
Characterization of Interactions between CTX-M-15 and Clavulanic Acid, Desfuroylceftiofur, Ceftiofur, Ampicillin, and Nitrocefin

 ,
,
Abstract
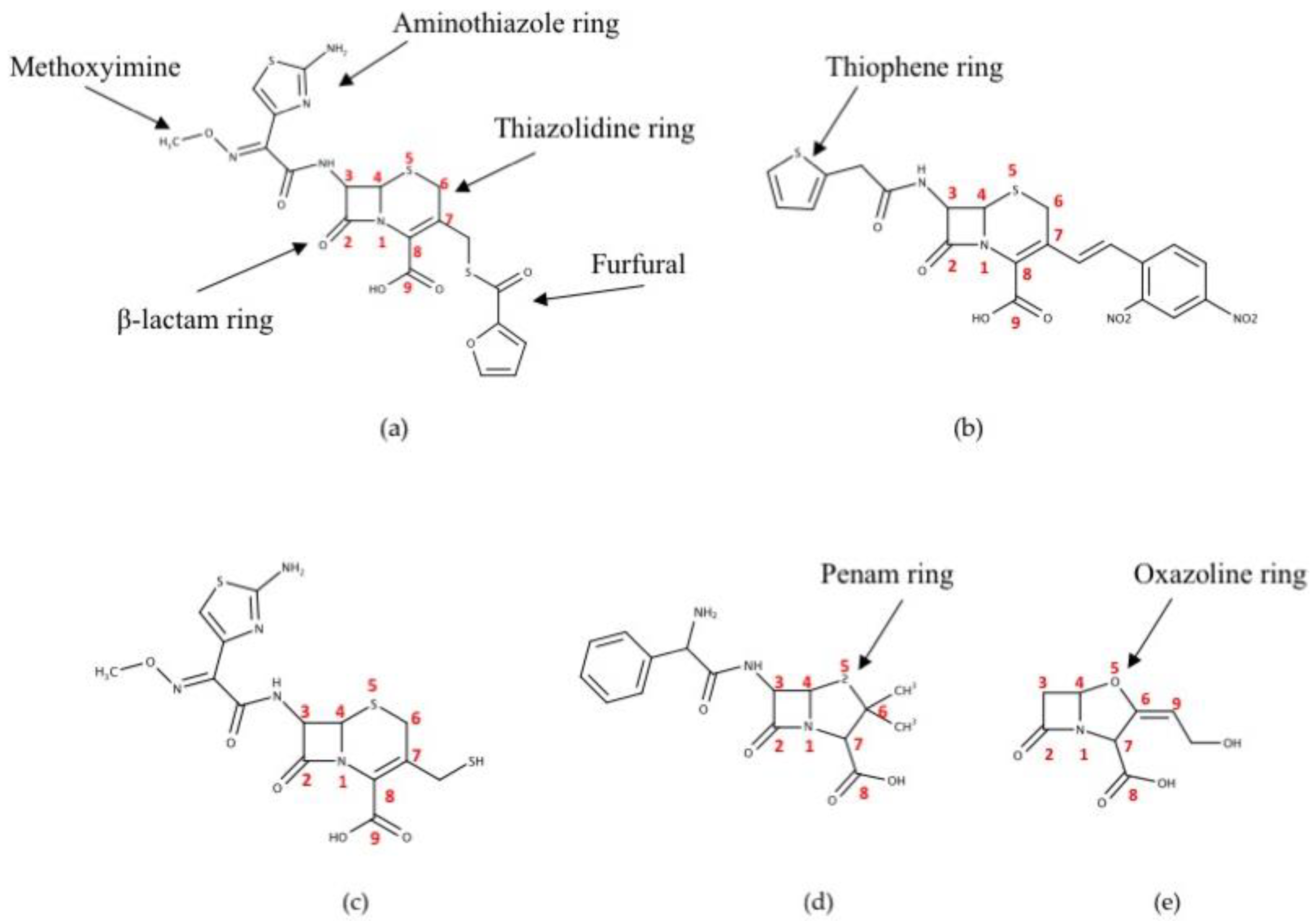
:1. Introduction
2. Results
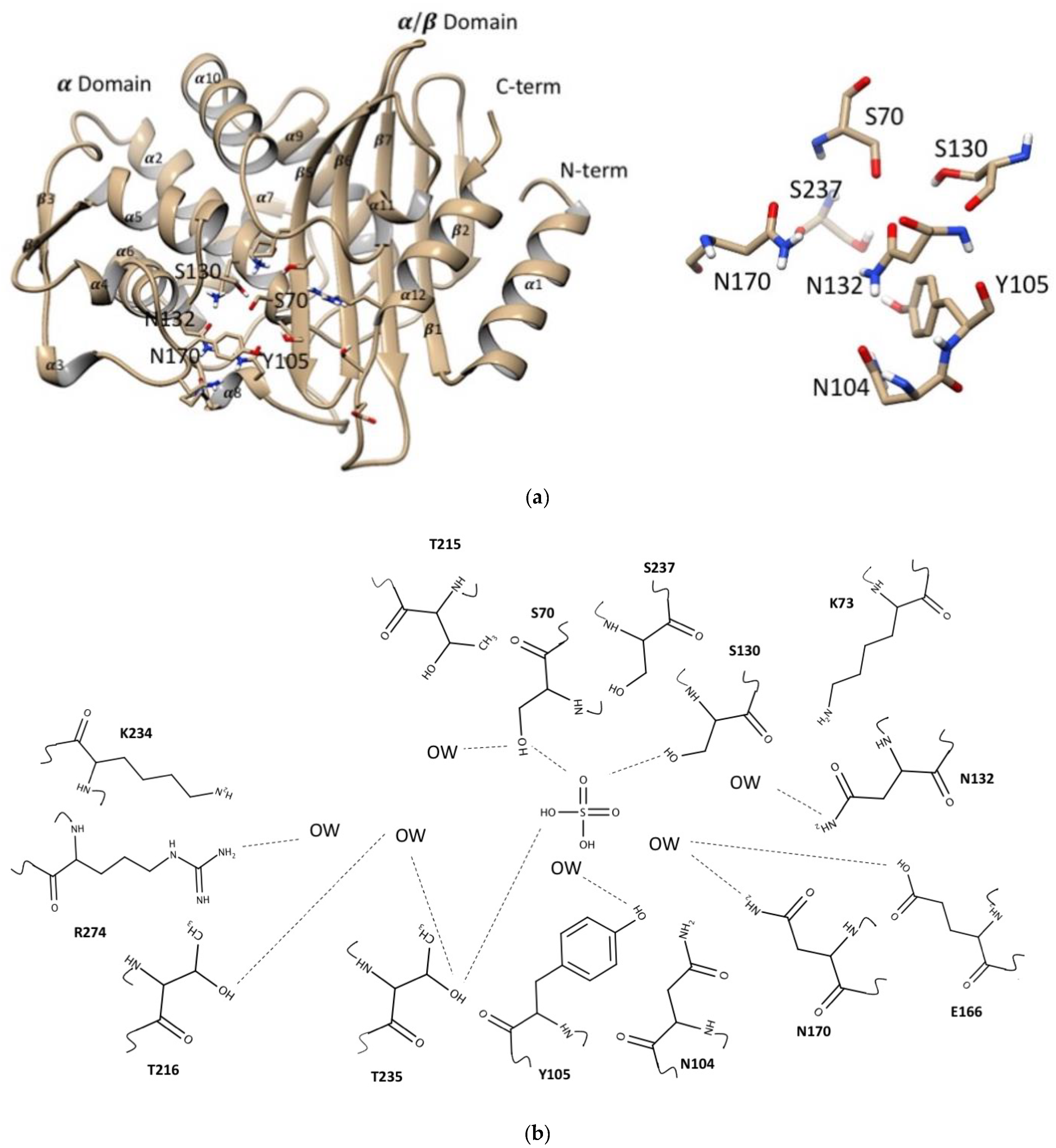
2.1. Global Structure of CTX-M-15
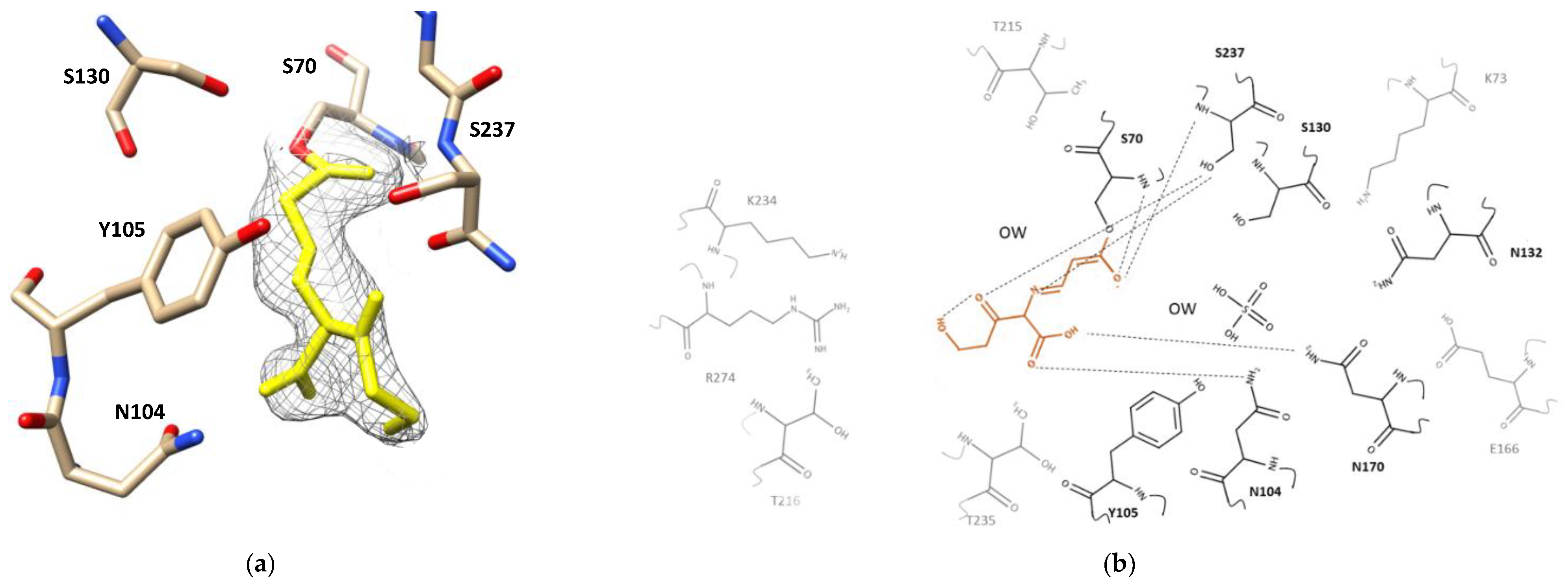
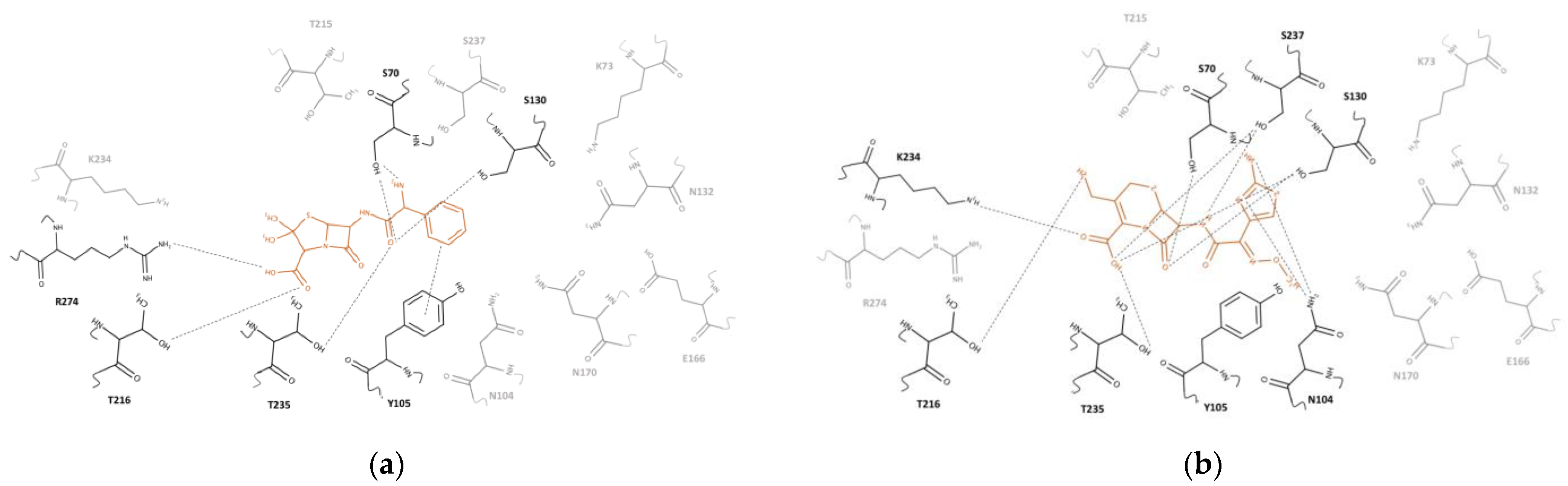
2.2. Clavulanic Acid Complex
2.3. Ampicillin Complex
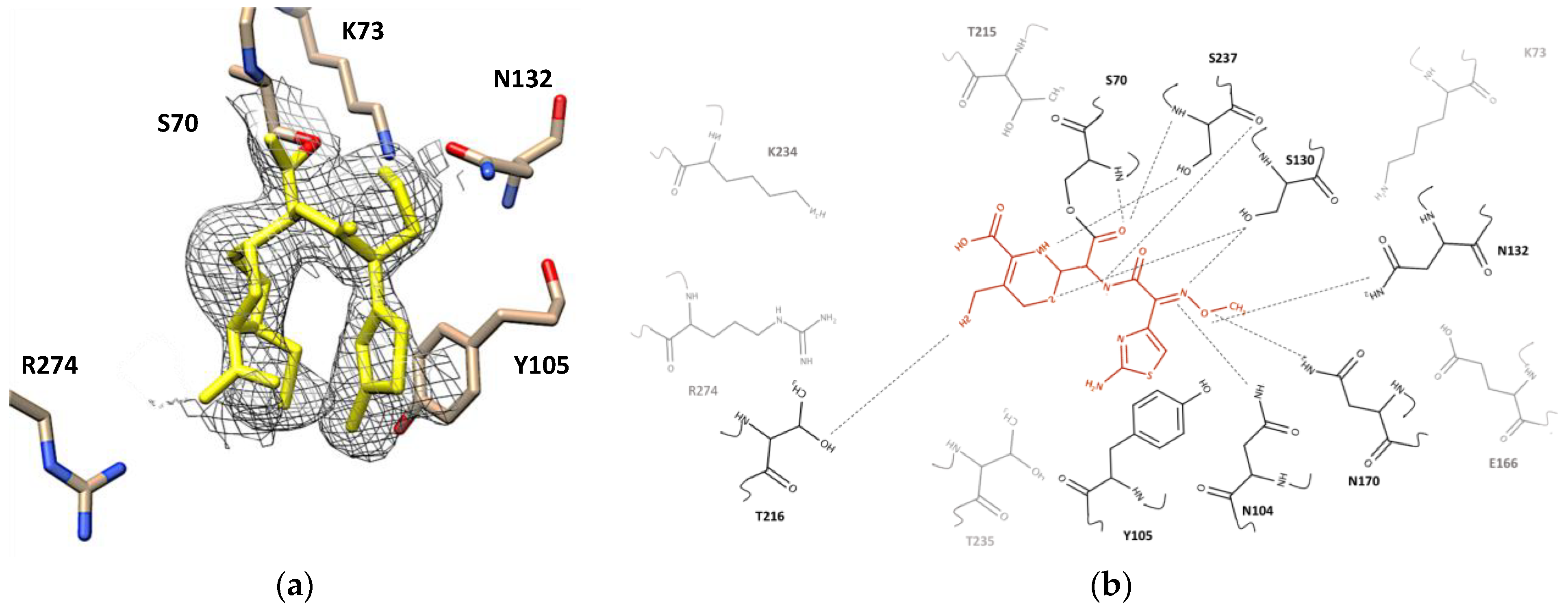
2.4. Desfuroylceftiofur (DFC) Complex
2.5. Steady-State Kinetics of CTX-M-15
2.6. Isothermal Titration Calorimetry
2.7. Molecular Docking
3. Discussion
3.1. Structural, Kinetic, and Thermodynamic Analysis for Complex Formation
3.2. Enzyme Catalytic Mechanism and Inhibition Mechanism
3.2.1. Hydrolysis Mechanism
3.2.2. Inhibition Mechanism
4. Materials and Methods
4.1. Accession Numbers
4.2. Chemicals and General Reagents
4.3. Expression and Purification of Recombinant Protein
4.4. Crystallization
4.5. Structure Determination
4.6. Site-Directed Mutagenesis
4.7. Steady-State Kinetics Experiment
4.8. Isothermal Calorimetry Titrations (ITC)
4.9. Molecular Docking
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC. Antibiotic Resistance Threats in the United States; U.S. Department of Health and Human Services, Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019.
- Jacoby, G.A.; Munoz-Price, L.S. The New Beta-Lactamases. N. Engl. J. Med. 2015, 352, 380–391. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, M.M.; Arena, F.; Pallecchi, L.; Rossolini, G.M. CTX-M-Type Beta-Lactamases: A Successful Story of Antibiotic Resistance. Int. J. Med. Microbiol. 2013, 303, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Mulvey, M.R.; Bryce, E.; Boyd, D.; Ofner-Agostini, M.; Christianson, S.; Simor, A.E.; Paton, S. Ambler Class A Extended-Spectrum Beta-Lactamase-Producing Escherichia coli and Klebsiella spp. in Canadian Hospitals. Antimicrobe Agents Chemother. 2004, 48, 1204–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moland, E.S.; Black, J.A.; Hossain, A.; Hanson, N.D.; Thomson, K.S.; Pottumarthy, S. Discovery of CTX-M-like Extended-Spectrum Beta-Lactamases in Escherichia coli Isolates from Five U.S. States. Antimicrobe Agents Chemother. 2003, 47, 2382–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanheira, M.; Mendes, R.E.; Rhomberg, P.R.; Jones, R.N. Rapid Emergence of BlaCTX-M among Enterobacteriaceae in U.S. Medical Centers: Molecular Evaluation from the MYSTIC Program (2007). Microb Drug Resist. 2008, 14, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Hasan, F.; Ahmed, S.; Hameed, A. Extended-Spectrum Beta-Lactamases (ESbLs): Characterization, Epidemiology and Detection. Crit. Rev. Microbiol. 2004, 30, 25–32. [Google Scholar] [CrossRef]
- Zhao, W.H.; Hu, Z.Q. Epidemiology and Genetics of CTX-M Extended-Spectrum Beta-Lactamases in Gram-Negative Bacteria. Crit. Rev. Microbiol. 2013, 39, 79–101. [Google Scholar] [CrossRef]
- Livermore, D.M. The β-Lactamase Threat in Enterobacteriaceae, Pseudomonas and Acinetobacter. Trends Microbiol. 2006, 14, 413–420. [Google Scholar] [CrossRef]
- Reading, C.; Cole, M. ClavulanicAcid: A Beta-Lactamase-Inhibiting Beta-Lactam from Streptomyces Clavuligerus. Am. Soc. Microbiol. 1977, 11, 852–857. [Google Scholar]
- Sulton, D.; Pagan-Rodriguez, D. Clavulanic Acid Inactivation of SHV-1 and the Inhibitor-Resistant S130G SHV-1 b-Lactamase. J. Biol. Chem. 2005, 280, 33528–33536. [Google Scholar] [CrossRef] [Green Version]
- Lovering, A.L.; Safadi, S.S.; Strynadka, N.C. Structural Perspective of Peptidoglycan Biosynthesis and Assembly. Ann. Rev. Biochem. 2012, 81, 451–478. [Google Scholar] [CrossRef] [PubMed]
- Pazos, M.; Peters, K. Peptidoglycan Bacterial Cell Walls and Membranes; Springer: Berlin/Heidelberg, Germany, 2019; pp. 127–168. [Google Scholar]
- Rang, H.P.; Dale, M.M.; Ritter, J.M.; Flower, R.J. Drugs Used in the Treatment of Infections and Cancer and Antibacterial Drugs. In Textbook of Pharmacology, 6th ed.; Church Hill Living Stone: London, UK, 2006; pp. 650–665. [Google Scholar]
- Fisher, J.F.; Meroueh, S.O.; Mobashery, S. Bacterial Resistance to Beta-Lactam Antibiotics: Compelling Opportusism, Compelling Opportunity. Chem. Rev. 2005, 105, 395–424. [Google Scholar] [CrossRef] [PubMed]
- Giedraitiene, A.; Vitkauskiene, A.; Naginiene, R.; Pavilonis, A. Antibiotic Resistance Mechanisms of Clinically Important Bacteria. Medicina 2011, 47, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Meroueh, S.O.; Minasov, G.; Lee, W.; Shoichet, B.K.; Mobashery, S. Structural Aspects for Evolution of Beta-Lactamases from Penicillin-Binding Proteins. J. Am. Chem. Soc. 2003, 125, 9612–9618. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A. AmpC Beta-Lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitout, J.D.; Laupland, K.B. Extended-Spectrum b-Lactamase-Producing Enterobacteriaceae: An Emerging Public Health Concern. Lancet Infect. Dis. 2008, 8, 159–166. [Google Scholar] [CrossRef]
- Paterson, D.L.; Bonomo, R.A. Extended-Spectrum Beta-Lactamases: A Clinical Update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [Green Version]
- Thomson, J.M.; Bonomo, R.A. The Threat of Antibiotic Resistance in Gram-Negative Pathogenic Bacteria: β-Lactams in Peril. Curr. Opin. Microbiol. 2005, 8, 518–524. [Google Scholar] [CrossRef]
- Nikaido, H.; Pagès, J.M. Broad-Specificityeffluxpumpsand Their Role in Multidrug Resistance of Gram-Negative Bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [Green Version]
- Delaire, M.; Lenfant, F.; Labia, R.; Masson, J.M. Site-Directed Mutagenesis on TEM-1 β-Lactamase: Role of Glu166 in Catalysis and Substrate Binding. Protein Eng. 1991, 4, 805–810. [Google Scholar] [CrossRef]
- Trynadka, N.C.J.; Adachi, H.; Jensen, S.E.; Johns, K.; Sielecki, A.; Betzel, C. Molecular Structure of the Acyl-Enzyme Intermediate in β-Lactam Hydrolysis at 1.7 Å Resolution. Nature 1992, 359, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Minasov, G.; Wang, X.; Shoichet, B.K. An Ultrahigh Resolution Structure of TEM-1 Beta-Lactamase Suggests a Role for Glu166 as the General Base in Acylation. J. Am. Chem. Soc. 2002, 124, 5333–5340. [Google Scholar] [CrossRef] [PubMed]
- Katsung, B.G.; Masters, S.B.; Trevor, A.J. Beta Lactam and Other Cell Wall- and Membrane-Active Antibiotics: Textbook on Basics and Clinical Pharmacology, 11th ed.; Tata Mcgraw Hill Education Private Limited: New York, NY, USA, 2009; p. 775. [Google Scholar]
- Palzkill, T. Metallo-β-Lactamase Structure and Function. Ann. N. Y. Acad. Sci. 2013, 1277, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical Relevance of the ESKAPE Pathogens. Expert Rev. Anti Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. b-Lactams and b-Lactamase Inhibitors: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, A025247. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Fisher, J.F. Epidemiological Expansion, Structural Studies, and Clinical Challenges of New β-Lactamases from Gram-Negative Bacteria. Ann. Rev. Microbiol. 2011, 65, 455–478. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated Functional Classification of b-Lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.A.; Sischo, W.M.; Jones, L.P.; Moore, D.A.; Ahmed, S.; Short, D.M.; Besser, T.E. Recent Emergence of Escherichia coli with Cephalosporin Resistance Conferred by BlaCTX-M on Washington State Dairy Farms. Appl. Environ. Microbiol. 2015, 81, 4403–4410. [Google Scholar] [CrossRef] [Green Version]
- Hugonnet, J.E.; Blanchard, J.S. Irreversible Inhibition of the Mycobacterium Tuberculosis β-Lactamase by Clavulanate. Biochemistry 2007, 46, 11998–12004. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Delmas, J.; Sirot, J.; Shoichet, B.; Bonnet, R. Atomic Resolution Structures of CTX-M β-Lactamases: Extended Spectrum Activities from Increased Mobility and Decreased Stability. J. Mol. Biol. 2005, 348, 349–362. [Google Scholar] [CrossRef]
- Kuzin, A.P.; Nukaga, M.; Nukaga, Y.; Hujer, A.M.; Bonomo, R.A.; Knox, J.R. Structure of the SHV-1 β-Lactamase. Biochemistry 1999, 38, 5720–5727. [Google Scholar] [CrossRef] [PubMed]
- Stec, B.; Holtz, K.M.; Wojciechowski, C.L.; Kantrowitz, E.R. Structure of the Wild-Type TEM-1 [Beta]-Lactamase at 1.55 A and the Mutant Enzyme Ser70Ala at 2.1 A Suggest the Mode of Noncovalent Catalysis for the Mutant Enzyme. Acta Crystallogr. Sect. D 2005, 61, 1072–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahiri, S.D.; Mangani, S.; Reville, T.D.; Benvenuti, M.; Luca, F.D.; Sanyal, G.; Docquier, J.D. Structural Insight into Potent Broad-Spectrum Inhibition with Reversible Recyclization Mechanism: Avibactam in Complex with CTX-M-15 and Pseudomonas Aeruginosa AmpC b-Lactamases. Antimicrob. Agents Chemother. 2013, 57, 2496–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michio, I.; Noriyoshi, I. Hypervalent Nonbonded Interactions of a Divalent Sulfur Atom. Implications in Protein Architecture and the Functions. Molecules 2012, 17, 7266–7283. [Google Scholar]
- Cantón, R.; Coque, T.M. The CTX-M β-Lactamase Pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Carter, P.; Wells, J.A. Dissecting the Catalytic Triad of a Serine Protease. Nature 1988, 332, 564–568. [Google Scholar] [CrossRef] [PubMed]
- Mader, M.M.; Bartlett, P.A. Binding Energy and Catalysis: The Implications for Transition-State Analogs and Catalytic An-Tibodies. Chem. Rev. 1997, 97, 1281–1302. [Google Scholar] [CrossRef]
- Hornish, R.E.; Katarski, S.F. Cephalosporins in Veterinary Medicine—Ceftiofur Use in Food Animals. Curr. Top. Med. Chem. 2002, 2, 717–731. [Google Scholar] [CrossRef]
- Schuler, A.M.; Rice, C.; Gorden, P.J. Survey of Treatment Practices on Midwest Dairy Farms. Bov. Pract. 2017, 51, 190–199. [Google Scholar]
- Beyer, A.; Baumann, S.; Scherz, G.; Stahl, J.; Von Bergen, M.; Friese, A.; Roesler, U.; Kietzmann, M.; Honscha, W. Effects of Ceftiofur Treatment on the Susceptibility of Commensal Porcine E. Coli—Comparison between Treated and Untreated Ani-Mals Housed in the Same Stable. BMC Vet. Res. 2015, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mclaughlin, C.L.; Stanisiewski, E.P.; Risco, C.A.; Santos, J.E.P.; Dahl, G.E.; Chebel, R.C.; LaGrow, C.; Daugherty, C.; Bryson, L.; Weigel, D. Evaluation of Ceftiofur Crystalline Free Acid Sterile Suspension for Control of Metritis in High-Risk Lac-Tating Dairy Cows. Theriogenology 2013, 79, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Vandavasi, V.G.; Weiss, K.L.; Cooper, J.B.; Erskine, P.T.; Tomanicek, S.J.; Ostermann, A.; Schrader, T.E.; Ginell, S.L.; Coates, L. Exploring the Mechanism of Beta-Lactam Ring Protonation in the Class A Beta-Lactamase Acylation Mechanism Using Neutron and X-ray Crystallography. J. Med. Chem. 2016, 59, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, L.W.; Fan, F.; Blanchard, J.S. Biochemical and Structural Characterization of Mycobacterium Tuberculosis β-Lactamase with the Carbapenems Ertapenem and Doripenem. Biochemistry 2010, 49, 3766–3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avillan, J.; Ahmadvand, P.; Lu, S.Y.; Horton, J.; Liu, J.; Lofgren, E.T.; Davis, M.; Kang, C.H.; Call, D. Excreted Antibiotics May Be Key to Emergence of Increasingly Efficient Antimicrobial Resistance. Appl. Environ. Microbiol. 2022; under review. [Google Scholar]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A Comprehensive Python-Based System for Macromolecular Structure Solution. Acta Cryst. D Biol. Cryst. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and Development of Coot. Acta Cryst. D Biol. Cryst. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Apo-CTX-M-15 (7U57) | Clavulanic Acid-CTX-M-15 (7U48) | Ampicillin-CTX-M-15 (7U4B) | DFC-CTX-M-15 (7U49) | |
|---|---|---|---|---|
| Data collection | ||||
| Space group | C 1 2 1 | C 1 2 1 | C 1 2 1 | C 1 2 1 |
| Cell dimensions | ||||
| a, b, c (Å) | 171.079, 51.033, 106.863 | 170.692, 50.976, 106.699 | 170.663, 50.9, 106.859 | 170.955, 50.933, 106.597 |
| α, β, γ (°) | 90.00, 112.589, 90.00 | 90.00, 113.232, 90.00 | 90.00, 112.535, 90.00 | 90.00, 113.446, 90.00 |
| Resolution (Å) | 51.71–2.37 (2.455–0.37) | 48.48–1.67 (1.73–1.67) | 49.35–1.92 (1.989–1.92) | 48.9–1.96 (2.03–1.96) |
| Rmeas | 0.2228 (1.029) | 0.185 (1.579) | 0.0815 (0.2692) | 0.1696 (1.378) |
| Rpim | 0.08877 (0.4186) | 0.1012 (0.9726) | 0.0326 (0.1081) | 0.0664 (0.5471) |
| Wavelength (Å) | 1.000 | 1.000 | 1.000 | 1.000 |
| Unique reflections | 34970 (3456) | 83673 (2177) | 64491 (6470) | 60844 (3489) |
| Completeness (%) | 99.09 (96.59) | 99.08 (92.07) | 99.54 (98.11) | 91.95 (57.41) |
| <I>/σI | 4.49 (0.68) | 6.27 (0.35) | 11.04 (1.04) | 5.29 (0.49) |
| CC1/2 | 0.988 (0.704) | 0.975 (0.287) | 0.998 (0.977) | 0.996 (0.6) |
| Redundancy | 6.2 (5.9) | 3.0 (1.7) | 6.0 (5.7) | 6.4 (6.1) |
| Refinement | ||||
| Rwork/Rfree | 0.2228/0.3239 (0.2598/0.3449) | 0.2119/0.2699 (0.4972/0.6601) | 0.1870/0.2508 (0.2541/0.3326) | 0.2400/0.3162 (0.3901/0.4214) |
| Number of atoms | ||||
| Protein and ligand | 6201 | 6439 | 6899 | 6199 |
| Water | 195 | 174 | 117 | 127 |
| B-factors (Å2) | ||||
| All atoms | 42.3 | 37.66 | 28.27 | 40.53 |
| Solvent | 42.15 | 40.53 | 35.8 | 38.88 |
| R.m.s deviations | ||||
| Bonds (Å) | 0.013 | 0.013 | 0.012 | 0.015 |
| Angles (°) | 1.37 | 1.28 | 1.24 | 1.41 |
| Ramachandrans | ||||
| % Favored | 92.88 | 95.93 | 95.93 | 91.86 |
| % Outliers | 0.76 | 0.76 | 1.15 | 0.42 |
| Clashscore | 18.32 | 8.78 | 9.98 | 15.76 |
| Substrate | β-Lactamase | Vmax (μmol/s) | Km (μM) | kcat (s−1) | kcat/Km (μM−1 s−1) |
|---|---|---|---|---|---|
| Nitrocefin | WT-CTX-M-15 | 3.38 ± 0.12 | 1047.0 ± 75.8 | 338.3 ± 16.4 | 3.23 ± 0.02 |
| S70A-CTX-M-15 | 0.031 ± 0.003 | 45.6 ± 1.38 | 3.15 ± 0.17 | 0.07 ± 0.00 | |
| Ceftiofur | WT-CTX-M-15 | 3.14 ± 0.07 | 330.0 ± 14.1 | 313.7 ± 13.0 | 0.95 ± 0.01 |
| S70A-CTX-M-15 | 0.027 ± 0.00 | 35.0 ± 0.42 | 2.73 ± 0.08 | 0.08 ± 0.00 | |
| Ampicillin | WT-CTX-M-15 | 1.01 ± 0.06 | 197.8 ± 13.0 | 101.9 ± 5.6 | 0.52 ± 0.01 |
| S70A-CTX-M-15 | 0.007 ± 0.00 | 26.4 ± 3.7 | 0.67 ± 0.04 | 0.03 ± 0.00 | |
| DFC | WT-CTX-M-15 | 4.89 ± 0.2 | 141.2 ± 11.2 | 489.8 ± 16.2 | 3.46 ± 0.14 |
| S70A-CTX-M-15 | 0.006 ± 0.00 | 7.8 ± 0.03 | 0.59 ± 0.02 | 0.08 ± 0.00 |
| S70A-CTX-M-15 | Kd (μM) | ΔH (cal mol−1) | ΔS (cal mol−1 K−1) | ΔG (kcal mol−1) |
|---|---|---|---|---|
| Clavulanic acid | 288.6 | −1333 | 11.7 | −4.8 |
| Nitrocefin | 71.3 | −2025 | 12.1 | −5.7 |
| Ceftiofur | 38.2 | −1157 | 16.3 | −6.0 |
| Ampicillin | 28.8 | −509 | 19.1 | −6.2 |
| DFC | 10.7 | −2246 | 15.2 | −6.7 |
| Substrate | ΔG Binding CTX-M-15 (kcal mol−1) | ΔG Binding CTX-M-15 S70A (kcal mol−1) |
|---|---|---|
| Clavulanic acid | −6.2 | −6.0 |
| Nitrocefin | −6.7 | −7.4 |
| Ceftiofur | −6.8 | −7.6 |
| Ampicillin | −6.9 | −7.4 |
| DFC | −7.5 | −8.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmadvand, P.; Avillan, J.J.; Lewis, J.A.; Call, D.R.; Kang, C. Characterization of Interactions between CTX-M-15 and Clavulanic Acid, Desfuroylceftiofur, Ceftiofur, Ampicillin, and Nitrocefin. Int. J. Mol. Sci. 2022, 23, 5229. https://doi.org/10.3390/ijms23095229
Ahmadvand P, Avillan JJ, Lewis JA, Call DR, Kang C. Characterization of Interactions between CTX-M-15 and Clavulanic Acid, Desfuroylceftiofur, Ceftiofur, Ampicillin, and Nitrocefin. International Journal of Molecular Sciences. 2022; 23(9):5229. https://doi.org/10.3390/ijms23095229
Chicago/Turabian StyleAhmadvand, Parvaneh, Johannetsy J. Avillan, Jacob A. Lewis, Douglas R. Call, and ChulHee Kang. 2022. "Characterization of Interactions between CTX-M-15 and Clavulanic Acid, Desfuroylceftiofur, Ceftiofur, Ampicillin, and Nitrocefin" International Journal of Molecular Sciences 23, no. 9: 5229. https://doi.org/10.3390/ijms23095229
APA StyleAhmadvand, P., Avillan, J. J., Lewis, J. A., Call, D. R., & Kang, C. (2022). Characterization of Interactions between CTX-M-15 and Clavulanic Acid, Desfuroylceftiofur, Ceftiofur, Ampicillin, and Nitrocefin. International Journal of Molecular Sciences, 23(9), 5229. https://doi.org/10.3390/ijms23095229