
The SCO2102 Protein Harbouring a DnaA II Protein-Interaction Domain Is Essential for the SCO2103 Methylenetetrahydrofolate Reductase Positioning at Streptomyces Sporulating Hyphae, Enhancing DNA Replication during Sporulation

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
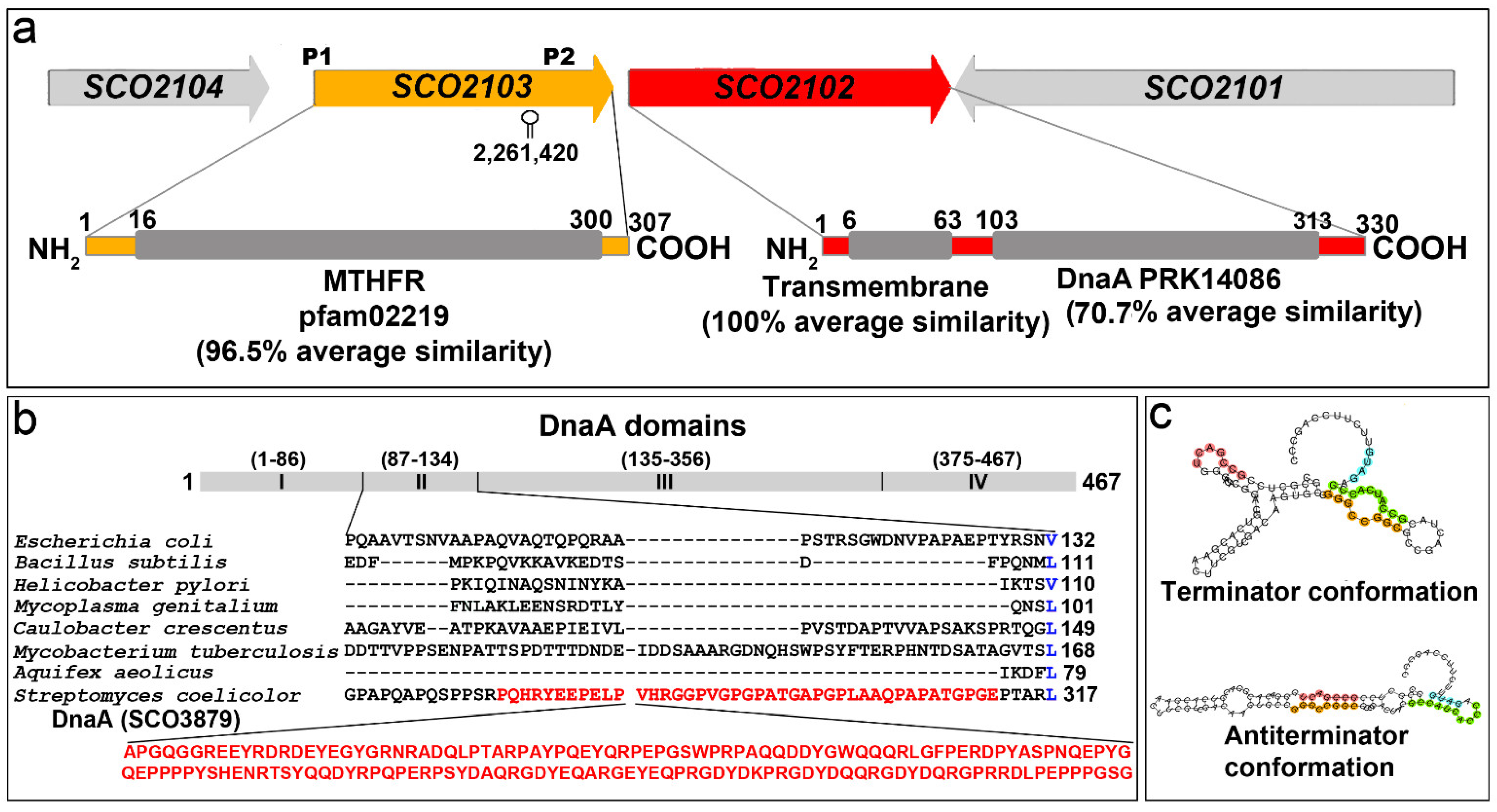
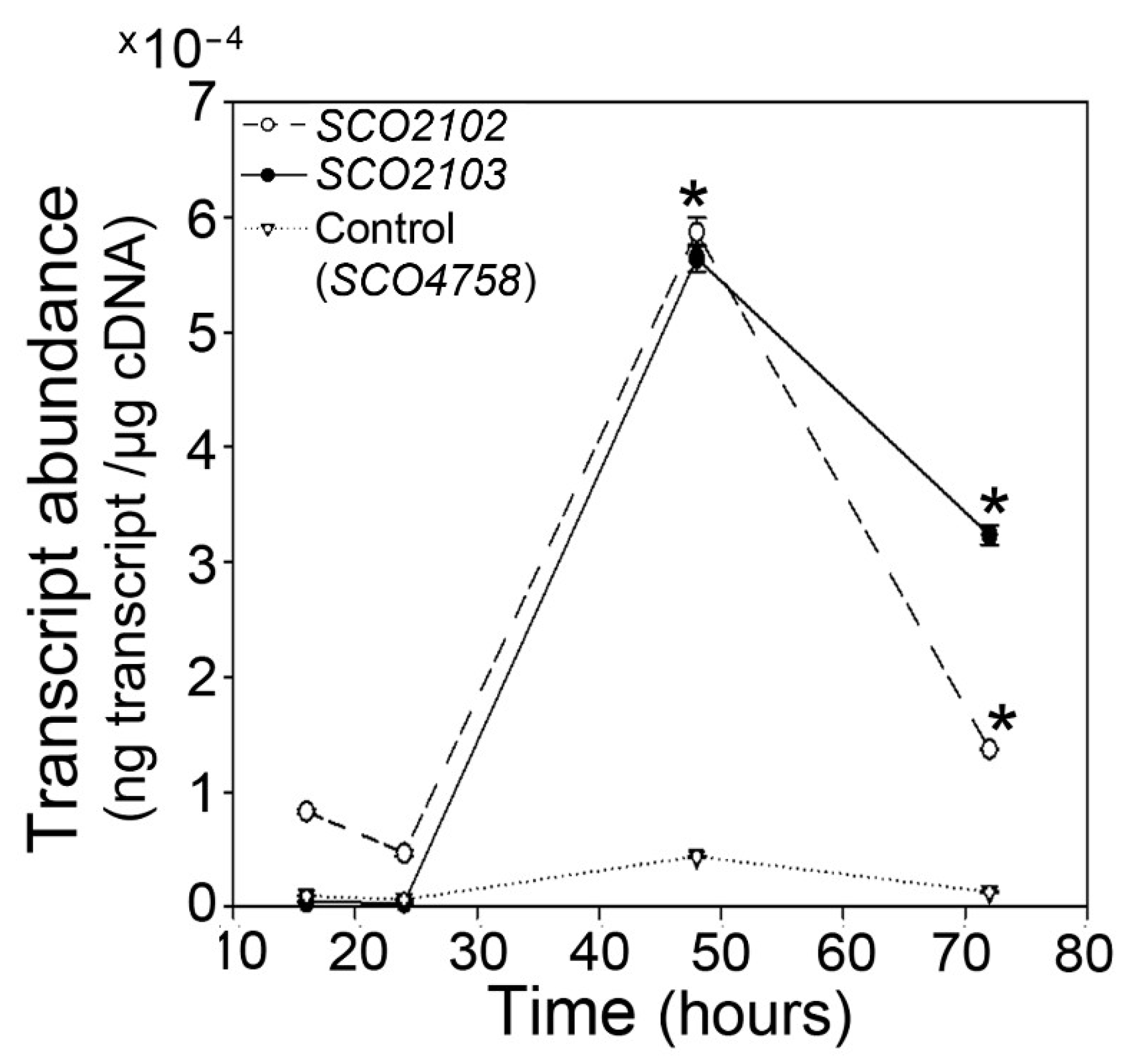
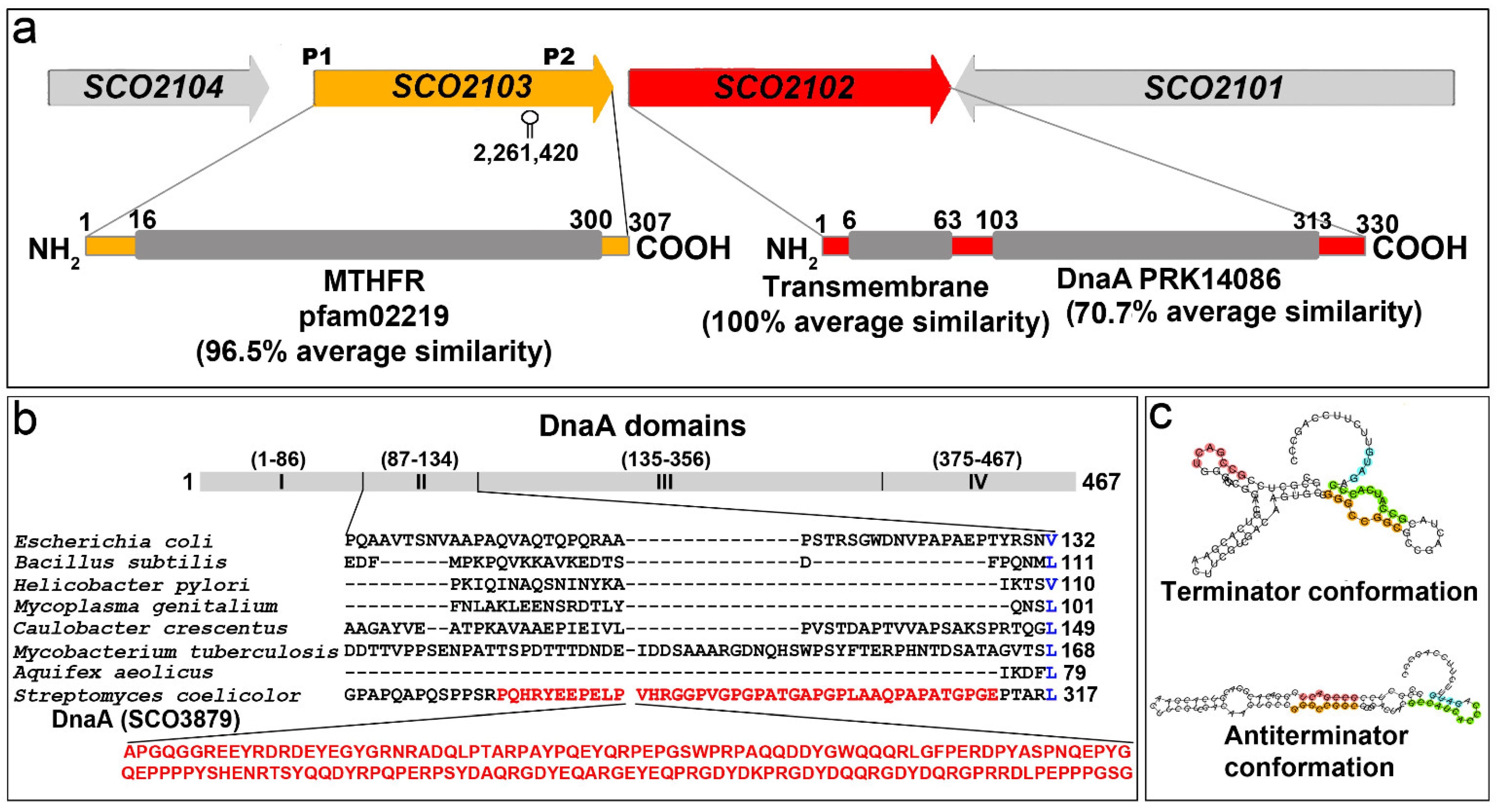
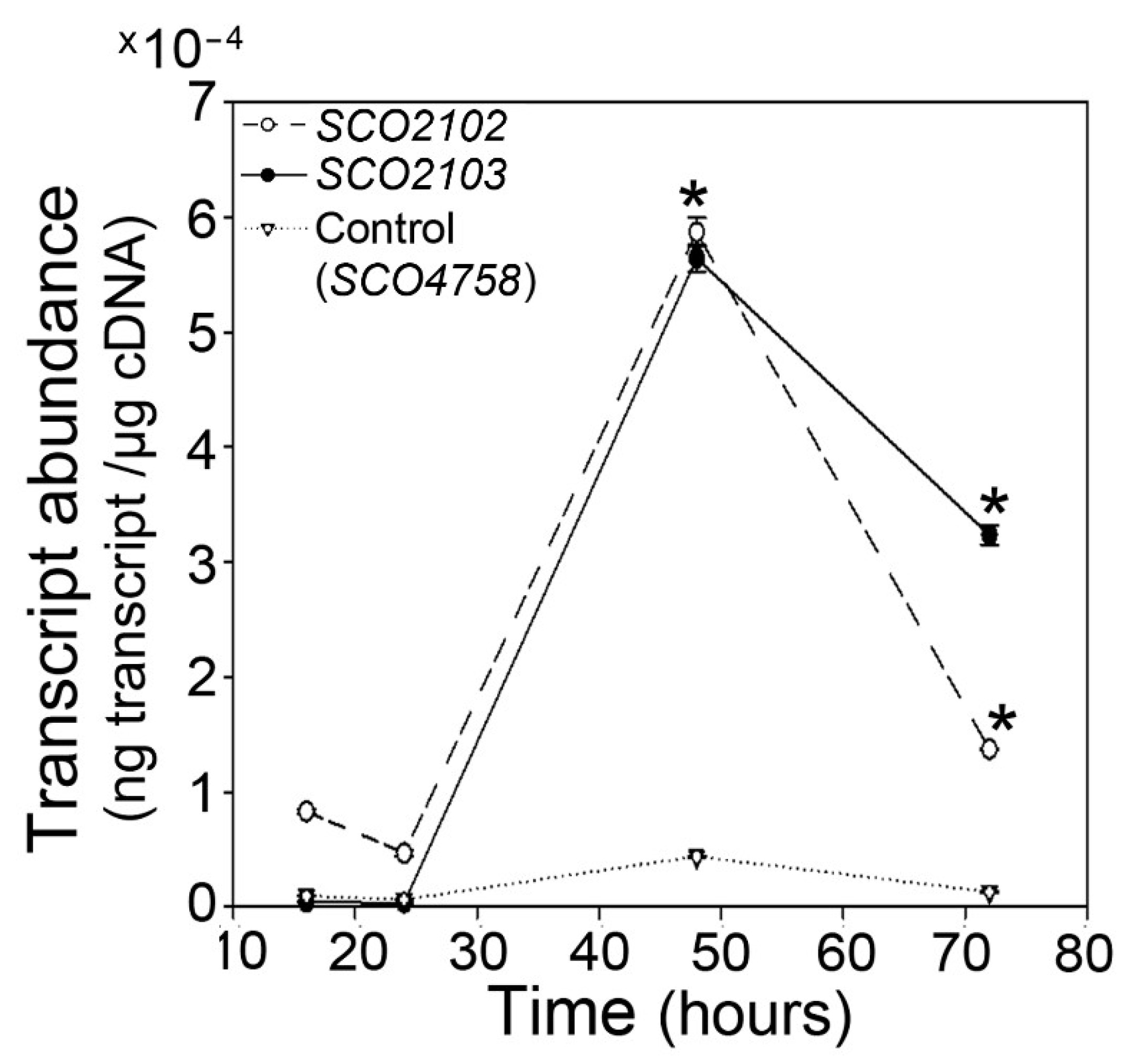
2.1. SCO2102/2103 Genes Are Highly Conserved in Streptomyces and Expressed during Sporulation
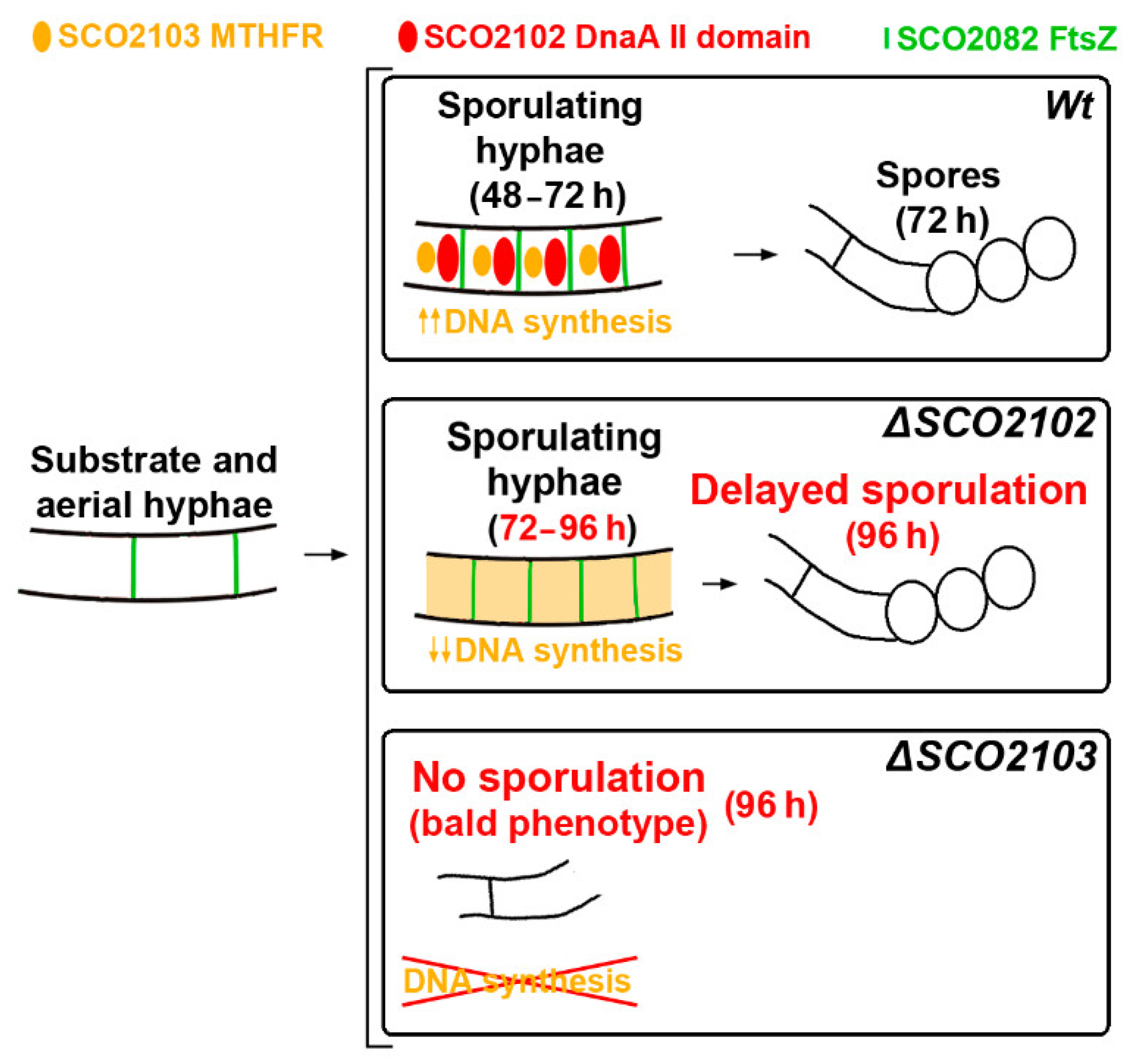
2.2. ΔSCO2103 Knockout Shows a Bald Phenotype
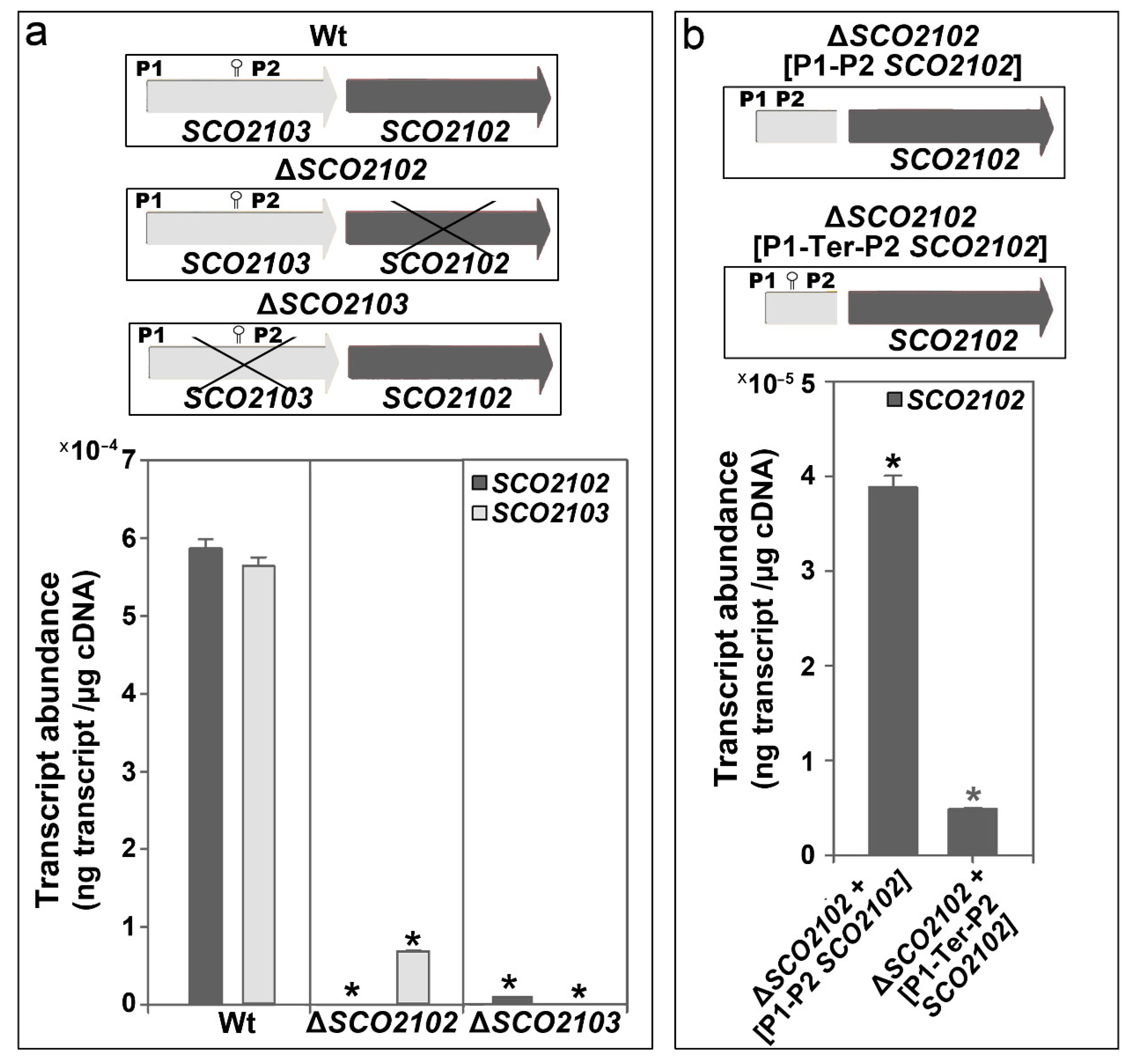
2.3. SCO2102 and SCO2103 Gene Expression Is Highly Regulated, Involving at Least Two Promoters and a Conditional Transcriptional Terminator
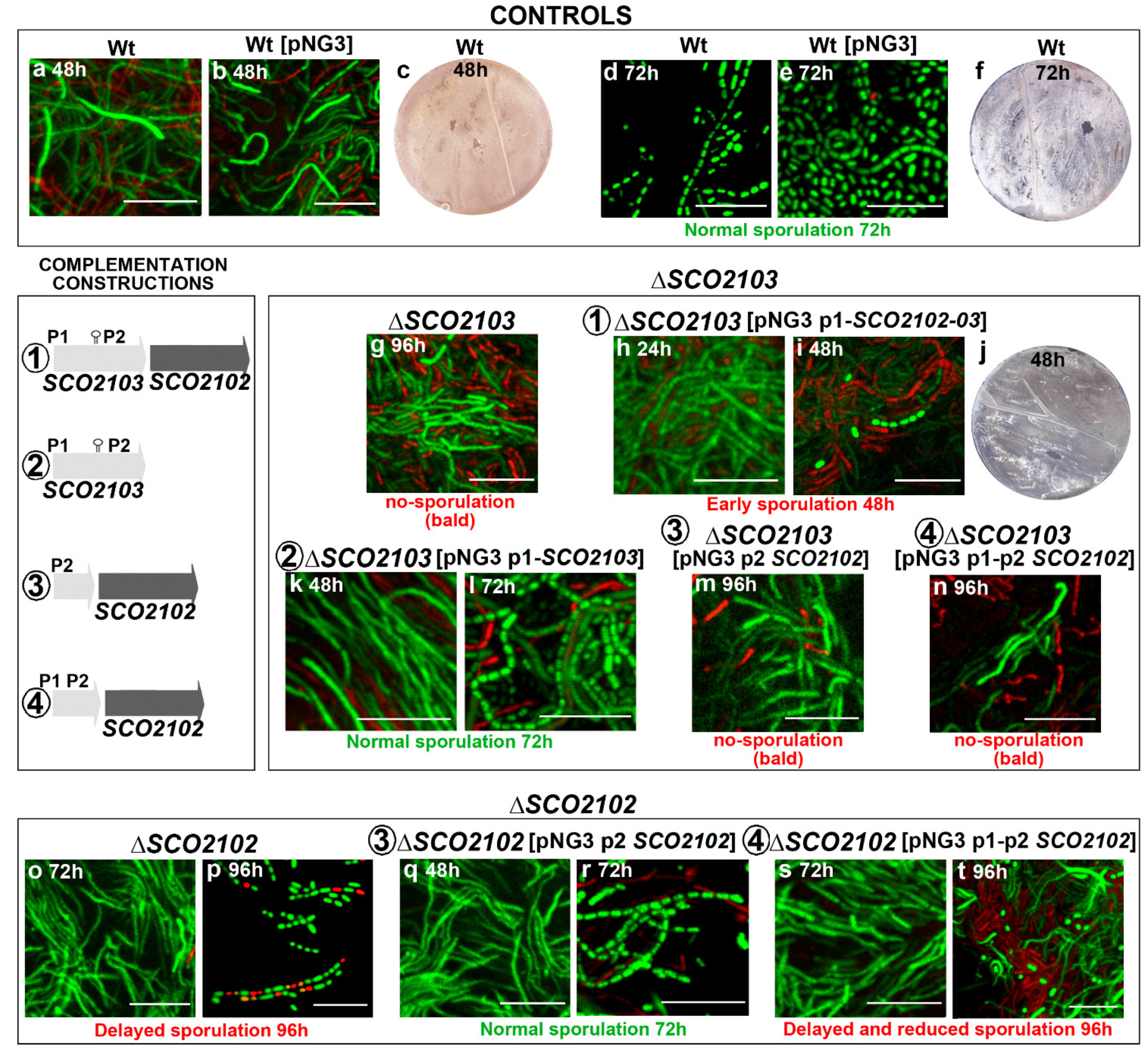
2.4. ΔSCO2103 Knockout Mutant Sporulation Is Restored by the SCO2103 Gene
2.5. ΔSCO2102 Knockout Shows a Delay in Sporulation, Which Is Complemented by the SCO2102 Gene
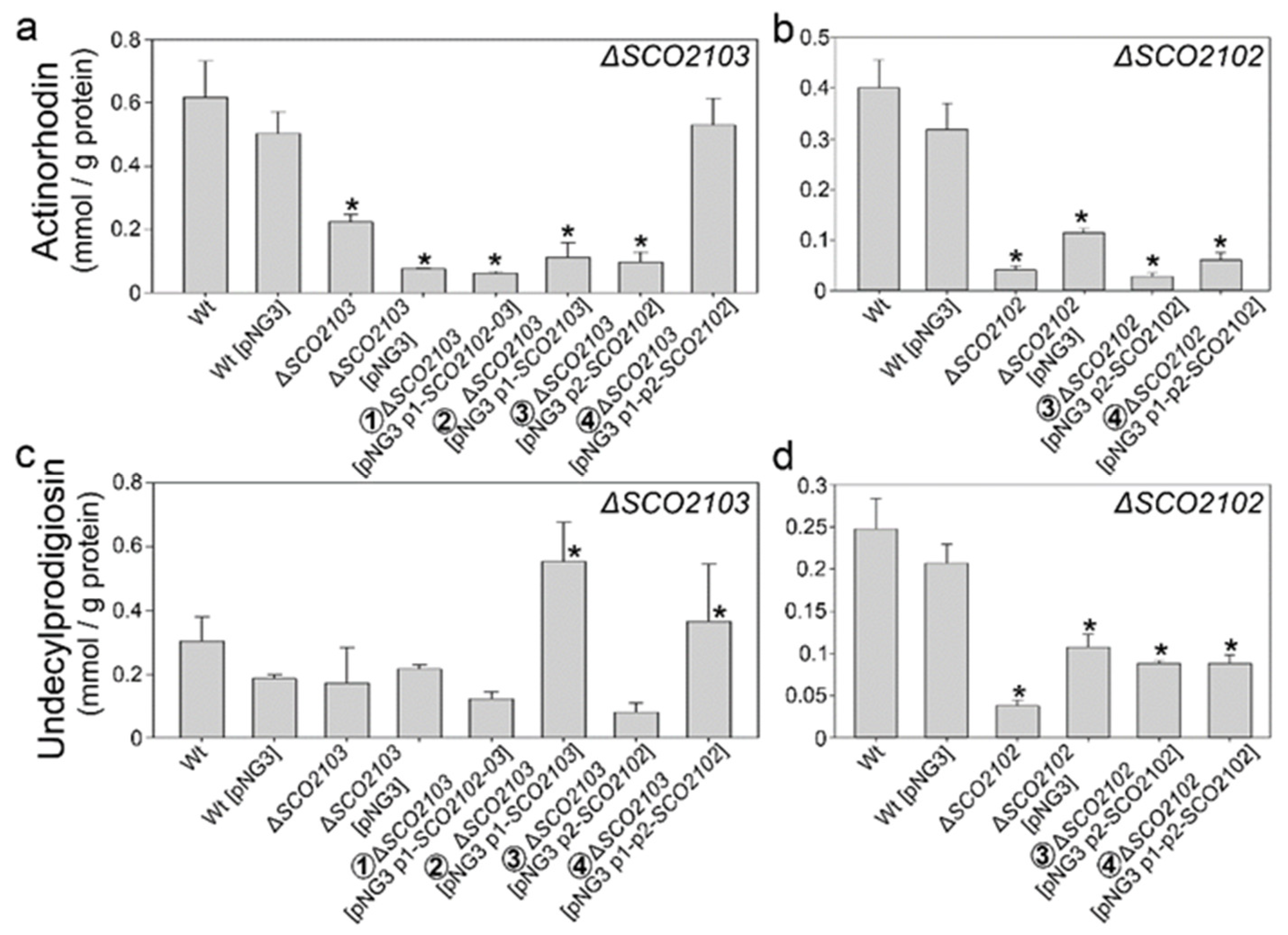
2.6. Actinorhodin and Undecylprodigiosin Productions Were Altered in the ΔSCO2102 and ΔSCO2103 Knockout Mutants
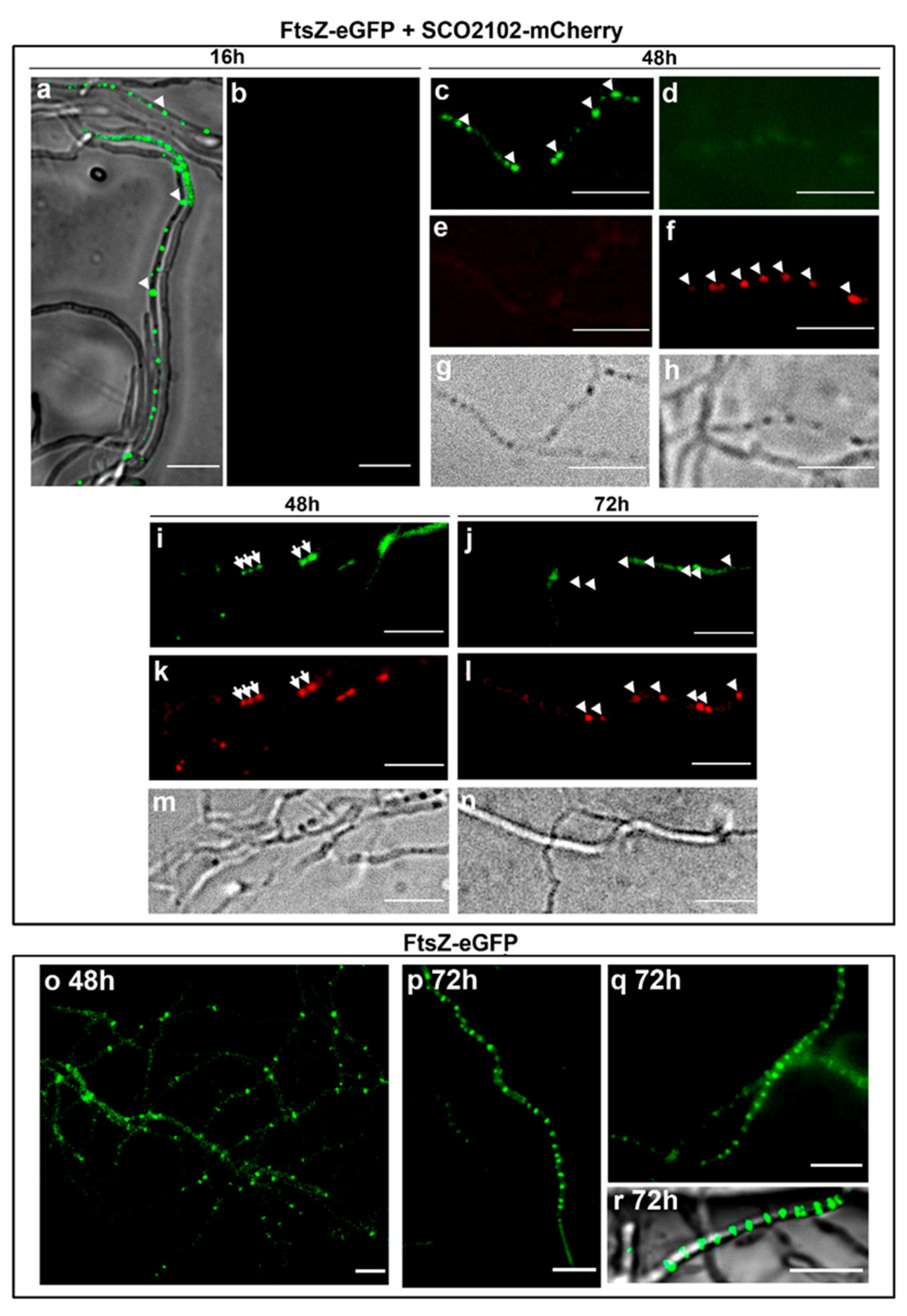
2.7. SCO2102 Co-Localises with FtsZ during Sporulation
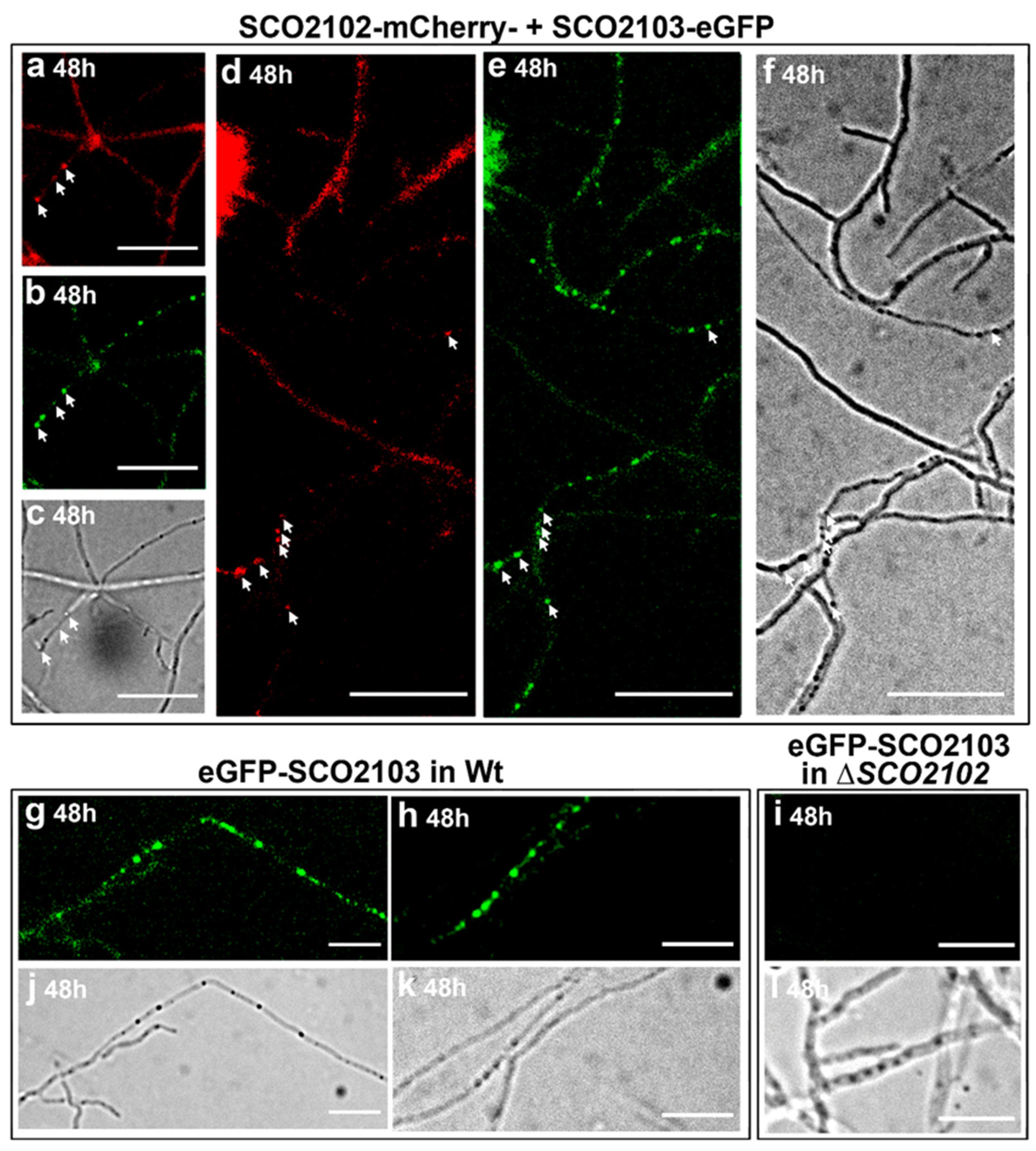
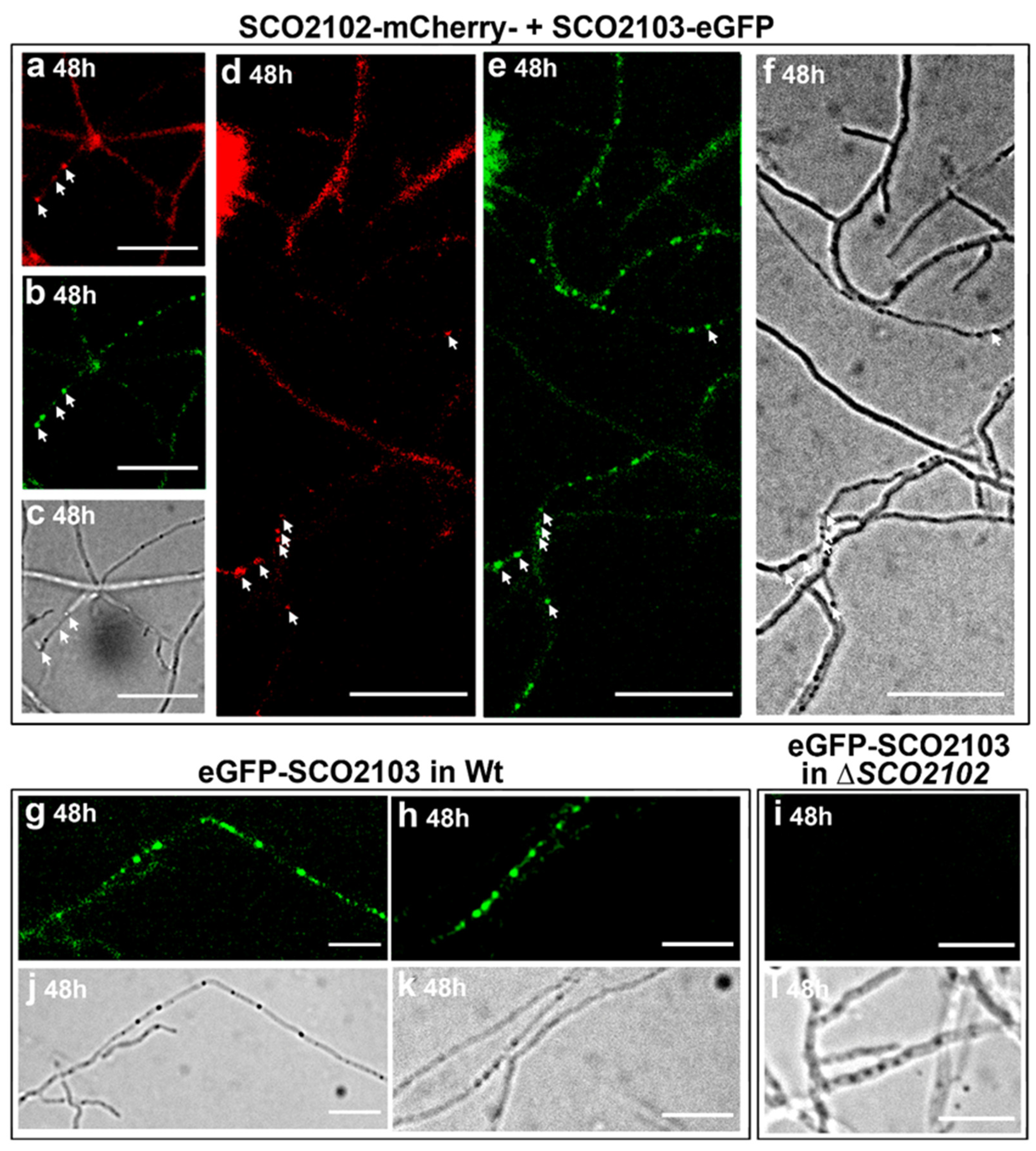
2.8. SCO2102 and SCO2103 Co-Localise during Sporulation
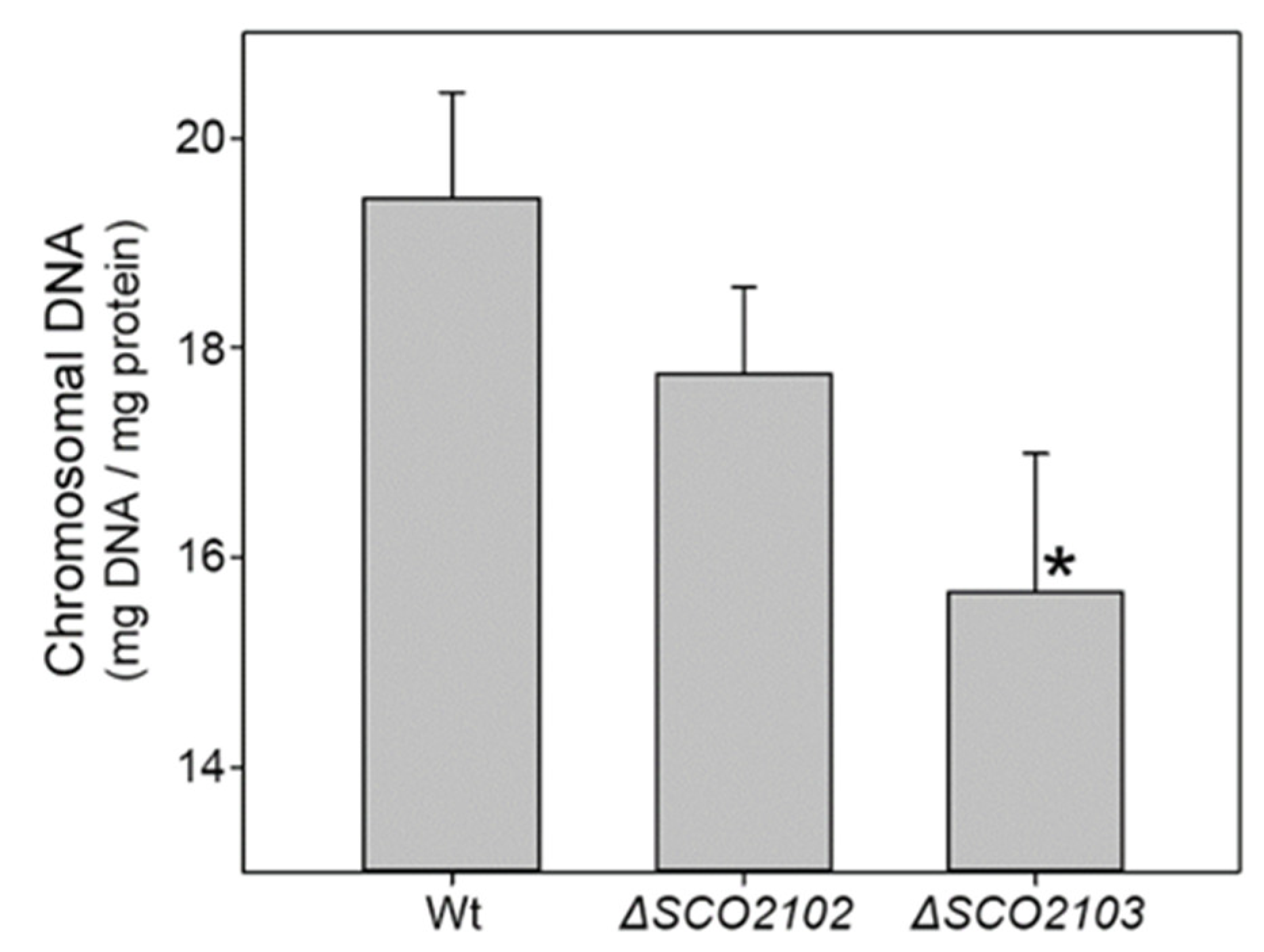
2.9. Chromosomal DNA Is Reduced in the Hyphae of the ΔSCO2103 Knockout Compared to the ΔSCO2102 Knockout and the Wild-Type Strains
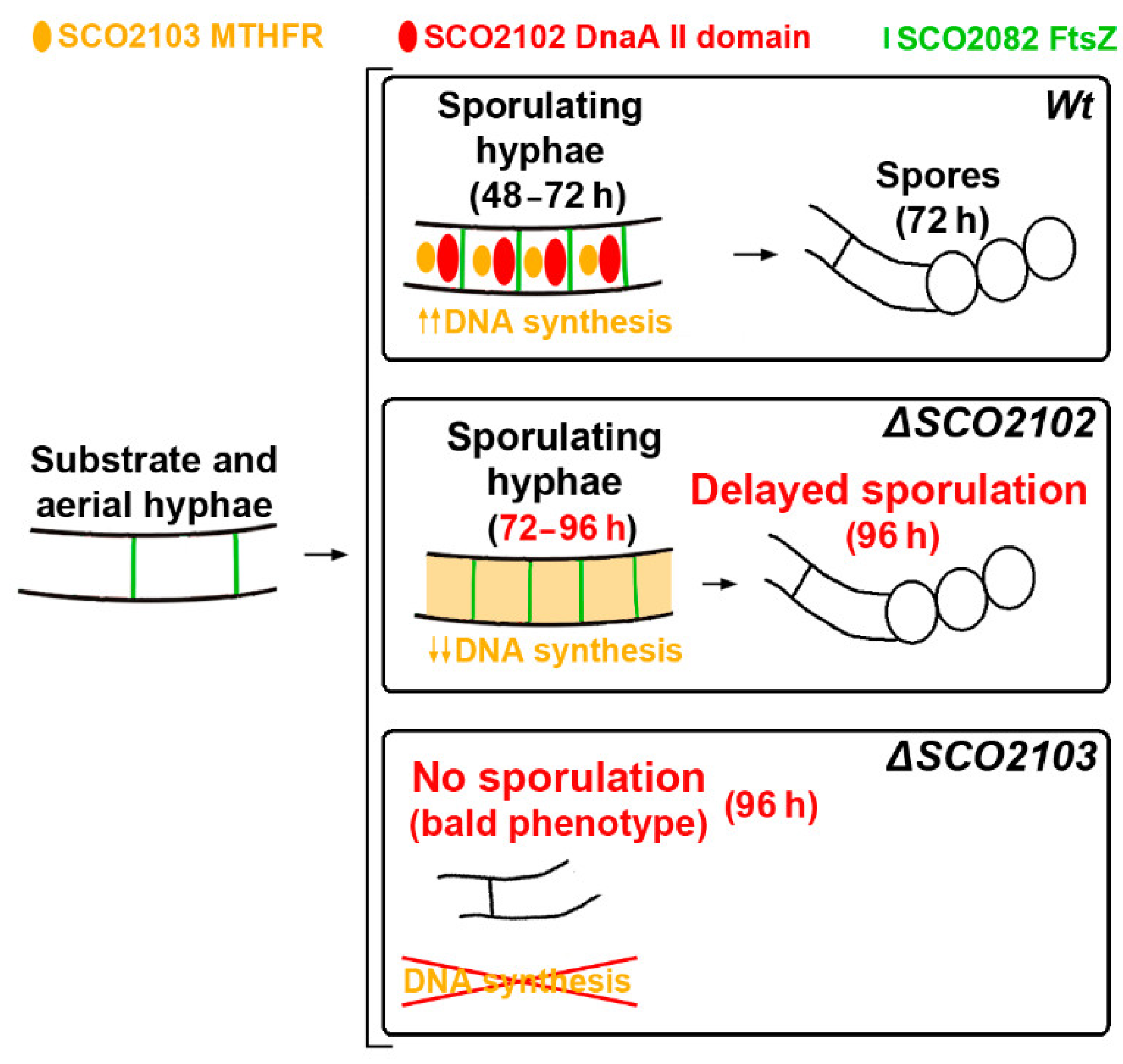
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. SCO2103 Mutagenesis
4.3. SCO2102 Mutagenesis
4.4. Complementation of ΔSCO2102 and ΔSCO2103 Mutations
4.5. SCO2102/2103 and DnaA Sequence Analyses
4.6. DNA and RNA Extraction
4.7. Real-Time Quantitative Reverse-Transcription PCR (qRT-PCR)
4.8. Antibiotic Quantification
4.9. Protein Quantification
4.10. S. coelicolor SCO2102 mCherry/eGFP and SCO2103 eGFP Constructions
4.11. Viability Staining, mCherry and eGFP Visualisation
4.12. Quantification of Chromosomal DNA
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berdy, J. Bioactive microbial metabolites. J. Antibiot. (Tokyo) 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopwood, D.A. Streptomyces in Nature and Medicine: The Antibiotic Makers; Oxford University Press: New York, NY, USA; Oxford, UK, 2007. [Google Scholar]
- Flardh, K.; Buttner, M.J. Streptomyces morphogenetics: Dissecting differentiation in a filamentous bacterium. Nat. Rev. Microbiol. 2009, 7, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Yague, P.; Lopez-Garcia, M.T.; Rioseras, B.; Sanchez, J.; Manteca, A. Pre-sporulation stages of Streptomyces differentiation: State-of-the-art and future perspectives. FEMS Microbiol. Lett. 2013, 342, 79–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakimowicz, D.; van Wezel, G.P. Cell division and DNA segregation in Streptomyces: How to build a septum in the middle of nowhere? Mol. Microbiol. 2012, 85, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Fingland, N.; Patil, D.; Sharma, A.K.; Crooke, E. Crosstalk between DnaA protein, the initiator of escherichia coli chromosomal replication, and acidic phospholipids present in bacterial membranes. Int. J. Mol. Sci. 2013, 14, 8517–8537. [Google Scholar] [CrossRef] [Green Version]
- Zawilak-Pawlik, A.; Nowaczyk, M.; Zakrzewska-Czerwińska, J. The role of the N-terminal domains of bacterial initiator DnaA in the assembly and regulation of the bacterial replication initiation complex. Genes 2017, 8, 136. [Google Scholar] [CrossRef]
- Lebkowski, T.; Wolanski, M.; Oldziej, S.; Flardh, K.; Zakrzewska-Czerwinska, J. AfsK-mediated site-specific phosphorylation regulates DnaA initiator protein activity in streptomyces coelicolor. J. Bacteriol. 2020, 202, e00597-19. [Google Scholar] [CrossRef]
- Blaauwen, T.d.; Hamoen, L.W.; Levin, P.A. The divisome at 25: The road ahead. Curr. Opin. Microbiol. 2017, 36, 85–94. [Google Scholar] [CrossRef]
- Plachetka, M.; Zyla-Uklejewicz, D.; Weigel, C.; Donczew, R.; Donczew, M.; Jakimowicz, D.; Zawilak-Pawlik, A.; Zakrzewska-Czerwinska, J. Streptomycete origin of chromosomal replication with two putative unwinding elements. Microbiology 2019, 165, 1365–1375. [Google Scholar] [CrossRef]
- McCormick, J.R.; Su, E.P.; Driks, A.; Losick, R. Growth and viability of Streptomyces coelicolor mutant for the cell division gene ftsZ. Mol. Microbiol. 1994, 14, 243–254. [Google Scholar] [CrossRef]
- McCormick, J.R. Cell division is dispensable but not irrelevant in Streptomyces. Curr. Opin. Microbiol. 2009, 12, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Willemse, J.; Borst, J.W.; de Waal, E.; Bisseling, T.; van Wezel, G.P. Positive control of cell division: FtsZ is recruited by SsgB during sporulation of Streptomyces. Genes Dev. 2011, 25, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Sol, R.; Mullins, J.G.; Grantcharova, N.; Flardh, K.; Dyson, P. Influence of CrgA on assembly of the cell division protein FtsZ during development of Streptomyces coelicolor. J. Bacteriol. 2006, 188, 1540–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantlay, S.; Sen, B.C.; Flärdh, K.; McCormick, J.R. Influence of core divisome proteins on cell division in Streptomyces venezuelae ATCC 10712. Microbiology 2021, 167, 001015. [Google Scholar] [CrossRef]
- Blanco, J.; Coque, J.J.; Martin, J.F. The folate branch of the methionine biosynthesis pathway in Streptomyces lividans: Disruption of the 5,10-methylenetetrahydrofolate reductase gene leads to methionine auxotrophy. J. Bacteriol. 1998, 180, 1586–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, S.; Varshney, U. Regulation of translation by one-carbon metabolism in bacteria and eukaryotic organelles. J. Biol. Chem. 2020, 296, 100088. [Google Scholar] [CrossRef]
- Millman, A.; Dar, D.; Shamir, M.; Sorek, R. Computational prediction of regulatory, premature transcription termination in bacteria. Nucleic Acids Res. 2017, 45, 886–893. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.; Kim, J.-N.; Kim, M.W.; Bucca, G.; Cho, S.; Yoon, Y.J.; Kim, B.-G.; Roe, J.-H.; Kim, S.C.; Smith, C.P.; et al. The dynamic transcriptional and translational landscape of the model antibiotic producer Streptomyces coelicolor A3(2). Nat. Commun. 2016, 7, 11605. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, W.; Li, X.; Fan, K.; Yang, K. Genome-wide identification and characterization of reference genes with different transcript abundances for Streptomyces coelicolor. Sci. Rep. 2015, 5, 15840. [Google Scholar] [CrossRef] [Green Version]
- Willemse, J.; van Wezel, G.P. Imaging of Streptomyces coelicolor A3 with reduced autofluorescence reveals a novel stage of FtsZ localization. PLoS ONE 2009, 4, e4242. [Google Scholar] [CrossRef] [Green Version]
- Yagüe, P.; Willemse, J.; Koning, R.I.; Rioseras, B.; López-García, M.T.; Gonzalez-Quiñonez, N.; Lopez-Iglesias, C.; Shliaha, P.V.; Rogowska-Wrzesinska, A.; Koster, A.J.; et al. Subcompartmentalization by cross-membranes during early growth of Streptomyces hyphae. Nat. Commun. 2016, 7, 12467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Huh, J.-H.; Yang, Y.-Y.; Kang, C.-M.; Lee, I.-H.; Hyun, C.-G.; Hong, S.-K.; Suh, J.-W. Accumulation of S -Adenosyl -l- Methionine Enhances Production of Actinorhodin but Inhibits Sporulation in Streptomyces lividans TK23. J. Bacteriol. 2003, 185, 592–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manteca, A.; Yague, P. Streptomyces Differentiation in Liquid Cultures as a Trigger of Secondary Metabolism. Antibiotics 2018, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plachetka, M.; Krawiec, M.; Zakrzewska-Czerwinska, J.; Wolanski, M. AdpA positively regulates morphological differentiation and chloramphenicol biosynthesis in Streptomyces venezuelae. Microbiol. Spectr. 2021, 9, e0198121. [Google Scholar] [CrossRef]
- Kieser, T. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Novella, I.S.; Barbés, C.; Sánchez, J. Sporulation of Streptomyces antibioticus ETHZ 7451 in submerged culture. Can. J. Microbiol. 1992, 38, 769–773. [Google Scholar] [CrossRef]
- Sanchez, C.; Butovich, I.A.; Brana, A.F.; Rohr, J.; Mendez, C.; Salas, J.A. The biosynthetic gene cluster for the antitumor rebeccamycin: Characterization and generation of indolocarbazole derivatives. Chem. Biol. 2002, 9, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Aparicio, J.F.; Colina, A.J.; Ceballos, E.; Martin, J.F. The biosynthetic gene cluster for the 26-membered ring polyene macrolide pimaricin. A new polyketide synthase organization encoded by two subclusters separated by functionalization genes. J. Biol. Chem. 1999, 274, 10133–10139. [Google Scholar] [CrossRef] [Green Version]
- MacNeil, D.J.; Gewain, K.M.; Ruby, C.L.; Dezeny, G.; Gibbons, P.H.; MacNeil, T. Analysis of Streptomyces avermitilis genes required for avermectin biosynthesis utilizing a novel integration vector. Gene 1992, 111, 61–68. [Google Scholar] [CrossRef]
- Flett, F.; Mersinias, V.; Smith, C.P. High efficiency intergeneric conjugal transfer of plasmid DNA from Escherichia coli to methyl DNA-restricting streptomycetes. FEMS Microbiol. Lett. 1997, 155, 223–229. [Google Scholar] [CrossRef]
- Gonzalez-Quinonez, N.; Lopez-Garcia, M.T.; Yague, P.; Rioseras, B.; Pisciotta, A.; Alduina, R.; Manteca, A. New PhiBT1 site-specific integrative vectors with neutral phenotype in Streptomyces. Appl. Microbiol. Biotechnol. 2016, 100, 2797–2808. [Google Scholar] [CrossRef]
- Perez-Redondo, R.; Santamarta, I.; Bovenberg, R.; Martin, J.F.; Liras, P. The enigmatic lack of glucose utilization in Streptomyces clavuligerus is due to inefficient expression of the glucose permease gene. Microbiology 2010, 156, 1527–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Y.; Charusanti, P.; Zhang, L.; Weber, T.; Lee, S.Y. CRISPR-Cas9 Based Engineering of Actinomycetal Genomes. ACS Synth. Biol. 2015, 4, 1020–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rioseras, B.; Yagüe, P.; García, M.T.L.; Gonzalez-Quiñonez, N.; Binda, E.; Marinelli, F.; Manteca, A. Characterization of SCO4439, a D-alanyl-D-alanine carboxypeptidase involved in spore cell wall maturation, resistance and germination in Streptomyces coelicolor. Sci. Rep. 2016, 6, 21659. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Shin, M.K.; Ryu, D.K.; Kim, S.; Ryu, W.-S. Insertion and deletion mutagenesis by overlap extension PCR. In sertion and deletion mutagenesis by overlap extension PCR. In In Vitro Mutagenesis Protocols, 3rd ed.; Braman, J., Ed.; Methods in Molecular Biology (Methods and Protocols); Humana: Totowa, NJ, USA, 2010; Volume 634. [Google Scholar]
- Gregory, M.A.; Till, R.; Smith, M.C. Integration site for Streptomyces phage phiBT1 and development of site-specific integrating vectors. J. Bacteriol. 2003, 185, 5320–5323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutledge, R.G.; Cote, C. Mathematics of quantitative kinetic PCR and the application of standard curves. Nucleic Acids Res. 2003, 31, e93. [Google Scholar] [CrossRef] [PubMed]
- Tsao, S.W.; Rudd, B.A.; He, X.G.; Chang, C.J.; Floss, H.G. Identification of a red pigment from Streptomyces coelicolor A3 as a mixture of prodigiosin derivatives. J. Antibiot. (Tokyo) 1985, 38, 128–131. [Google Scholar] [CrossRef] [Green Version]
- Bystrykh, L.V.; Fernandez-Moreno, M.A.; Herrema, J.K.; Malpartida, F.; Hopwood, D.A.; Dijkhuizen, L. Production of actinorhodin-related "blue pigments" by Streptomyces coelicolor A3. J. Bacteriol. 1996, 178, 2238–2244. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Combes, P.; Till, R.; Bee, S.; Smith, M.C. The streptomyces genome contains multiple pseudo-attB sites for the (phi)C31-encoded site-specific recombination system. J. Bacteriol. 2002, 184, 5746–5752. [Google Scholar] [CrossRef] [Green Version]
- Manteca, A.; Alvarez, R.; Salazar, N.; Yague, P.; Sanchez, J. Mycelium differentiation and antibiotic production in submerged cultures of Streptomyces coelicolor. Appl. Environ. Microbiol. 2008, 74, 3877–3886. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, K. A study of the conditions and mechanism of the diphenylamine reaction for the colorimetric estimation of deoxyribonucleic acid. Biochem. J. 1956, 62, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterborg, J.H.; Matthews, H.R. The burton assay for DNA. Methods Mol. Biol. 1985, 2, 1–3. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Reference |
|---|---|---|
| S.coelicolor M145 | SCP1- SCP2-, reference strain | [26] |
| S. coelicolor ΔSCO2102 | SCO2102 eliminated by CRISPR−Cas9 | This study |
| S. coelicolor ΔSCO2103 | SCO2102 eliminated by CRISPR−Cas9 | This study |
| E. coli TOP10 | F- mcrA Δ (mrr−hsdRMS−mcrBC) φ80lacZΔM15 ΔlacX74 recA1 araD139 Δ (ara-leu)7697 galU galK rpsL endA1 nupG | Invitrogen |
| E. coli ET12567 | dam-13::Tn9, dcm-6, hsdM, hsdR | [30] |
| E. coli ET12567/pUZ8002 | E. coli ET12567 harbouring pUZ8002, a non-self-transmissible plasmid which can mobilize oriT−containing plasmids by conjugation | [31] |
| Plasmids | Description | Reference |
| pMV | Cloning vector; AmpR | BGI |
| pKMV | Cloning vector; KmR | BGI |
| pNG3 | Integrative and conjugative vector, HygroR | [32] |
| pRAS | pRA modified by Antonio Rodríguez and Alberto−Sola−Landa | [33] |
| pNG3 p2−SCO2102 | pNG3 harbouring p2−SCO2102, HygroR | This study |
| pNG3 p1−p2−SCO2102 | pNG3 harbouring p1−p2−SCO2102, HygroR | This study |
| pNG3 SCO2103 | pNG3 harbouring SCO2103, HygroR | This study |
| pNG3 SCO2102-03 | pNG3 harbouring SCO2102 and SCO2103, HygroR | This study |
| PCR™-Blunt II−TOPO® | Zero Blunt® TOPO® PCR Cloning Kit, KanR | Invitrogen |
| pCRISPR−Cas9 | Conjugative and thermosensitive plasmid harbouring Cas9 | [34] |
| pCRISPR−2102 | pCRISPR harbouring the target SCO2102 sequence and the 2.13 kb fragment used to create the SCO2102 knockout | This study |
| pCRISPR−2103 | pCRISPR harbouring the target SCO2103 sequence and the 2.61 kb fragment used to create the SCO2103 knockout | This study |
| pNG3−p2−SCO2102−mCherry | pNG3 harbouring SCO2102, mCherry and p2 | This study |
| pNG3−p2−SCO2102-eGFP | pNG3 harbouring SCO2102, eGFP and p2 | This study |
| pRAS−p1−SCO2103−eGFP | pRAS harbouring p1, SCO2103 and eGFP | This study |
| pNG3−p1−conditional terminator−p2−SCO2102 | pNG3 harbouring p1, the conditional terminator, p2 and SCO2102 | This study |
| Primer | Sequence | Reference |
| SCO2102F | AAAAAGATATCCTCACAGCCAAGGACGATCC | This study |
| SCO2102R | CCCACTAGTGGACTCGTGGTGGAGGGG | This study |
| SCO4848F | CGTCGTATCCCCTCGGTTG | [35] |
| pMS82R | GAGCCGGGAAAGCTCATTCA | [35] |
| M13 F | CAGGAAACAGCTATGA | Invitrogen |
| M13 R | CTGGCCGTCGTTTTAC | Invitrogen |
| 2102 leftF | CGGTTGGGTGACCGCCGCA | This study |
| 2102leftR | GCGGCTCCGGCTTCCTCTTCCGTACGCCCCTCTCCCAGTGGC | This study |
| 2102 rightF | GCCACTGGGAGAGGGGCGTACGGAAGAGGAAGCCGGAGCCGC | This study |
| 2102 rightR | AGGAGCCGCTGTGGCCCAAC | This study |
| sg2102F | GGCTTCCTCTTCCTCAGAAG | This study |
| sgRNA−R | ACGCCTACGTAAAAAAAGCACCGACTCGGTGCC | This study |
| CAS91R | GTAGTACGGGATGCGGAAGG | This study |
| CAS9−1F | ATTACTGGACCGGATCGGG | This study |
| 2103F | AACCTGGACGAGGTGCTGG | This study |
| 2103R | CGTTACGCAGTGTGTCGCAAAT | This study |
| 210203R | TGCGCGACCGTCTCCTGT | This study |
| sg2103F | CCGGCGGCTGTACGCTCGAT | This study |
| 2103 LeftF | GTAGCCAGGTTTCCGACTGT | This study |
| 2103 LeftR | GATCGTCCTTGGCTGTGAGATGGGCAGGTTAGCCAGGGT | This study |
| 2103 RightF | ACCCTGGCTAACCTGCCCAT CTCACAGCCAAGGACGATC | This study |
| 2103 RightR | TCTTCCTCAGAAGCGGTACT | This study |
| TerF | AACTGGTCAGGCTCATCAAGG | This study |
| TerR | TTCCGCGGCTCCCAGT | This study |
| 2103R2 | GGCCCGCGGTGGGTGG | This study |
| q2103F | GGTGACCAGTGTGAAGATG | This study |
| q2103R | GAGTTGTTGAGCGTGATGA | This study |
| q2102F | AACTCGTACGGCGTCTA | This study |
| q2102R | ATGTACGGGTCGGAGTAG | This study |
| q4758F | ATCACCGACCGGATGCCCTT | [20] |
| q4758R | GCCGAGCCCCGCTTCTTC | [20] |
| SCO3798intF | CAGCTCGTCCTTGGTGTTCA | This study |
| SCO3798intR | TCAGGTCCATGACGTTTCCC | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-García, G.; González-Quiñónez, N.; Rioseras, B.; Alonso-Fernández, S.; Fernández, J.; Lombó, F.; Manteca, Á. The SCO2102 Protein Harbouring a DnaA II Protein-Interaction Domain Is Essential for the SCO2103 Methylenetetrahydrofolate Reductase Positioning at Streptomyces Sporulating Hyphae, Enhancing DNA Replication during Sporulation. Int. J. Mol. Sci. 2022, 23, 4984. https://doi.org/10.3390/ijms23094984
Fernández-García G, González-Quiñónez N, Rioseras B, Alonso-Fernández S, Fernández J, Lombó F, Manteca Á. The SCO2102 Protein Harbouring a DnaA II Protein-Interaction Domain Is Essential for the SCO2103 Methylenetetrahydrofolate Reductase Positioning at Streptomyces Sporulating Hyphae, Enhancing DNA Replication during Sporulation. International Journal of Molecular Sciences. 2022; 23(9):4984. https://doi.org/10.3390/ijms23094984
Chicago/Turabian StyleFernández-García, Gemma, Nathaly González-Quiñónez, Beatriz Rioseras, Sergio Alonso-Fernández, Javier Fernández, Felipe Lombó, and Ángel Manteca. 2022. "The SCO2102 Protein Harbouring a DnaA II Protein-Interaction Domain Is Essential for the SCO2103 Methylenetetrahydrofolate Reductase Positioning at Streptomyces Sporulating Hyphae, Enhancing DNA Replication during Sporulation" International Journal of Molecular Sciences 23, no. 9: 4984. https://doi.org/10.3390/ijms23094984
APA StyleFernández-García, G., González-Quiñónez, N., Rioseras, B., Alonso-Fernández, S., Fernández, J., Lombó, F., & Manteca, Á. (2022). The SCO2102 Protein Harbouring a DnaA II Protein-Interaction Domain Is Essential for the SCO2103 Methylenetetrahydrofolate Reductase Positioning at Streptomyces Sporulating Hyphae, Enhancing DNA Replication during Sporulation. International Journal of Molecular Sciences, 23(9), 4984. https://doi.org/10.3390/ijms23094984