
Quantitative and Functional Assessment of the Influence of Routinely Used Cryopreservation Media on Mononuclear Leukocytes for Medical Research

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Results
2.1. The Choice of Cryopreservation Medium Affects Recovery and Viability of MNCs after Thawing
2.2. Cryopreservation Alters the Relative Distribution of Leukocyte Populations
2.3. Analysis of the Effect of Cryopreservation Media on the Relative Frequency of Individual Leukocyte Subsets
2.4. A 1-h Resting Period after Thawing Does Not Dramatically Alter Leukocyte Population Amounts
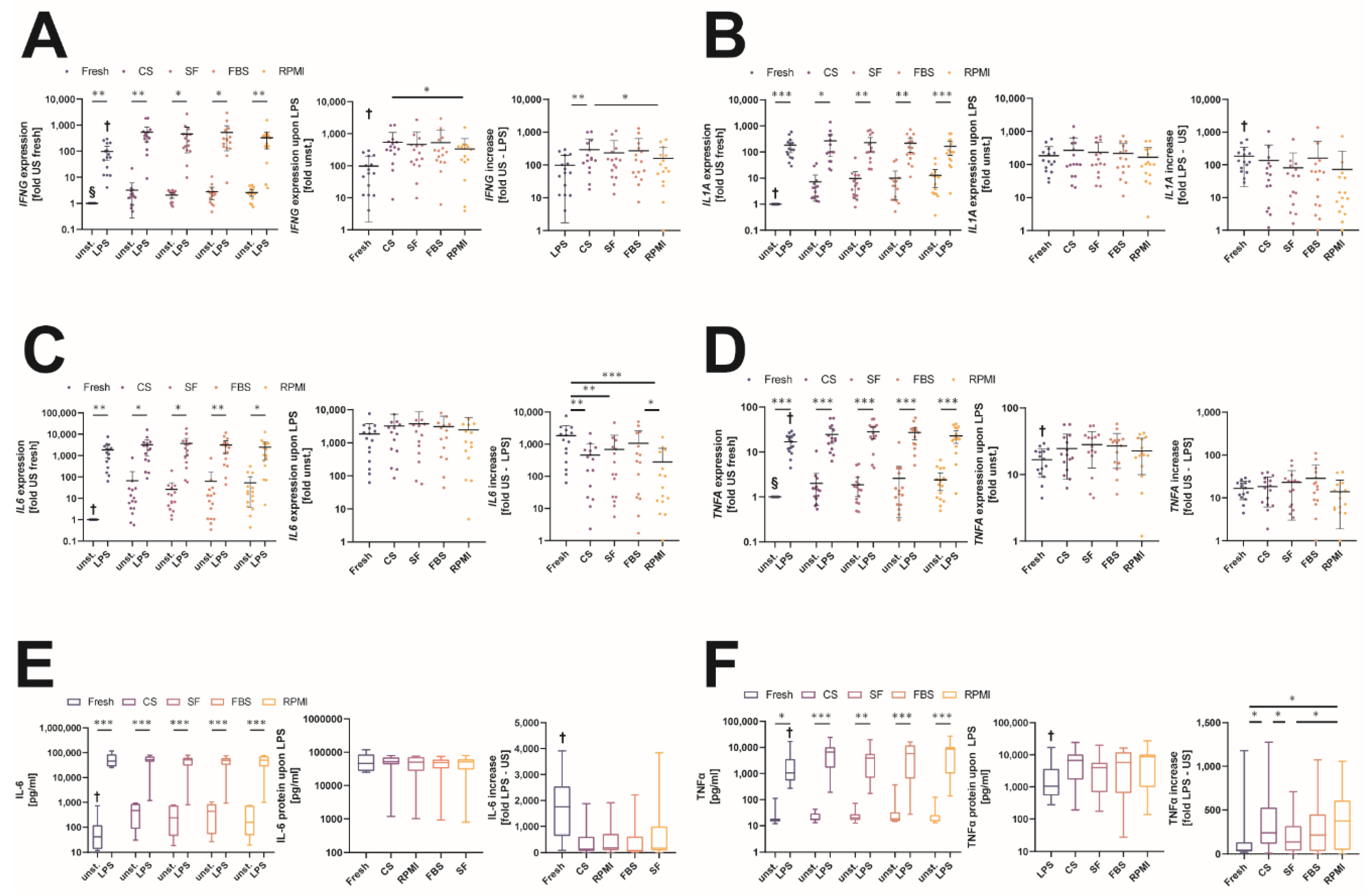
2.5. Changes in the Immunological Response to Bacterial Lipopolysaccharide after Storage in Different Cryopreservation Media
2.6. Cryopreservation Affects the Activatability of the CD11b Surface Integrin
2.7. Comparison of the Individual Cryopreservation Media for Different Experimental Research Questions
3. Discussion
4. Materials and Methods
4.1. Study Samples
4.2. MNC Isolation
4.3. Cryopreservation
4.4. Measurement of Absolute Cell Counts and Viability after Thawing
4.5. In Vitro MNC Stimulation
4.6. Flow Cytometry
4.7. Gene Expression Analysis
4.8. Protein Quantification
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hong, M.; Clubb, J.D.; Chen, Y.Y. Engineering CAR-T Cells for Next-Generation Cancer Therapy. Cancer Cell 2020, 38, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Labanieh, L.; Majzner, R.G.; Mackall, C.L. Programming CAR-T Cells to Kill Cancer. Nat. Biomed. Eng. 2018, 2, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Reinhardt, P.; Hus, I.; Tabarkiewicz, J.; Roliñski, J.; Barth, T.; Giannopoulos, K.; Dmoszyñska, A.; Wiesneth, M.; Schmitt, M. Large-Scale Generation of Autologous Dendritic Cells for Immunotherapy in Patients with Acute Myeloid Leukemia. Transfusion 2007, 47, 1588–1594. [Google Scholar] [CrossRef] [PubMed]
- Fesnak, A.; Lin, C.; Siegel, D.L.; Maus, M.V. CAR-T Cell Therapies From the Transfusion Medicine Perspective. Transfus. Med. Rev. 2016, 30, 139–145. [Google Scholar] [CrossRef]
- Krychtiuk, K.A.; Lenz, M.; Richter, B.; Hohensinner, P.J.; Kastl, S.P.; Mangold, A.; Huber, K.; Hengstenberg, C.; Wojta, J.; Heinz, G.; et al. Monocyte Subsets Predict Mortality after Cardiac Arrest. J. Leukoc. Biol. 2021, 109, 1139–1146. [Google Scholar] [CrossRef]
- Pfluecke, C.; Wydra, S.; Berndt, K.; Tarnowski, D.; Cybularz, M.; Jellinghaus, S.; Mierke, J.; Ende, G.; Poitz, D.M.; Barthel, P.; et al. Mon2-Monocytes and Increased CD-11b Expression before Transcatheter Aortic Valve Implantation Are Associated with Earlier Death. Int. J. Cardiol. 2020, 318, 115–120. [Google Scholar] [CrossRef]
- Shantsila, E.; Ghattas, A.; Griffiths, H.R.; Lip, G.Y.H. Mon2 Predicts Poor Outcome in ST-Elevation Myocardial Infarction. J. Intern. Med. 2019, 285, 301–316. [Google Scholar] [CrossRef]
- Leers, M.P.G.; Keuren, J.F.W.; Frissen, M.E.P.W.; Huts, M.; Kragten, J.A.; Jie, K.-S.G. The Pro- and Anticoagulant Role of Blood-Borne Phagocytes in Patients with Acute Coronary Syndrome. Thromb. Haemost. 2013, 110, 101–109. [Google Scholar] [CrossRef]
- Tortola, L.; Jacobs, A.; Pohlmeier, L.; Obermair, F.-J.; Ampenberger, F.; Bodenmiller, B.; Kopf, M. High-Dimensional T Helper Cell Profiling Reveals a Broad Diversity of Stably Committed Effector States and Uncovers Interlineage Relationships. Immunity 2020, 53, 597–613.e6. [Google Scholar] [CrossRef]
- Peng, Y.; Mentzer, A.J.; Liu, G.; Yao, X.; Yin, Z.; Dong, D.; Dejnirattisai, W.; Rostron, T.; Supasa, P.; Liu, C.; et al. Broad and Strong Memory CD4+ and CD8+ T Cells Induced by SARS-CoV-2 in UK Convalescent Individuals Following COVID-19. Nat. Immunol. 2020, 21, 1336–1345. [Google Scholar] [CrossRef]
- Newell, E.W.; Sigal, N.; Bendall, S.C.; Nolan, G.P.; Davis, M.M. Cytometry by Time-of-Flight Shows Combinatorial Cytokine Expression and Virus-Specific Cell Niches within a Continuum of CD8+ T Cell Phenotypes. Immunity 2012, 36, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Cossarizza, A.; Chang, H.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the Use of Flow Cytometry and Cell Sorting in Immunological Studies (Second Edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [PubMed]
- Mallone, R.; Mannering, S.I.; Brooks-Worrell, B.M.; Durinovic-Belló, I.; Cilio, C.M.; Wong, F.S.; Schloot, N.C.; on behalf of the Immunology of Diabetes Society T-Cell Workshop Committee. Isolation and Preservation of Peripheral Blood Mononuclear Cells for Analysis of Islet Antigen-Reactive T Cell Responses: Position Statement of the T-Cell Workshop Committee of the Immunology of Diabetes Society: PBMC Preparation for T Cell Assays. Clin. Exp. Immunol. 2011, 163, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Trück, J.; Mitchell, R.; Thompson, A.J.; Morales-Aza, B.; Clutterbuck, E.A.; Kelly, D.F.; Finn, A.; Pollard, A.J. Effect of Cryopreservation of Peripheral Blood Mononuclear Cells (PBMCs) on the Variability of an Antigen-Specific Memory B Cell ELISpot. Hum. Vaccin. Immunother. 2014, 10, 2490–2496. [Google Scholar] [CrossRef]
- Stevens, V.L.; Patel, A.V.; Feigelson, H.S.; Rodriguez, C.; Thun, M.J.; Calle, E.E. Cryopreservation of Whole Blood Samples Collected in the Field for a Large Epidemiologic Study. Cancer Epidemiol. Biomark. Prev. 2007, 16, 2160–2163. [Google Scholar] [CrossRef][Green Version]
- Martikainen, M.-V.; Roponen, M. Cryopreservation Affected the Levels of Immune Responses of PBMCs and Antigen-Presenting Cells. Toxicol. In Vitro 2020, 67, 104918. [Google Scholar] [CrossRef]
- Verschoor, C.P.; Kohli, V.; Balion, C. A Comprehensive Assessment of Immunophenotyping Performed in Cryopreserved Peripheral Whole Blood: Testing Cryopreserved Blood for Immunophenotyping. Cytometry 2018, 94, 818–826. [Google Scholar] [CrossRef]
- Draxler, D.F.; Madondo, M.T.; Hanafi, G.; Plebanski, M.; Medcalf, R.L. A Flowcytometric Analysis to Efficiently Quantify Multiple Innate Immune Cells and T Cell Subsets in Human Blood: Flowcytometric Analysis of Human Blood. Cytometry 2017, 91, 336–350. [Google Scholar] [CrossRef]
- Lemieux, J.; Jobin, C.; Simard, C.; Néron, S. A Global Look into Human T Cell Subsets before and after Cryopreservation Using Multiparametric Flow Cytometry and Two-Dimensional Visualization Analysis. J. Immunol. Methods 2016, 434, 73–82. [Google Scholar] [CrossRef]
- Weber, C.; Shantsila, E.; Hristov, M.; Caligiuri, G.; Guzik, T.; Heine, G.H.; Hoefer, I.E.; Monaco, C.; Peter, K.; Rainger, E.; et al. Role and Analysis of Monocyte Subsets in Cardiovascular Disease. Joint Consensus Document of the European Society of Cardiology (ESC) Working Groups “Atherosclerosis & Vascular Biology” and “Thrombosis.”. Thromb. Haemost. 2016, 116, 626–637. [Google Scholar] [CrossRef]
- Barcelo, H.; Faul, J.; Crimmins, E.; Thyagarajan, B. A Practical Cryopreservation and Staining Protocol for Immunophenotyping in Population Studies. Curr. Protoc. Cytom. 2018, 84, e35. [Google Scholar] [CrossRef] [PubMed]
- Njai, H.F.; Gombe, B.; Khamis, T.; Birungi, J.; Ruzagira, E.; Admassu, D.; Tarragona-Fiol, T.; Porter, K.; Stevens, G.; Mugisha, J.; et al. Setting Up a Standardized Peripheral Blood Mononuclear Cells Processing Laboratory to Support Multi-Center HIV/AIDS Vaccine and Intervention Trials. Lab. Med. 2011, 42, 711–718. [Google Scholar] [CrossRef]
- Krychtiuk, K.A.; Lenz, M.; Koller, L.; Honeder, M.C.; Wutzlhofer, L.; Zhang, C.; Chi, L.; Maurer, G.; Niessner, A.; Huber, K.; et al. Monocyte Subset Distribution Is Associated with Mortality in Critically Ill Patients. Thromb. Haemost. 2016, 116, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Weng, N.-P.; Akbar, A.N.; Goronzy, J. CD28− T Cells: Their Role in the Age-Associated Decline of Immune Function. Trends Immunol. 2009, 30, 306–312. [Google Scholar] [CrossRef]
- Kazem, N.; Sulzgruber, P.; Thaler, B.; Baumgartner, J.; Koller, L.; Laufer, G.; Steinlechner, B.; Hohensinner, P.; Wojta, J.; Niessner, A. CD8+CD28null T Lymphocytes Are Associated with the Development of Atrial Fibrillation after Elective Cardiac Surgery. Thromb. Haemost. 2020, 120, 1182–1187. [Google Scholar] [CrossRef]
- Sulzgruber, P.; Koller, L.; Winter, M.-P.; Richter, B.; Blum, S.; Korpak, M.; Hülsmann, M.; Goliasch, G.; Wojta, J.; Niessner, A. The Impact of CD4+CD28null T-Lymphocytes on Atrial Fibrillation and Mortality in Patients with Chronic Heart Failure. Thromb. Haemost. 2017, 117, 349–356. [Google Scholar] [CrossRef]
- Hanley, P.J. Fresh versus Frozen: Effects of Cryopreservation on CAR T Cells. Mol. Ther. 2019, 27, 1213–1214. [Google Scholar] [CrossRef]
- Schmid, M.C.; Khan, S.Q.; Kaneda, M.M.; Pathria, P.; Shepard, R.; Louis, T.L.; Anand, S.; Woo, G.; Leem, C.; Faridi, M.H.; et al. Integrin CD11b Activation Drives Anti-Tumor Innate Immunity. Nat. Commun. 2018, 9, 5379. [Google Scholar] [CrossRef]
- Zhou, X.; Gao, X.-P.; Fan, J.; Liu, Q.; Anwar, K.N.; Frey, R.S.; Malik, A.B. LPS Activation of Toll-like Receptor 4 Signals CD11b/CD18 Expression in Neutrophils. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 288, L655–L662. [Google Scholar] [CrossRef]
- Waller, K.; James, C.; de Jong, A.; Blackmore, L.; Ma, Y.; Stagg, A.; Kelsell, D.; O’Dwyer, M.; Hutchins, R.; Alazawi, W. ADAM17-Mediated Reduction in CD14++CD16+ Monocytes Ex Vivo and Reduction in Intermediate Monocytes with Immune Paresis in Acute Pancreatitis and Acute Alcoholic Hepatitis. Front. Immunol. 2019, 10, 1902. [Google Scholar] [CrossRef]
- Thaler, B.; Hohensinner, P.J.; Krychtiuk, K.A.; Matzneller, P.; Koller, L.; Brekalo, M.; Maurer, G.; Huber, K.; Zeitlinger, M.; Jilma, B.; et al. Differential In Vivo Activation of Monocyte Subsets during Low-Grade Inflammation through Experimental Endotoxemia in Humans. Sci. Rep. 2016, 6, 30162. [Google Scholar] [CrossRef] [PubMed]
- Braudeau, C.; Salabert-Le Guen, N.; Chevreuil, J.; Rimbert, M.; Martin, J.C.; Josien, R. An Easy and Reliable Whole Blood Freezing Method for Flow Cytometry Immuno-Phenotyping and Functional Analyses. Cytom. B Clin. Cytom. 2021, 100, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Toh, Z.Q.; Reitsma, A.; Do, L.A.H.; Nathanielsz, J.; Licciardi, P.V. Effect of Peripheral Blood Mononuclear Cell Cryopreservation on Innate and Adaptive Immune Responses. J. Immunol. Methods 2019, 465, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Sart, S.; Ma, T.; Li, Y. Cryopreservation of Pluripotent Stem Cell Aggregates in Defined Protein-Free Formulation. Biotechnol. Prog. 2013, 29, 143–153. [Google Scholar] [CrossRef]
- Clarke, D.M.; Yadock, D.J.; Nicoud, I.B.; Mathew, A.J.; Heimfeld, S. Improved Post-Thaw Recovery of Peripheral Blood Stem/Progenitor Cells Using a Novel Intracellular-like Cryopreservation Solution. Cytotherapy 2009, 11, 472–479. [Google Scholar] [CrossRef]
- Campbell, L.H.; Brockbank, K.G.M. Serum-Free Solutions for Cryopreservation of Cells. In Vitro Cell. Dev. Biol. Anim. 2007, 43, 269–275. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, P.; Liu, H.; Zhu, Z.; Li, C.; Gao, Y. The State of T Cells before Cryopreservation: Effects on Post-Thaw Proliferation and Function. Cryobiology 2017, 79, 65–70. [Google Scholar] [CrossRef]
- Appleby, L.J.; Nausch, N.; Midzi, N.; Mduluza, T.; Allen, J.E.; Mutapi, F. Sources of Heterogeneity in Human Monocyte Subsets. Immunol. Lett. 2013, 152, 32–41. [Google Scholar] [CrossRef]
- Kaiser, D.; Otto, N.M.; McCallion, O.; Hoffmann, H.; Zarrinrad, G.; Stein, M.; Beier, C.; Matz, I.; Herschel, M.; Hester, J.; et al. Freezing Medium Containing 5% DMSO Enhances the Cell Viability and Recovery Rate After Cryopreservation of Regulatory T Cell Products Ex Vivo and In Vivo. Front. Cell Dev. Biol. 2021, 9, 750286. [Google Scholar] [CrossRef]
- Ong, S.-M.; Hadadi, E.; Dang, T.-M.; Yeap, W.-H.; Tan, C.T.-Y.; Ng, T.-P.; Larbi, A.; Wong, S.-C. The Pro-Inflammatory Phenotype of the Human Non-Classical Monocyte Subset Is Attributed to Senescence. Cell Death Dis. 2018, 9, 266. [Google Scholar] [CrossRef]
- Fendl, B.; Weiss, R.; Eichhorn, T.; Spittler, A.; Fischer, M.B.; Weber, V. Storage of Human Whole Blood, but Not Isolated Monocytes, Preserves the Distribution of Monocyte Subsets. Biochem. Biophys. Res. Commun. 2019, 517, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Rundgren, I.M.; Bruserud, Ø.; Ryningen, A.; Ersvær, E. Standardization of Sampling and Sample Preparation for Analysis of Human Monocyte Subsets in Peripheral Blood. J. Immunol. Methods 2018, 461, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Zarife, M.A.S.; Reis, E.A.G.; Carmo, T.M.A.; Lopes, G.B.; Brandão, E.C.M.; Silva, H.R.; Santana, N.; Martins-Filho, O.A.; Reis, M.G. Increased Frequency of CD56Bright NK-Cells, CD3− CD16+ CD56− NK-Cells and Activated CD4 + T-Cells or B-Cells in Parallel with CD4+ CDC25High T-Cells Control Potentially Viremia in Blood Donors with HCV: Lymphocyte Subsets in Blood Donors with HCV. J. Med. Virol. 2009, 81, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Hayden, H.; Friedl, J.; Dettke, M.; Sachet, M.; Hassler, M.; Dubsky, P.; Bachleitner-Hofmann, T.; Gnant, M.; Stift, A. Cryopreservation of Monocytes Is Superior to Cryopreservation of Immature or Semi-Mature Dendritic Cells for Dendritic Cell-Based Immunotherapy. J. Immunother. 2009, 32, 638–654. [Google Scholar] [CrossRef]
- Ferdowsi, S.; Abbasi-Malati, Z.; Pourfathollah, A.A. Leukocyte Reduction Filters as an Alternative Source of Peripheral Blood Leukocytes for Research. Hematol. Transfus. Cell Ther. 2021, 43, 494–498. [Google Scholar] [CrossRef]
- Haider, P.; Kral-Pointner, J.B.; Mayer, J.; Richter, M.; Kaun, C.; Brostjan, C.; Eilenberg, W.; Fischer, M.B.; Speidl, W.S.; Hengstenberg, C.; et al. Neutrophil Extracellular Trap Degradation by Differently Polarized Macrophage Subsets. ATVB 2020, 40, 2265–2278. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Research Question | CS | SF | FBS | RPMI |
|---|---|---|---|---|
| Recovery and Viability (Figure 1) | 2 | 4 | 1 | 3 |
| Cell population relative amount (Figure 2) | 1 | 3 | 2 | 4 |
| Cell population dead cells (Figure 2) | 1 | 4 | 2 | 3 |
| Monocyte subsets (Figure 3) | 1 | 2 | 4 | 3 |
| T cell subsets (Figure 3) | 4 | 1 | 2 | 2 |
| Cytokine measurement (Figure 5) | 2 | 1 | 4 | 3 |
| Monocyte CD11b activation (Figure 6) | 1 | 3 | 2 | 4 |
| Cumulative performance (overall rank) | 1 | 3 | 2 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haider, P.; Hoberstorfer, T.; Salzmann, M.; Fischer, M.B.; Speidl, W.S.; Wojta, J.; Hohensinner, P.J. Quantitative and Functional Assessment of the Influence of Routinely Used Cryopreservation Media on Mononuclear Leukocytes for Medical Research. Int. J. Mol. Sci. 2022, 23, 1881. https://doi.org/10.3390/ijms23031881
Haider P, Hoberstorfer T, Salzmann M, Fischer MB, Speidl WS, Wojta J, Hohensinner PJ. Quantitative and Functional Assessment of the Influence of Routinely Used Cryopreservation Media on Mononuclear Leukocytes for Medical Research. International Journal of Molecular Sciences. 2022; 23(3):1881. https://doi.org/10.3390/ijms23031881
Chicago/Turabian StyleHaider, Patrick, Timothy Hoberstorfer, Manuel Salzmann, Michael B. Fischer, Walter S. Speidl, Johann Wojta, and Philipp J. Hohensinner. 2022. "Quantitative and Functional Assessment of the Influence of Routinely Used Cryopreservation Media on Mononuclear Leukocytes for Medical Research" International Journal of Molecular Sciences 23, no. 3: 1881. https://doi.org/10.3390/ijms23031881
APA StyleHaider, P., Hoberstorfer, T., Salzmann, M., Fischer, M. B., Speidl, W. S., Wojta, J., & Hohensinner, P. J. (2022). Quantitative and Functional Assessment of the Influence of Routinely Used Cryopreservation Media on Mononuclear Leukocytes for Medical Research. International Journal of Molecular Sciences, 23(3), 1881. https://doi.org/10.3390/ijms23031881