
The Ameliorative Effect of Dexamethasone on the Development of Autoimmune Lung Injury and Mediastinal Fat-Associated Lymphoid Clusters in an Autoimmune Disease Mouse Model

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Analysis of the Impact of Dexamethasone on the Degree of Lung Injury and the Development of MFALCs and the Systemic Autoimmunity
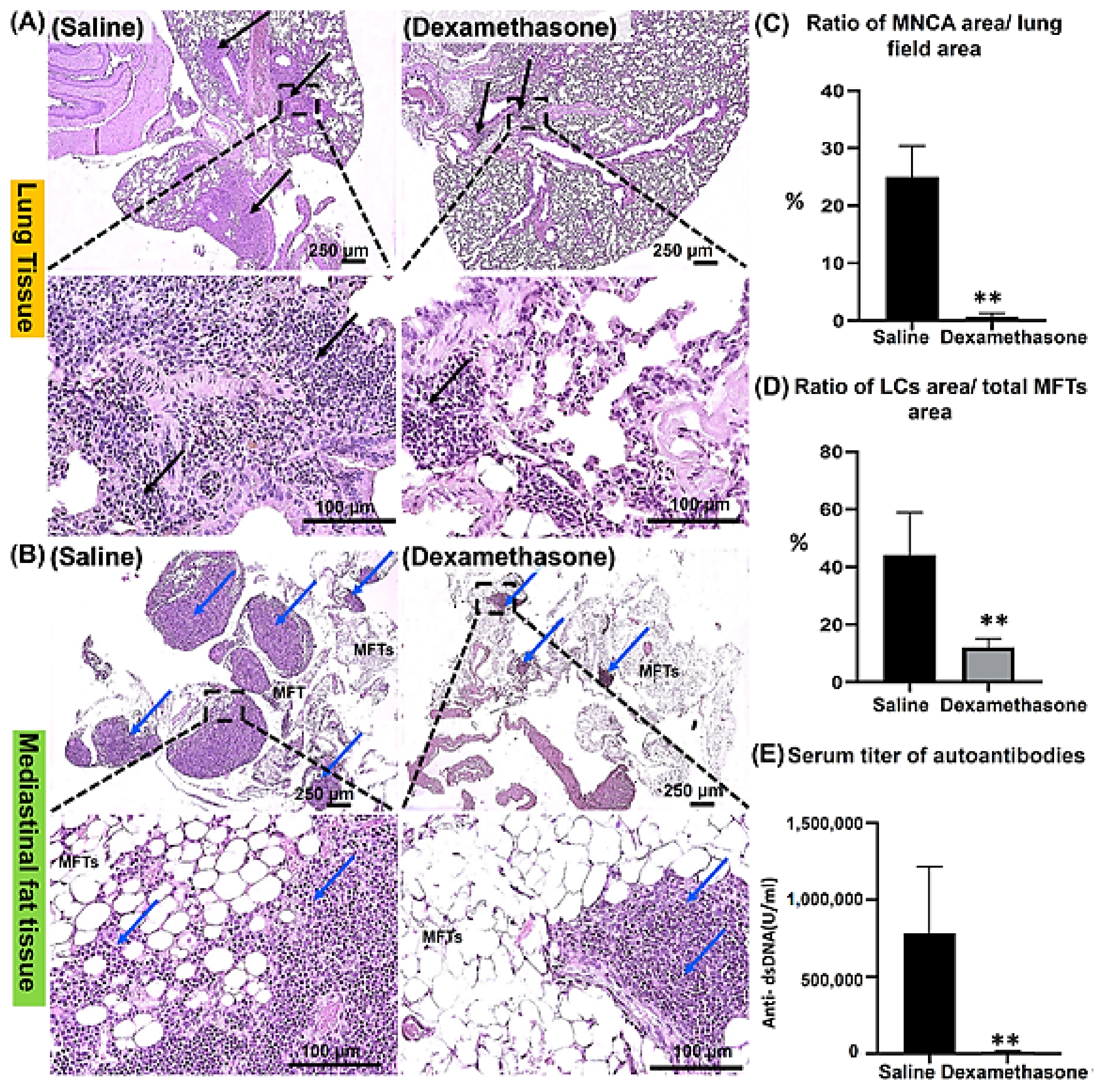
2.1.1. Histopathological Features of the Lungs and MFALCs in the Saline and Dexamethasone Groups
2.1.2. The Degree of Systemic Autoimmunity in the Saline and Dexamethasone Groups
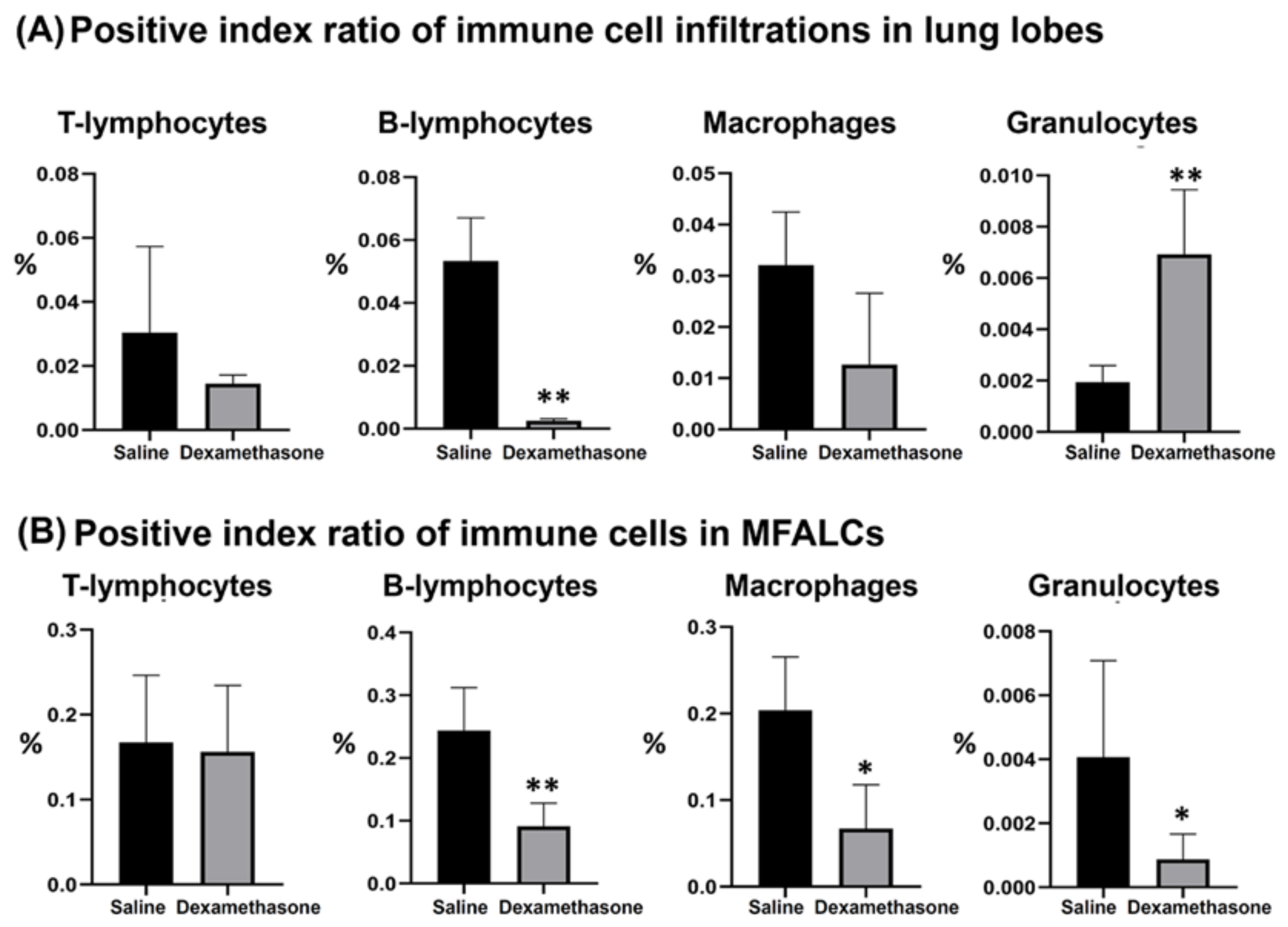
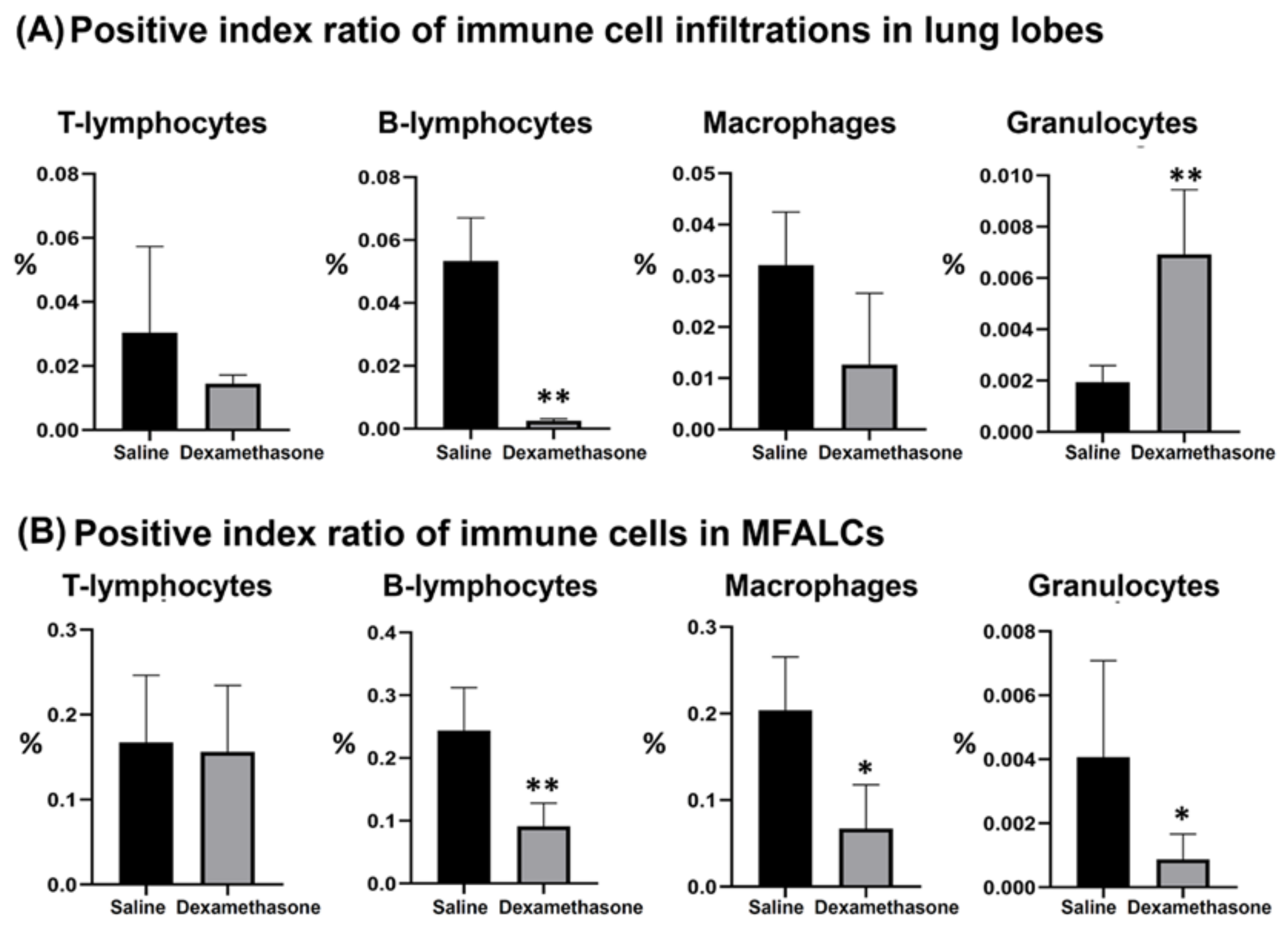
2.2. Immune Cells in the Lung and MFALCs of Saline and Dexamethasone Groups
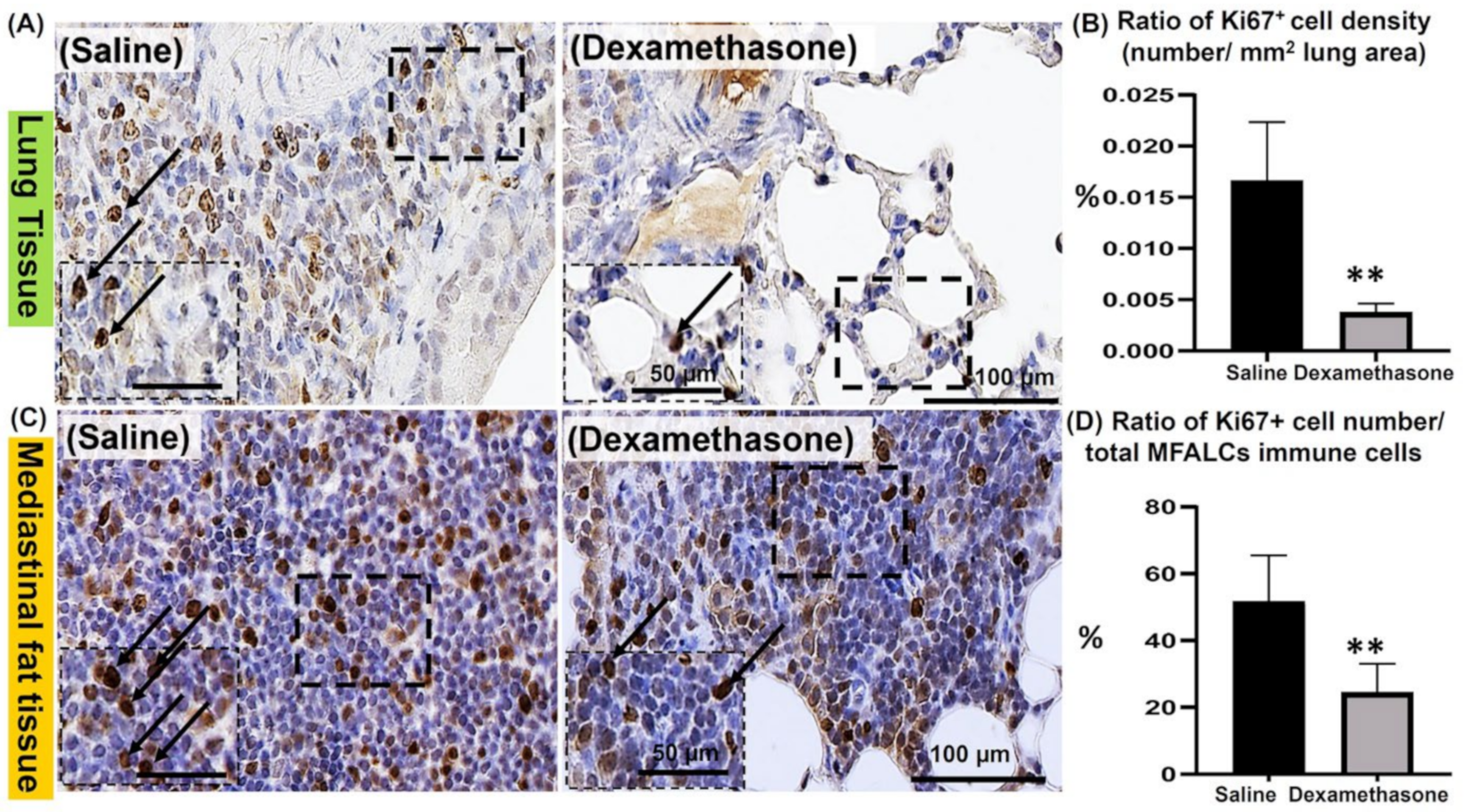
2.3. Analysis of the Degree of Proliferation of Immune Cells in the Lungs and MFALCs between the Saline and Dexamethasone Groups
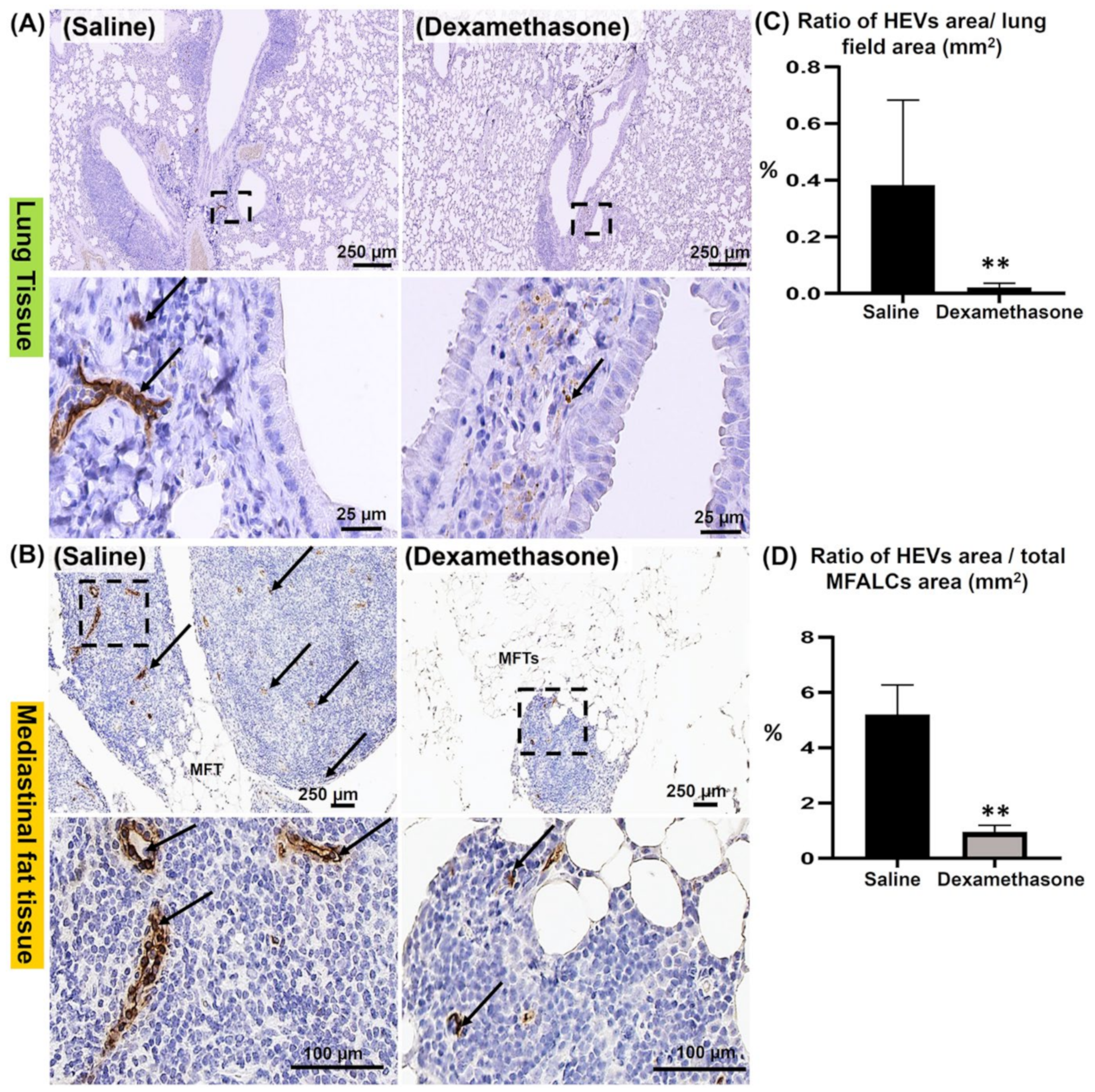
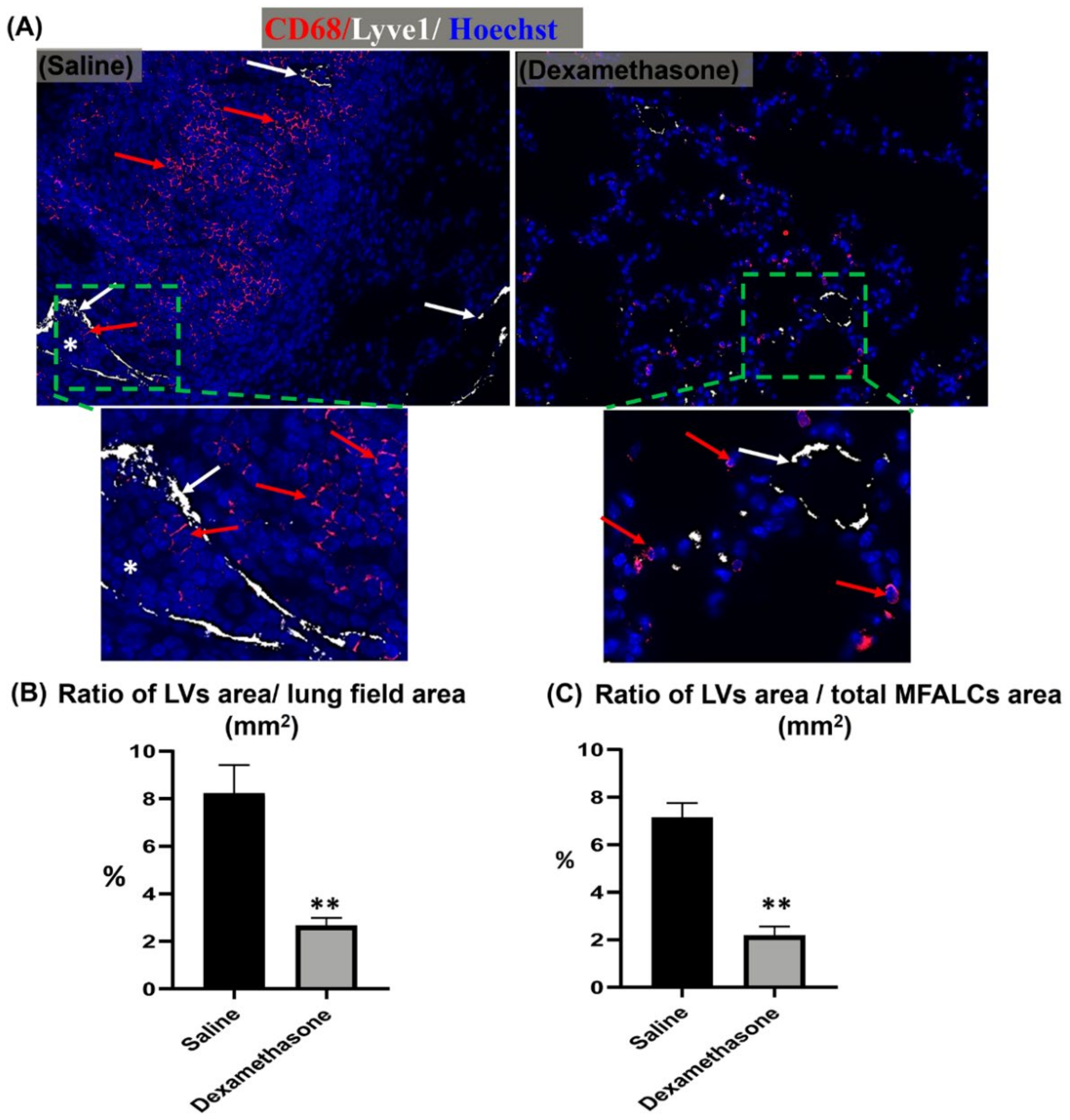
2.4. Comparison of HEVs and the Development of Lymphatic Vessels (LVs) in the Lung and MFALCs between the Saline and Dexamethasone Groups
2.5. Analysis of the Impact of Dexamethasone on Chemokine Expression in the Lungs between the Saline and Dexamethasone Groups
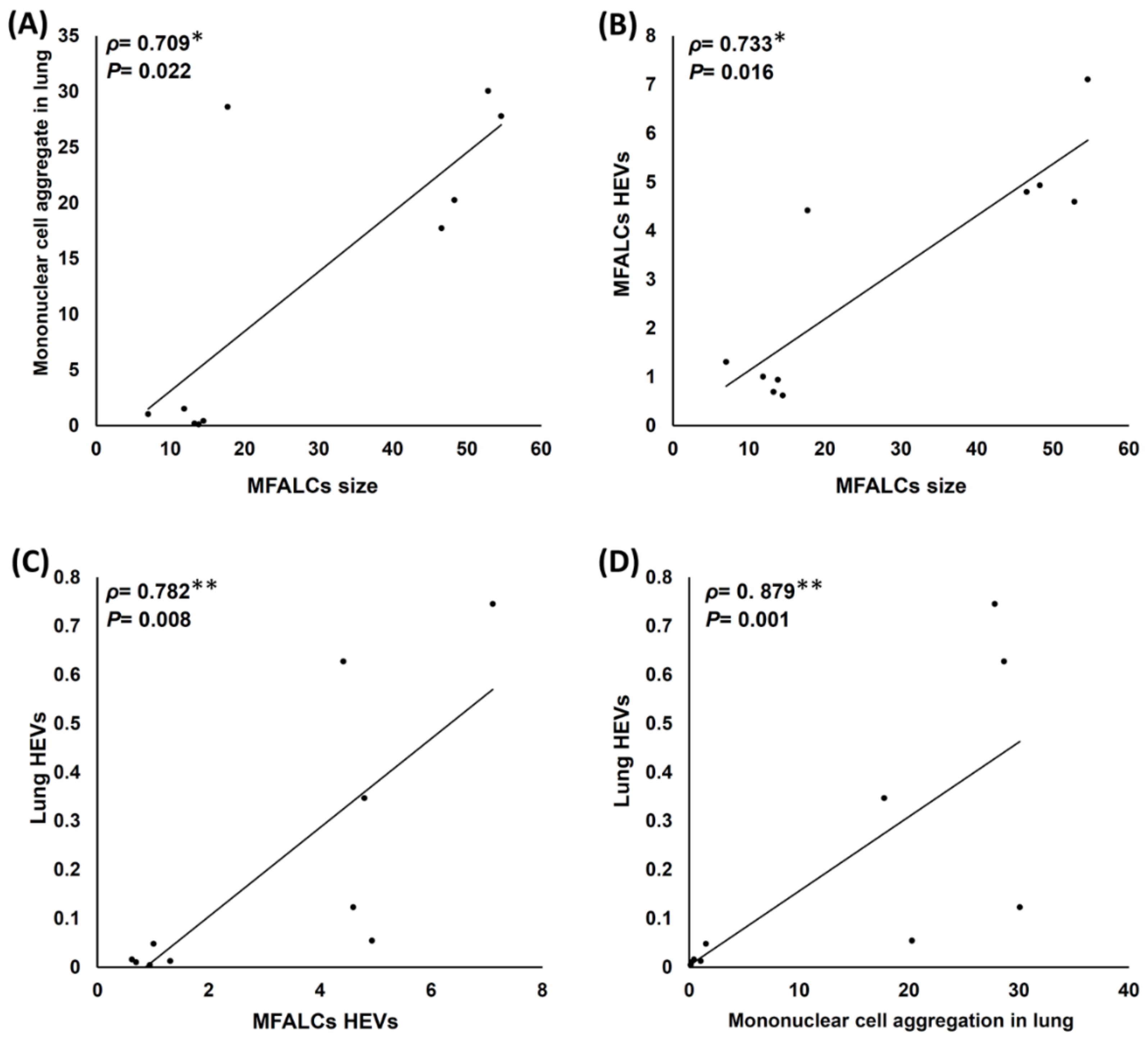
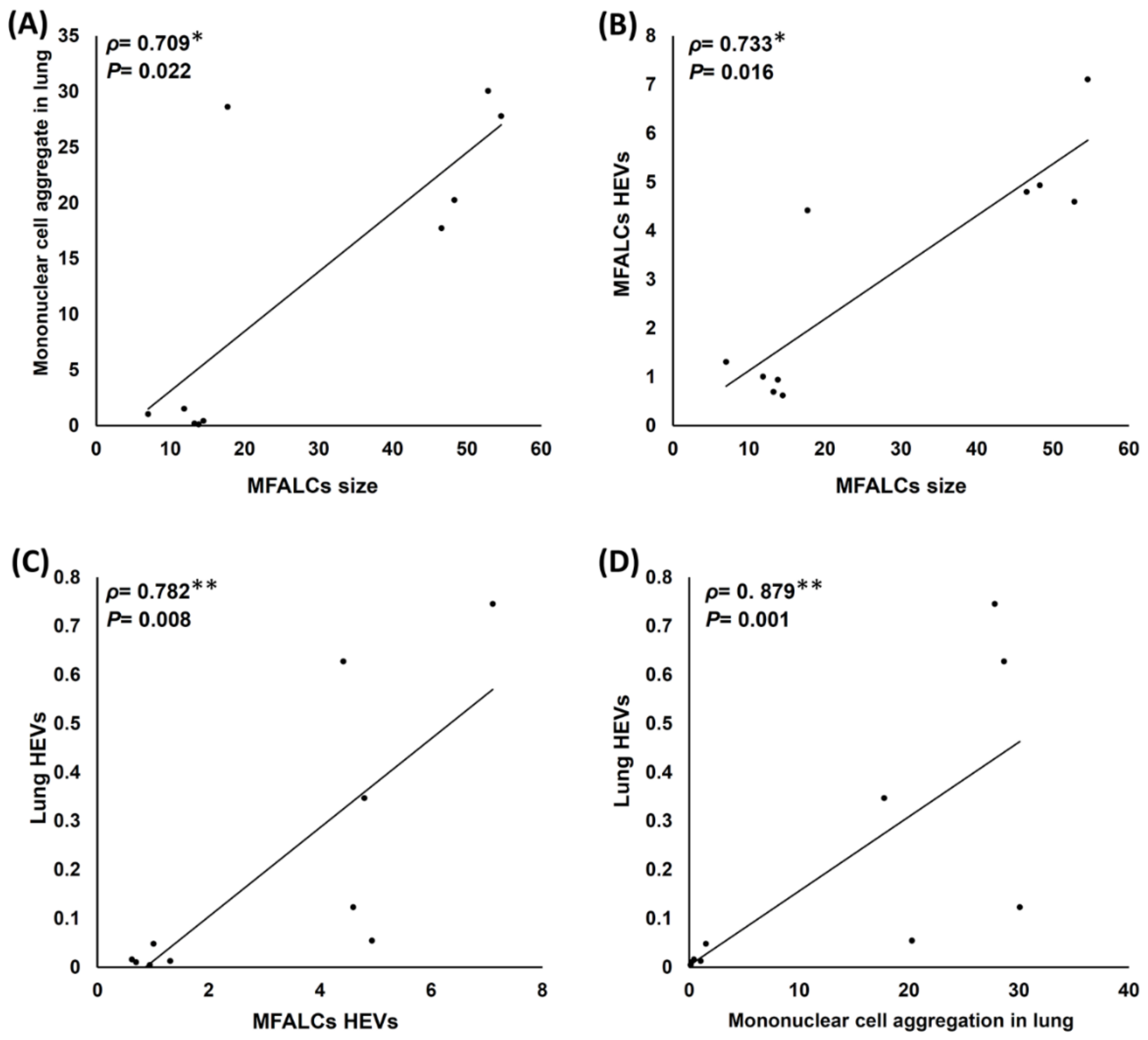
2.6. Analysis of Histopathological Correlations among Different Parameters of the Lungs and MFALCs in Both the Saline and Dexamethasone Groups
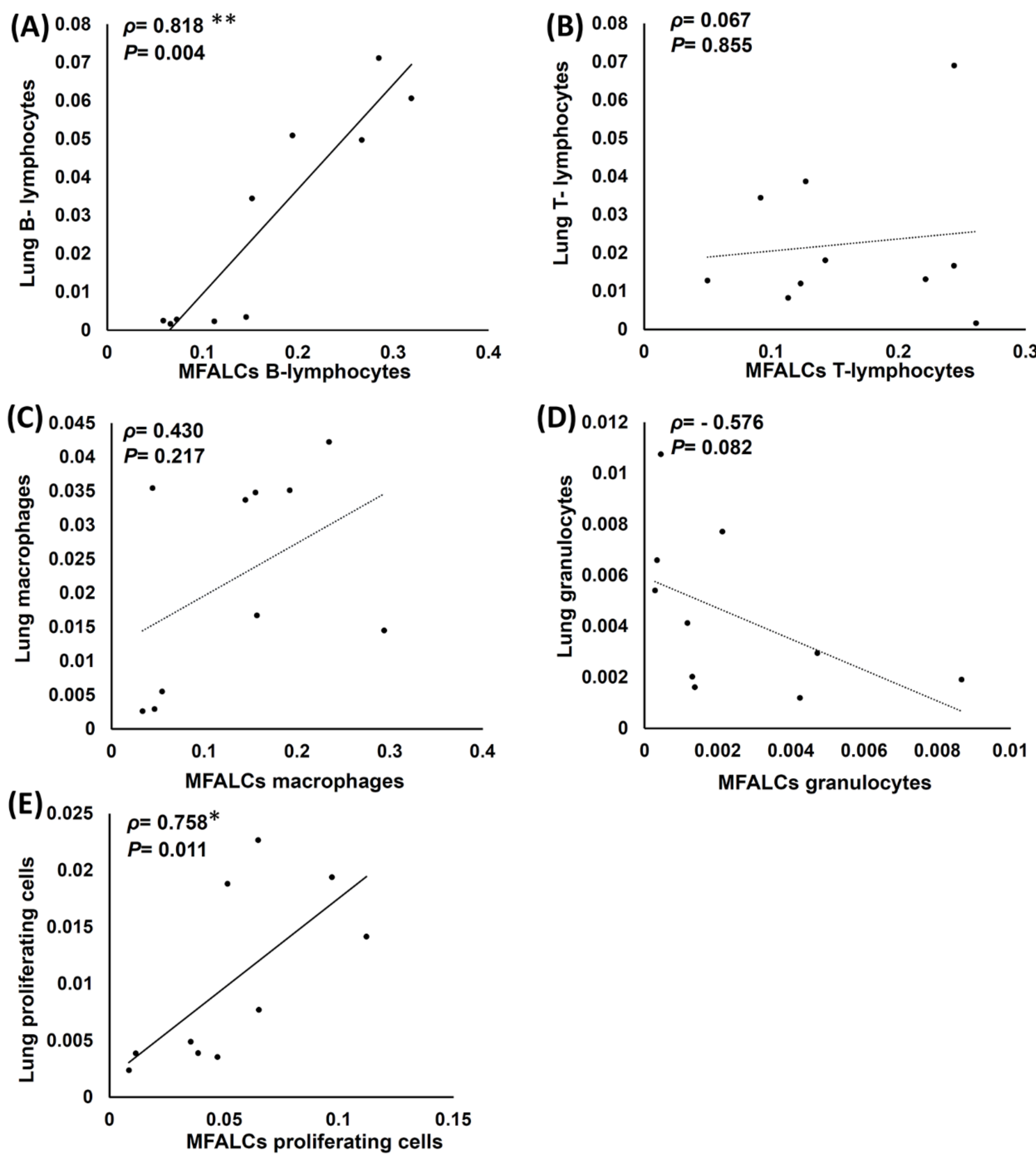
2.7. Analysis of Histopathological Correlations between Immune Cell Populations and the Proliferating of Cells in the Lungs and MFALCs of Both Saline and Dexamethasone Group Mice
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Ethical Statement
4.2. Experimental Design and Sample Preparation
4.3. Serum Autoantibody Measurement
4.4. Tissue Preparation for Histopathological and Immunohistochemical Analysis
4.5. Morphometrical Measurements
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Angum, F.; Khan, T.; Kaler, J.; Siddiqui, L.; Hussain, A. The Prevalence of Autoimmune Disorders in Women: A Narrative Review. Cureus 2020, 12, e8094. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.D., Jr. Rheumatoid arthritis. Pathophysiology and implications for therapy. N. Engl. J. Med. 1990, 322, 1277–1289. [Google Scholar] [PubMed]
- Baumgart, D.C.; Carding, S.R. Inflammatory bowel disease: Cause and immunobiology. Lancet 2007, 369, 1627–1640. [Google Scholar] [CrossRef]
- Wenzel, J.; Schmidt, R.; Proelss, J.; Zahn, S.; Bieber, T.; Tüting, T. Type I interferon-associated skin recruitment of CXCR3+ lymphocytes in dermatomyositis. Clin. Exp. Dermatol. Exp. Dermatol. 2006, 31, 576–582. [Google Scholar] [CrossRef]
- Askanase, A.; Shum, K.; Mitnick, H. Systemic Lupus Erythematosus: An Overview. Soc. Work Health Care 2012, 51, 576–586. [Google Scholar] [CrossRef]
- Jindal, S.K.; Agarwal, R. Autoimmunity and interstitial lung disease. Curr. Opin. Pulm. Med. 2005, 11, 438–446. [Google Scholar] [CrossRef]
- Fairweather, D.; Frisancho-Kiss, S.; Rose, N.R. Sex differences in autoimmune disease from a pathological perspective. Am. J. Pathol. 2008, 173, 600–609. [Google Scholar] [CrossRef] [Green Version]
- Gershwin, L.J. Autoimmune diseases in small animals. Vet. Clin. Small Anim. Pract. 2010, 40, 439–457. [Google Scholar] [CrossRef]
- Reilly, C.M.; Gilkeson, G.S. Use of genetic knockouts to modulate disease expression in a murine model of lupus, MRL/lpr mice. Immunol. Res. 2002, 25, 143–153. [Google Scholar] [CrossRef]
- Perry, D.; Sang, A.; Yin, Y.; Zheng, Y.-Y.; Morel, L. Murine models of systemic lupus erythematosus. J. Biomed. Biotechnol. 2011, 2011, 271694. [Google Scholar] [CrossRef] [Green Version]
- Elewa, Y.H.; Ichii, O.; Kon, Y. Comparative analysis of mediastinal fat-associated lymphoid cluster development and lung cellular infiltration in murine autoimmune disease models and the corresponding normal control strains. Immunology 2016, 147, 30–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, M.; Zemans, R.L.; Jeyaseelan, S. Role of chemokines in the pathogenesis of acute lung injury. Am. J. Respir. Cell Mol. Biol. 2012, 46, 566–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimatcheva, E.; Pandina, T.; Reilly, C.; Torno, S.; Bussler, H.; Scrivens, M.; Jonason, A.; Mallow, C.; Doherty, M.; Paris, M.; et al. CXCL13 antibody for the treatment of autoimmune disorders. BMC Immunol. 2015, 16, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kameda, M.; Otsuka, M.; Chiba, H.; Kuronuma, K.; Hasegawa, T.; Takahashi, H.; Takahashi, H. CXCL9, CXCL10, and CXCL11; biomarkers of pulmonary inflammation associated with autoimmunity in patients with collagen vascular diseases-associated interstitial lung disease and interstitial pneumonia with autoimmune features. PLoS ONE 2020, 15, e0241719. [Google Scholar] [CrossRef] [PubMed]
- Claeys, E.; Vermeire, K. Immunosuppressive drugs in organ transplantation to prevent allograft rejection: Mode of action and side effects. J. Immunol. Sci. 2019, 3, 14–21. [Google Scholar] [CrossRef]
- Horby, P.; Lim, W.S.; Emberson, J.; Mafham, M.; Bell, J.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; Elmahi, E.; et al. Effect of Dexamethasone in Hospitalized Patients with COVID-19—Preliminary Report. medRxiv 2020. [Google Scholar] [CrossRef]
- Masum, M.A.; Ichii, O.; Elewa, Y.H.A.; Otani, Y.; Namba, T.; Kon, Y. Vasculature-Associated Lymphoid Tissue: A Unique Tertiary Lymphoid Tissue Correlates with Renal Lesions in Lupus Nephritis Mouse Model. Front. Immunol. 2020, 11, 3211. [Google Scholar] [CrossRef]
- Elewa, Y.H.A.; Ichii, O.; Kon, Y. Sex-related differences in autoimmune-induced lung lesions in MRL/MpJ-faslpr mice are mediated by the development of mediastinal fat-associated lymphoid clusters. Autoimmunity 2017, 50, 306–316. [Google Scholar] [CrossRef]
- Elewa, Y.H.A.; Ichii, O.; Takada, K.; Nakamura, T.; Masum, M.A.; Kon, Y. Histopathological Correlations between Mediastinal Fat-Associated Lymphoid Clusters and the Development of Lung Inflammation and Fibrosis following Bleomycin Administration in Mice. Front. Immunol. 2018, 9, 271. [Google Scholar] [CrossRef] [Green Version]
- Elewa, Y.H.A.; Ichii, O.; Nakamura, T.; Kon, Y. Pathological Alternations of Mediastinal Fat-Associated Lymphoid Cluster and Lung in a Streptozotocin-Induced Diabetic Mouse Model. Microsc. Microanal. 2021, 27, 187–200. [Google Scholar] [CrossRef]
- Tokunaga, R.; Zhang, W.; Naseem, M.; Puccini, A.; Berger, M.D.; Soni, S.; McSkane, M.; Baba, H.; Lenz, H.-J. CXCL9, CXCL10, CXCL11/CXCR3 axis for immune activation—A target for novel cancer therapy. Cancer Treat. Rev. 2018, 63, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, E.; Sa, Q.; Brogli, M.; Kudo, T.; Wang, X.; Dubey, J.P.; Suzuki, Y. CXCL9 is important for recruiting immune T cells into the brain and inducing an accumulation of the T cells to the areas of tachyzoite proliferation to prevent reactivation of chronic cerebral infection with Toxoplasma gondii. Am. J. Pathol. 2015, 185, 314–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legler, D.F.; Loetscher, M.; Roos, R.S.; Clark-Lewis, I.; Baggiolini, M.; Moser, B. B cell-attracting chemokine 1, a human CXC chemokine expressed in lymphoid tissues, selectively attracts B lymphocytes via BLR1/CXCR5. J. Exp. Med. 1998, 187, 655–660. [Google Scholar] [CrossRef]
- Zhu, D.; Ye, W.; Jiang, J. Clinical significance of CXCL13/CXCR5 axis in human cancers. Transl. Cancer Res. 2018, 7, 1737–1742. [Google Scholar] [CrossRef]
- Mokra, D.; Mikolka, P.; Kosutova, P.; Mokry, J. Corticosteroids in Acute Lung Injury: The Dilemma Continues. Int. J. Mol. Sci. 2019, 20, 4765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.J.; Yong, S.H.; Leem, A.Y.; Lee, S.H.; Kim, S.Y.; Lee, S.H.; Kim, E.Y.; Chung, K.S.; Jung, J.Y.; Kang, Y.A.; et al. Corticosteroid responsiveness in patients with acute exacerbation of interstitial lung disease admitted to the emergency department. Sci. Rep. 2021, 11, 5762. [Google Scholar] [CrossRef]
- Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; Elmahi, E.; et al. Dexamethasone in Hospitalized Patients with Covid-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar]
- Villar, J.; Añón, J.M.; Ferrando, C.; Aguilar, G.; Muñoz, T.; Ferreres, J.; Ambrós, A.; Aldecoa, C.; Suárez-Sipmann, F.; Thorpe, K.E.; et al. Efficacy of dexamethasone treatment for patients with the acute respiratory distress syndrome caused by COVID-19: Study protocol for a randomized controlled superiority trial. Trials 2020, 21, 717. [Google Scholar] [CrossRef]
- Ruddle, N.H. High Endothelial Venules and Lymphatic Vessels in Tertiary Lymphoid Organs: Characteristics, Functions, and Regulation. Front. Immunol. 2016, 7, 491. [Google Scholar] [CrossRef] [Green Version]
- Kazanietz, M.G.; Durando, M.; Cooke, M. CXCL13 and Its Receptor CXCR5 in Cancer: Inflammation, Immune Response, and Beyond. Front. Endocrinol. 2019, 10, 471. [Google Scholar] [CrossRef] [Green Version]
- Jenh, C.-H.; Cox, M.A.; Hipkin, W.; Lu, T.; Pugliese-Sivo, C.; Gonsiorek, W.; Chou, C.-C.; Narula, S.K.; Zavodny, P.J. Human B Cell-Attracting Chemokine 1 (Bca-1; Cxcl13) Is an Agonist for the Human Cxcr3 Receptor. Cytokine 2001, 15, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ye, X.; Pitmon, E.; Lu, M.; Wan, J.; Jellison, E.R.; Adler, A.J.; Vella, A.T.; Wang, K. IL-17 inhibits CXCL9/10-mediated recruitment of CD8+ cytotoxic T cells and regulatory T cells to colorectal tumors. J. Immunother. Cancer 2019, 7, 324. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tang, J.; Shuai, W.; Meng, J.; Feng, J.; Han, Z. Macrophage polarization and its role in the pathogenesis of acute lung injury/acute respiratory distress syndrome. Inflamm. Res. 2020, 69, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K.; Chittezhath, M.; Shalova, I.N.; Lim, J.Y. Macrophage polarization and plasticity in health and disease. Immunol. Res. 2012, 53, 11–24. [Google Scholar] [CrossRef]
- Aggarwal, N.R.; King, L.S.; D’Alessio, F.R. Diverse macrophage populations mediate acute lung inflammation and resolution. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L709–L725. [Google Scholar] [CrossRef]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Grommes, J.; Soehnlein, O. Contribution of neutrophils to acute lung injury. Mol. Med. 2011, 17, 293–307. [Google Scholar] [CrossRef]
- Tavares, L.P.; Peh, H.Y.; Tan, W.S.D.; Pahima, H.; Maffia, P.; Tiligada, E.; Levi-Schaffer, F. Granulocyte-targeted therapies for airway diseases. Pharmacol. Res. 2020, 157, 104881. [Google Scholar] [CrossRef]
- Serhan, C.N. Resolution phase of inflammation: Novel endogenous anti-inflammatory and proresolving lipid mediators and pathways. Annu. Rev. Immunol. 2007, 25, 101–137. [Google Scholar] [CrossRef] [Green Version]
- Wofsy, D.; Ledbetter, J.A.; Roubinian, J.R.; Seaman, W.E.; Talal, N. Thymic influences on autoimmunity in MRL-lpr mice. Scand. J. Immunol. 1982, 16, 51–58. [Google Scholar] [CrossRef]
- Yamauchi, K.; Kasuya, Y.; Kuroda, F.; Tanaka, K.; Tsuyusaki, J.; Ishizaki, S.; Matsunaga, H.; Iwamura, C.; Nakayama, T.; Tatsumi, K. Attenuation of lung inflammation and fibrosis in CD69-deficient mice after intratracheal bleomycin. Respir. Res. 2011, 12, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horkowitz, A.P.; Schwartz, A.V.; Alvarez, C.A.; Herrera, E.B.; Thoman, M.L.; Chatfield, D.A.; Osborn, K.G.; Feuer, R.; George, U.Z.; Phillips, J.A. Acetylcholine Regulates Pulmonary Pathology During Viral Infection and Recovery. ImmunoTargets Ther. 2020, 9, 333–350. [Google Scholar] [CrossRef] [PubMed]
- Pierezan, F.; Mansell, J.; Ambrus, A.; Rodrigues Hoffmann, A. Immunohistochemical expression of ionized calcium binding adapter molecule 1 in cutaneous histiocytic proliferative, neoplastic and inflammatory disorders of dogs and cats. J. Comp. Pathol. 2014, 151, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Rakaee, M.; Busund, L.-T.R.; Jamaly, S.; Paulsen, E.-E.; Richardsen, E.; Andersen, S.; Al-Saad, S.; Bremnes, R.M.; Donnem, T.; Kilvaer, T.K. Prognostic Value of Macrophage Phenotypes in Resectable Non-Small Cell Lung Cancer Assessed by Multiplex Immunohistochemistry. Neoplasia 2019, 21, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Ichii, O.; Sunden, Y.; Elewa, Y.H.A.; Yoshiyasu, T.; Hattori, H.; Tatsumi, O.; Kon, Y.; Nagasaki, K.-I. Slc:Wistar/ST rats develop unilateral thyroid dysgenesis: A novel animal model of thyroid hemiagenesis. PLoS ONE 2019, 14, e0221939. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Source | Dilution | Antigen Retrieval | Heating Condition |
|---|---|---|---|---|
| Rabbit anti-CD3 | Nichirei (Tokyo, Japan) | 1:200 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Rat anti-B220 | Cedarlane (Ontario, Canada) | 1:1600 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rabbit anti-Iba1 | Wako (Osaka, Japan) | 1:1200 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rat anti-Gr1 | R and D system (Minneapolis, MN, USA) | 1:800 | 0.1% pepsin/0.2 N HCl | 37 °C, 5 min |
| Rabbit anti-Ki67 | Abcam (Tokyo, Japan) | 1:800 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Rabbit anti-LYVE-1 | Adipogen (San Diego, CA, USA) | 1:500 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rat anti-PNAd | BioLegend (San Diego, CA, USA) | 1:500 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Goat anti-CXCL9 | R and D system (Minneapolis, MN, USA) | 1:400 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Goat anti-CXCL13 | R and D system (Minneapolis, MN, USA) | 1:200 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Goat anti-Iba1 | Abcam (Tokyo, Japan) | 1:600 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Rabbit anti-CD68 | Abcam (Cambridge, UK) | 1:600 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Mouse anti-CD204 | TransGenic (Hyogo, Japan) | 1:400 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Rat anti-CD68 | Biolegend (San Diego, CA, USA) | 1:100 | 10 mM citrate buffer (pH 6.0) | 105 °C, 20 min |
| Rat anti-B220 (FITC) | Southern Biothech (Birmingham, AL, USA) | 1:300 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Rat anti-CD4 | Southern Biothech (Birmingham, AL, USA) | 1:200 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Rat anti-CD8 | Southern Biothech (Birmingham, AL, USA) | 1:400 | 20 mM Tris-HCl buffer (pH 9) | 105 °C, 20 min |
| Mouse anti-CD79a | Novus Biologicals (Centennial, CO, USA) | 1:500 | 10 mM citrate buffer (pH 6.0) | 90 °C, 30 min |
| Rat anti-CXCR5 | BD Pharmingen (Tokyo, Japan) | 1:100 | 10 mM citrate buffer (pH 6.0) | 90 °C, 30 min |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elewa, Y.H.A.; Masum, M.A.; Mohamed, S.K.A.; Islam, M.R.; Nakamura, T.; Ichii, O.; Kon, Y. The Ameliorative Effect of Dexamethasone on the Development of Autoimmune Lung Injury and Mediastinal Fat-Associated Lymphoid Clusters in an Autoimmune Disease Mouse Model. Int. J. Mol. Sci. 2022, 23, 4449. https://doi.org/10.3390/ijms23084449
Elewa YHA, Masum MA, Mohamed SKA, Islam MR, Nakamura T, Ichii O, Kon Y. The Ameliorative Effect of Dexamethasone on the Development of Autoimmune Lung Injury and Mediastinal Fat-Associated Lymphoid Clusters in an Autoimmune Disease Mouse Model. International Journal of Molecular Sciences. 2022; 23(8):4449. https://doi.org/10.3390/ijms23084449
Chicago/Turabian StyleElewa, Yaser Hosny Ali, Md Abdul Masum, Sherif Kh. A. Mohamed, Md Rashedul Islam, Teppei Nakamura, Osamu Ichii, and Yasuhiro Kon. 2022. "The Ameliorative Effect of Dexamethasone on the Development of Autoimmune Lung Injury and Mediastinal Fat-Associated Lymphoid Clusters in an Autoimmune Disease Mouse Model" International Journal of Molecular Sciences 23, no. 8: 4449. https://doi.org/10.3390/ijms23084449
APA StyleElewa, Y. H. A., Masum, M. A., Mohamed, S. K. A., Islam, M. R., Nakamura, T., Ichii, O., & Kon, Y. (2022). The Ameliorative Effect of Dexamethasone on the Development of Autoimmune Lung Injury and Mediastinal Fat-Associated Lymphoid Clusters in an Autoimmune Disease Mouse Model. International Journal of Molecular Sciences, 23(8), 4449. https://doi.org/10.3390/ijms23084449