
Global Noncoding microRNA Profiling in Mice Infected with Partial Human Mouth Microbes (PAHMM) Using an Ecological Time-Sequential Polybacterial Periodontal Infection (ETSPPI) Model Reveals Sex-Specific Differential microRNA Expression

 ,
, 
 , ,
, ,
Abstract
1. Introduction
2. Results
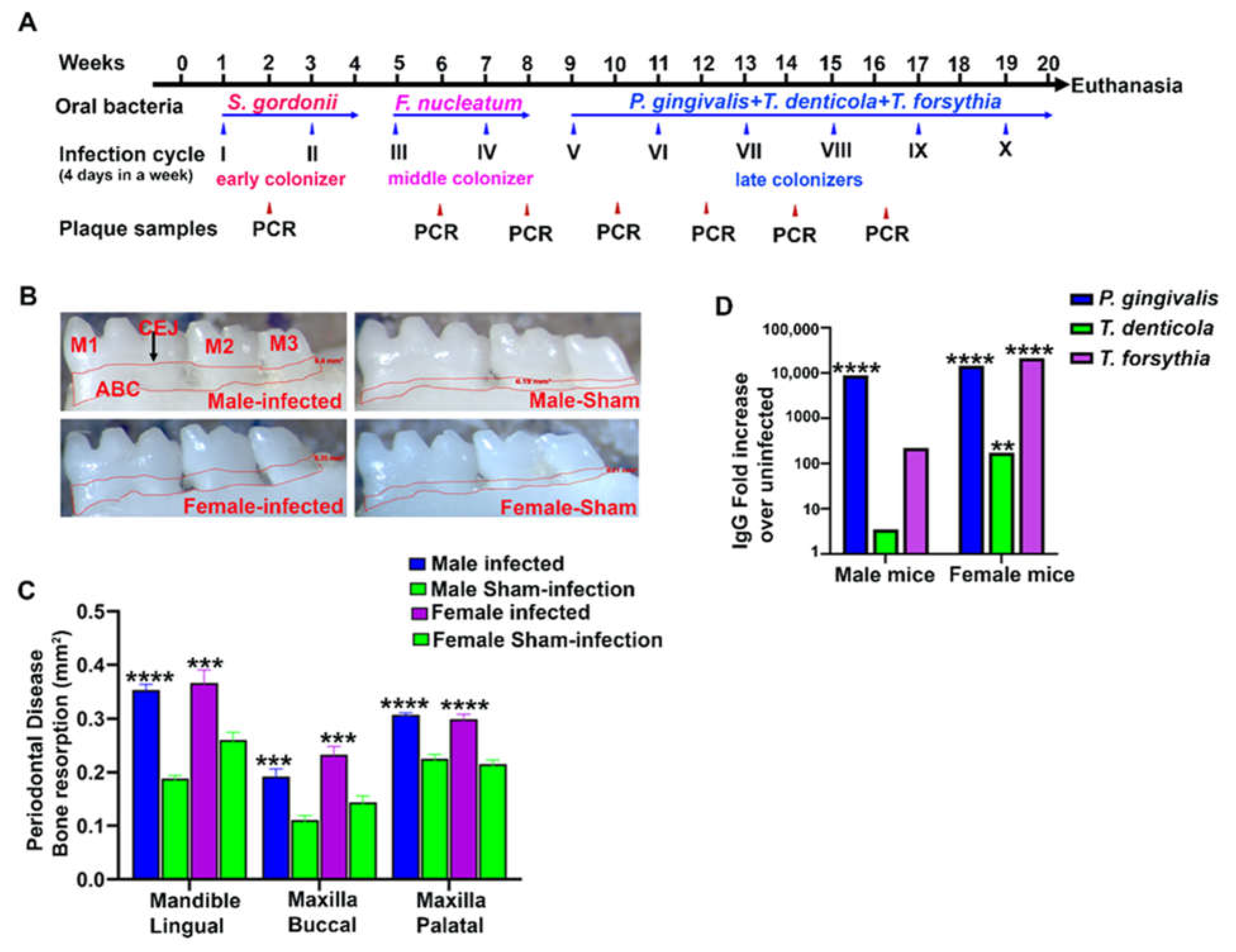
2.1. ETSPPI-Induced PAHMM Colonization in C57BL/6J Mice
2.2. ETSPPI Increased Alveolar Bone Resorption, IgG Antibody Response, and Dissemination of Bacteria to Distal Organs
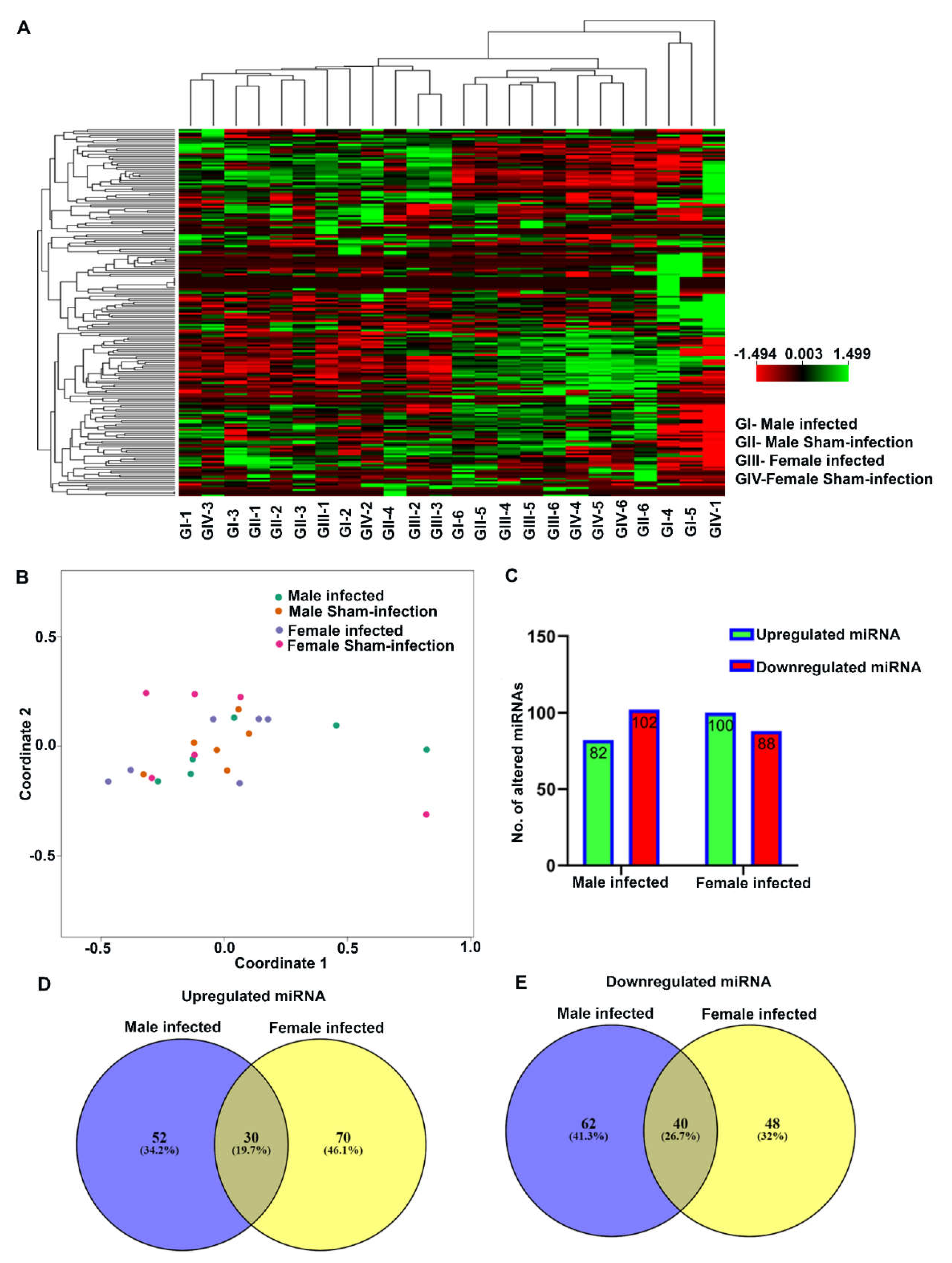
2.3. Nanostring Analysis of miRNAs in Bacteria-Infected Mandibles
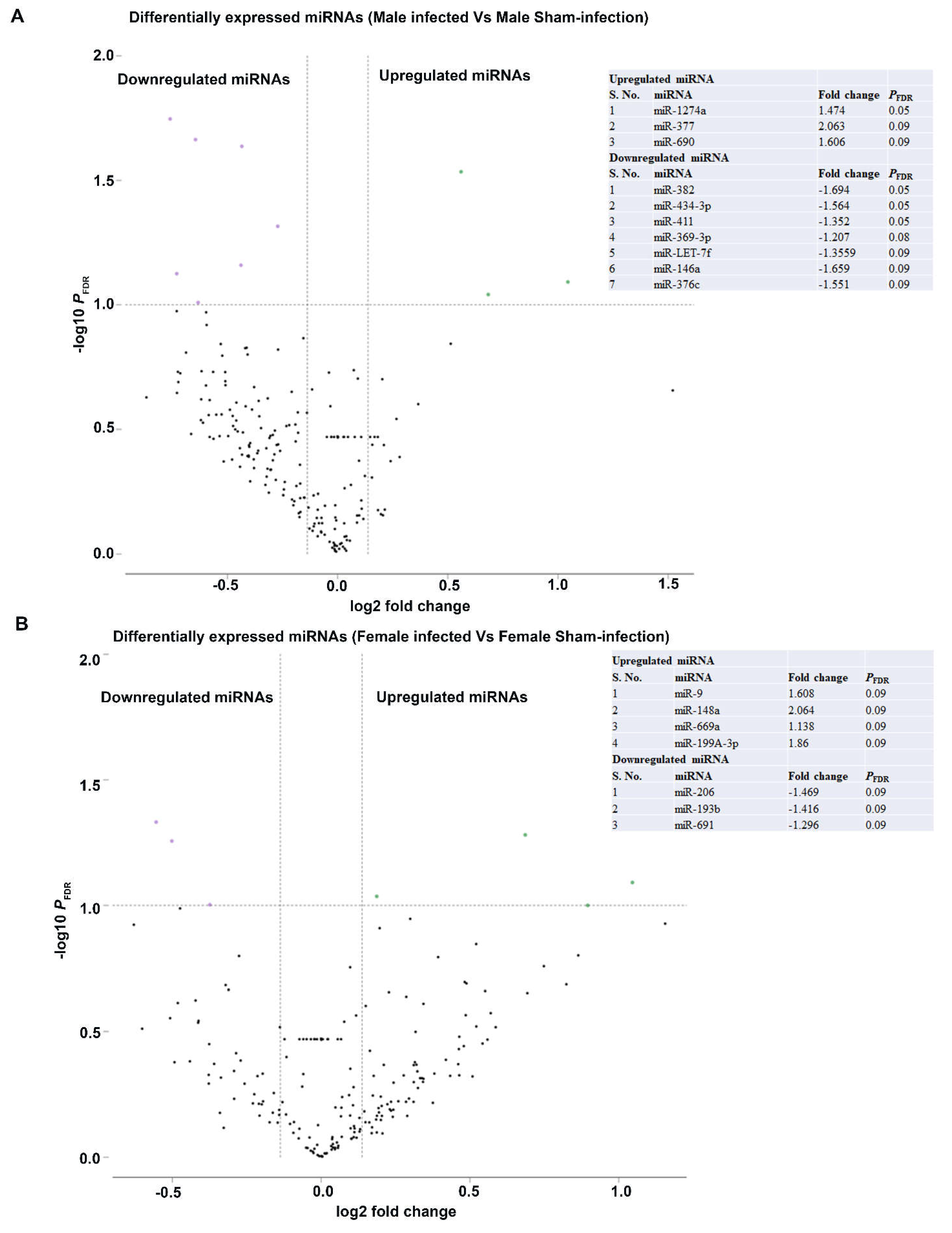
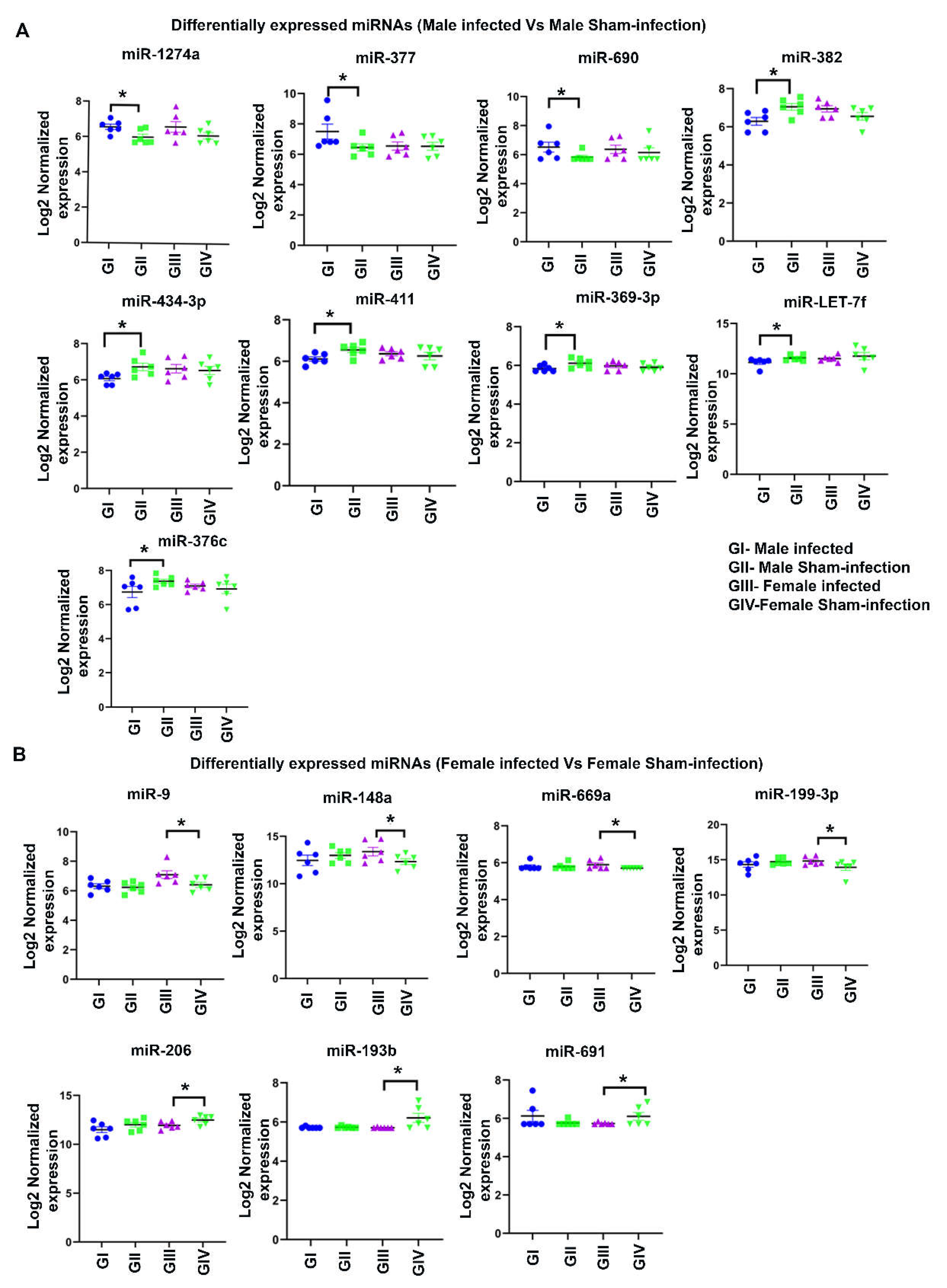
2.4. Differentially Expressed miRNAs
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Growth Conditions, and Polybacterial Preparations
4.2. Ecological Time-Sequential Polybacterial Periodontal Infection in C57BL/6J Mice
4.3. Bacterial Genomic DNA Detection in Gingival Plaque Using PCR
4.4. Bacterial Systemic Dissemination to Distal Organs
4.5. Antibacterial Serum Immunoglobulin G Antibody Analysis by Enzyme-Linked Immunosorbent Assay
4.6. Alveolar Bone Resorption by Morphometry
4.7. Extraction and Quantification of RNA for Nanostring Analysis
4.8. nCounter® miRNA Expression Profiling
4.9. Nanostring Data Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xie, Y.F.; Shu, R.; Jiang, S.Y.; Liu, D.L.; Zhang, X.L. Comparison of microRNA profiles of human periodontal diseased and healthy gingival tissues. Int. J. Oral. Sci. 2011, 3, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Nahid, M.A.; Rivera, M.; Lucas, A.; Chan, E.K.; Kesavalu, L. Polymicrobial infection with periodontal pathogens specifically enhances microRNA miR-146a in ApoE−/− mice during experimental periodontal disease. Infect. Immun. 2011, 79, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Motedayyen, H.; Ghotloo, S.; Saffari, M.; Sattari, M.; Amid, R. Evaluation of MicroRNA-146a and Its Targets in Gingival Tissues of Patients with Chronic Periodontitis. J. Periodontol. 2015, 86, 1380–1385. [Google Scholar] [CrossRef]
- Jiang, S.Y.; Xue, D.; Xie, Y.F.; Zhu, D.W.; Dong, Y.Y.; Wei, C.C.; Deng, J.Y. The negative feedback regulation of microRNA-146a in human periodontal ligament cells after Porphyromonas gingivalis lipopolysaccharide stimulation. Inflamm. Res. 2015, 64, 441–451. [Google Scholar] [CrossRef]
- Perri, R.; Nares, S.; Zhang, S.; Barros, S.P.; Offenbacher, S. MicroRNA modulation in obesity and periodontitis. J. Dent. Res. 2012, 91, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Stoecklin-Wasmer, C.; Guarnieri, P.; Celenti, R.; Demmer, R.T.; Kebschull, M.; Papapanou, P.N. MicroRNAs and their target genes in gingival tissues. J. Dent. Res. 2012, 91, 934–940. [Google Scholar] [CrossRef]
- Yue, J.; Song, D.; Lu, W.; Lu, Y.; Zhou, W.; Tan, X.; Zhang, L.; Huang, D. Expression Profiles of Inflammation-associated microRNAs in Periapical Lesions and Human Periodontal Ligament Fibroblasts Inflammation. J. Endod. 2016, 42, 1773–1778. [Google Scholar] [CrossRef]
- Chen, S.C.; Constantinides, C.; Kebschull, M.; Papapanou, P.N. MicroRNAs Regulate Cytokine Responses in Gingival Epithelial Cells. Infect. Immun. 2016, 84, 3282–3289. [Google Scholar] [CrossRef]
- Eke, P.I.; Dye, B.A.; Wei, L.; Slade, G.D.; Thornton-Evans, G.O.; Borgnakke, W.S.; Taylor, G.W.; Page, R.C.; Beck, J.D.; Genco, R.J. Update on Prevalence of Periodontitis in Adults in the United States: NHANES 2009 to 2012. J. Periodontol. 2015, 86, 611–622. [Google Scholar] [CrossRef]
- Shiau, H.J.; Aichelmann-Reidy, M.E.; Reynolds, M.A. Influence of sex steroids on inflammation and bone metabolism. Periodontology 2000 2014, 64, 81–94. [Google Scholar] [CrossRef]
- McAbee, J.; Li, Q.; Yu, H.; Kirkwood, K.L. Sexual dimorphism in periapical inflammation and bone loss from mitogen-activated protein kinase phosphatase-1 deficient mice. J. Endod. 2012, 38, 1097–1100. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Niessen, L.C.; Gibson, G.; Kinnunen, T.H. Women’s oral health: Why sex and gender matter. Dent. Clin. N. Am. 2013, 57, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Glowacki, J.; Christoph, K. Gender differences in the growing, abnormal, and aging jaw. Dent. Clin. N. Am. 2013, 57, 263–280. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, B.; Duffy, D.; Urrutia, A.; Quach, H.; Patin, E.; Posseme, C.; Bergstedt, J.; Charbit, B.; Rouilly, V.; MacPherson, C.R.; et al. Distinctive roles of age, sex, and genetics in shaping transcriptional variation of human immune responses to microbial challenges. Proc. Natl. Acad. Sci. USA 2018, 115, E488–E497. [Google Scholar] [CrossRef]
- Zhao, Y.Q.; Zhou, Y.H.; Zhao, J.; Feng, Y.; Gao, Z.R.; Ye, Q.; Liu, Q.; Chen, Y.; Zhang, S.H.; Tan, L.; et al. Sex Variations in the Oral Microbiomes of Youths with Severe Periodontitis. J. Immunol. Res. 2021, 2021, 8124593. [Google Scholar] [CrossRef]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef]
- Mutha, N.V.R.; Mohammed, W.K.; Krasnogor, N.; Tan, G.Y.A.; Choo, S.W.; Jakubovics, N.S. Transcriptional responses of Streptococcus gordonii and Fusobacterium nucleatum to coaggregation. Mol. Oral. Microbiol. 2018, 33, 450–464. [Google Scholar] [CrossRef]
- Park, Y.; Simionato, M.R.; Sekiya, K.; Murakami, Y.; James, D.; Chen, W.; Hackett, M.; Yoshimura, F.; Demuth, D.R.; Lamont, R.J. Short fimbriae of Porphyromonas gingivalis and their role in coadhesion with Streptococcus gordonii. Infect. Immun. 2005, 73, 3983–3989. [Google Scholar] [CrossRef]
- Brown, J.L.; Yates, E.A.; Bielecki, M.; Olczak, T.; Smalley, J.W. Potential role for Streptococcus gordonii-derived hydrogen peroxide in heme acquisition by Porphyromonas gingivalis. Mol. Oral. Microbiol. 2018, 33, 322–335. [Google Scholar] [CrossRef]
- Chukkapalli, S.S.; Easwaran, M.; Rivera-Kweh, M.F.; Velsko, I.M.; Ambadapadi, S.; Dai, J.; Larjava, H.; Lucas, A.R.; Kesavalu, L. Sequential colonization of periodontal pathogens in induction of periodontal disease and atherosclerosis in LDLRnull mice. Pathog. Dis. 2017, 75, ftx003. [Google Scholar] [CrossRef]
- Chukkapalli, S.S.; Velsko, I.M.; Rivera-Kweh, M.F.; Zheng, D.; Lucas, A.R.; Kesavalu, L. Polymicrobial Oral Infection with Four Periodontal Bacteria Orchestrates a Distinct Inflammatory Response and Atherosclerosis in ApoE null Mice. PLoS ONE 2015, 10, e0143291. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Na, H.S.; Jeong, S.Y.; Jeong, S.H.; Park, H.R.; Chung, J. Comparison of inflammatory microRNA expression in healthy and periodontitis tissues. Biocell 2011, 35, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, S.S.; Rivera, M.F.; Velsko, I.M.; Lee, J.Y.; Chen, H.; Zheng, D.; Bhattacharyya, I.; Gangula, P.R.; Lucas, A.R.; Kesavalu, L. Invasion of oral and aortic tissues by oral spirochete Treponema denticola in ApoE(−/−) mice causally links periodontal disease and atherosclerosis. Infect. Immun. 2014, 82, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, S.S.; Ambadapadi, S.; Varkoly, K.; Jiron, J.; Aguirre, J.I.; Bhattacharyya, I.; Morel, L.M.; Lucas, A.R.; Kesavalu, L. Impaired innate immune signaling due to combined Toll-like receptor 2 and 4 deficiency affects both periodontitis and atherosclerosis in response to polybacterial infection. Pathog. Dis. 2018, 76, fty076. [Google Scholar] [CrossRef]
- Rivera, M.F.; Lee, J.Y.; Aneja, M.; Goswami, V.; Liu, L.; Velsko, I.M.; Chukkapalli, S.S.; Bhattacharyya, I.; Chen, H.; Lucas, A.R.; et al. Polymicrobial infection with major periodontal pathogens induced periodontal disease and aortic atherosclerosis in hyperlipidemic ApoE(null) mice. PLoS ONE 2013, 8, e57178. [Google Scholar] [CrossRef]
- Velsko, I.M.; Chukkapalli, S.S.; Rivera-Kweh, M.F.; Zheng, D.; Aukhil, I.; Lucas, A.R.; Larjava, H.; Kesavalu, L. Periodontal pathogens invade gingiva and aortic adventitia and elicit inflammasome activation in alphavbeta6 integrin-deficient mice. Infect. Immun. 2015, 83, 4582–4593. [Google Scholar] [CrossRef]
- Rodean, I.P.; Lazar, L.; Halatiu, V.B.; Biris, C.; Benedek, I.; Benedek, T. Periodontal Disease Is Associated with Increased Vulnerability of Coronary Atheromatous Plaques in Patients Undergoing Coronary Computed Tomography Angiography-Results from the Atherodent Study. J. Clin. Med. 2021, 10, 1290. [Google Scholar] [CrossRef]
- Singer, R.H.; Stoutenberg, M.; Feaster, D.J.; Cai, J.; Hlaing, W.M.; Metsch, L.R.; Salazar, C.R.; Beaver, S.M.; Finlayson, T.L.; Talavera, G.; et al. The association of periodontal disease and cardiovascular disease risk: Results from the Hispanic Community Health Study/Study of Latinos. J. Periodontol. 2018, 89, 840–857. [Google Scholar] [CrossRef]
- Lockhart, P.B.; Bolger, A.F.; Papapanou, P.N.; Osinbowale, O.; Trevisan, M.; Levison, M.E.; Taubert, K.A.; Newburger, J.W.; Gornik, H.L.; Gewitz, M.H.; et al. Periodontal disease and atherosclerotic vascular disease: Does the evidence support an independent association? A scientific statement from the American Heart Association. Circulation 2012, 125, 2520–2544. [Google Scholar] [CrossRef]
- Aravindraja, C.; Sakthivel, R.; Liu, X.; Goodwin, M.; Veena, P.; Godovikova, V.; Fenno, J.C.; Levites, Y.; Golde, T.E.; Kesavalu, L. Intracerebral but Not Peripheral Infection of Live Porphyromonas gingivalis Exacerbates Alzheimer’s Disease like Amyloid Pathology in APP-TgCRND8 Mice. Int. J. Mol. Sci. 2022, 23, 3328. [Google Scholar] [CrossRef]
- Mosailova, N.; Truong, J.; Dietrich, T.; Ashurst, J. Streptococcus gordonii: A Rare Cause of Infective Endocarditis. Case Rep. Infect. Dis. 2019, 2019, 7127848. [Google Scholar] [CrossRef] [PubMed]
- Park, O.J.; Kwon, Y.; Park, C.; So, Y.J.; Park, T.H.; Jeong, S.; Im, J.; Yun, C.H.; Han, S.H. Streptococcus gordonii: Pathogenesis and Host Response to Its Cell Wall Components. Microorganisms 2020, 8, 1852. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Hu, H.; Fang, W.; Shi, D.; Liang, C.; Sun, Y.; Gao, G.; Wang, H.; Zhang, Q.; Wang, L.; et al. Detection of pathogens from resected heart valves of patients with infective endocarditis by next-generation sequencing. Int. J. Infect. Dis. 2019, 83, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, K.; Joseph, R.; Vo, A.; Patel, T.; Chaudhry, S.; Nguyen, U.; Trevor, A.; Robinson, E.; Campbell, M.; McLennan, J.; et al. Children with severe early childhood caries: Streptococci genetic strains within carious and white spot lesions. J. Oral Microbiol. 2014, 6, 25805. [Google Scholar] [CrossRef]
- Zandi, H.; Kristoffersen, A.K.; Ørstavik, D.; Rôças, I.N.; Siqueira, J.F., Jr.; Enersen, M. Microbial Analysis of Endodontic Infections in Root-filled Teeth with Apical Periodontitis before and after Irrigation Using Pyrosequencing. J. Endod. 2018, 44, 372–378. [Google Scholar] [CrossRef]
- Dong, J.; Li, W.; Wang, Q.; Chen, J.; Zu, Y.; Zhou, X.; Guo, Q. Relationships Between Oral Microecosystem and Respiratory Diseases. Front. Mol. Biosci. 2021, 8, 718222. [Google Scholar] [CrossRef]
- Kelly, N.; Winning, L.; Irwin, C.; Lundy, F.T.; Linden, D.; McGarvey, L.; Linden, G.J.; El Karim, I.A. Periodontal status and chronic obstructive pulmonary disease (COPD) exacerbations: A systematic review. BMC Oral. Health 2021, 21, 425. [Google Scholar] [CrossRef]
- Hajishengallis, G. Immunomicrobial pathogenesis of periodontitis: Keystones, pathobionts, and host response. Trends Immunol. 2014, 35, 3–11. [Google Scholar] [CrossRef]
- Essandoh, K.; Li, Y.; Huo, J.; Fan, G.C. MiRNA-Mediated Macrophage Polarization and its Potential Role in the Regulation of Inflammatory Response. Shock 2016, 46, 122–131. [Google Scholar] [CrossRef]
- Fang, Y.; Xu, X.Y.; Shen, Y.; Li, J. miR-148 targets CiGadd45ba and CiGadd45bb to modulate the inflammatory response to bacterial infection in grass carp. Dev. Comp. Immunol. 2020, 106, 103611. [Google Scholar] [CrossRef]
- Dong, W.; Gao, W.; Cui, J.; Xu, T.; Sun, Y. microRNA-148 is involved in NF-kappaB signaling pathway regulation after LPS stimulation by targeting IL-1beta in miiuy croaker. Fish Shellfish Immunol. 2021, 118, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Barbano, R.; Pasculli, B.; Rendina, M.; Fontana, A.; Fusilli, C.; Copetti, M.; Castellana, S.; Valori, V.M.; Morritti, M.; Graziano, P.; et al. Stepwise analysis of MIR9 loci identifies miR-9-5p to be involved in Oestrogen regulated pathways in breast cancer patients. Sci. Rep. 2017, 7, 45283. [Google Scholar] [CrossRef] [PubMed]
- Bardhan, K.; Liu, K. Epigenetics and colorectal cancer pathogenesis. Cancers 2013, 5, 676–713. [Google Scholar] [CrossRef]
- Cai, L.; Cai, X. Up-regulation of miR-9 expression predicate advanced clinicopathological features and poor prognosis in patients with hepatocellular carcinoma. Diagn. Pathol. 2014, 9, 1000. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, L.; Ma, Z.; Sun, G.; Luo, X.; Li, M.; Zhai, S.; Li, P.; Wang, X. Oncogenic miR-9 is a target of erlotinib in NSCLCs. Sci. Rep. 2015, 5, 17031. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Li, Q.; Xu, Q.; Liu, L.; Jiang, B. MiR-148a inhibits angiogenesis by targeting ERBB3. J. Biomed. Res. 2011, 25, 170–177. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, L.; Xing, W.; Sun, H. microRNA-148a-3p enhances the effects of sevoflurane on hepatocellular carcinoma cell progression via ROCK1 repression. Cell. Signal. 2021, 83, 109982. [Google Scholar] [CrossRef]
- Liu, J.; Si, L.; Tian, H. MicroRNA-148a inhibits cell proliferation and cell cycle progression in lung adenocarcinoma via directly targeting transcription factor E2F3. Exp. Ther. Med. 2018, 16, 5400–5409. [Google Scholar] [CrossRef]
- Fu, J.; Li, T.; Jiang, X.; Xia, B.; Hu, L. MicroRNA-199-3p targets Sp1 transcription factor to regulate proliferation and epithelial to mesenchymal transition of human lung cancer cells. 3 Biotech 2021, 11, 352. [Google Scholar] [CrossRef]
- Ren, B.; Yang, B.; Li, P.; Ge, L. Upregulation of MiR-1274a is Correlated with Survival Outcomes and Promotes Cell Proliferation, Migration, and Invasion of Colon Cancer. OncoTargets Ther. 2020, 13, 6957–6966. [Google Scholar] [CrossRef]
- Zhu, S.J.; Wang, X.; Hu, S.L.; Fang, Y.; Guan, B.X.; Li, J.; Li, G.; Xu, J.Y. Clinical Significance and Biological Function of miR-1274a in Non-small Cell Lung Cancer. Mol. Biotechnol. 2021, 64, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Ding, S.; Sun, Y.; Zhao, B.; Pan, Y.; Wan, J. MiR-377 Regulates Inflammation and Angiogenesis in Rats After Cerebral Ischemic Injury. J. Cell. Biochem. 2018, 119, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Garcia, R.I.; Janket, S.J.; Jones, J.A.; Mascarenhas, A.K.; Scott, T.E.; Nunn, M.E. The association between cumulative periodontal disease and stroke history in older adults. J. Periodontol. 2006, 77, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.L.; Hu, H.Y.; Huang, N.; Hwang, D.K.; Chou, P.; Chu, D. Dental prophylaxis and periodontal treatment are protective factors to ischemic stroke. Stroke 2013, 44, 1026–1030. [Google Scholar] [CrossRef]
- Kim, H.D.; Sim, S.J.; Moon, J.Y.; Hong, Y.C.; Han, D.H. Association between periodontitis and hemorrhagic stroke among Koreans: A case-control study. J. Periodontol. 2010, 81, 658–665. [Google Scholar] [CrossRef]
- Grau, A.J.; Becher, H.; Ziegler, C.M.; Lichy, C.; Buggle, F.; Kaiser, C.; Lutz, R.; Bultmann, S.; Preusch, M.; Dorfer, C.E. Periodontal disease as a risk factor for ischemic stroke. Stroke 2004, 35, 496–501. [Google Scholar] [CrossRef]
- Wu, T.; Trevisan, M.; Genco, R.J.; Dorn, J.P.; Falkner, K.L.; Sempos, C.T. Periodontal disease and risk of cerebrovascular disease: The first national health and nutrition examination survey and its follow-up study. Arch. Intern Med. 2000, 160, 2749–2755. [Google Scholar] [CrossRef]
- Jimenez, M.; Krall, E.A.; Garcia, R.I.; Vokonas, P.S.; Dietrich, T. Periodontitis and incidence of cerebrovascular disease in men. Ann. Neurol. 2009, 66, 505–512. [Google Scholar] [CrossRef]
- Guo, Y.; Huang, S.; Ma, Y.; Zhang, J.; Wen, Y.; Zhou, L.; Yuan, G.; Cheng, J. MiR-377 mediates the expression of Syk to attenuate atherosclerosis lesion development in ApoE(−/−) mice. Biomed. Pharmacother. 2019, 118, 109332. [Google Scholar] [CrossRef]
- Xin, M.; Liang, H.; Wang, H.; Wen, D.; Wang, L.; Zhao, L.; Sun, M.; Wang, J. Mirt2 functions in synergy with miR-377 to participate in inflammatory pathophysiology of Sjogren’s syndrome. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2473–2480. [Google Scholar] [CrossRef]
- Menikdiwela, K.R.; Ramalingam, L.; Abbas, M.M.; Bensmail, H.; Scoggin, S.; Kalupahana, N.S.; Palat, A.; Gunaratne, P.; Moustaid-Moussa, N. Role of microRNA 690 in Mediating Angiotensin II Effects on Inflammation and Endoplasmic Reticulum Stress. Cells 2020, 34, 1327. [Google Scholar] [CrossRef]
- Al-Atrache, Z.; Lopez, D.B.; Hingley, S.T.; Appelt, D.M. Astrocytes infected with Chlamydia pneumoniae demonstrate altered expression and activity of secretases involved in the generation of beta-amyloid found in Alzheimer disease. BMC Neurosci. 2019, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Kesavalu, L.; Sathishkumar, S.; Bakthavatchalu, V.; Matthews, C.; Dawson, D.; Steffen, M.; Ebersole, J.L. Rat model of polymicrobial infection, immunity, and alveolar bone resorption in periodontal disease. Infect. Immun. 2007, 75, 1704–1712. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, S.S.; Velsko, I.M.; Rivera-Kweh, M.F.; Larjava, H.; Lucas, A.R.; Kesavalu, L. Global TLR2 and 4 deficiency in mice impacts bone resorption, inflammatory markers and atherosclerosis to polymicrobial infection. Mol. Oral. Microbiol. 2017, 32, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Vukmirovic, M.; Herazo-Maya, J.D.; Blackmon, J.; Skodric-Trifunovic, V.; Jovanovic, D.; Pavlovic, S.; Stojsic, J.; Zeljkovic, V.; Yan, X.; Homer, R.; et al. Identification and validation of differentially expressed transcripts by RNA-sequencing of formalin-fixed, paraffin-embedded (FFPE) lung tissue from patients with Idiopathic Pulmonary Fibrosis. BMC Pulm. Med. 2017, 17, 15. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Bacteria and Oral Infection | Sex | Number of Mice |
|---|---|---|---|
| I | C57BL/6J mice—ETSPPI (Sg + Fn + Pg/Td/Tf) | Male | 10 |
| II | C57BL/6J mice—sham-infection (Vehicle control) | Male | 10 |
| III | C57BL/6J mice—ETSPPI (Sg + Fn + Pg/Td/Tf) | Female | 10 |
| IV | C57BL/6J mice—sham-infection (vehicle control) | Female | 9 |
| Group/Mice/Infection | Positive Gingival Plaque Samples (n = 10 Mice) | ||||
|---|---|---|---|---|---|
| 2 weeks (Sg) | 6 weeks (Fn) | 8 weeks (Fn) | 10 weeks Pg/Td/Tf | 14 weeks Pg/Td/Tf | |
| Group I/Male C57BL/6J mice/ETSPPI | 10 | 5 | 10 | 10/5/5 | 10/6/10 |
| Group II/Male C57BL/6J mice/Sham-infection | 0 | 0 | NC | 0/0/0 | NC |
| Group III/Female C57BL/6J mice/ETSPPI | 10 | 7 | 10 | 8/7/6 | 10/9/10 |
| Group IV/Female C57BL/6J mice/Sham-infection | 0 | 0 | NC | 0/0/0 | NC |
| Positive Systemic Tissue Specimens (n = 10) | |||||
|---|---|---|---|---|---|
| Bacterial Infection | Sex (Male/Female) | Heart | Lungs | Kidney | Liver |
| S. gordonii | M | 9 | 0 | 6 | 9 |
| F | 4 | 7 * | 4 | 9 | |
| F. nucleatum | M | 9* | 0 | 0 | 0 |
| F | 3 | 9 * | 0 | 0 | |
| P. gingivalis | M | 4 | 1 | 0 | 0 |
| F | 2 | 8 * | 0 | 0 | |
| T. denticola | M | 10 * | 0 | 0 | 0 |
| F | 0 | 1 | 0 | 0 | |
| T. forsythia | M | 9 | 0 | 0 | 0 |
| F | 5 | 9 * | 0 | 0 | |
| This Study | Human PD | Other Diseases |
|---|---|---|
| Female Mice | ||
| miR-9 | Inflamed gingival tissues [1,22] | Gene regulators in many human cancers (e.g., breast, colorectal, hepatocellular, non-small-cell lung) [42,43,44,45] |
| miR-148a | Inflamed gingival tissues [6] | Gene regulators in many human cancers (e.g., breast, hepatocellular, lung) [46,47,48] |
| miR-669a | Not reported | Not reported |
| miR-199a-3p | Not reported | Tumor-suppressor miRNA in lung cancer [49] |
| Male mice | ||
| miR-1274a | Not reported | Oncogene and prognostic marker for colon cancer [35] Non-small-cell lung cancer [51] |
| miR-377 | Not reported | Cerebral inflammation and angiogenesis in ischemic stroke animal models [52] Atherosclerosis [62] and Sjogren’s syndrome [60] |
| miR-690 | Not reported | Inflammation and endoplasmic reticulum stress in obese animal models [61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aravindraja, C.; Kashef, M.R.; Vekariya, K.M.; Ghanta, R.K.; Karanth, S.; Chan, E.K.L.; Kesavalu, L. Global Noncoding microRNA Profiling in Mice Infected with Partial Human Mouth Microbes (PAHMM) Using an Ecological Time-Sequential Polybacterial Periodontal Infection (ETSPPI) Model Reveals Sex-Specific Differential microRNA Expression. Int. J. Mol. Sci. 2022, 23, 5107. https://doi.org/10.3390/ijms23095107
Aravindraja C, Kashef MR, Vekariya KM, Ghanta RK, Karanth S, Chan EKL, Kesavalu L. Global Noncoding microRNA Profiling in Mice Infected with Partial Human Mouth Microbes (PAHMM) Using an Ecological Time-Sequential Polybacterial Periodontal Infection (ETSPPI) Model Reveals Sex-Specific Differential microRNA Expression. International Journal of Molecular Sciences. 2022; 23(9):5107. https://doi.org/10.3390/ijms23095107
Chicago/Turabian StyleAravindraja, Chairmandurai, Matteen R. Kashef, Krishna Mukesh Vekariya, Ravi K. Ghanta, Shama Karanth, Edward K. L. Chan, and Lakshmyya Kesavalu. 2022. "Global Noncoding microRNA Profiling in Mice Infected with Partial Human Mouth Microbes (PAHMM) Using an Ecological Time-Sequential Polybacterial Periodontal Infection (ETSPPI) Model Reveals Sex-Specific Differential microRNA Expression" International Journal of Molecular Sciences 23, no. 9: 5107. https://doi.org/10.3390/ijms23095107
APA StyleAravindraja, C., Kashef, M. R., Vekariya, K. M., Ghanta, R. K., Karanth, S., Chan, E. K. L., & Kesavalu, L. (2022). Global Noncoding microRNA Profiling in Mice Infected with Partial Human Mouth Microbes (PAHMM) Using an Ecological Time-Sequential Polybacterial Periodontal Infection (ETSPPI) Model Reveals Sex-Specific Differential microRNA Expression. International Journal of Molecular Sciences, 23(9), 5107. https://doi.org/10.3390/ijms23095107