
PRDX2 Knockdown Inhibits Extracellular Matrix Synthesis of Chondrocytes by Inhibiting Wnt5a/YAP1/CTGF and Activating IL-6/JAK2/STAT3 Pathways in Deer Antler


Abstract
:1. Introduction
2. Results
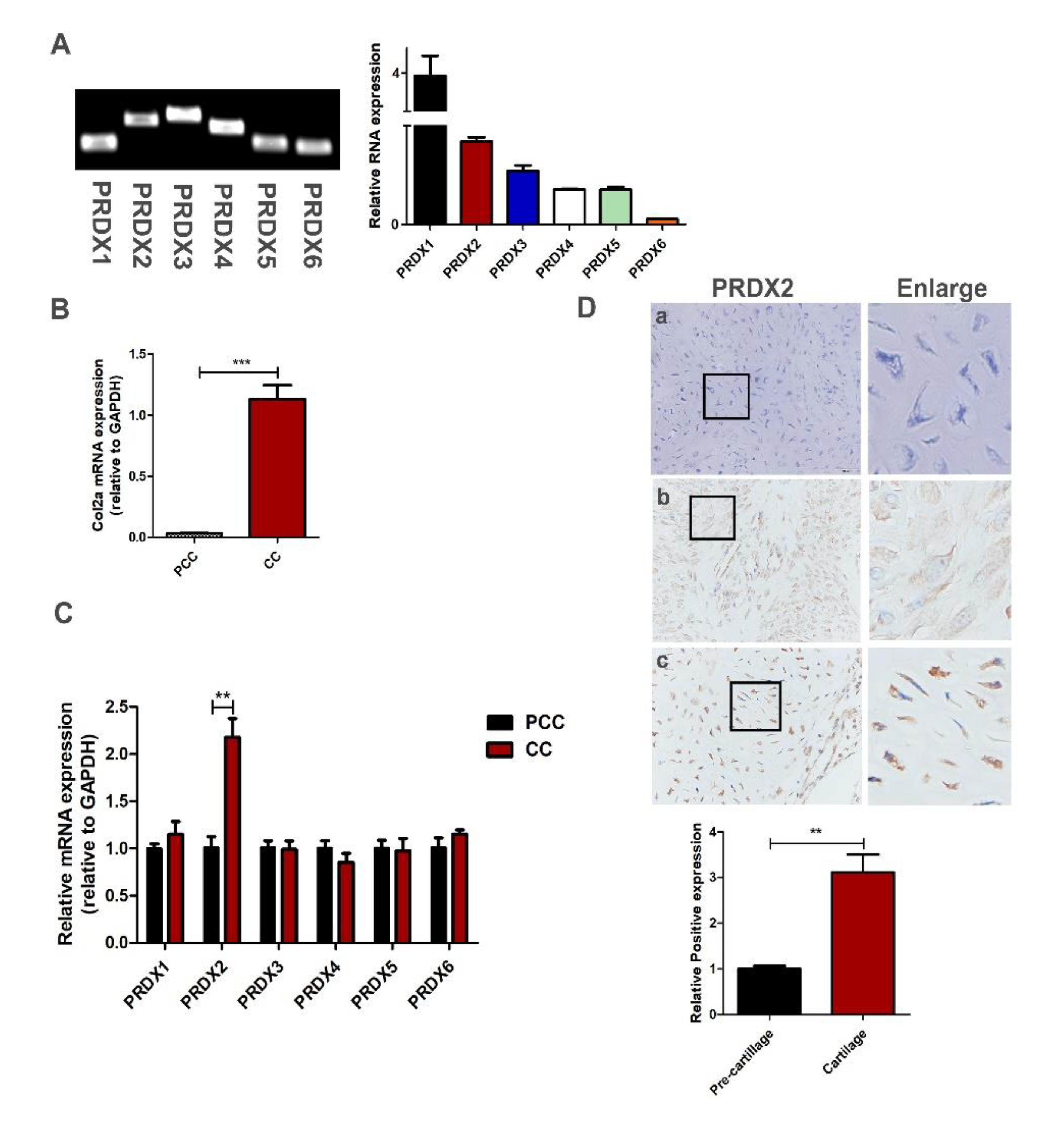
2.1. Antler Chondrocytes Can Highly Express a Variety of Peroxiredoxins, and the Expression of PRDX2 Upregulates as the Chondrocytes Mature
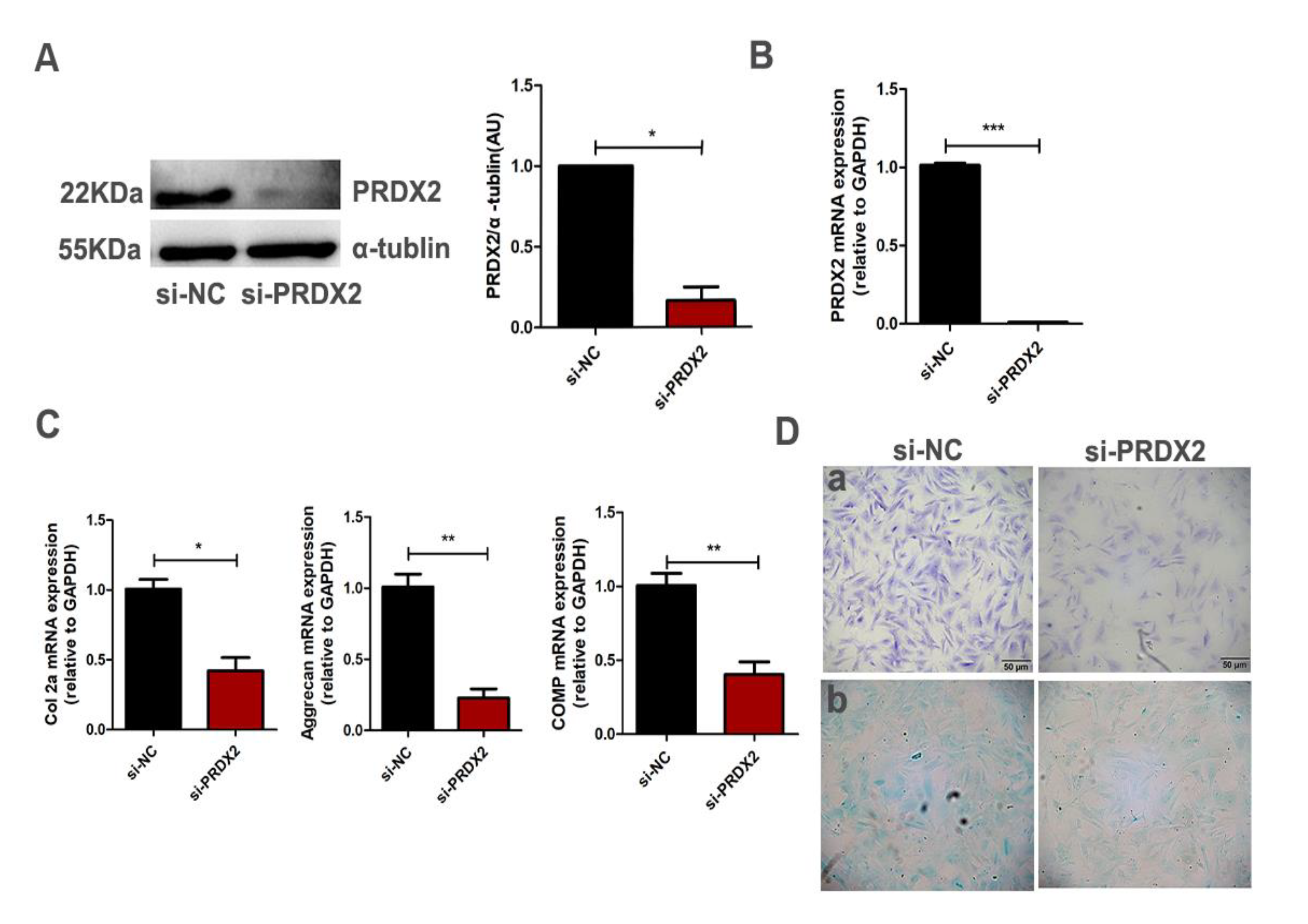
2.2. PRDX2 Knockdown Inhibits the Synthesis of Cartilage Matrix Proteins
2.3. PRDX2 Knockdown Inhibits the Expression of CTGF
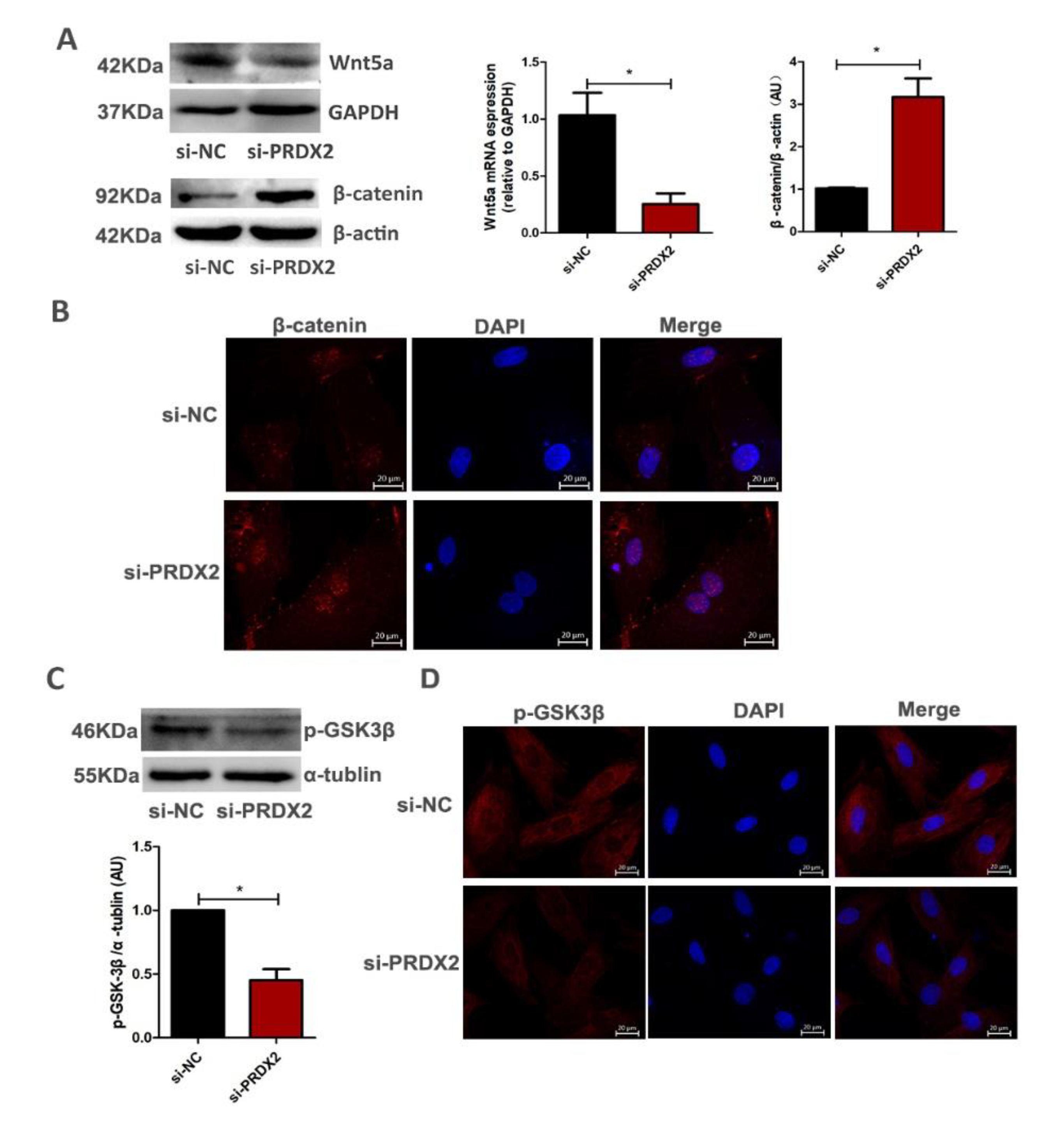
2.4. PRDX2 Knockdown Inhibits the Expression of Wnt5a and Promotes Nuclear Translocation of β-Catenin
2.5. PRDX2 Knockdown Reduces the Expression of CTGF by Inhibiting the Activity of YAP1
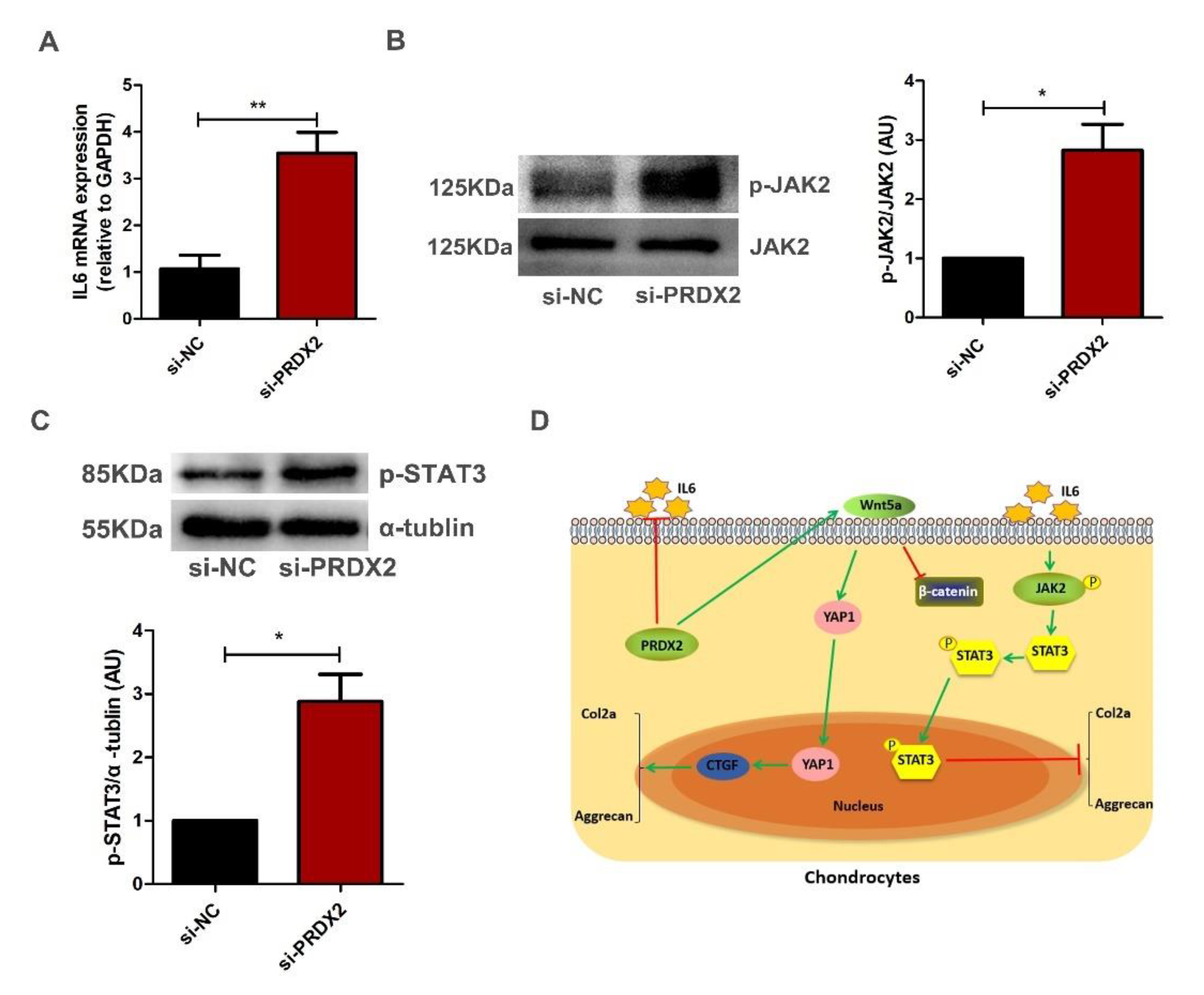
2.6. PRDX2 Knockdown Activates IL6-Induced JAK2/STAT3 Signaling Pathway in Chondrocytes
3. Discussion
4. Materials and Methods
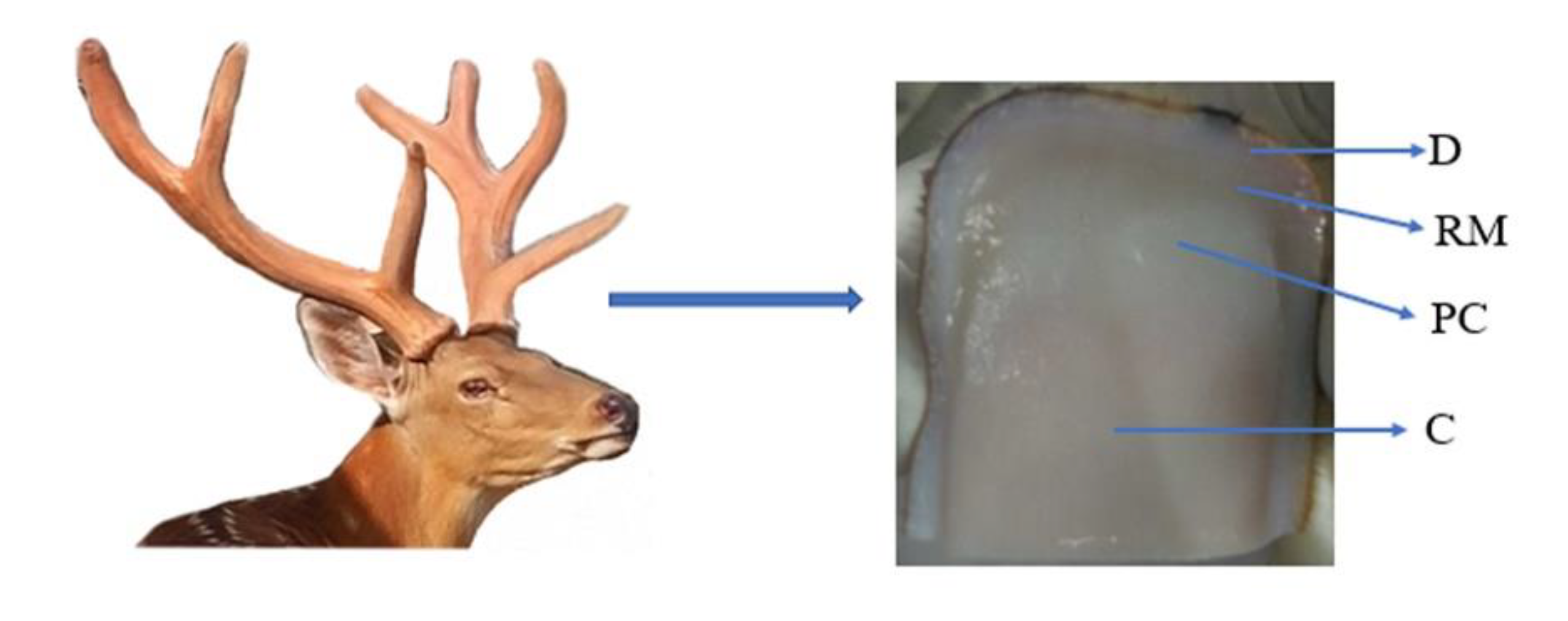
4.1. Antler Issue Collection and Cell Culture
4.2. Immunohistochemistry Staining
4.3. RNA Interference
4.4. Total RNA and Quantitative Real-Time PCR (qRT-PCR)
4.5. Western Blot Assay
4.6. Immunofluorescence Assay
4.7. Toluidine Blue and Alcian Blue Staining
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Price, J.S.; Oyajobi, B.O.; Nalin, A.M.; Frazer, A.; Russell, R.G.; Sandell, L.J. Chondrogenesis in the regenerating antler tip in red deer: Expression of collagen types I, IIA, IIB, and X demonstrated by in situ nucleic acid hybridization and immunocytochemistry. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1996, 205, 332–347. [Google Scholar] [CrossRef]
- Li, C.; Clark, D.E.; Lord, E.A.; Stanton, J.A.; Suttie, J.M. Sampling technique to discriminate the different tissue layers of growing antler tips for gene discovery. Anat. Rec. 2002, 268, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; He, C.; Sun, X.; Wang, J.; Luo, C.; Liu, G.; Yang, L.; Xiong, J.; Huo, L. The Regulatory Mechanism of MLT/MT1 Signaling on the Growth of Antler Mesenchymal Cells. Molecules 2017, 22, 1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, C.; Yik, J.H.; Kishore, A.; Van Dinh, V.; Di Cesare, P.E.; Haudenschild, D.R. Cartilage oligomeric matrix protein and its binding partners in the cartilage extracellular matrix: Interaction, regulation and role in chondrogenesis. Matrix Biol. J. Int. Soc. Matrix Biol. 2014, 37, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Andoh, M.; Ueno-Kudoh, H.; Sato, T.; Miyaki, S.; Asahara, H. Sox9 directly promotes Bapx1 gene expression to repress Runx2 in chondrocytes. Exp. Cell Res. 2009, 315, 2231–2240. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, R.; Wakabayashi, M.; Hata, K.; Matsubara, T.; Honma, S.; Wakisaka, S.; Kiyonari, H.; Shioi, G.; Yamaguchi, A.; Tsumaki, N.; et al. Osterix regulates calcification and degradation of chondrogenic matrices through matrix metalloproteinase 13 (MMP13) expression in association with transcription factor Runx2 during endochondral ossification. J. Biol. Chem. 2012, 287, 33179–33190. [Google Scholar] [CrossRef] [Green Version]
- Rahmati, M.; Nalesso, G.; Mobasheri, A.; Mozafari, M. Aging and osteoarthritis: Central role of the extracellular matrix. Ageing Res. Rev. 2017, 40, 20–30. [Google Scholar] [CrossRef]
- Li, Y.J.; Zhao, Y.H.; Yang, Q. Development of cartilage extracellular matrix in cartilage tissue engineering. Hua Xi Kou Qiang Yi Xue Za Zhi = Huaxi Kouqiang Yixue Zazhi = West China J. Stomatol. 2019, 37, 220–223. [Google Scholar]
- Kong, J.; Wang, J.; Gong, X.; Zheng, X.; Chen, T. Punicalagin Inhibits Tert-Butyl Hydroperoxide-Induced Apoptosis and Extracellular Matrix Degradation in Chondrocytes by Activating Autophagy and Ameliorates Murine Osteoarthritis. Drug Des. Dev. Ther. 2020, 14, 5521–5533. [Google Scholar] [CrossRef]
- Peskin, A.V.; Meotti, F.C.; de Souza, L.F.; Anderson, R.F.; Winterbourn, C.C.; Salvador, A. Intra-dimer cooperativity between the active site cysteines during the oxidation of peroxiredoxin 2. Free Radic. Biol. Med. 2020, 158, 115–125. [Google Scholar] [CrossRef]
- Checconi, P.; Salzano, S.; Bowler, L.; Mullen, L.; Mengozzi, M.; Hanschmann, E.M.; Lillig, C.H.; Sgarbanti, R.; Panella, S.; Nencioni, L.; et al. Redox proteomics of the inflammatory secretome identifies a common set of redoxins and other glutathionylated proteins released in inflammation, influenza virus infection and oxidative stress. PLoS ONE 2015, 10, e0127086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; He, J.; Tang, M.; Sun, H. Prdx2 Upregulation Promotes the Growth and Survival of Gastric Cancer Cells. Pathol. Oncol. Res. POR 2020, 26, 1869–1877. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, Z.; Zhu, S.; Lu, H.; Peng, D.; Soutto, M.; Naz, H.; Peek, R., Jr.; Xu, H.; Zaika, A.; et al. PRDX2 protects against oxidative stress induced by H. pylori and promotes resistance to cisplatin in gastric cancer. Redox Biol. 2020, 28, 101319. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, C.; Wang, W.; Liu, L.; Zhang, Q.; Zhang, J.; Wang, B.; Wang, S.; Hou, L.; Gao, C.; et al. PRDX2 Protects Against Atherosclerosis by Regulating the Phenotype and Function of the Vascular Smooth Muscle Cell. Front. Cardiovasc. Med. 2021, 8, 624796. [Google Scholar] [CrossRef]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef]
- Vallée, A.; Guillevin, R.; Vallée, J.N. Vasculogenesis and angiogenesis initiation under normoxic conditions through Wnt/β-catenin pathway in gliomas. Rev. Neurosci. 2018, 29, 71–91. [Google Scholar] [CrossRef]
- Hou, L.; Shi, H.; Wang, M.; Liu, J.; Liu, G. MicroRNA-497-5p attenuates IL-1β-induced cartilage matrix degradation in chondrocytes via Wnt/β-catenin signal pathway. Int. J. Clin. Exp. Pathol. 2019, 12, 3108–3118. [Google Scholar]
- Li, Z.; Wang, Y.; Xiang, S.; Zheng, Z.; Bian, Y.; Feng, B.; Weng, X. Chondrocytes-derived exosomal miR-8485 regulated the Wnt/β-catenin pathways to promote chondrogenic differentiation of BMSCs. Biochem. Biophys. Res. Commun. 2020, 523, 506–513. [Google Scholar] [CrossRef]
- Wang, W.; Smits, R.; Hao, H.; He, C. Wnt/β-Catenin Signaling in Liver Cancers. Cancers 2019, 11, 926. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Dong, X.; Liu, Y.; Ni, B.; Sai, N.; You, L.; Sun, M.; Yao, Y.; Qu, C.; Yin, X.; et al. Itraconazole exerts anti-liver cancer potential through the Wnt, PI3K/AKT/mTOR, and ROS pathways. Biomed. Pharmacother. 2020, 131, 110661. [Google Scholar] [CrossRef]
- Zhang, P.; Hu, C.; Li, Y.; Wang, Y.; Gao, L.; Lu, K.; Chang, G.; Li, Y.; Qin, S.; Zhang, D. Vangl2 is essential for myocardial remodeling activated by Wnt/JNK signaling. Exp. Cell Res. 2018, 365, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Scholz, B.; Korn, C.; Wojtarowicz, J.; Mogler, C.; Augustin, I.; Boutros, M.; Niehrs, C.; Augustin, H.G. Endothelial RSPO3 Controls Vascular Stability and Pruning through Non-canonical WNT/Ca2+/NFAT Signaling. Dev. Cell 2016, 36, 79–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yang, J.; Bao, M.; Zeng, K.; Fu, S.; Wang, C.; Ye, L. Wnt signaling in bone metastasis: Mechanisms and therapeutic opportunities. Life Sci. 2018, 208, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Park, H.W.; Kim, Y.C.; Yu, B.; Moroishi, T.; Mo, J.S.; Plouffe, S.W.; Meng, Z.; Lin, K.C.; Yu, F.X.; Alexander, C.M.; et al. Alternative Wnt Signaling Activates YAP/TAZ. Cell 2015, 162, 780–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Lei, D.; Huang, S.; Yang, Y.; Jiang, C.; Shi, H.; Chen, W.; Zhao, Q.; You, Z.; Ye, X. A novel biodegradable external stent regulates vein graft remodeling via the Hippo-YAP and mTOR signaling pathways. Biomaterials 2020, 258, 120254. [Google Scholar] [CrossRef]
- Melhuish, T.A.; Kowalczyk, I.; Manukyan, A.; Zhang, Y.; Shah, A.; Abounader, R.; Wotton, D. Myt1 and Myt1l transcription factors limit proliferation in GBM cells by repressing YAP1 expression. Biochim. Et Biophys. Acta. Gene Regul. Mech. 2018, 1861, 983–995. [Google Scholar] [CrossRef]
- Chai, J.; Xu, S.; Guo, F. TEAD1 mediates the oncogenic activities of Hippo-YAP1 signaling in osteosarcoma. Biochem. Biophys. Res. Commun. 2017, 488, 297–302. [Google Scholar] [CrossRef]
- Moon, S.; Lee, S.; Caesar, J.A.; Pruchenko, S.; Chaqour, B.J.S.E.J. A CTGF-YAP Regulatory Pathway is Essential for Angiogenesis and Barriergenesis in the Retina. iScience 2020, 23, 101184. [Google Scholar] [CrossRef]
- Woods, A.; Pala, D.; Kennedy, L.; McLean, S.; Rockel, J.S.; Wang, G.; Leask, A.; Beier, F. Rac1 signaling regulates CTGF/CCN2 gene expression via TGFbeta/Smad signaling in chondrocytes. Osteoarthr. Cartil. 2009, 17, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Kanaan, R.A.; Aldwaik, M.; Al-Hanbali, O.A. The role of connective tissue growth factor in skeletal growth and development. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2006, 12, Ra277–Ra281. [Google Scholar]
- Jiang, W.; Takeshita, N.; Maeda, T.; Sogi, C.; Oyanagi, T.; Kimura, S.; Yoshida, M.; Sasaki, K.; Ito, A.; Takano-Yamamoto, T. Connective tissue growth factor promotes chemotaxis of preosteoblasts through integrin α5 and Ras during tensile force-induced intramembranous osteogenesis. Sci. Rep. 2021, 11, 2368. [Google Scholar] [CrossRef] [PubMed]
- Hall-Glenn, F.; Aivazi, A.; Akopyan, L.; Ong, J.R.; Baxter, R.R.; Benya, P.D.; Goldschmeding, R.; van Nieuwenhoven, F.A.; Hunziker, E.B.; Lyons, K.M. CCN2/CTGF is required for matrix organization and to protect growth plate chondrocytes from cellular stress. J. Cell Commun. Signal. 2013, 7, 219–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Tao, P.; Zhou, Q.; Li, J.; Yu, Z.; Wang, X.; Li, J.; Li, C.; Yan, M.; Zhu, Z.; et al. IL-6 secreted by cancer-associated fibroblasts promotes epithelial-mesenchymal transition and metastasis of gastric cancer via JAK2/STAT3 signaling pathway. Oncotarget 2017, 8, 20741–20750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Ding, Z.; Liu, F.; Shan, W.; Cheng, C.; Xu, J.; He, W.; Huang, W.; Ma, J.; Yin, Z. Dopamine delays articular cartilage degradation in osteoarthritis by negative regulation of the NF-κB and JAK2/STAT3 signaling pathways. Biomed. Pharmacother. 2019, 119, 109419. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; He, J.; Sun, Y.; Dong, X.; Yao, J.; Gu, H.; Liu, L. Leptin Induced TLR4 Expression via the JAK2-STAT3 Pathway in Obesity-Related Osteoarthritis. Oxidative Med. Cell. Longev. 2021, 2021, 7385160. [Google Scholar] [CrossRef]
- Sun, X.; Gu, X.; Li, K.; Li, M.; Peng, J.; Zhang, X.; Yang, L.; Xiong, J. Melatonin Promotes Antler Growth by Accelerating MT1-Mediated Mesenchymal Cell Differentiation and Inhibiting VEGF-Induced Degeneration of Chondrocytes. Int. J. Mol. Sci. 2022, 23, 759. [Google Scholar] [CrossRef]
- Tu, B.; Yao, J.; Ferri-Borgogno, S.; Zhao, J.; Chen, S.; Wang, Q.; Yan, L.; Zhou, X.; Zhu, C.; Bang, S.; et al. YAP1 oncogene is a context-specific driver for pancreatic ductal adenocarcinoma. JCI Insight 2019, 4, e130811. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, T.; Hattori, T.; Ono, M.; Uehara, J.; Kubota, S.; Kuboki, T.; Takigawa, M. CCN family 2/connective tissue growth factor (CCN2/CTGF) stimulates proliferation and differentiation of auricular chondrocytes. Osteoarthr. Cartil. 2008, 16, 787–795. [Google Scholar] [CrossRef] [Green Version]
- Yosimichi, G.; Kubota, S.; Nishida, T.; Kondo, S.; Yanagita, T.; Nakao, K.; Takano-Yamamoto, T.; Takigawa, M. Roles of PKC, PI3K and JNK in multiple transduction of CCN2/CTGF signals in chondrocytes. Bone 2006, 38, 853–863. [Google Scholar] [CrossRef]
- Hori, A.; Nishida, T.; Takashiba, S.; Kubota, S.; Takigawa, M. Regulatory mechanism of CCN2 production by serotonin (5-HT) via 5-HT2A and 5-HT2B receptors in chondrocytes. PLoS ONE 2017, 12, e0188014. [Google Scholar] [CrossRef]
- Xing, X.; Li, Z.; Yu, Z.; Cheng, G.; Li, D.; Li, Z. Effects of connective tissue growth factor (CTGF/CCN2) on condylar chondrocyte proliferation, migration, maturation, differentiation and signalling pathway. Biochem. Biophys. Res. Commun. 2018, 495, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Thongon, N.; Castiglioni, I.; Zucal, C.; Latorre, E.; Provenzani, A.J.O. The GSK3β inhibitor BIS I reverts YAP-dependent EMT signature in PDAC cell lines by decreasing SMADs expression level. Oncotarget 2016, 7, 26551–26566. [Google Scholar] [CrossRef] [PubMed]
- Ford, C.E.; Punnia-Moorthy, G.; Henry, C.E.; Llamosas, E.; Nixdorf, S.; Olivier, J.; Caduff, R.; Ward, R.L.; Heinzelmann-Schwarz, V. The non-canonical Wnt ligand, Wnt5a, is upregulated and associated with epithelial to mesenchymal transition in epithelial ovarian cancer. Gynecol. Oncol. 2014, 134, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M. Canonical and non-canonical WNT signaling in cancer stem cells and their niches: Cellular heterogeneity, omics reprogramming, targeted therapy and tumor plasticity (Review). Int. J. Oncol. 2017, 51, 1357–1369. [Google Scholar] [CrossRef] [Green Version]
- Flores-Hernández, E.; Velázquez, D.M.; Castañeda-Patlán, M.C.; Fuentes-García, G.; Fonseca-Camarillo, G.; Yamamoto-Furusho, J.K.; Romero-Avila, M.T.; García-Sáinz, J.A.; Robles-Flores, M. Canonical and non-canonical Wnt signaling are simultaneously activated by Wnts in colon cancer cells. Cell. Signal. 2020, 72, 109636. [Google Scholar] [CrossRef]
- Curto, J.; Del Valle-Pérez, B.; Villarroel, A.; Fuertes, G.; Vinyoles, M.; Peña, R.; García de Herreros, A.; Duñach, M. CK1ε and p120-catenin control Ror2 function in noncanonical Wnt signaling. Mol. Oncol. 2018, 12, 611–629. [Google Scholar] [CrossRef] [Green Version]
- Bin, S.; Xin, L.; Lin, Z.; Jinhua, Z.; Rui, G.; Xiang, Z. Targeting miR-10a-5p/IL-6R axis for reducing IL-6-induced cartilage cell ferroptosis. Exp. Mol. Pathol. 2021, 118, 104570. [Google Scholar] [CrossRef]
- Wang, T.; He, C. Pro-inflammatory cytokines: The link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018, 44, 38–50. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, P.; Sun, X.; Zhou, L.; Zhao, J. Long non-coding RNA DANCR regulates proliferation and apoptosis of chondrocytes in osteoarthritis via miR-216a-5p-JAK2-STAT3 axis. Biosci. Rep. 2018, 38, BSR20181228. [Google Scholar] [CrossRef] [Green Version]
- Jo, H.A.; Kim, J.Y.; Yang, S.H.; Han, S.S.; Joo, K.W.; Kim, Y.S.; Kim, D.K. The role of local IL6/JAK2/STAT3 signaling in high glucose-induced podocyte hypertrophy. Kidney Res. Clin. Pract. 2016, 35, 212–218. [Google Scholar] [CrossRef]
- Mianehsaz, E.; Mirzaei, H.R.; Mahjoubin-Tehran, M.; Rezaee, A.; Sahebnasagh, R.; Pourhanifeh, M.H.; Mirzaei, H.; Hamblin, M.R. Mesenchymal stem cell-derived exosomes: A new therapeutic approach to osteoarthritis? Stem Cell Res. Ther. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, J.H.; Fu, Q.W.; Li, L.X.; Zhou, R.; Liu, N.; Peng, J.H.; Chen, Y. Prx II reduces oxidative stress and cell senescence in chondrocytes by activating the p16-CDK4/6-pRb-E2F signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3448–3458. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) |
|---|---|
| Col2a | F: GAGGCAGCCGGCAACCTGAG |
| R: TGCGAGCTGGGTTCTTGCGG | |
| YAP-1 | F: GTTCCAACCAGCAGCAACAG |
| R: GGTAACTGGCTACGGAGAGC | |
| PRDX2 | F: GCTGAACATTCCCCTGCTG |
| R: CGTCCACATTGGGCTTGAT | |
| Aggrecan | F: CAACCTCCTGGGAGTGAGGA |
| R: GCTTTGCCGTGAGGATCAC | |
| Wnt5a | F: CTCCTTCGCCCAGGTTGTAAT |
| R: GGAACTGATACTGGCACTCCT | |
| GAPDH | F: GAAGGGTGGCGCCAAGAGGG |
| R: GGGGGCCAAGCAGTTGGTGG | |
| CTGF | F: CAAGGGCCTCTTCTGCGACT |
| R: ACGTGCACTGGTATTTGCAG | |
| IL-6 | F: GCATTCCCTCCTCTGGTCA |
| R: AAAACATTCAAGCCGCACA | |
| COMP | F: GATGCGGACAAGGTGGTAGA |
| R:TCCTGGTAGCCAAAGATGAAA | |
| PRDX1 | F:CCCAAGAAACAAGGAGGACTG |
| R:GCCCCTGAATGAGATGCC | |
| PRDX3 | F:GAGCCCTGCATAACGAAGATG |
| R:GAACTGGTGCTAAAGGCGAAT | |
| PRDX4 | F:TGATTCACAGTTCACCCATTTG |
| R:CACGGGAAGGTCATTCAGAGTA | |
| PRDX5 | F:CCGTCGGTGGAGGTATTTG |
| R:GGCAGGTGGGTCTTGGAAC | |
| PRDX6 | F:CTGGCAGGAACTTTGATGAGAT |
| R:CCTCTTCAGGGATGGTTGGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Gu, X.; Peng, J.; Yang, L.; Zhang, X.; Ran, Z.; Xiong, J. PRDX2 Knockdown Inhibits Extracellular Matrix Synthesis of Chondrocytes by Inhibiting Wnt5a/YAP1/CTGF and Activating IL-6/JAK2/STAT3 Pathways in Deer Antler. Int. J. Mol. Sci. 2022, 23, 5232. https://doi.org/10.3390/ijms23095232
Sun X, Gu X, Peng J, Yang L, Zhang X, Ran Z, Xiong J. PRDX2 Knockdown Inhibits Extracellular Matrix Synthesis of Chondrocytes by Inhibiting Wnt5a/YAP1/CTGF and Activating IL-6/JAK2/STAT3 Pathways in Deer Antler. International Journal of Molecular Sciences. 2022; 23(9):5232. https://doi.org/10.3390/ijms23095232
Chicago/Turabian StyleSun, Xuyang, Xiaoying Gu, Jingna Peng, Liguo Yang, Xinxin Zhang, Zaohong Ran, and Jiajun Xiong. 2022. "PRDX2 Knockdown Inhibits Extracellular Matrix Synthesis of Chondrocytes by Inhibiting Wnt5a/YAP1/CTGF and Activating IL-6/JAK2/STAT3 Pathways in Deer Antler" International Journal of Molecular Sciences 23, no. 9: 5232. https://doi.org/10.3390/ijms23095232
APA StyleSun, X., Gu, X., Peng, J., Yang, L., Zhang, X., Ran, Z., & Xiong, J. (2022). PRDX2 Knockdown Inhibits Extracellular Matrix Synthesis of Chondrocytes by Inhibiting Wnt5a/YAP1/CTGF and Activating IL-6/JAK2/STAT3 Pathways in Deer Antler. International Journal of Molecular Sciences, 23(9), 5232. https://doi.org/10.3390/ijms23095232