3. Discussion
Persons suffering from celiac disease, wheat allergy, and non-celiac gluten-sensitivity show very severe intestinal effects due to prolamins. Each milligram of gluten plays a decisive role for the well-being of such people. In particular, prolamins derived from wheat, barley, and rye cause the most severe problems. Other species of Poaceae (=Gramineae), however, may also contain gluten. Among them are spelt (
Triticum aestivum subsp.
spelta), Kamut (Khorasan wheat,
Triticum turgidum ×
polonicum), the generic hybrids of wheat and rye called
Triticale or
Triticumsecale and
Secalotri(ti)cum, as well as einkorn (
Triticum monococcum) and emmer (
Tritium diococcum), both of which are ancient grains that are very popular in modern diets [
6,
18]. The processing of all of these crops has a negligible impact on the hazardous effect of the gluten proteins. Avoiding those irritants by strictly following a gluten-free diet is of crucial relevance, as there is no cure for these diseases yet. However, traces of prolamins can be found even in products classified as gluten free, and, therefore, gluten cannot be completely avoided by affected individuals. Several steps, derived from agricultural techniques from harvesting to food processing, can be responsible for the contamination [
6]. Thus, a dietary supplement binding the toxic plant peptides might be of great relevance for patients suffering from gluten proteins. This approach is particularly important because the gluten-free diet offers fewer vitamins and minerals, when compared to conventional food consumption. Therefore, slightly less-restrictive nutrition could offer great benefits concerning diet-related deficiency signs. Moreover, trapping triggers might help patients to recover and lead less painful lives.
For all experiments presented in this study, a special purified clinoptilolite-tuff (PCT), intensively tested and approved for prolonged human application [
14,
15,
19], was used. Due to a patented purification process, PCT is free of toxic elements and safe for supplemental oral intake [
13].
Clinoptilolite belongs to the family of natural zeolites, and has been used for decades for farm animals as well as for pets. In the European Union, clinoptilolite-tuff is allowed as feed additive [
12,
16,
17,
20]. In the United States of America, the raw material of PCT, clinoptilolite of sedimentary origin, is generally recognized as safe “for use as an anticaking agent in diets for cattle, swine, goats, sheep, poultry, cats and dogs at a level up to 1% by weight in complete diets” (GRAS Notice No. AGRN 29, 30 August 2019). Interestingly, the first specific tests on animals were performed in Japan in 1965, when natural zeolites were used as dietary supplements for ruminants, swine, and poultry. Subsequent scientific investigations showed a significant increase in feed efficiency and in the general health of livestock, with no apparent side effects [
12,
21,
22]. Various studies applying low doses of zeolite-tuff (0.2–5% clinoptilolite-tuff/kg feed), combined with mycotoxin-contaminated feed, revealed remarkable results by preventing mycotoxin-related issues (mycotoxicoses; [
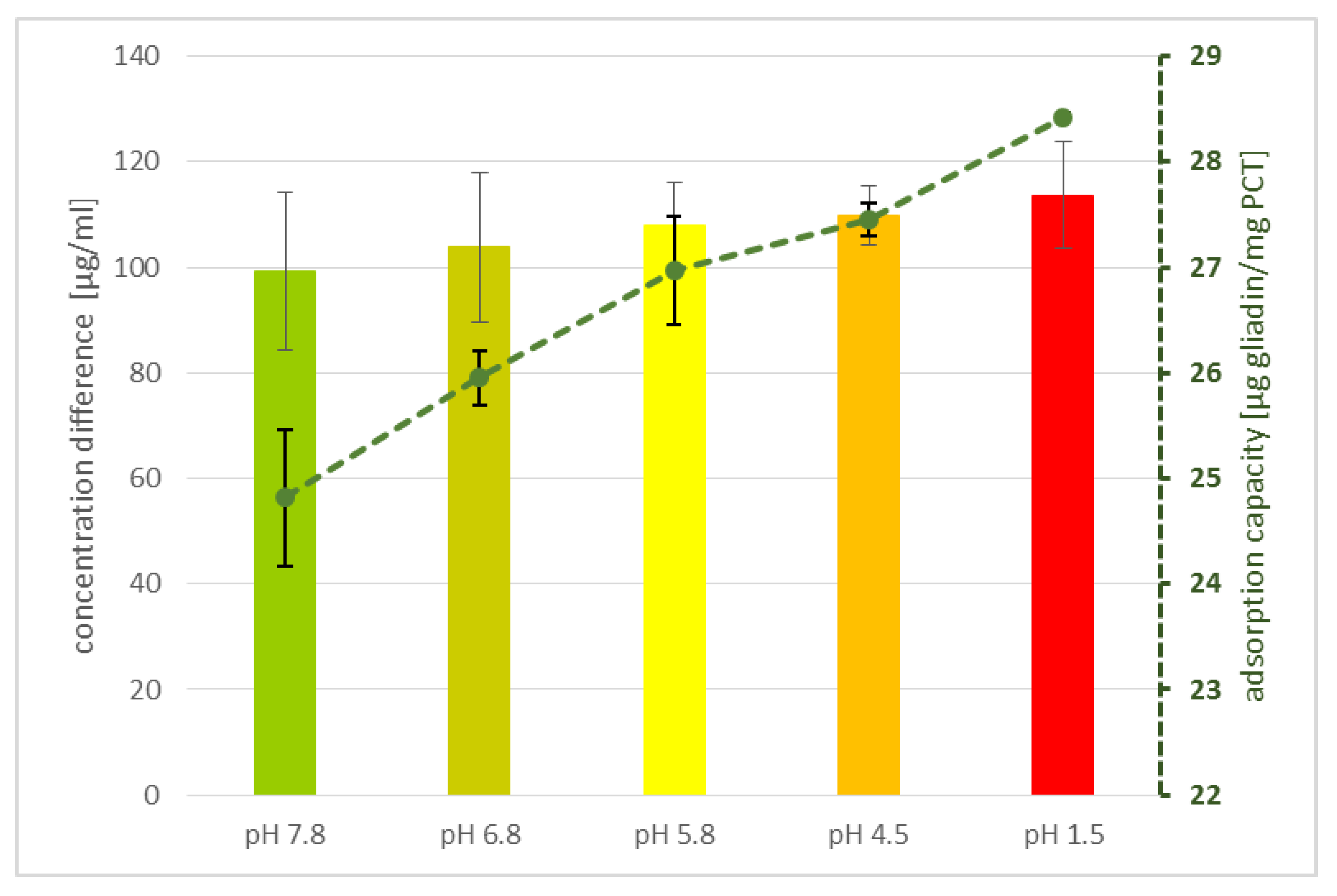
22]). For an average person with 80 kg bodyweight, a daily application of 2 g PCT corresponds to 0.0025% PCT/kg, compared with an animal receiving 0.2–5% zeolite-tuff/kg at each feed. The in vitro experiments of this study showed a binding capacity of more than 20 mg gliadin per gram of PCT, which accords with a daily application of 2 g PCT for 40 mg gliadin, or an 80 mg gluten minimum-sorption capacity. Taking into consideration that less than 20 µg/g are defined as “gluten-free” (≙10 µg/g gliadin), 2 g of PCT can bind traces of prolamins sufficiently and could contribute to a safe diet.
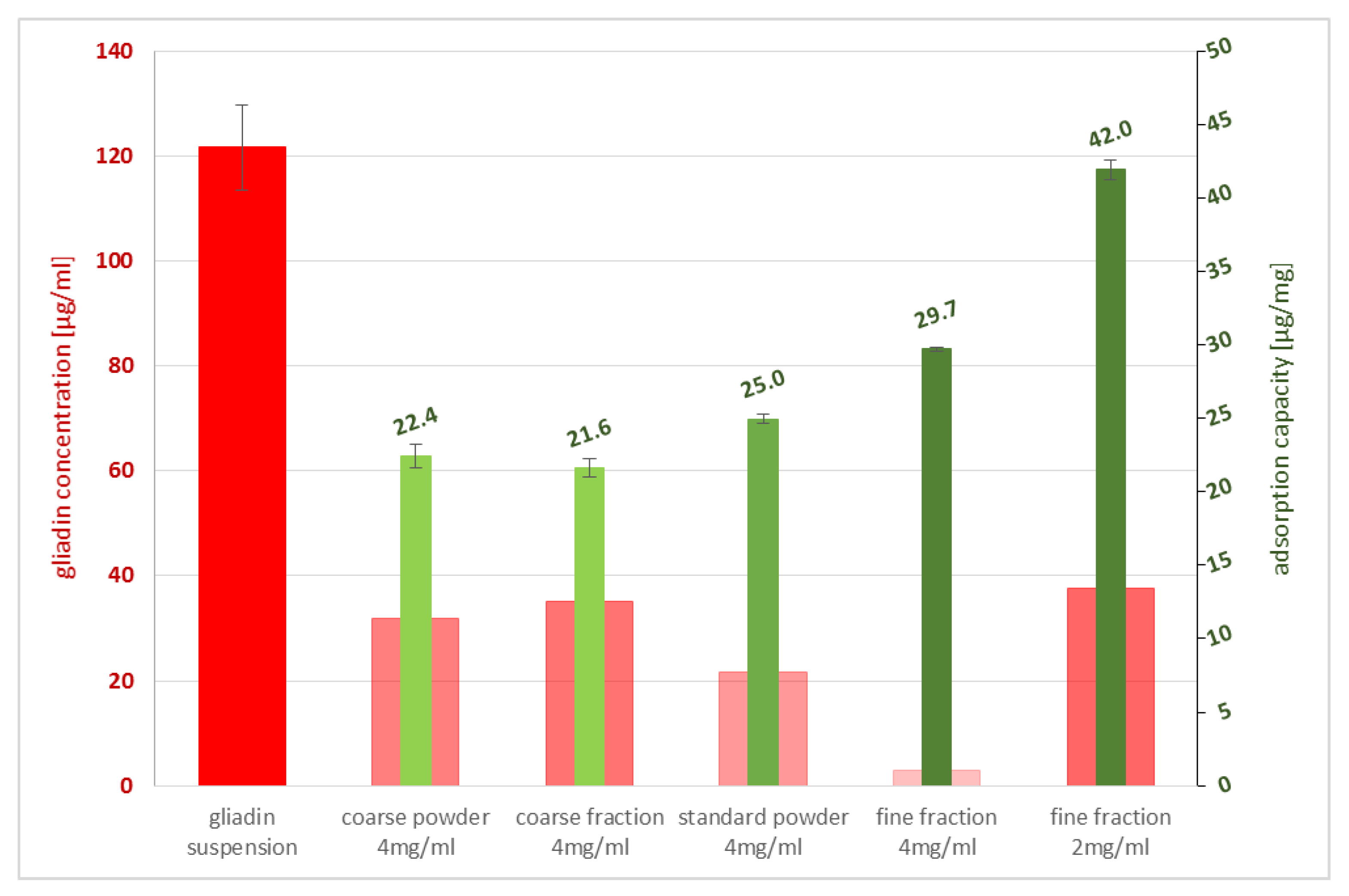
The size of a particle’s surface is directly related to its adsorption capacity—the larger the surface, the higher the capability of sorption. Studying materials with varying particle-size distributions confirmed the highest gluten sorption capacity for the finest grain-size fraction. Hence, milling is a crucial step during the manufacturing of zeolite-based materials, as high-impact grinding tends to destroy the clinoptilolite’s crystal framework, and therefore lowers its efficacy. PCT is micronized with a special jetmill technique, resulting in a homogenous, narrow, and fine-grained range of particle-sizes with fully intact crystal structures; this allows the finished product to have the optimal binding characteristics. The lowest sorption capacity was obtained by the coarse fraction, which was used for the calculation above to demonstrate the impressive quality of gluten adsorption even under non-optimal particle sizes of PCT. Due to the effective pore size of PCT, corresponding to 0.4 nm [
23], and our findings that the same weight of different fractions but divergent sizes of particles show incoherent sorption capacities, we strongly propose adsorption mechanisms, rather than absorption of prolamins onto PCT. This proposal is supported by the fact that the average particle size of gliadin is 3.88 nm, while that of hordein is 4.32 nm and that of secalin is 5.79 nm [
24]. In addition, in-house XRD measurements before and after adsorption of gluten to PCT revealed no changes in the diffraction pattern. This was expected, as the relatively big prolamin proteins are not capable of either entering the pores or the crystal lattice.
Zeolites have been applied for decades in storage mold management. Bodroža-Solarov et al. [
25] reported that the addition of zeolite to wheat dough through clinoptilolite-tuff containing residues in flour was problematic in changing the rheological properties of the wheat dough; they referred to a Russian patent that described the sensory properties of bread as deteriorating when exceeding 5% zeolite-tuff. Seen from this point of view, adding zeolite to prolamin-containing food seems to be inferior to oral administration. Moreover, the changing of the flow characteristics of the dough when zeolite is added reveals an action between the mineral and the gluten, as prolamins define the breadcrumb and are responsible for the texture and the elasticity of the dough. Casually speaking, gluten glues the ingredients of the dough together. This finding serves as a further indication of a specific binding of gluten to zeolite.
Another aspect of celiac disease is a deviation in the intestinal microflora, which is indicated by low diversity and aberrant composition [
1]. When analyzing various manifestations of celiac disease in biopsy samples of patients showing intestinal and extraintestinal (anemia, dermatitis herpetiformis = Duhring’s disease) forms, the expression in the gastrointestinal tract or anemia was linked to lower diversity, moreso than when dermatitis herpetiformis was the main clinical symptom. Patients with such dermatological characteristic had similar intestinal microbiota as control subjects [
26]. Another human study revealed that even treated celiac patients obeying a strict gluten-free diet showed persistent symptoms when their duodenial microbiota were altered. Proteobacteria were increased, and Bacterioidetes and Firmicutes were reduced, as their whole microbial richness was decreased [
27]. Interestingly, Prasai et al. described the effects of natural zeolite-enriched feedstuff in poultry, when cloacal swabs indicated a significant reduction of proteobacteria without influencing the beneficial bacteria [
28].
In this microbial context, bacteria—especially some of those isolated from curd and sourdough—are subjects of investigation regarding their ability to break down gluten by their metabolic activity. Scientific research on this topic started some decades ago, and deals with the definition of the special bacterial species responsible for this process, as high microbial diversity is found in both sourdough and curd [
29].
Bacillus sp. [
30], as well as species of Enterobacteriaceae and
Streptococcus [
31], were identified as showing proteolytic activity towards gluten proteins. These bacteria use gluten as carbon and nitrogen sources [
31]. Interestingly, Enterobacteriaceae belong to the group of proteobacteria, while the genera
Streptococcus,
Enterococcus, and
Lactobacillus are Firmicutes, which are described as advantageous for human health. The idea is either to administer gluten-hydrolyzing bacteria as probiotics (like
Bacillus sp.) to reduce the gluten content of food directly in the human digestive tract or to add them to ferment the gluten-containing dough [
30,
31,
32]. However, research is still at the pre-clinical level.
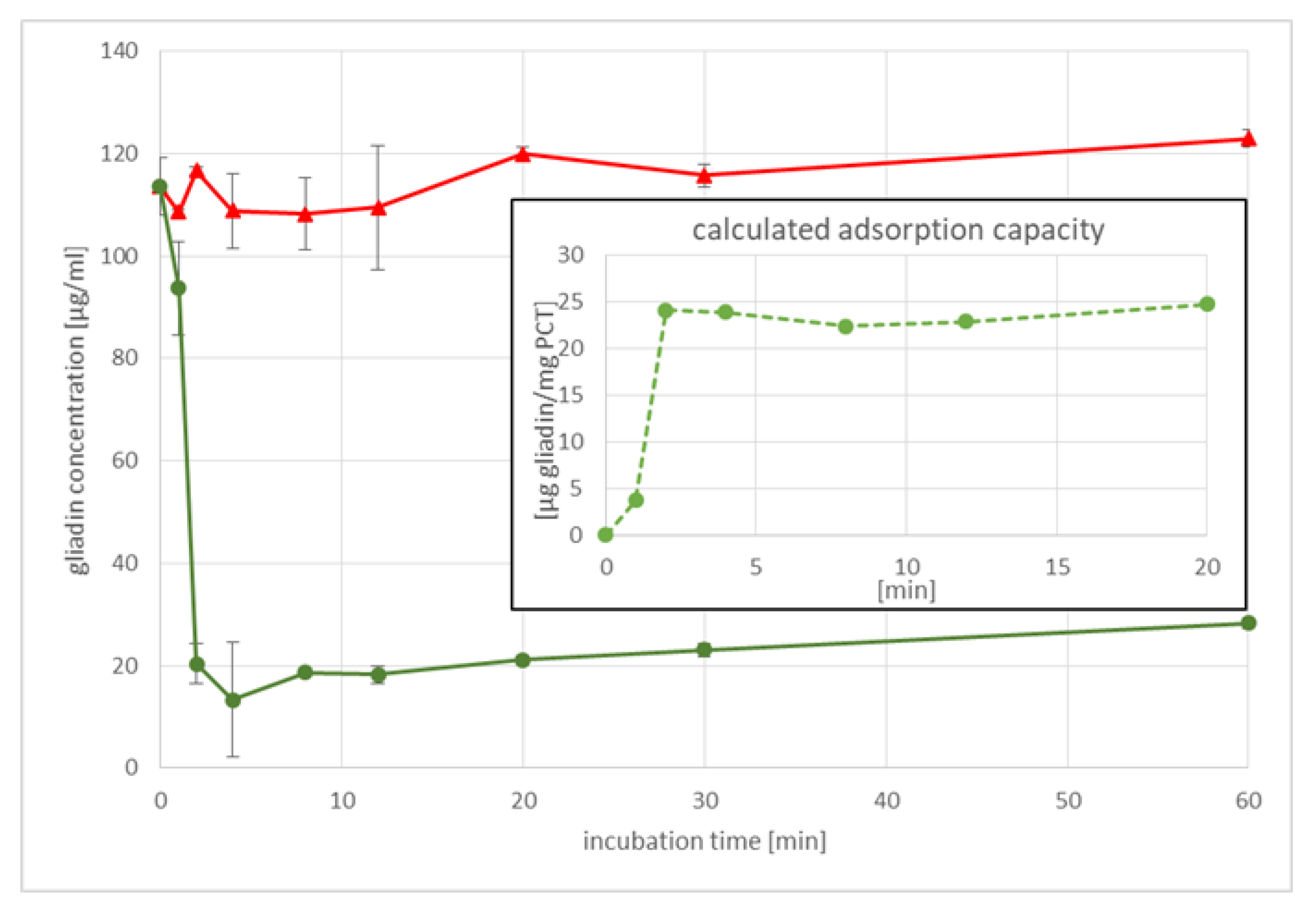
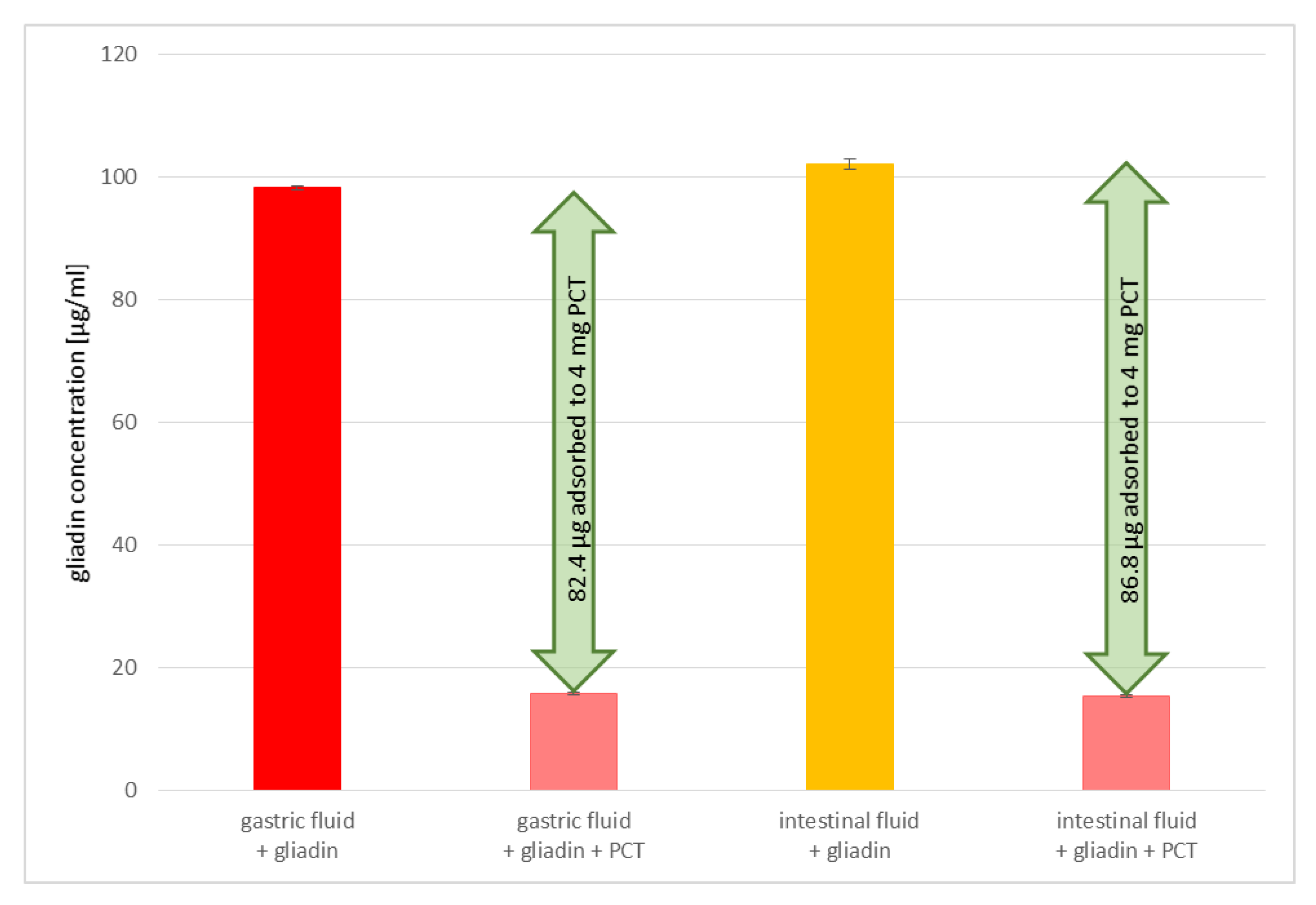
Contrary to the intended use of bacteria in gluten degradation, the intake of PCT is well-established and easy to handle. Our results demonstrate the constant efficiency of gluten binding under different conditions (stomach/intestine); the mineral remains sorptive for gluten at various pH levels and artificial fluids, while bacteria have to survive the low pH of the stomach to reach their target location in the intestine, where the pH is in the alkaline range. In addition to changing pH levels, bile acid is another factor that bacteria have to overcome, as high levels of it are found in the small intestine. Furthermore, most bacteria are stored best under refrigeration for only a certain amount of time; PCT, however, can be kept at room temperature and in a dry environment with, theoretically, no expiration date. Using bacteria in sourdough for gluten degradation, a certain incubation time and temperature are necessary to gain an acceptable reduction of gluten, a process that must be considered in terms of profitability. PCT not only binds gluten irreversibly, as the studies on sorption kinetics revealed, but it is also extremely fast; within the very first 4 min, gluten adhered to zeolite-tuff. This reduces the risk of gluten-induced damage caused by a long retention time in the intestine.
A further approach to break down gluten is the use of enzymes—derived from either bacteria, fungi, insects, humans, or plants—in the gastro-duodenal tract before reaching the small intestine [
33,
34]. Although the main interest is on endopeptidases, there are some exopeptidases documented in the literature, as well [
33]. Among the endopeptidases are glutenases, subtilisins, and propyl endopeptidases; synthetic glutenases are also created. Even human saliva is under investigation, as it contains a wide variety of bacteria, most of them descending from the phyla of Bacteroidetes and Actinobacteria [
34]. To be useful for oral enzyme therapy, the potential candidates must be safe for human consumption and cleave prolin- and glutamine-rich domains; their structural stability and enzymatic activity over a broad pH range (highly acidic in the stomach and mildly alkaline in the intestine) must be maintained; and, most importantly, no increase of immunogenicity by producing small immunogenic peptides may occur [
33,
34]. Enzymes must work fast, as gluten should be degraded before they can cause harm in the small intestine. The utilization of several different enzymes combined as a drug could result in a synergistic effect, as some enzymes are already active at low pH in the stomach, whereas others show highest efficacy in the duodenum, which would increase the degradation time [
33,
34]. In some cases, coating with PEG and microencapsulation with PLGA helps to retain the enzymes in their active state. Research also focusses on molecular modeling through genetic modification to transform natural proteins into more stable, pH-resistant enzymes with higher substrate specificity and increased activity. However, as many factors must be taken into consideration, obtaining acceptable results from clinical studies has proved to be a difficult task [
34]. An in vitro study by Tanner was published using nine commercially available digestive enzyme supplements to test their potential in the removal of celiac-relevant epitopes [
35]. The test conditions included two pH levels, at 3.5 and 7.0. Gliadin was isolated and purified from a single wheat cultivar (cv. Baxter) and afterwards digested with trypsin and pepsin to gain immunoactive peptides, as takes place in human digestion. The results demonstrated unequal activity and reaction times of the single supplements at the pH values tested, as well as strong to no activity in rendering gliadin that were harmless. Only one, an oleoresin- (caricain) containing supplement, could efficiently and rapidly degrade the trypsin/pepsin pre-digested gliadin at pH 7 [
35]. Sometimes, a mixture of enzymes must be applied, because only in combination can they offer their full, non-human-toxic action as described for
Pseudomonas/
Lactobacillus (while the first one cleaves gluten, resulting in smaller immunogenic peptides, the second one fragmentizes them into non-immunogenic gluten-residues) [
34] or
Aspergillus niger/
Aspergillus oryzae (aspergillo-pepsin and dipeptidyl peptidase IV cooperatively degrade gluten) [
33,
34]. Taking all noted factors into account, enzymes can provide help in the future for gluten-sensitive persons. Regarding the unrestricted gluten-binding capacity throughout the various pH levels in the whole intestinal tract, as tested in this work (starting at pH 1.5 and ending at pH 7.8), the PCT bears some significant advantages toward enzymes. Furthermore, the straightforward administration, without any indispensable coating, also contributes to PCT being favored over enzymes.
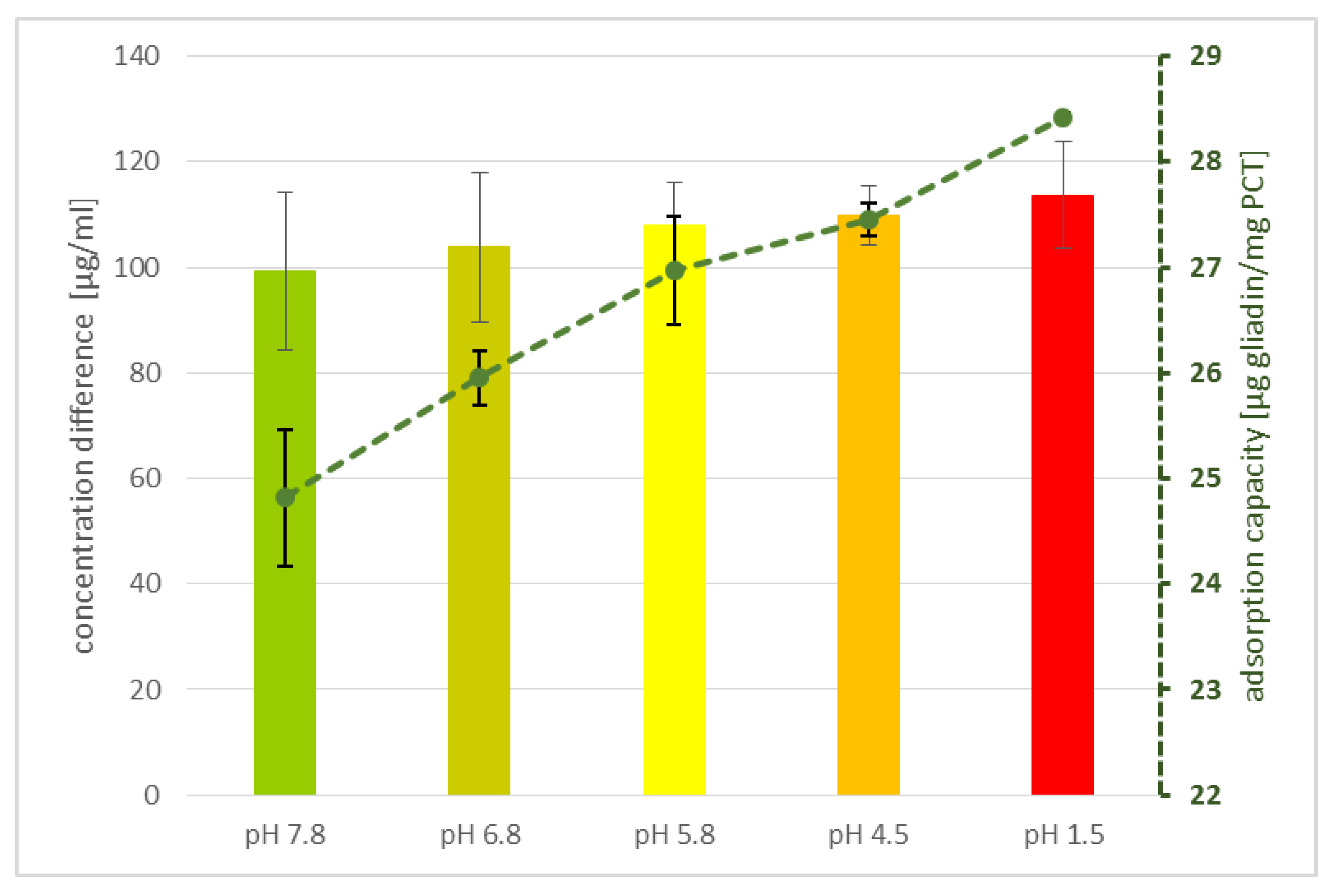
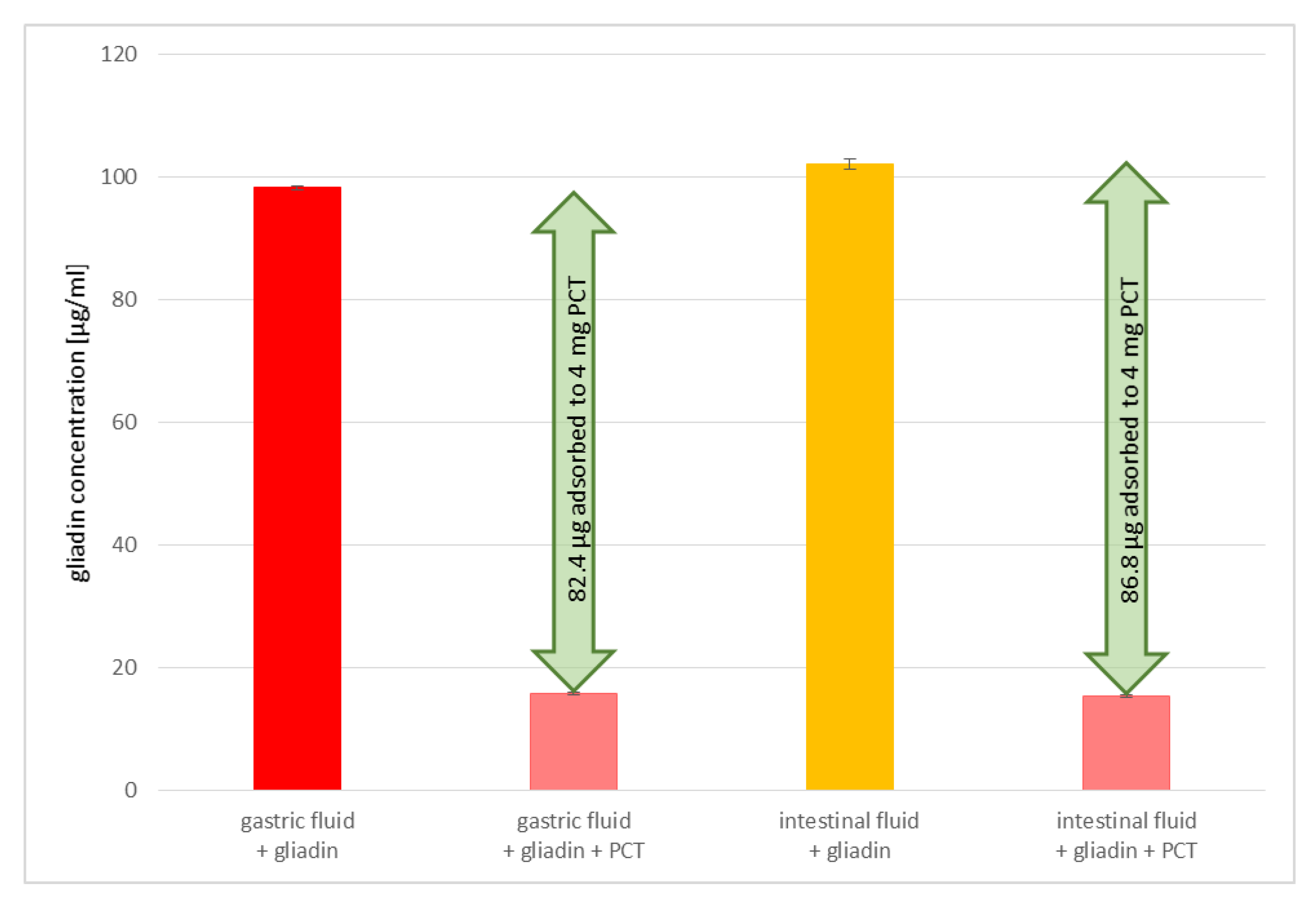
In addition, the adsorption of gluten to PCT seems to be relatively specific, as demonstrated in experiments with artificial fluids. Although there is a higher concentration of denaturated proteins (trypsin and pepsin, among others) than gluten in the solution, the binding of prolamins to PCT is not hindered, as revealed by ELISA. Perhaps a partial charge of the prolamins and/or their poor solubility in aqueous solution are responsible for the effect observed. Previous findings endorsed this presumption, as shown, e.g., for
Clostridioides difficile toxins A and B, which were bound to PCT and thus neutralized [
19]. Previously, it was known that these toxins have an affinity with an anionic resin called tolevamer, a polymer used for the treatment of
C. difficile-associated diarrhea in humans [
36]. Zeolites are composed of SiO
4 tetrahedra with an isomorphic substitution of aluminum ions, thereby inducing a negative framework charge and generating a cation-binding capacity [
37].
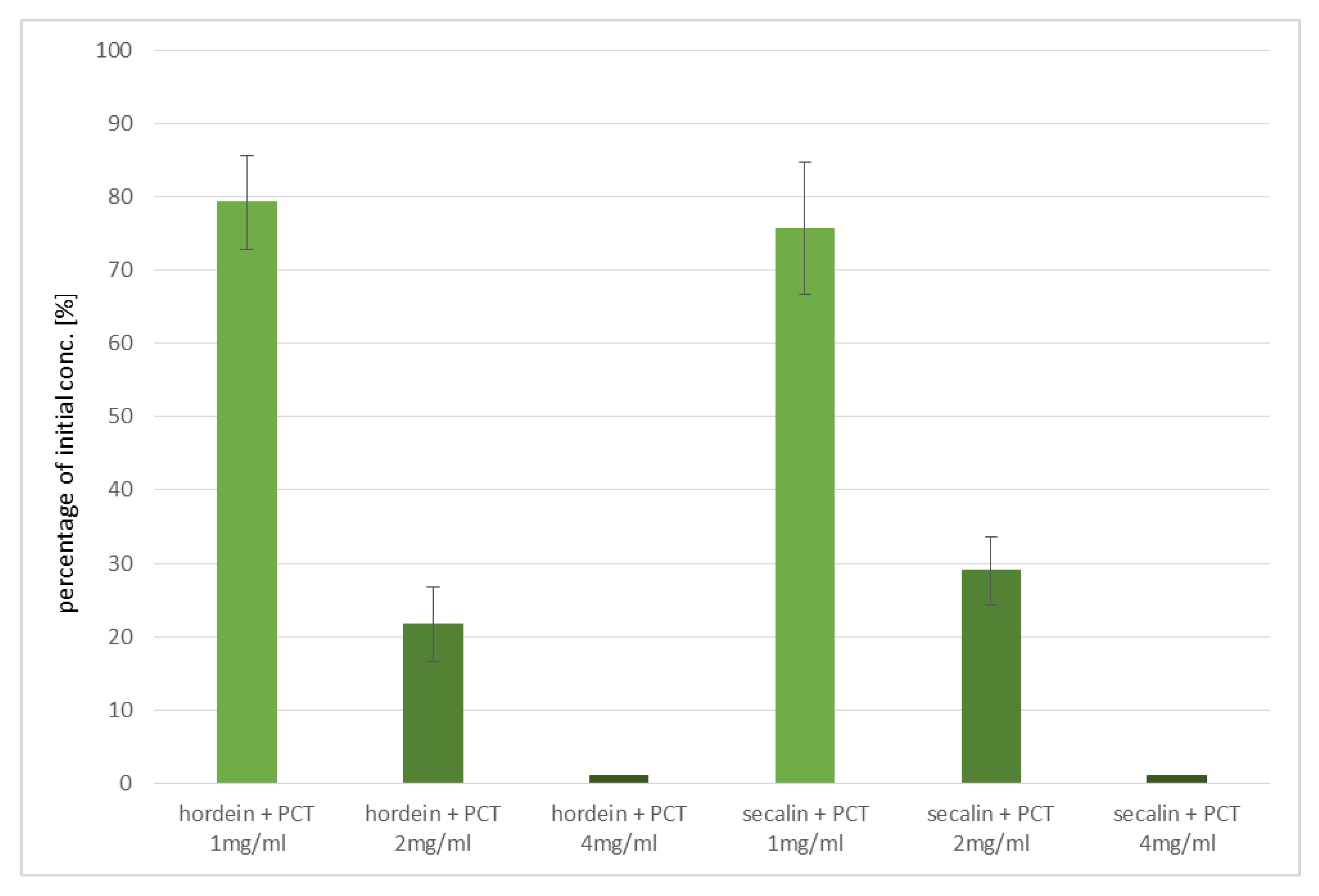
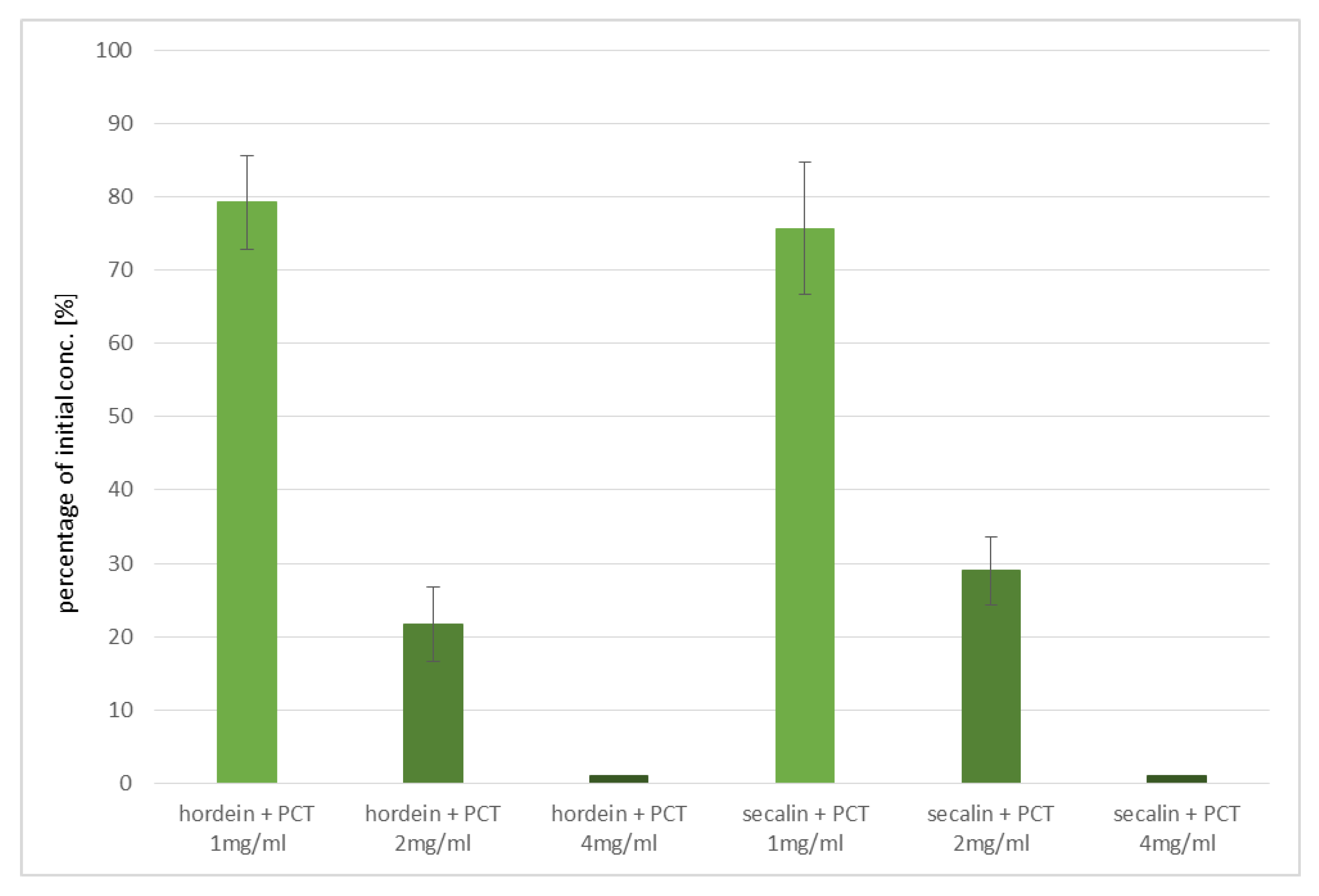
In the experiments performed, purified zeolite-tuff effectively sorbed the most celiac disease-critical prolamins of wheat (gliadins), barley (hordeins), and rye (secalins). As bread is often made of mixtures of wheat and rye, this result is a key advantage. Binding of barley’s hordeins is another asset of purified clinoptilolite-tuff, because pearl barley is a popular food. Since cross contaminations of the three different grains can occur easily during processing, a universal and non-specific prolamin-adsorbing capacity is a decisive positive factor in favor of PCT, especially since there are no negative side-effects documented, even when taken over longer periods.
Concerning the distinctive adsorption of the single prolamins, this effect might be based on their unique characteristics. Rani et al. vigorously studied the particular forms of prolamins of wheat, barley, and rye. They found, by applying scanning as well as transmission electron microscopy, that both the surface morphology and the shape of single gluten proteins differed specifically [
24]. Further, Lexhaller et al. compared prolamins and glutelins derived from wheat (cv. Akteur), barley (cv. Marthe), and rye (cv. Visello) with five sandwich ELISA kits [
38]. Thus, in addition to the different structures of gliadins, hordeins, and secalins [
24], inconsistent gluten values might be due to different affinities of the R5 monoclonal antibody of the ELISA kit used for detection, as shown by studies using various ELISA kits and the RP-HPLC technique. A higher sensitivity of the R5 monoclonal antibody toward secalins and hordeins, when compared to the sensitivity to gliadin, was demonstrated [
38].
A collaborative study concerning the “determination of gliadin as a measure of gluten in food by R5 Sandwich ELISA RIDASCREEN gliadin matrix extension” included 14 participating laboratories. The recovery rate was 80–130% and the relative reproducibility standard deviations for the contaminated samples were between 9.8% and 27.7% [
39]. In a review, Shewry noted that even wheat cultivars might give different gliadin results, as the number of epitopes for ELISA antibodies can vary among the single varieties when summarized in comparative studies of ancient and modern wheats. In fact, wheat samples grown under different conditions might exhibit other characteristics, and thus adverse effects for sensitive persons [
40]. Taking all these findings together, our results show the obviously prolamin-binding of PCT. Whether or not the higher adsorption values are attributable to the higher affinity of secalins and hordeins onto PCT, or are based on more binding sites (epitopes) than found in gliadin, is not deducible.
Taking all these considerations into account, our in vitro data clearly suggest that the intake of PCT could be highly beneficial for persons suffering from gluten-induced diseases. However, we are aware that only a clinical trial can thoroughly elaborate the positive effects of PCT administration on gluten-linked disorders.
4. Materials and Methods
4.1. The Chemical Composition of Buffers Used in the Experiments Performed
Buffers of different kinds were used for various experiments. In
Table 2, an overview of the specific buffers, including their single components, concentrations, and pH values, is provided (according to European Pharmacopoeia 10.0, Chapter 2.9.3, [
16]).
4.2. Synthetic Fluids Preparations
Both artificial fluids were prepared according to the recommendations of the European Pharmacopoeia [
16] and as noted in the EURL Evaluation Report [
17] on the Analytical Methods submitted in connection with the Application for Authorisation of a Feed Additive according to Regulation (EC) No 1831/2003 [
16,
17].
4.2.1. Preparation of Artificial Gastric Fluid
Pepsin (P7000, Sigma, Kawasaki, Japan) in the amount of 3.2 g was dissolved in 500 mL of distilled water prior to inactivation at 95 °C for 1 h. This was crucial in performing ELISA testing, as previous experiments revealed that pepsin altered the structures of prolamins so that they were no longer captured by the antibodies provided in the ELISA kit. Afterwards, 2.0 g NaCl (sodium chloride, 71380, Sigma-Aldrich, St. Louis, MO, USA) were added and dissolved before 80 mL of HCl [1 M] (hydrochloric acid 25%, CVH Chemie-Vertrieb GmbH & Co, Hannover, Germany) were pipetted to the solution. Finally, distilled water was added to reach a final volume of 1000 mL. The measured pH value was 1.3 at room temperature (pH 3210i, WTW).
4.2.2. Preparation of Artificial Intestinal Fluid
Pancreatin (P1750, Sigma, Kawasaki, Japan) in the amount of 10 g was dissolved in 500 mL distilled water before inactivation at 95 °C for 1 h. As described for the preparation of synthetic gastric fluid, this step was important for analyzing samples by ELISA. Next, 250 mL KH2PO4 [0.2 M] were mixed with 77 mL NaOH [0.2 M] (sodium hydroxide solution, ≥32%, extra pure, Carl Roth, Karlsruhe, Germany) and added to the dissolved pancreatin solution. Then, the pH was fixed to 6.8 and distilled water was used to reach an end volume of 1000 mL.
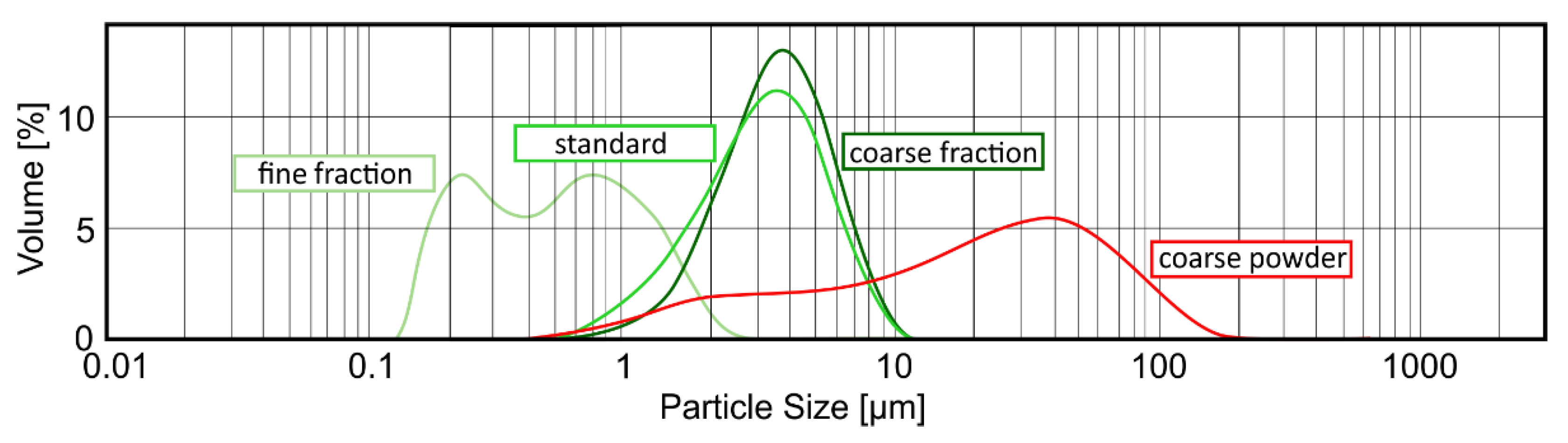
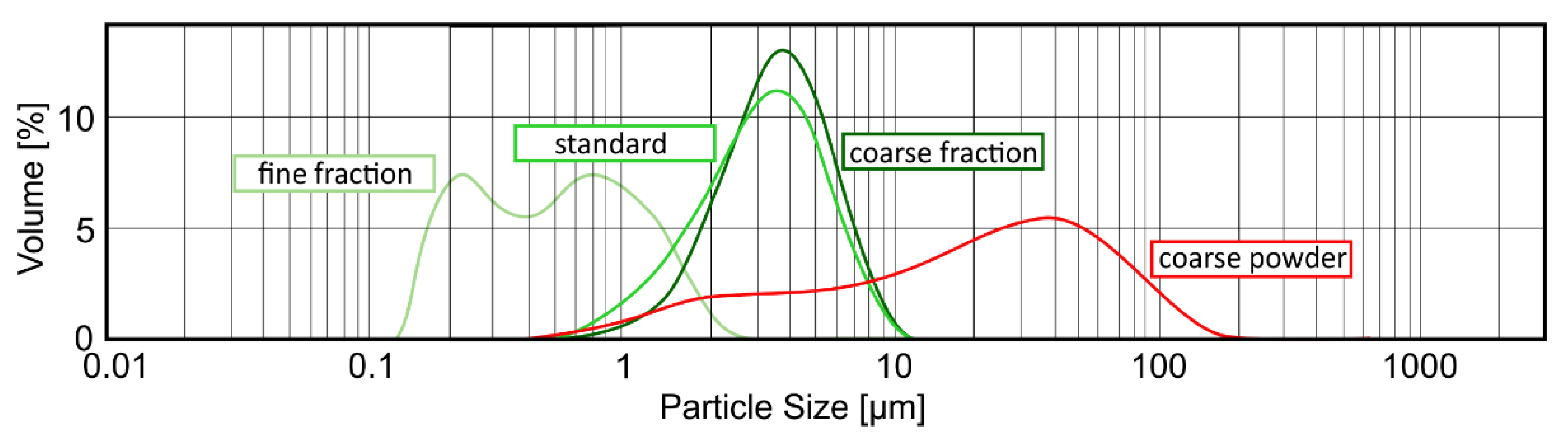
4.3. Measurement of Particle Size Distribution
The determination of the sorption capacity is a crucial step in analyzing the characteristics of purified clinoptilolite-tuff. Generally, the smaller the particle is, the larger the surface, and hence the sorption capacity, are (valid for molecules bigger in size than the pores of the PCT). To study this phenomenon in detail, three different grain-size fractions of G-PUR
® were examined. First, G-PUR
® was tested as a whole (standard), containing all particle sizes (d50 = 3.18 µm); furthermore, fine (d50 = 0.52 µm) and coarse (d50 = 3.57 µm) fractions of G-PUR
®, which were separated by centrifugation from the initial product, were analyzed. The coarse powder (d50 = 20.90 µm) corresponding to the purified material of which G-PUR
® was produced, but this was carried out before micronization. The distribution of the examined single PCT samples is listed in
Table 3 and illustrated in
Figure 6.
The measurement of the particle size distribution using the Malvern instruments device Mastersizer 2000 was performed as follows. The sample was added till the light attenuation in the analyzer reached 10–20%. After 10 s of ultra-sonication and a 1 min incubation time, the sample was measured in triplicate and averaged. Three consecutive samples of each particle fraction were analyzed (i.e., nine measurements per PCT fraction). Data processing and evaluation was carried out via the Mastersizer 2000 software, version 5.31.
The procedure followed a quality-controlled, standardized, and validated method for routine analysis of PCT particle fractions.
4.4. The Kinetics of Gliadin Adsorption to PCT
To analyze the binding of gliadin to PCT in a time-dependent manner, an ELISA assay (RIDASCREEN® Gliadin ELISA kit, R7001, R-Biopharm AG, Darmstadt, Germany) was used. Gliadin (gliadin from wheat, G3375, Sigma-Aldrich) was diluted in phosphate buffer pH 6.8 containing 250 mL of 0.2 M KH2PO4 (potassium dihydrogen phosphate, 26,931.263, VWR) and 112 mL of 0.2 M NaOH (sodium hydroxide solution, ≥32%, extra pure, Carl Roth) in a final volume of 1000 mL to a final concentration of 125 µg/mL. Purified clinoptilolite-tuff was diluted in the same buffer to gain an end-concentration of 4 mg/mL. Controls containing only gliadin in suspension were performed in duplets, while gliadin and PCT mixtures were set up in quadruplets. All samples were incubated at 37 °C under rotation (25 rpm/min, rotator SB3, Stuart) between 1 h as maximum and 1 min as minimum. The samples were centrifuged (MEGA STAR 1.6R, VWR) at 4100× g for 2 min at room temperature prior to determining the content of gliadin in the supernatants by ELISA, following the manufacturer’s instructions. Measurements were performed at a wavelength of λ = 450 nm by the use of a plate reader (Biotek synergy HT, BioTek Instruments, Inc., Winooski, VT, USA).
4.5. Adsorption of Gliadin to Different PCT Fractions
The prolamin solution containing gliadin from wheat (G3375, Sigma) was extracted according to the ELISA’s manufacturer’s advice, using the patented cocktail developed by Dr. Enrique Mendez (R7006, R-Biopharm AG, Darmstadt, Germany), which is the official method according to the AOAC. Then, the extract was diluted to a final concentration of 150 µg/mL in phosphate buffer pH 6.8 containing 250 mL of 0.2 M KH2PO4 and 112 mL of 0.2 M NaOH in a final volume of 1000 mL.
The different fractions (coarse, standard = G-PUR
®, and fine) and a coarse powder of PCT were used in various amounts, as noted in
Table 3. They were also diluted in phosphate buffer pH 6.8.
Prior to mixing, the various solutions were pre-heated to 37 °C to simulate human body temperature.
After 30 min of incubation rotating (25 rpm/min, rotator SB3, Stuart) at 37 °C, the samples were centrifuged at 4100× g (MEGA STAR 1.6R, VWR) for 2 min and the supernatants were analyzed by ELISA (R7001, R-Biopharm AG, Darmstadt, Germany), as recommended by the manufacturer, to examine their adsorption value. Measurements were performed at a wavelength of λ = 450 nm by using a plate reader (Biotek synergy HT).
4.6. The Influence of pH
The determination of a possible influence of acidity or alkalinity on gliadin adsorption to purified clinoptilolite-tuff was performed by incubation of 125 µg/mL gliadin and 0.1 g/mL PCT for 30 min at 37 °C under rotation (25 rpm/min, rotator SB3, Stuart) in different buffers, including pH 7.8, 6.8, 5.8, 4.5, and a solution of pH 1.5 (
Table 2). Then, the samples were centrifuged at 4100×
g (MEGA STAR 1.6R, VWR) for 2 min and the supernatants were analyzed by ELISA (R7001, R-Biopharm AG, Darmstadt, Germany), according to the manufacturer’s guidelines, to examine their adsorption value. Measurements were performed at a wavelength of λ = 450 nm by using a plate reader (Biotek synergy HT).
4.7. Determination of Gluten Content via ELISA
Gluten were detected by using the RIDASCREEN® Gliadin ELISA kit (R7001, R-Biopharm AG, Darmstadt, Germany), which is especially designed for the quantitative analysis of wheat, rye, and barley prolamins in food declared as gluten-free. It is certified as the Codex Alimentarius Method (Type I) and by the AOAC. It has a limit of detection according to 0.5 mg/kg gliadin (≙1.0 mg/kg gluten) and a limit of quantification referring to 2.5 mg/kg gliadin (≙5.0 mg/kg gluten). Prolamins react with the monoclonal R5 antibody offered in the kit.
The gliadin content of the supernatants was determined using the RIDASCREEN® Gliadin ELISA kit (R7001, R-Biopharm AG, Darmstadt, Germany) before measurement of the samples in a spectrophotometer (Biotek synergy HT) at a wavelength of λ = 450 nm.
It is important to note that the content of gluten refers to twice of the amount determined for gliadin/hordein/secalin. This is based on the assumption that the ratio of prolamin:glutelin is 1:1.
4.8. The Isolation of Secalins and Hordeins of Flour
Flour of barley and rye was used for the isolation of hordeins and secalins, respectively.
Organic barley and organic rye were purchased from different providers, either as grain or as flour. Organic barley (Rosenfellner Mühle, St. Peter in der Au, Austria, MHD 080720219 and Spar Österreich Warenhandels-AG, Salzburg, Austria, L22013S0421, MHD 31.01.2023) and organic rye (Rosenfellner Mühle, MHD 02042021) grain were rough-ground in a kitchen appliance (Russel Hobbs, Multifunctional Blender) before milling in a vibrational ball mill (MM 400, Retsch, Hahn, Germany) at a frequency of 30 Hz for 1 min. The resulting powder of the milled barley and rye was sieved (250 µm sieve) in a vibratory sieve shaker (AS 200 control, Retsch) and the fine fraction (<250 µm) was used for the extraction. The single powders or the purchased rye flour (Spar Österreich Warenhandels-AG, Typ 960, L215087, MHD 17.10.2022) were then incubated with the patented cocktail (R7006, R-Biopharm AG, Darmstadt, Germany) according to the manufacturer’s guidelines to isolate the specific prolamins. In short, 500 mg of the barley or rye samples were extracted by the addition of 5 mL of the patented cocktail at 50 °C in a water bath (WB 10, Medingen, Germany) for 40 min. Afterwards, a cooldown of the samples at room temperature lasted for 3 min. Then, 15 mL of 80% EtOH were added per sample and mixed thoroughly (vortex mixer SA8, Stuart). Another incubation rotating (25 rpm/min, rotator SB3, Stuart) at room temperature lasted for 1 h. Finally, the samples were centrifuged two times, first at 3000× g (MEGA STAR 1.6R, VWR) for 10 min and subsequently at 21,885× g (Biofuge primo R, Heraeus, Hanau, Germany) for another 10 min. The resulting supernatants containing the extracts were transferred to a new vial and stored at room temperature in the dark.
The concentration of prolamins in the resulting extracts was determined by RIDASCREEN® Gliadin ELISA (R7001, R-Biopharm AG, Darmstadt, Germany).
Binding experiments of secalins and hordeins to purified clinoptilolite-tuff were performed, as described for gliadins.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}