vinarsky, V.; Bartholomew, J.C.; Ames, B.N. ( R )-α-Lipoic Acid-supplemented Old Rats Have Improved Mitochondrial Function, Decreased Oxidative Damage, and Increased Metabolic Rate. FASEB J. 1999, 13, 411–418. [Google Scholar] [CrossRef]
- Suh, J.H.; Shigeno, E.T.; Morrow, J.D.; Cox, J.B.; Rocha, A.E.; Frei, B.; Hagen, T.M. Oxidative Stress in the Aging Rat Heart Is Reversed by Dietary Supplementation with ( R )-α-lipoic Acid. FASEB J. 2001, 15, 700–706. [Google Scholar] [CrossRef]
- Ou, P.; Tritschler, H.J.; Wolff, S.P. Thioctic (Lipoic) Acid: A Therapeutic Metal-Chelating Antioxidant? Biochem. Pharmacol. 1995, 50, 123–126. [Google Scholar] [CrossRef]
- Yaworsky, K.; Somwar, R.; Ramlal, T.; Tritschler, H.J.; Klip, A. Engagement of the Insulin-Sensitive Pathway in the Stimulation of Glucose Transport by α-Lipoic Acid in 3T3-L1 Adipocytes. Diabetologia 2000, 43, 294–303. [Google Scholar] [CrossRef]
- Konrad, D.; Somwar, R.; Sweeney, G.; Yaworsky, K.; Hayashi, M.; Ramlal, T.; Klip, A. The Antihyperglycemic Drug α-Lipoic Acid Stimulates Glucose Uptake via Both GLUT4 Translocation and GLUT4 Activation: Potential Role of P38 Mitogen-Activated Protein Kinase in GLUT4 Activation. Diabetes 2001, 50, 1464–1471. [Google Scholar] [CrossRef]
- Pick, U.; Haramaki, N.; Constantinescu, A.; Handelman, G.J.; Tritschler, H.J.; Packer, L. Glutathione Reductase and Lipoamide Dehydrogenase Have Opposite Stereospecificities for α-Lipoic Acid Enantiomers. Biochem. Biophys. Res. Commun. 1995, 206, 724–730. [Google Scholar] [CrossRef]
- Bilska, A.; Włodek, L. Lipoic Acid-The Drug of the Future? Pharmacol. Rep. 2005, 57, 570–577. [Google Scholar]
- May, J.M.; Qu, Z.C.; Nelson, D.J. Uptake and Reduction of α-Lipoic Acid by Human Erythrocytes. Clin. Biochem. 2007, 40, 1135–1142. [Google Scholar] [CrossRef]
- Lisovskaya, I.L.; Shcherbachenko, I.M.; Volkova, R.I.; Ataullakhanov, F.I. Clotrimazole Enhances Lysis of Human Erythrocytes Induced by T-BHP. Chem. Biol. Interact. 2009, 180, 433–439. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Yamada, T.; Hashida, K.; Takarada-Iemata, M.; Matsugo, S.; Hori, O. α-Lipoic Acid (LA) Enantiomers Protect SH-SY5Y Cells against Glutathione Depletion. Neurochem. Int. 2011, 59, 1003–1009. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione Synthesis. Biochim. Biophys. Acta-Gen. Subj. 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Tibullo, D.; Li Volti, G.; Giallongo, C.; Grasso, S.; Tomassoni, D.; Anfuso, C.D.; Lupo, G.; Amenta, F.; Avola, R.; Bramanti, V. Biochemical and Clinical Relevance of Alpha Lipoic Acid: Antioxidant and Anti-Inflammatory Activity, Molecular Pathways and Therapeutic Potential. Inflamm. Res. 2017, 66, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Handelman, G.; Marcocci, L.; Sen, C.K.; Roy, S.; Kobuchi, H.; Tritschler, H.J.; Flohé, L.; Packer, L. Lipoic Acid Increases de Novo Synthesis of Cellular Glutathione by Improving Cystine Utilization. BioFactors 1997, 6, 321–338. [Google Scholar] [CrossRef]
- Qasim, N.; Mahmood, R. Diminution of Oxidative Damage to Human Erythrocytes and Lymphocytes by Creatine: Possible Role of Creatine in Blood. PLoS ONE 2015, 10, e0141975. [Google Scholar] [CrossRef]
- Hemnani, T.; Parihar, M.S. Reactive Oxygen Species and Oxidative DNA Damage. Indian J. Physiol. Pharmacol. 1998, 42, 440–452. [Google Scholar]
- Mirjana, M.; Jelena, A.; Aleksandra, U.; Svetlana, D.; Nevena, G.; Jelena, M.; Goran, P.; Melita, V. Alpha-Lipoic Acid Preserves the Structural and Functional Integrity of Red Blood Cells by Adjusting the Redox Disturbance and Decreasing O-GlcNAc Modifications of Antioxidant Enzymes and Heat Shock Proteins in Diabetic Rats. Eur. J. Nutr. 2012, 51, 975–986. [Google Scholar] [CrossRef]
- Hackel, C.; Beiguelman, B. Chromosomal Aberrations in Cultures of Skin Fibroblasts of Leprosy Patients. Int. J. Lepr. 1985, 53, 533–539. [Google Scholar]
- Chen, J.; Li, Q. Lipoic Acid Decreases the Expression of Poly ADP-Ribose Polymerase and Inhibits Apoptosis in Diabetic Rats. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 1725–1731. [Google Scholar] [CrossRef]
- Ikuta, N.; Okamoto, H.; Furune, T.; Uekaji, Y.; Terao, K.; Uchida, R.; Iwamoto, K.; Miyajima, A.; Hirota, T.; Sakamoto, N. Bioavailability of an R-α-Lipoic Acid/γ-Cyclodextrin Complex in Healthy Volunteers. Int. J. Mol. Sci. 2016, 17, 949. [Google Scholar] [CrossRef]
- McMillan, D.C.; Simson, J.V.; Budinsky, R.A.; Jollow, D.J. Dapsone-Induced Hemolytic Anemia: Effect of Dapsone Hydroxylamine on Sulfhydryl Status, Membrane Skeletal Proteins and Morphology of Human and Rat Erythrocytes. J. Pharmacol. Exp. Ther. 1995, 274, 540–547. [Google Scholar]
- Pandey, K.B.; Rizvi, S.I. Protective Effect of Resveratrol on Markers of Oxidative Stress in Human Erythrocytes Subjected to in Vitro Oxidative Insult. Phyther. Res. 2010, 24, S11–S14. [Google Scholar] [CrossRef] [PubMed]
- Reilly, T.P.; Bellevue, F.H.; Woster, P.M.; Svensson, C.K. Comparison of the in Vitro Cytotoxicity of Hydroxylamine Metabolites of Sulfamethoxazole and Dapsone. Biochem. Pharmacol. 1998, 55, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Hegesh, E. A Sensitive Micromethod for the Determination of Methemoglobin in Blood. Clin. Chim. Acta 1970, 30, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Zhang, J.; Wang, Y.; Du, L.Q.; Xu, C.; Wang, Q.; Liu, J.X.; Su, X.; Fan, F.Y.; Liu, Q.; et al. Cytogenetic Abnormalities in Lymphocytes from Victims Exposed to Cobalt-60 Radiation. Int. J. Mol. Sci. 2013, 14, 17525–17535. [Google Scholar] [CrossRef]
- Anderson, D.; Yu, T.W.; Phillips, B.J.; Schmezer, P. The Effect of Various Antioxidants and Other Modifying Agents on Oxygen-Radical-Generated DNA Damage in Human Lymphocytes in the COMET Assay. Mutat. Res. -Fundam. Mol. Mech. Mutagen. 1994, 307, 261–271. [Google Scholar] [CrossRef]
- Tedesco, I.; Russo, M.; Russo, P.; Iacomino, G.; Russo, G.L.; Carraturo, A.; Faruolo, C.; Moio, L.; Palumbo, R. Antioxidant Effect of Red Wine Polyphenols on Red Blood Cells. J. Nutr. Biochem. 2000, 11, 114–119. [Google Scholar] [CrossRef]
- Jung, U.; Zheng, X.; Yoon, S.O.; Chung, A.S. Se-Methylselenocysteine Induces Apoptosis Mediated by Reactive Oxygen Species in HL-60 Cells. Free Radic. Biol. Med. 2001, 31, 479–489. [Google Scholar] [CrossRef]
- Diehn, M.; Cho, R.W.; Lobo, N.A.; Kalisky, T.; Dorie, M.J.; Kulp, A.N.; Qian, D.; Lam, J.S.; Ailles, L.E.; Wong, M.; et al. Association of Reactive Oxygen Species Levels and Radioresistance in Cancer Stem Cells. Nature 2009, 458, 780–783. [Google Scholar] [CrossRef]
- Myhre, O.; Andersen, J.M.; Aarnes, H.; Fonnum, F. Evaluation of the Probes 2′,7′-Dichlorofluorescin Diacetate, Luminol, and Lucigenin as Indicators of Reactive Species Formation. Biochem. Pharmacol. 2003, 65, 1575–1582. [Google Scholar] [CrossRef]
- Gomes, A.; Fernandes, E.; Lima, J.L.F.C. Fluorescence Probes Used for Detection of Reactive Oxygen Species. J. Biochem. Biophys. Methods 2005, 65, 45–80. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos SM, L.; Goulart MO, F.; Moura JB, D.F.; Manfredini, V.; Benfato MD, S.; Kubota, L.T. Espécies reativas de oxigênio e de nitrogênio, antioxidantes e marcadores de dano oxidativo em sangue humano: Principais métodos analíticos para sua determinação. Quim. Nova 2007, 30, 1323–1338. [Google Scholar] [CrossRef]
- McCords, J.M.; Fridovich, I. Superoxide Dismutase. J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeerschd, T.; Zurek, E.; Hutchison, G.R. Avogadro: An Advanced Semantic Chemical Editor, Visualization, and Analysis Platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef]
- Revision, D.; Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; et al. Gaussian 09, Revision; Gaussian, Inc.: Wallingford, CT, USA, 2015; pp. 1–20. [Google Scholar]
- Hohenberg, P.; Kohn, W. Physical Review Physical Review. Phys. Rev. 1964, 136, 864–871. [Google Scholar] [CrossRef]
- Lee, C.; Yang, W.; Parr, R.G. Development of the Colle-Salvetti Correlation-Energy Formula into a Functional of the Electron Density. Phys. Rev. B 1988, 37, 785–789. [Google Scholar] [CrossRef]
- RCSB-Protein Data Bank. Available online: https://www.rcsb.org/ (accessed on 19 July 2022).
- Yi, J.; Soares, A.S.; Richter-Addo, G.B. Crystallographic Characterization of the Nitric Oxide Derivative of R-State Human Hemoglobin. Nitric Oxide-Biol. Chem. 2014, 39, 46–50. [Google Scholar] [CrossRef]
- Schrödinger, L.; DeLano, W. Polymol. Available online: www.pymol.org. (accessed on 19 July 2022).
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated Docking Using a Lamarckian Genetic Algorithm and an Empirical Binding Free Energy Function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef]
- Jacob, G.; Schorr, M.; Moist, L.M. Methemoglobinemia in a 28-Year-Old Woman Treated with Dapsone. Can. Med. Assoc. J. 2022, 194, E1062–E1065. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comp | HOMO (eV) | LUMO (eV) | GAP (eV) |
|---|---|---|---|
| 1 | −5.58 | −0.97 | 4.61 |
| 2 | −5.56 | −1.00 | 4.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espíndola, K.M.M.; Varela, E.L.P.; de Albuquerque, R.d.F.V.; Figueiredo, R.A.; dos Santos, S.M.; Malcher, N.S.; da S. Seabra, P.S.; Fonseca, A.d.N.; de Azevedo Sousa, K.M.; de Oliveira, S.B.B.; et al. Alpha-Lipoic Acid and Its Enantiomers Prevent Methemoglobin Formation and DNA Damage Induced by Dapsone Hydroxylamine: Molecular Mechanism and Antioxidant Action. Int. J. Mol. Sci. 2023, 24, 57. https://doi.org/10.3390/ijms24010057
Espíndola KMM, Varela ELP, de Albuquerque RdFV, Figueiredo RA, dos Santos SM, Malcher NS, da S. Seabra PS, Fonseca AdN, de Azevedo Sousa KM, de Oliveira SBB, et al. Alpha-Lipoic Acid and Its Enantiomers Prevent Methemoglobin Formation and DNA Damage Induced by Dapsone Hydroxylamine: Molecular Mechanism and Antioxidant Action. International Journal of Molecular Sciences. 2023; 24(1):57. https://doi.org/10.3390/ijms24010057
Chicago/Turabian StyleEspíndola, Kaio Murilo Monteiro, Everton Luiz Pompeu Varela, Rosyana de Fátima Vieira de Albuquerque, Rosiane Araújo Figueiredo, Sávio Monteiro dos Santos, Nívea Silva Malcher, Pamela Suelen da S. Seabra, Andréia do Nascimento Fonseca, Karla Marcely de Azevedo Sousa, Susan Beatriz Batista de Oliveira, and et al. 2023. "Alpha-Lipoic Acid and Its Enantiomers Prevent Methemoglobin Formation and DNA Damage Induced by Dapsone Hydroxylamine: Molecular Mechanism and Antioxidant Action" International Journal of Molecular Sciences 24, no. 1: 57. https://doi.org/10.3390/ijms24010057
APA StyleEspíndola, K. M. M., Varela, E. L. P., de Albuquerque, R. d. F. V., Figueiredo, R. A., dos Santos, S. M., Malcher, N. S., da S. Seabra, P. S., Fonseca, A. d. N., de Azevedo Sousa, K. M., de Oliveira, S. B. B., Carneiro, A. d. S., Coleman, M. D., & Monteiro, M. C. (2023). Alpha-Lipoic Acid and Its Enantiomers Prevent Methemoglobin Formation and DNA Damage Induced by Dapsone Hydroxylamine: Molecular Mechanism and Antioxidant Action. International Journal of Molecular Sciences, 24(1), 57. https://doi.org/10.3390/ijms24010057