
Antigen Presentation and Autophagy in Teleost Adaptive Immunity


Abstract
:1. Introduction
Antigen Processing and Presentation (APP) in Adaptive Immunity
2. Antigen Processing and Presentation and Autophagy in Higher Vertebrates
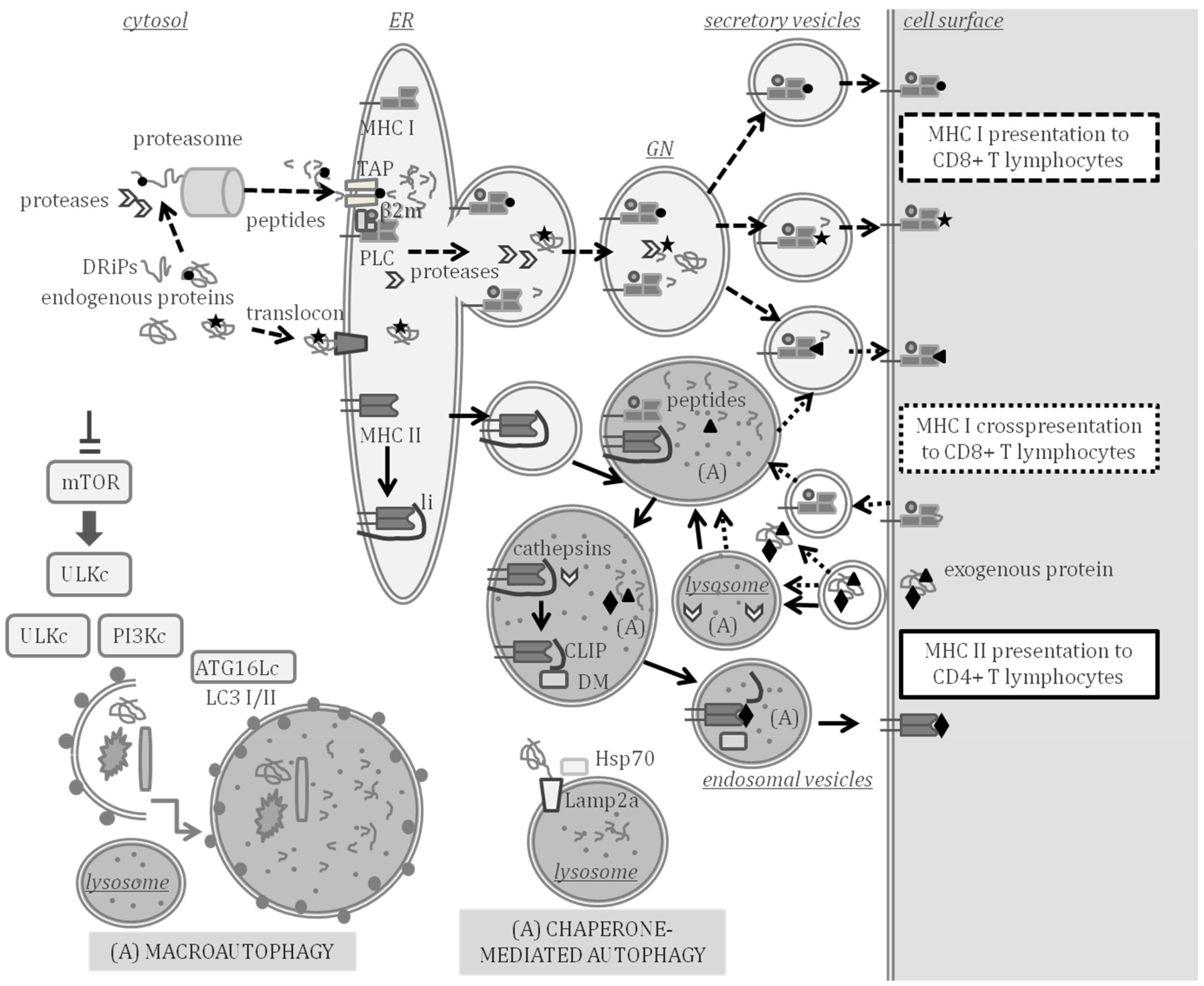
2.1. Antigen Processing and Presentation in Adaptive Immunity
2.2. Autophagy in Adaptive Immunity
3. Antigen Processing and Presentation in Teleost Fish
3.1. Antigen Processing and Presentation Genes Conserved in Teleost Fish
3.2. Antigen-Presenting Cells (APC) and Professional APC (pAPC) in Teleost Fish
3.3. Peptide Processing, Loading and Transport by MHC in Teleost Fish
4. Autophagy and Related LC3-Associated Phagocytosis in Teleost Fish
4.1. Chaperone-Mediated Autophagy in Teleost Fish
4.2. Macroautophagy in Teleost Fish
4.3. LC3-Associated Phagocytosis in Teleost Fish
5. Regulation of Antigen Processing and Presentation and Autophagy during Infection of Teleost Fish
5.1. Modulation of Antigen Processing and Presentation and Autophagy during Viral Infection

{kind=link}
| Virus 1 | Host Species | MHC I 2 APP | MHC II 2 APP | Autophagy 2 | Organ or Cell | Reference |
|---|---|---|---|---|---|---|
| VHSV | Lateolabrax japonicus | n.d. | − | n.d. | Brain cell line. | [105] |
| Sander vitreus | + | n.d. | n.d. | Skin fibroblast cell line. | [106] | |
| Oncorhynchus mykiss | + | n.d. | n.d. | Skin fibroblast cell line. | [107] | |
| + | + | + | Red blood cells and head-kidney. | [108,109] | ||
| n.d. | + | n.d. | Blood and spleen B cells. | [84] | ||
| Scophthalmus maximus | n.d. | n.d. | + | Red blood cells. | [73] | |
| Paralichthys olivaceus | n.d. | n.d. | + | Kidney and spleen. | [110] | |
| NNV | Epinephelus coioides | + | n.d. | n.d. | Liver and spleen. | [47] |
| Sparus aurata | + | − | −/+ | Larvae. | [111] | |
| CSV | Oncorhynchus mykiss | + | n.d. | n.d. | Monocyte/macrophage cell line. | [44] |
| GCRV | Ctenopharyngodon idella | n.d. | n.d. | + | Kidney cell line. | [112] |
| SGIV | Epinephelus coioides | + | n.d. | n.d. | Liver and spleen. | [47] |
| IHNV | Oncorhynchus mykiss | + | − | n.d. | Spleen, intestines and head-kidney. | [113] |
| RBIV | Cynoglossus semilaevis | n.d. | + | n.d. | Kidney and spleen. | [61] |
| SAV3 | Salmo salar | + | n.d. | n.d. | Pancreas, heart, spleen, head-kidney, liver and gills. | [114] |
| ISKNV | Paralichthys olivaceus | + | n.d. | n.d. | Head-kidney, spleen and liver. | [115] |
5.2. Modulation of Antigen Processing and Presentation and Autophagy during Bacterial or Parasite Infection
| Pathogen 1 | Host Species | MHC I 2 APP | MHC II 2 APP | Autophagy 2 | Organ or Cell | Reference |
|---|---|---|---|---|---|---|
| Piscirickettsia salmonis | Salmo salar | +/(=) | n.d. | n.d. | Heart, spleen, head-kidney, liver and gill. | [114] |
| Vibrio anguillarum | Sebastes schlegelii | + | n.d. | − | Lymphocytes from peripheral blood, head-kidney and spleen. | [120] |
| Sparus aurata | n.d. | + | n.d. | Head-kidney leucocytes. | [53] | |
| Cynoglossus semilaevis | n.d. | + | n.d. | Kidney and liver. | [61] | |
| Miichthys miiuy | n.d. | + | n.d. | Liver, spleen and kidney. | [64] | |
| Paralichthys olivaceus | + | n.d. | n.d. | Head-kidney, spleen and liver. | [115] | |
| Yersinia ruckeri | Oncorhynchus mykiss | n.d. | + | n.d. | Head-kidney and spleen. | [121] |
| Edwarsiella ictaluri | Ictalurus furcatus | + | n.d. | n.d. | Liver. | [119] |
| Ictalurus punctatus | n.d. | + | n.d. | Skin, gill and intestine. | [65] | |
| Edwardsiella tarda | Paralichthys olivaceus | + | n.d. | n.d. | Head-kidney, spleen and liver. | [115] |
| Trachinotus ovatus | n.d. | + | n.d. | Liver, spleen and head-kidney. | [66] | |
| Citrobacter freundii | Danio rerio | + | − | + | Skin. | [122] |
| Streptococcus agalactiae3 | Danio rerio | − | − | n.d. | Intestine and skin. | [123] |
| Staphylococcus aureus3 | Danio rerio | n.d. | n.d. | + | Macrophages and neutrophils. | [98,103] |
| Salmonella typhimurium3 | Danio rerio | n.d. | n.d. | + | Macrophages and neutrophils. | [98] |
| n.d. | n.d. | +/− | Macrophages and larvae. | [102,124] | ||
| Saccharomyces cerevisiae | Sparus aurata | n.d. | + | n.d. | Head-kidney leucocytes. | [53] |
| Enteromyxum scophthalmi | Scophthalmus maximus | − | n.d. | n.d. | Blood. | [72] |
| Neoparamoeba perurans | Salmo salar | − | n.d. | n.d. | Gills. | [125] |
| Lepeophtheirus salmonis | Salmo salar | − | − | n.d. | Skin and head-kidney. | [126] |
| Gyrodactylus bullatarudis | Poecilia reticulata | + | = | n.d. | Skin. | [127] |
| Cryptocaryon irritans | Epinephelus coioides | + | + | n.d. | Skin. | [80] |
| Sparus aurata | n.d. | −/= | n.d. | Gills and head-kidney. | [128] |
5.3. Modulation of Antigen Processing and Presentation and Autophagy upon Vaccination
| Vaccine 1 | Host Species | MHC I 2 APP | MHC II 2 APP | Autophagy 2 | Organ or Cell | Reference |
|---|---|---|---|---|---|---|
| Vibrio harvey | Cynoglossus semilaevis | n.d. | + | n.d. | Kidney. | [61] |
| Edwardsiella tarda | Danio rerio | + | − | n.d. | Liver and spleen. | [135,136] |
| Vibrio anguillarum and Edwardsiella tarda | Scophthalmus maximus | + | + | n.d. | Liver and spleen, kidney. | [137] |
| Danio rerio | + | + | n.d. | Liver and spleen. | [137] | |
| Vibrio parahaemolyticus, Vibrio alginolyticus and Aeromonas hydrophila | Larimichthys crocea | + | + | n.d. | Spleen. | [138] |
| VHSV | Danio rerio | n.d. | n.d. | + | Fibroblast cell line and muscle. | [139] |
| SVCV | Danio rerio | n.d. | n.d. | + | Fibroblast cell line and muscle. | [139] |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galluzzi, L.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cecconi, F.; Choi, A.M.; Chu, C.T.; Codogno, P.; Colombo, M.I.; et al. Molecular definitions of autophagy and related processes. EMBO J. 2017, 36, 1811–1836. [Google Scholar] [CrossRef]
- Münz, C. The macroautophagy machinery in MHC restricted antigen presentation. Front. Immunol. 2021, 12, 1–7. [Google Scholar] [CrossRef]
- Münz, C. Non-canonical functions of autophagy proteins in immunity and infection. Mol. Asp. Med. 2021, 82, 100987. [Google Scholar] [CrossRef]
- Rock, K.L.; Reits, E.; Neefjes, J. Present yourself! By MHC class I and MHC class II molecules. Trends Immunol. 2016, 37, 724–737. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.B. MHC and adaptive immunity in teleost fishes. Immunogenetics 2017, 69, 521–528. [Google Scholar] [CrossRef]
- Yewdell, J.W.; Antón, L.C.; Bennink, J.R. Defective ribosomal products (DRiPs): A major source of antigenic peptides for MHC class I molecules? J. Immunol. 1996, 157, 1823–1826. [Google Scholar]
- Wu, Y.; Zhang, N.; Hashimoto, K.; Xia, C.; Dijkstra, J.M. Structural comparison between MHC classes I and II; in evolution, a Class-II-like molecule probably came first. Front. Immunol. 2021, 12, 621153. [Google Scholar] [CrossRef]
- De Groot, A.S.; Moise, L.; Terry, F.; Gutierrez, A.H.; Hindocha, P.; Richard, G.; Hoft, D.F.; Ross, T.M.; Noe, A.R.; Takahashi, Y.; et al. Better epitope discovery, precision immune engineering, and accelerated vaccine design using Immunoinformatics tools. Front. Immunol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Abi Habib, J.; Lesenfants, J.; Vigneron, N.; Van den Eynde, B.J. Functional differences between proteasome subtypes. Cells 2022, 11, 421. [Google Scholar] [CrossRef]
- Blees, A.; Januliene, D.; Hofmann, T.; Koller, N.; Schmidt, C.; Trowitzsch, S.; Moeller, A.; Tampé, R. Structure of the human MHC-I peptide-loading complex. Nature 2017, 551, 525–528. [Google Scholar] [CrossRef]
- Thomas, C.; Tampé, R. MHC I assembly and peptide editing—chaperones, clients, and molecular plasticity in immunity. Curr. Opin. Immunol. 2021, 70, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, C.; Del Val, M. Traffic of proteins and peptides across membranes for immunosurveillance by CD8+ T lymphocytes: A topological challenge. Traffic 2007, 8, 1486–1494. [Google Scholar] [CrossRef] [PubMed]
- Del-Val, M.; López, D. Multiple proteases process viral antigens for presentation by MHC class I molecules to CD8+ T lymphocytes. Mol. Immunol. 2002, 39, 235–247. [Google Scholar] [CrossRef]
- Serwold, T.; Gonzalez, F.; Kim, J.; Jacob, R.; Shastri, N. ERAAP customizes peptides for MHC class I molecules in the endoplasmic reticulum. Nature 2002, 419, 480–483. [Google Scholar] [CrossRef]
- Chang, S.-C.; Momburg, F.; Bhutani, N.; Goldberg, A.L. The ER aminopeptidase, ERAP1, trims precursors to lengths of MHC class I peptides by a “molecular ruler” mechanism. Proc. Natl. Acad. Sci. USA 2005, 102, 17107–17112. [Google Scholar] [CrossRef] [Green Version]
- Gil-Torregrosa, B.C.; Castaño, A.R.; Del Val, M. Major histocompatibility complex class I viral antigen processing in the secretory pathway defined by the trans-Golgi network protease furin. J. Exp. Med. 1998, 188, 1105–1116. [Google Scholar] [CrossRef]
- Grommé, M.; Uytdehaag, F.G.C.M.; Janssen, H.; Calafat, J.; van Binnendijk, R.S.; Kenter, M.J.H.; Tulp, A.; Verwoerd, D.; Neefjes, J. Recycling MHC class I molecules and endosomal peptide loading. Proc. Natl. Acad. Sci. USA 1999, 96, 10326–10331. [Google Scholar] [CrossRef] [Green Version]
- Rock, K.L.; Shen, L. Cross-presentation: Underlying mechanisms and role in immune surveillance. Immunol. Rev. 2005, 207, 166–183. [Google Scholar] [CrossRef]
- Possamaï, D.; Hanafi, L.A.; Bellemare-Pelletier, A.; Hamelin, K.; Thébault, P.; Hébert, M.J.; Gagnon, É.; Leclerc, D.; Lapointe, R. MHC class I antigen cross-presentation mediated by PapMV nanoparticles in human antigen-presenting cells is dependent on autophagy. PLoS ONE 2021, 16, e0261987. [Google Scholar] [CrossRef]
- Johnstone, C.; Ramos, M.; García-Barreno, B.; López, D.; Melero, J.A.; Del Val, M. Exogenous, TAP-independent lysosomal presentation of a respiratory syncytial virus CTL epitope. Immunol. Cell Biol. 2012, 90, 978–982. [Google Scholar] [CrossRef] [Green Version]
- Schröder, B. The multifaceted roles of the invariant chain CD74—More than just a chaperone. Biochim. Biophys. Acta 2016, 1863, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.A. Endosomal proteolysis and MHC class II function. Curr. Opin. Immunol. 1998, 10, 93–102. [Google Scholar] [CrossRef]
- Romagnoli, P.; Germain, R.N. The CLIP region of invariant chain plays a critical role in regulating major histocompatibility complex class II folding, transport, and peptide occupancy. J. Exp. Med. 1994, 180, 1107–1113. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Vargas, E.; Barker, A.P.; Zhou, Z.; He, X.; Jensen, P.E. HLA-DM catalytically enhances peptide dissociation by sensing peptide-MHC class II interactions throughout the peptide-binding cleft. J. Biol. Chem. 2020, 295, 2959–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Maben, Z.J.; Becerra, A.; Stern, L.J. Evaluating the role of HLA-DM in MHC class II-peptide association reactions. J. Immunol. 2015, 195, 706–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.K.; Mattei, L.M.; Steinberg, B.E.; Alberts, P.; Lee, Y.H.; Chervonsky, A.; Mizushima, N.; Grinstein, S.; Iwasaki, A. In vivo requirement for Atg5 in antigen presentation by dendritic cells. Immunity 2010, 32, 227–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, D.; Münz, C. Innate and adaptive immunity through autophagy. Immunity 2007, 27, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Paludan, C.; Schmid, D.; Landthaler, M.; Vockerodt, M.; Kube, D.; Tuschl, T.; Münz, C. Endogenous MHC class II processing of a viral nuclear antigen after autophagy. Science 2005, 307, 593–596. [Google Scholar] [CrossRef]
- Kaushik, S.; Cuervo, A.M. Chaperone-mediated autophagy: A unique way to enter the lysosome world. Trends Cell Biol. 2012, 22, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Gomes, L.C.; Dikic, I. Autophagy in antimicrobial immunity. Mol. Cell 2014, 54, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy 2021, 17, 1–382. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, A.N.; Miller, N.W.; Clem, L.W. Phylogeny of immune recognition: Role of alloantigens in antigen presentation in channel catfish immune responses. Immunology 1991, 74, 165–168. [Google Scholar] [PubMed]
- Vallejo, A.N.; Miller, N.W.; Harvey, N.E.; Cuchens, M.A.; William Clem, L. Cellular pathway(s) of antigen processing and presentation in fish APC: Endosomal involvement and cell-free antigen presentation. Dev. Immunol. 1992, 3, 51–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Dijkstra, J.M. Major histocompatibility complex (MHC) genes and disease resistance in fish. Cells 2019, 8, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radwan, J.; Babik, W.; Kaufman, J.; Lenz, T.L.; Winternitz, J. Advances in the evolutionary understanding of MHC polymorphism. Trends Genet. 2020, 36, 298–311. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, J. Unfinished business: Evolution of the MHC and the adaptive immune system of jawed vertebrates. Annu. Rev. Immunol. 2018, 36, 383–409. [Google Scholar] [CrossRef]
- Nonaka, M.; Naruse, K.; Matsuo, M.; Shima, A. Comparative genomics of medaka: The major histocompatibility complex (MHC). Mar. Biotechnol. 2001, 3, S141–S144. [Google Scholar] [CrossRef]
- Kim, D.-H.; Lee, S.-M.; Hong, B.-Y.; Kim, Y.-T.; Choi, T.-J. Cloning and sequence analysis of cDNA for the proteasome activator PA28-β subunit of flounder (Paralichthys olivaceus). Mol. Immunol. 2003, 40, 611–616. [Google Scholar] [CrossRef]
- Liu, G.; Zheng, W.; Chen, X. Molecular cloning of proteasome activator PA28-β subunit of large yellow croaker (Pseudosciana crocea) and its coordinated up-regulation with MHC class I α-chain and β2-microglobulin in poly I:C-treated fish. Mol. Immunol. 2007, 44, 1190–1197. [Google Scholar] [CrossRef]
- Kasthuri, S.R.; Umasuthan, N.; Whang, I.; Kim, E.; Park, H.-C.; Lee, J. Genomic structural characterization and transcriptional expression analysis of proteasome activator PA28α and PA28β subunits from Oplegnathus fasciatus. Fish Shellfish Immunol. 2013, 35, 1224–1234. [Google Scholar] [CrossRef]
- Pinto, R.D.; Randelli, E.; Buonocore, F.; Pereira, P.J.B.; dos Santos, N.M.S. Molecular cloning and characterization of sea bass (Dicentrarchus labrax, L.) MHC class I heavy chain and β2-microglobulin. Dev. Comp. Immunol. 2013, 39, 234–254. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.D.; Moreira, A.R.; Pereira, P.J.B.; dos Santos, N.M.S. Two thioredoxin-superfamily members from sea bass (Dicentrarchus labrax, L.): Characterization of PDI (PDIA1) and ERp57 (PDIA3). Fish Shellfish Immunol. 2013, 35, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Sever, L.; Bols, N.C.; Dixon, B. The cloning and inducible expression of the rainbow trout ERp57 gene. Fish Shellfish Immunol. 2013, 34, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Sever, L.; Vo, N.T.K.; Bols, N.C.; Dixon, B. Expression of tapasin in rainbow trout tissues and cell lines and up regulation in a monocyte/macrophage cell line (RTS11) by a viral mimic and viral infection. Dev. Comp. Immunol. 2014, 44, 86–93. [Google Scholar] [CrossRef]
- Wilson, W.H.; Gilg, I.C.; Moniruzzaman, M.; Field, E.K.; Koren, S.; Lecleir, G.R.; Martínez-Martínez, J.; Poulton, N.J.; Swan, B.K.; Stepanauskas, R.; et al. Genomic exploration of individual giant ocean viruses. ISME J. 2017, 11, 1736–1745. [Google Scholar] [CrossRef]
- Nonaka, M.I.; Aizawa, K.; Mitani, H.; Bannai, H.P.; Nonaka, M. Retained orthologous relationships of the MHC class I genes during euteleost evolution. Mol. Biol. Evol. 2011, 28, 3099–3112. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, L.; Huang, J.; Li, X.; Guan, L.; Wang, Q.; Yang, M.; Qin, Q. Functional analysis of a novel MHC-Iα genotype in orange-spotted grouper: Effects on Singapore grouper iridovirus (SGIV) replication and apoptosis. Fish Shellfish Immunol. 2022, 121, 487–497. [Google Scholar] [CrossRef]
- Hashimoto, K.; Nakanishi, T.; Kurosawa, Y. Isolation of carp genes encoding major histocompatibility complex antigens. Proc. Natl. Acad. Sci. USA 1990, 87, 6863–6867. [Google Scholar] [CrossRef] [Green Version]
- McConnell, S.C.; Hernández, K.M.; Wcisel, D.J.; Kettleborough, R.N.; Stemple, D.L.; Yoder, J.A.; Andrade, J.; de Jong, J.L.O. Alternative haplotypes of antigen processing genes in zebrafish diverged early in vertebrate evolution. Proc. Natl. Acad. Sci. USA 2016, 113, E5014–E5023. [Google Scholar] [CrossRef] [Green Version]
- Grimholt, U. Whole genome duplications have provided teleosts with many roads to peptide loaded MHC class I molecules. BMC Evol. Biol. 2018, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Dirscherl, H.; Yoder, J.A. A nonclassical MHC class I U lineage locus in zebrafish with a null haplotypic variant. Immunogenetics 2015, 67, 501–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimholt, U.; Fosse, J.H.; Sundaram, A.Y.M. Selective stimulation of duplicated Atlantic salmon MHC pathway genes by interferon-gamma. Front. Immunol. 2020, 11, 571650–571665. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, A.; Ángeles Esteban, M.; Meseguer, J. Cloning, distribution and up-regulation of the teleost fish MHC class II alpha suggests a role for granulocytes as antigen-presenting cells. Mol. Immunol. 2006, 43, 1275–1285. [Google Scholar] [CrossRef]
- Pilstrom, L.; Warr, G.W.; Stromberg, S. Why is the antibody response of Atlantic cod so poor? The search for a genetic explanation. Fish. Sci. 2005, 71, 961–971. [Google Scholar] [CrossRef]
- Haase, D.; Roth, O.; Kalbe, M.; Schmiedeskamp, G.; Scharsack, J.P.; Rosenstiel, P.; Reusch, T.B.H. Absence of major histocompatibility complex class II mediated immunity in pipefish, Syngnathus typhle: Evidence from deep transcriptome sequencing. Biol. Lett. 2013, 9, 20130044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkstra, J.M.; Yamaguchi, T. Ancient features of the MHC class II presentation pathway, and a model for the possible origin of MHC molecules. Immunogenetics 2019, 71, 233–249. [Google Scholar] [CrossRef]
- Dijkstra, J.M.; Grimholt, U.; Leong, J.; Koop, B.F.; Hashimoto, K. Comprehensive analysis of MHC class II genes in teleost fish genomes reveals dispensability of the peptide-loading DM system in a large part of vertebrates. BMC Evol. Biol. 2013, 13, 260. [Google Scholar] [CrossRef] [Green Version]
- Fujiki, K. Alternate forms of MHC class II-associated invariant chain are not produced by alternative splicing in rainbow trout (Oncorhynchus mykiss) but are encoded by separate genes. Dev. Comp. Immunol. 2003, 27, 377–391. [Google Scholar] [CrossRef]
- Semple, S.L.; Heath, G.; Christie, D.; Braunstein, M.; Kales, S.C.; Dixon, B. Immune stimulation of rainbow trout reveals divergent regulation of MH class II-associated invariant chain isoforms. Immunogenetics 2019, 71, 407–420. [Google Scholar] [CrossRef]
- Li, M.; Li, Q.; Yang, Z.; Hu, G.; Li, T.; Chen, X.; Ao, J. Identification of cathepsin B from large yellow croaker (Pseudosciaena crocea) and its role in the processing of MHC class II-associated invariant chain. Dev. Comp. Immunol. 2014, 45, 313–320. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, M.; Sun, L. Identification and expressional analysis of two cathepsins from half-smooth tongue sole (Cynoglossus semilaevis). Fish. Shellfish Immunol. 2011, 31, 1270–1277. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ao, J.; Mu, Y.; Yang, Z.; Li, T.; Zhang, X.; Chen, X. Cathepsin S, but not cathepsin L, participates in the MHC class II-associated invariant chain processing in large yellow croaker (Larimichthys crocea). Fish. Shellfish Immunol. 2015, 47, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xu, T.; Wang, J.; Cheng, Y.; Wang, R. Sequence and expression analysis of cathepsin S gene in the miiuy croaker Miichthys miiuy. Fish. Physiol. Biochem. 2011, 37, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Che, R.; Wang, R.; Xu, T. Comparative genomic of the teleost cathepsin B and H and involvement in bacterial induced immunity of miiuy croaker. Fish. Shellfish Immunol. 2014, 41, 163–171. [Google Scholar] [CrossRef]
- Dong, X.; Ye, Z.; Song, L.; Su, B.; Zhao, H.; Peatman, E.; Li, C. Expression profile analysis of two cathepsin S in channel catfish (Ictalurus punctatus) mucosal tissues following bacterial challenge. Fish. Shellfish Immunol. 2016, 48, 112–118. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, H.; Zhou, Y.; Sun, Y.; Yang, H.; Cao, Z.; Qin, Q.; Liu, C.; Guo, W. Functional characterization of cathepsin B and its role in the antimicrobial immune responses in golden pompano (Trachinotus ovatus). Dev. Comp. Immunol. 2021, 123, 104128. [Google Scholar] [CrossRef]
- Criscitiello, M.F.; Ohta, Y.; Graham, M.D.; Eubanks, J.O.; Chen, P.L.; Flajnik, M.F. Shark class II invariant chain reveals ancient conserved relationships with cathepsins and MHC class II. Dev. Comp. Immunol. 2012, 36, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Dijkstra, J.M.; Köllner, B.; Aoyagi, K.; Sawamoto, Y.; Kuroda, A.; Ototake, M.; Nakanishi, T.; Fischer, U. The rainbow trout classical MHC class I molecule Onmy-UBA*501 is expressed in similar cell types as mammalian classical MHC class I molecules. Fish. Shellfish Immunol. 2003, 14, 1–23. [Google Scholar] [CrossRef]
- Chang, Y.T.; Kai, Y.H.; Chi, S.C.; Song, Y.L. Cytotoxic CD8α+ leucocytes have heterogeneous features in antigen recognition and class I MHC restriction in grouper. Fish. Shellfish Immunol. 2011, 30, 1283–1293. [Google Scholar] [CrossRef]
- Kanako, L. Lewis; Natasha, Del Cid; D.T. Perspectives on antigen presenting cells in zebrafish. Dev. Comp. Immunol. 2014, 46, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Iliev, D.B.; Thim, H.; Lagos, L.; Olsen, R.; Jørgensen, J.B. Homing of antigen-presenting cells in head kidney and spleen—salmon head kidney hosts diverse APC types. Front. Immunol. 2013, 4, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronza, P.; Álvarez-Dios, J.A.; Robledo, D.; Losada, A.P.; Romero, R.; Bermúdez, R.; Pardo, B.G.; Martínez, P.; Quiroga, M.I. Blood transcriptomics of turbot Scophthalmus maximus: A tool for health monitoring and disease studies. Animals 2021, 11, 1296. [Google Scholar] [CrossRef] [PubMed]
- Pereiro, P.; Romero, A.; Díaz-Rosales, P.; Estepa, A.; Figueras, A.; Novoa, B. Nucleated teleost erythrocytes play an Nk-lysin- and autophagy-dependent role in antiviral immunity. Front. Immunol. 2017, 8, 1458. [Google Scholar] [CrossRef] [PubMed]
- Lugo-Villarino, G.; Balla, K.M.; Stachura, D.L.; Bañuelos, K.; Werneck, M.B.F.; Traver, D. Identification of dendritic antigen-presenting cells in the zebrafish. Proc. Natl. Acad. Sci. USA 2010, 107, 15850–15855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, T.; Zhu, L.-Y.; Nie, L.; Shi, W.; Dong, W.-R.; Xiang, L.-X.; Shao, J.-Z. Characterization of surface phenotypic molecules of teleost dendritic cells. Dev. Comp. Immunol. 2015, 49, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Zoccola, E.; Delamare-Deboutteville, J.; Barnes, A.C. Identification of barramundi (Lates calcarifer) DC-SCRIPT, a specific molecular marker for dendritic cells in fish. PLoS ONE 2015, 10, e0132687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soleto, I.; Fischer, U.; Tafalla, C.; Granja, A.G. Identification of a potential common ancestor for mammalian cross-presenting dendritic cells in teleost respiratory surfaces. Front. Immunol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soleto, I.; Granja, A.G.; Simón, R.; Morel, E.; Díaz-Rosales, P.; Tafalla, C. Identification of CD8α+ dendritic cells in rainbow trout (Oncorhynchus mykiss) intestine. Fish. Shellfish Immunol. 2019, 89, 309–318. [Google Scholar] [CrossRef]
- Kato, G.; Miyazawa, H.; Nakayama, Y.; Ikari, Y.; Kondo, H.; Yamaguchi, T.; Sano, M.; Fischer, U. A novel antigen-sampling cell in the teleost gill epithelium with the potential for direct antigen presentation in mucosal tissue. Front. Immunol. 2018, 9, 2116–2128. [Google Scholar] [CrossRef]
- Hu, Y.; Li, A.; Xu, Y.; Jiang, B.; Lu, G.; Luo, X. Transcriptomic variation of locally-infected skin of Epinephelus coioides reveals the mucosal immune mechanism against Cryptocaryon irritans. Fish. Shellfish Immunol. 2017, 66, 398–410. [Google Scholar] [CrossRef]
- Wang, H.; Chen, X.; Li, S.; Zhou, C.; Xu, L.; Wu, Z.; Chen, X. Identification and expression analysis of Langerhans cells marker Langerin/CD207 in grass carp, Ctenopharyngodon idella. Gene 2021, 768, 145315. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Qin, Z.; Liu, H.; Lin, L.; Ye, J.; Li, J. Recent advances on phagocytic B cells in teleost fish. Front. Immunol. 2020, 11, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.; Wilson, M.; Bfuflten, E.; Stuge, T.; Warr, G.; Ciem, W. Functional and molecular characterization of teleost leukocytes. Immunol. Rev. 1998, 166, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Abós, B.; Castro, R.; González Granja, A.; Havixbeck, J.J.; Barreda, D.R.; Tafalla, C. Early activation of teleost B cells in response to rhabdovirus infection. J. Virol. 2015, 89, 1768–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, R.; Abós, B.; González, L.; Granja, A.G.; Tafalla, C. Expansion and differentiation of IgM+ B cells in the rainbow trout peritoneal cavity in response to different antigens. Dev. Comp. Immunol. 2017, 70, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Sunyer, J.O. Evolutionary and functional relationships of B cells from fish and mammals: Insights into their novel roles in phagocytosis and presentation of particulate antigen. Infect. Disord. Drug Targets 2012, 12, 200–212. [Google Scholar] [CrossRef]
- Rougeot, J.; Torraca, V.; Zakrzewska, A.; Kanwal, Z.; Jansen, H.J.; Sommer, F.; Spaink, H.P.; Meijer, A.H. RNAseq profiling of leukocyte populations in zebrafish larvae reveals a cxcl11 chemokine gene as a marker of macrophage polarization during mycobacterial infection. Front. Immunol. 2019, 10, 832. [Google Scholar] [CrossRef]
- Sever, L.; Vo, N.T.K.; Bols, N.C.; Dixon, B. Tapasin’s protein interactions in the rainbow trout peptide-loading complex. Dev. Comp. Immunol. 2018, 81, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Ji, C.; Cao, X.; Fu, Z.; Zhang, S.; Guo, X. Molecular and biological characterization of interferon-γ-inducible-lysosomal thiol reductase gene in zebrafish (Danio rerio). Fish. Shellfish Immunol. 2012, 33, 1133–1138. [Google Scholar] [CrossRef]
- Zheng, W.; Chen, X. Cloning and expression analysis of interferon-γ-inducible-lysosomal thiol reductase gene in large yellow croaker (Pseudosciaena crocea). Mol. Immunol. 2006, 43, 2135–2141. [Google Scholar] [CrossRef]
- Song, J.; Liu, H.; Ma, L.; Gao, C.; Zhang, S. Molecular cloning, expression and functional characterization of interferon-γ-inducible lysosomal thiol reductase (GILT) gene from mandarin fish (Siniperca chuatsi). Fish. Shellfish Immunol. 2014, 38, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Herpin, A.; Lescat, L.; Bobe, J.; Jenny, A.; Seiliez, I. Lighting chaperone-mediated autophagy (CMA) evolution with an ancient LAMP: The existence of a functional CMA activity in fish. Autophagy 2020, 16, 1918–1920. [Google Scholar] [CrossRef] [PubMed]
- Lescat, L.; Véeron, V.; Mourot, B.; Péron, S.; Chenais, N.; Dias, K.; Riera-Heredia, N.; Beaumatin, F.; Pinel, K.; Priault, M.; et al. Chaperone-mediated autophagy in the light of evolution: Insight from fish. Mol. Biol. Evol. 2020, 37, 2887–2899. [Google Scholar] [CrossRef] [PubMed]
- Lescat, L.; Herpin, A.; Mourot, B.; Véron, V.; Guiguen, Y.; Bobe, J.; Seiliez, I. CMA restricted to mammals and birds: Myth or reality? Autophagy 2018, 14, 1267–1270. [Google Scholar] [CrossRef] [PubMed]
- Yabu, T.; Imamura, S.; Mohammed, M.S.; Touhata, K.; Minami, T.; Terayama, M.; Yamashita, M. Differential gene expression of HSC70/HSP70 in yellowtail cells in response to chaperone-mediated autophagy. FEBS J. 2011, 278, 673–685. [Google Scholar] [CrossRef]
- Dubińska-Magiera, M.; Niedbalska-Tarnowska, J.; Migocka-Patrzałek, M.; Posyniak, E.; Daczewska, M. Characterization of Hspb8 in Zebrafish. Cells 2020, 9, 1562. [Google Scholar] [CrossRef]
- Pant, D.C.; Nazarko, T.Y. Selective autophagy: The rise of the zebrafish model. Autophagy 2021, 17, 3297–3305. [Google Scholar] [CrossRef]
- Muñoz-Sánchez, S.; van der Vaart, M.; Meijer, A.H. Autophagy and Lc3-associated phagocytosis in zebrafish models of bacterial infections. Cells 2020, 9, 2372. [Google Scholar] [CrossRef]
- Valionyte, E.; Yang, Y.; Griffiths, S.A.; Bone, A.T.; Barrow, E.R.; Sharma, V.; Lu, B.; Luo, S. The caspase-6–p62 axis modulates p62 droplets based autophagy in a dominant-negative manner. Cell Death Differ. 2021. [Google Scholar] [CrossRef]
- Dong, G.; Zhang, Z.; Duan, K.; Shi, W.; Huang, R.; Wang, B.; Luo, L.; Zhang, Y.; Ruan, H.; Huang, H. Beclin 1 deficiency causes hepatic cell apoptosis via endoplasmic reticulum stress in zebrafish larvae. FEBS Lett. 2020, 594, 1155–1165. [Google Scholar] [CrossRef]
- Mawed, S.A.; He, Y.; Zhang, J.; Mei, J. Strategy of hepatic metabolic defects induced by beclin1 heterozygosity in adult zebrafish. Int. J. Mol. Sci. 2020, 21, 1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masud, S.; Prajsnar, T.K.; Torraca, V.; Lamers, G.E.M.; Benning, M.; Van Der Vaart, M.; Meijer, A.H. Macrophages target Salmonella by Lc3-associated phagocytosis in a systemic infection model. Autophagy 2019, 15, 796–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prajsnar, T.K.; Serba, J.J.; Dekker, B.M.; Gibson, J.F.; Masud, S.; Fleming, A.; Johnston, S.A.; Renshaw, S.A.; Meijer, A.H. The autophagic response to Staphylococcus aureus provides an intracellular niche in neutrophils. Autophagy 2021, 17, 888–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, J.F.; Prajsnar, T.K.; Hill, C.J.; Tooke, A.K.; Serba, J.J.; Tonge, R.D.; Foster, S.J.; Grierson, A.J.; Ingham, P.W.; Renshaw, S.A.; et al. Neutrophils use selective autophagy receptor Sqstm1/p62 to target Staphylococcus aureus for degradation in vivo in zebrafish. Autophagy 2021, 17, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Li, W.; Guo, J.; Jia, P.; Zhang, W.; Yi, M.; Jia, K. N Protein of viral hemorrhagic septicemia virus suppresses STAT1-mediated MHC class II transcription to impair antigen presentation in sea perch, Lateolabrax japonicus. J. Immunol. 2022, 208, 1076–1084. [Google Scholar] [CrossRef]
- Abram, Q.H.; Vo, N.T.K.; Kellendonk, C.; Bols, N.C.; Katzenback, B.A.; Dixon, B. Regulation of endogenous antigen presentation in response to suboptimal temperatures in a walleye skin fibroblast cell line. Fish Shellfish Immunol. 2020, 98, 788–799. [Google Scholar] [CrossRef]
- Abram, Q.H.; Rodríguez-Ramos, T.; Bols, N.C.; Katzenback, B.A.; Dixon, B. Effect of suboptimal temperature on the regulation of endogenous antigen presentation in a rainbow trout hypodermal fibroblast cell line. Dev. Comp. Immunol. 2019, 100, 103423. [Google Scholar] [CrossRef]
- Nombela, I.; Requena-platek, R.; Morales-lange, B.; Chico, V.; Puente-Marín, S.; Ciordia, S.; Mena, M.C.; Coll, J.; Perez, L.; Mercado, L.; et al. Rainbow trout red blood cells exposed to viral hemorrhagic septicemia virus up-regulate antigen-processing mechanisms and MHC I&II, CD86, and CD83 antigen-presenting cell markers. Cells 2019, 8, 386. [Google Scholar] [CrossRef] [Green Version]
- Nombela, I.; López-Lorigados, M.; Salvador-Mira, M.E.; Puente-Marin, S.; Chico, V.; Ciordia, S.; Mena, M.C.; Mercado, L.; Coll, J.; Perez, L.; et al. Integrated transcriptomic and proteomic analysis of red blood cells from rainbow trout challenged with VHSV point towards novel immunomodulant targets. Vaccines 2019, 7, 63. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.J.; Moon, J.-Y.; Nam, B.-H.; Kim, Y.-O.; Kim, W.-J.; Lee, J.-H.; Kim, K.-K.; Kim, B.-S.; Yeo, S.-Y.; Lee, C.H.; et al. Molecular characterization of the autophagy-related gene Beclin-1 from the olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2011, 31, 189–195. [Google Scholar] [CrossRef]
- Peruzza, L.; Pascoli, F.; Dalla Rovere, G.; Franch, R.; Ferraresso, S.; Babbucci, M.; Biasini, L.; Abbadi, M.; Panzarin, V.; Toffan, A.; et al. Transcriptome analysis reveals a complex response to the RGNNV/SJNNV reassortant Nervous Necrosis Virus strain in sea bream larvae. Fish Shellfish Immunol. 2021, 114, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.; Wan, Q.; Su, H.; Xiao, X.; Liao, Z.; Ji, J.; Yang, C.; Lin, L.; Su, J. ROS-induced HSP70 promotes cytoplasmic translocation of high-mobility group box 1b and stimulates antiviral autophagy in grass carp kidney cells. J. Biol. Chem. 2018, 293, 17387–17401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, J.; La Patra, S. Induction of the rainbow trout MHC class I pathway during acute IHNV infection. Immunogenetics 2002, 54, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Svenning, S.; Gondek-Wyrozemska, A.T.; van der Wal, Y.A.; Robertsen, B.; Jensen, I.; Jørgensen, J.B.; Edholm, E.-S. Microbial danger signals control transcriptional induction of distinct MHC class I L lineage genes in Atlantic salmon. Front. Immunol. 2019, 10, 2425–2442. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Du, H.; Huang, H.; Xian, J.; Xia, Z.; Hu, Y. Major histocompatibility complex class I (MHC Iα) of Japanese flounder (Paralichthys olivaceus) plays a critical role in defense against intracellular pathogen infection. Fish Shellfish Immunol. 2019, 94, 122–131. [Google Scholar] [CrossRef]
- Ahn, D.-H.; Kang, S.; Park, H. Transcriptome analysis of immune response genes induced by pathogen agonists in the Antarctic bullhead notothen Notothenia coriiceps. Fish Shellfish Immunol. 2016, 55, 315–322. [Google Scholar] [CrossRef]
- Bello-Pérez, M.; Pereiro, P.; Coll, J.; Novoa, B.; Pérez, L.; Falco, A. Zebrafish C-reactive protein isoforms inhibit SVCV replication by blocking autophagy through interactions with cell membrane cholesterol. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Pereiro, P.; Figueras, A.; Novoa, B. Zebrafish pten genes play relevant but distinct roles in antiviral immunity. Vaccines 2020, 8, 199. [Google Scholar] [CrossRef]
- Peatman, E.; Terhune, J.; Baoprasertkul, P.; Xu, P.; Nandi, S.; Wang, S.; Somridhivej, B.; Kucuktas, H.; Li, P.; Dunham, R.; et al. Microarray analysis of gene expression in the blue catfish liver reveals early activation of the MHC class I pathway after infection with Edwardsiella ictaluri. Mol. Immunol. 2008, 45, 553–566. [Google Scholar] [CrossRef]
- Li, K.; Wei, X.; Zhang, L.; Chi, H.; Yang, J. Raptor/mTORC1 acts as a modulatory center to regulate anti-bacterial immune response in rockfish. Front. Immunol. 2019, 10, 2953. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.; Hummel, K.; Noebauer, K.; Welch, T.J.; Razzazi-Fazeli, E.; El-Matbouli, M. Proteome analysis reveals a role of rainbow trout lymphoid organs during Yersinia ruckeri infection process. Sci. Rep. 2018, 8, 13998. [Google Scholar] [CrossRef] [PubMed]
- Lü, A.; Hu, X.; Xue, J.; Zhu, J.; Wang, Y.; Zhou, G. Gene expression profiling in the skin of zebrafish infected with Citrobacter freundii. Fish Shellfish Immunol. 2012, 32, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.M.; Cao, L.; Hu, Y.W.; Chang, M.X. Transcriptomic characterization of adult zebrafish infected with Streptococcus agalactiae. Fish Shellfish Immunol. 2019, 94, 355–372. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, T.; Gao, S.; Xu, G.; Niu, H.; Huang, R.; Wu, S. Salmonella plasmid virulence gene spvB enhances bacterial virulence by inhibiting autophagy in a zebrafish infection model. Fish Shellfish Immunol. 2016, 49, 252–259. [Google Scholar] [CrossRef]
- Young, N.D.; Cooper, G.A.; Nowak, B.F.; Koop, B.F.; Morrison, R.N. Coordinated down-regulation of the antigen processing machinery in the gills of amoebic gill disease-affected Atlantic salmon (Salmo salar L.). Mol. Immunol. 2008, 45, 2581–2597. [Google Scholar] [CrossRef]
- Tadiso, T.M.; Krasnov, A.; Skugor, S.; Afanasyev, S.; Hordvik, I.; Nilsen, F. Gene expression analyses of immune responses in Atlantic salmon during early stages of infection by salmon louse (Lepeophtheirus salmonis) revealed bi-phasic responses coinciding with the copepod-chalimus transition. BMC Genom. 2011, 12, 141. [Google Scholar] [CrossRef] [Green Version]
- Konczal, M.; Ellison, A.R.; Phillips, K.P.; Radwan, J.; Mohammed, R.S.; Cable, J.; Chadzinska, M. RNA-Seq analysis of the guppy immune response against Gyrodactylus bullatarudis infection. Parasite Immunol. 2020, 42, e12782. [Google Scholar] [CrossRef]
- Cervera, L.; González-Fernández, C.; Arizcun, M.; Cuesta, A.; Chaves-Pozo, E. Severe natural outbreak of Cryptocaryon irritans in gilthead seabream produces leukocyte mobilization and innate immunity at the gill tissue. Int. J. Mol. Sci. 2022, 23, 937. [Google Scholar] [CrossRef]
- Sun, Z.; Shen, B.; Wu, H.; Zhou, X.; Wang, Q.; Xiao, J.; Zhang, Y. The secreted fructose 1,6-bisphosphate aldolase as a broad spectrum vaccine candidate against pathogenic bacteria in aquaculture. Fish Shellfish Immunol. 2015, 46, 638–647. [Google Scholar] [CrossRef]
- Darawiroj, D.; Kondo, H.; Hirono, I.; Aoki, T. Immune-related gene expression profiling of yellowtail (Seriola quinqueradiata) kidney cells stimulated with ConA and LPS using microarray analysis. Fish Shellfish Immunol. 2008, 24, 260–266. [Google Scholar] [CrossRef]
- Mostowy, S.; Boucontet, L.; Mazón-Moya, M.J.; Sirianni, A.; Boudinot, P.; Hollinshead, M.; Cossart, P.; Herbomel, P.; Levraud, J.P.; Colucci-Guyon, E. The zebrafish as a new model for the in vivo study of Shigella flexneri interaction with phagocytes and bacterial autophagy. PLoS Pathog. 2013, 9, e1003588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijer, A.H.; van der Vaart, M. DRAM1 promotes the targeting of mycobacteria to selective autophagy. Autophagy 2014, 10, 2389–2391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Vaart, M.; Korbee, C.J.; Lamers, G.E.M.; Tengeler, A.C.; Hosseini, R.; Haks, M.C.; Ottenhoff, T.H.M.; Spaink, H.P.; Meijer, A.H. The DNA damage-regulated autophagy modulator DRAM1 links mycobacterial recognition via TLP-MYD88 to authophagic defense. Cell Host Microbe 2014, 15, 753–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockhammer, O.W.; Rauwerda, H.; Wittink, F.R.; Breit, T.M.; Meijer, A.H.; Spaink, H.P. Transcriptome analysis of Traf6 function in the innate immune response of zebrafish embryos. Mol. Immunol. 2010, 48, 179–190. [Google Scholar] [CrossRef]
- Yang, D.; Liu, Q.; Yang, M.; Wu, H.; Wang, Q.; Xiao, J.; Zhang, Y. RNA-seq liver transcriptome analysis reveals an activated MHC-I pathway and an inhibited MHC-II pathway at the early stage of vaccine immunization in zebrafish. BMC Genom. 2012, 13, 319. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Liu, Q.; Ni, C.; Li, S.; Wu, H.; Wang, Q.; Xiao, J.; Zhang, Y. Gene expression profiling in live attenuated Edwardsiella tarda vaccine immunized and challenged zebrafish: Insights into the basic mechanisms of protection seen in immunized fish. Dev. Comp. Immunol. 2013, 40, 132–141. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, H.; Wang, Q.; Qu, J.; Liu, Q.; Xiao, J.; Zhang, Y. A live attenuated combination vaccine evokes effective immune-mediated protection against Edwardsiella tarda and Vibrio anguillarum. Vaccine 2014, 32, 5937–5944. [Google Scholar] [CrossRef]
- Zhang, X.; Mu, Y.; Mu, P.; Ao, J.; Chen, X. Transcriptome analysis reveals comprehensive insights into the early immune response of large yellow croaker (Larimichthys crocea) induced by trivalent bacterial vaccine. PLoS ONE 2017, 12, e0170958. [Google Scholar] [CrossRef] [Green Version]
- García-Valtanen, P.; Del Mar Ortega-Villaizan, M.; Martínez-López, A.; Medina-Gali, R.; Pérez, L.; Mackenzie, S.; Figueras, A.; Coll, J.M.; Estepa, A. Autophagy-inducing peptides from mammalian VSV and fish VHSV rhabdoviral G glycoproteins (G) as models for the development of new therapeutic molecules. Autophagy 2014, 10, 1666–1680. [Google Scholar] [CrossRef] [Green Version]
- Espín-Palazón, R.; Martínez-López, A.; Roca, F.J.; López-Muñoz, A.; Tyrkalska, S.D.; Candel, S.; García-Moreno, D.; Falco, A.; Meseguer, J.; Estepa, A.; et al. TNFα impairs rhabdoviral clearance by inhibiting the host autophagic antiviral response. PLoS Pathog. 2016, 12, e1005699. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnstone, C.; Chaves-Pozo, E. Antigen Presentation and Autophagy in Teleost Adaptive Immunity. Int. J. Mol. Sci. 2022, 23, 4899. https://doi.org/10.3390/ijms23094899
Johnstone C, Chaves-Pozo E. Antigen Presentation and Autophagy in Teleost Adaptive Immunity. International Journal of Molecular Sciences. 2022; 23(9):4899. https://doi.org/10.3390/ijms23094899
Chicago/Turabian StyleJohnstone, Carolina, and Elena Chaves-Pozo. 2022. "Antigen Presentation and Autophagy in Teleost Adaptive Immunity" International Journal of Molecular Sciences 23, no. 9: 4899. https://doi.org/10.3390/ijms23094899
APA StyleJohnstone, C., & Chaves-Pozo, E. (2022). Antigen Presentation and Autophagy in Teleost Adaptive Immunity. International Journal of Molecular Sciences, 23(9), 4899. https://doi.org/10.3390/ijms23094899