
Non-Coding RNAs in Tuberculosis Epidemiology: Platforms and Approaches for Investigating the Genome’s Dark Matter


Abstract
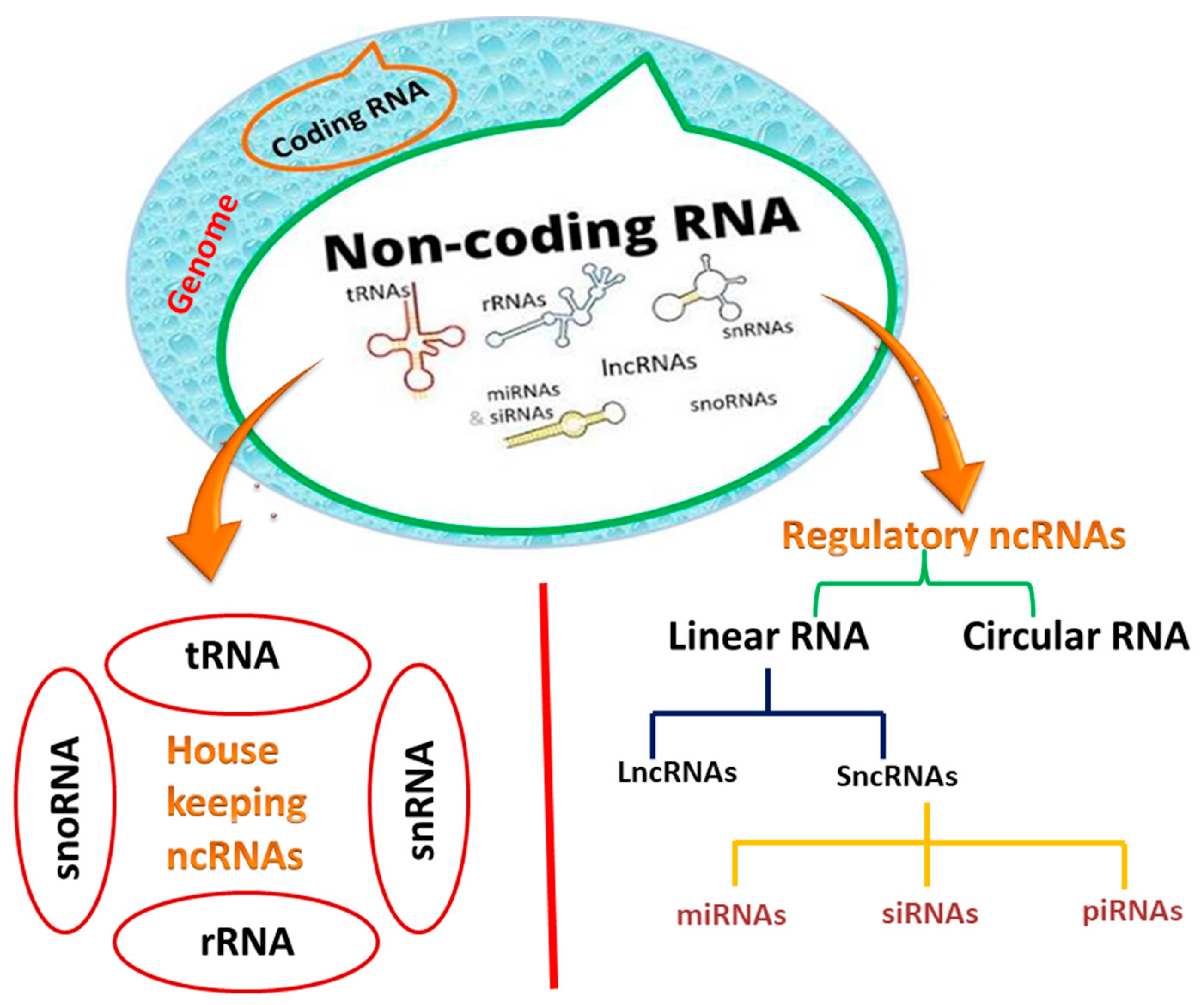
1. Background
2. Classification and Biogenesis of Noncoding RNAs
2.1. miRNA
2.2. lncRNAs
2.3. CircRNAs
3. Role of ncRNAs in the Molecular Epidemiologic Studies of Tuberculosis
3.1. The Role of miRNA
3.1.1. miRNA-29
3.1.2. miRNA-147
3.1.3. miRNA-21
3.1.4. miRNA-99b
3.1.5. miRNA-125b
3.1.6. miRNA-155
3.1.7. miRNA-144*
3.1.8. miRNAs-223 and 424
3.2. The Role of lncRNA
3.2.1. lncRNA MEG3
3.2.2. lncRNA CD244
3.2.3. lncRNA NEAT1
3.2.4. lncRNA PCED1B-AS1
3.3. The Role of circRNA
3.3.1. circRNA 051239
3.3.2. hsa-circRNA-100237
3.3.3. circAGFG1
3.3.4. CircTRAPPC6B
4. ncRNAs as a Diagnostic and Therapeutic Tool for Tuberculosis
4.1. The Potential of miRNAs as Diagnostic and Therapeutic Biomarkers for Tuberculosis
4.2. LncRNAs as Diagnostic and Therapeutic Biomarkers for Tuberculosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Noncoding RNA | Function in Tuberculosis | Action | References |
|---|---|---|---|
| miR-26-5p | Etiology | Inhibition of innate immunity | [145] |
| miR-132-3p | [146] | ||
| miR-155-5p | [147] | ||
| miR-29-3p | [148] | ||
| miR-21-5p | Suppression of inflammation | [149] | |
| miR-27b-3p | [150] | ||
| miR-99b-5p | [151] | ||
| miR-125-5p | [152] | ||
| miR-146a-5p | [153] | ||
| miR-223-3p | [154] | ||
| Let-7f | [155] | ||
| miR-20b-5p | [156] | ||
| miR-142-3p | [157] | ||
| miR-33 | Inhibition of phagosome maturation and autophagy | [158] | |
| miR-27a-5p | [159] | ||
| miR-125a-3p | [160] | ||
| miR-144-5p | [161] | ||
| miR-889-5p | [162] | ||
| miR-155-5p | Apoptosis Inhibition | [163] | |
| miR-582-5p | [20] | ||
| miR-769-5p | Diagnosis | Downregulation in TB patients | [164] |
| miR-320a | [165] | ||
| miR-22-3p | [166] | ||
| hsa_circ_0001380 | [167] | ||
| miR-423-5p | Upregulation in TB patients | [168] | |
| miR-17-5p | [169] | ||
| miR-20b-5p | [170] | ||
| lncRNA LOC152742 | [171] | ||
| hsa_circRNA_001937 | [172] | ||
| has_circRNA_051239 | [173] | ||
| hsa_circRNA_404022 | [174] | ||
| has_circRNA_029965 | [175] | ||
| lncRNAs NEAT1 | Treatment | Downregulation during drug treatment, linked with disease improvement | [176] |
| lncRNAs NEAT2 | [177] | ||
| lncRNA 152742 | [178] | ||
| circTRAPPC6B | [179] | ||
| lncRNAENST00000429730.1 | Downregulation during drug treatment, linked with entire inactivation of TB lesions from sputum negative patients | [180] | |
| lncRNA MSTRG.93125.4 | [181] |
4.3. CircRNAs as a Diagnostic and Therapeutic Biomarker for Tuberculosis
5. Methods for Profiling ncRNAs Expression
5.1. Microarray
5.2. Serial Analysis of Gene Expression
5.3. RNA Sequencing (RNA-seq)
5.4. In Silico Investigation
6. Methods for Validating ncRNA Expression
6.1. Northern Blotting (NB)
6.2. RT-qPCR
6.3. FISH
7. Potentially Beneficial Therapeutic Interventions
7.1. Noncoding RNAs Targeting: ASOs
7.2. Non-Coding RNAs Targeting: RNAi
7.3. ncRNAs Editing with CRISPR-Cas9
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kiazyk, S.; Ball, T. Tuberculosis (TB): Latent tuberculosis infection: An overview. Can. Commun. Dis. Rep. 2017, 43, 62. [Google Scholar] [CrossRef] [PubMed]
- Rehm, J.; Samokhvalov, A.V.; Neuman, M.G.; Room, R.; Parry, C.; Lönnroth, K.; Patra, J.; Poznyak, V.; Popova, S. The association between alcohol use, alcohol use disorders and tuberculosis (TB). A systematic review. BMC Public Health 2009, 9, 450. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Responding to Community Spread of COVID-19. Reference WHO/COVID-19/Community_Transmission/2020.1. 2020. Available online: https://www.who.int/publications/i/item/responding-to-community-spread-of-covid-19 (accessed on 1 April 2022).
- Zijenah, L.S. The world health organization recommended TB diagnostic tools. Tuberculosis 2018, 2, 71–90. [Google Scholar]
- Ottenhoff, T.H. Overcoming the global crisis: “Yes, we can”, but also for TB…? Eur. J. Immunol. 2009, 39, 2014–2020. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Mondal, R. Extensively drug-resistant tuberculosis: Current challenges and threats. FEMS Immunol. Med. Microbiol. 2008, 53, 145–150. [Google Scholar] [CrossRef]
- Koch, A.; Cox, H.; Mizrahi, V. Drug-resistant tuberculosis: Challenges and opportunities for diagnosis and treatment. Curr. Opin. Pharmacol. 2018, 42, 7–15. [Google Scholar] [CrossRef]
- Tahir ul Qamar, M.; Zhu, X.; Khan, M.S.; Xing, F.; Chen, L.L. Pan-genome: A promising resource for noncoding RNA discovery in plants. Plant Genome 2020, 13, e20046. [Google Scholar] [CrossRef]
- Jacques, P.-É.; Gervais, A.L.; Cantin, M.; Lucier, J.-F.o.; Dallaire, G.; Drouin, G.; Gaudreau, L.; Goulet, J.; Brzezinski, R. MtbRegList, a database dedicated to the analysis of transcriptional regulation in Mycobacterium tuberculosis. Bioinformatics 2005, 21, 2563–2565. [Google Scholar] [CrossRef][Green Version]
- Oliva, G.; Sahr, T.; Buchrieser, C. Small RNAs, 5′ UTR elements and RNA-binding proteins in intracellular bacteria: Impact on metabolism and virulence. FEMS Microbiol. Rev. 2015, 39, 331–349. [Google Scholar] [CrossRef]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, biology and functioning. In Non-Coding RNAs Colorectal Cancer; Springer: Berlin, Germany, 2016; pp. 3–17. [Google Scholar]
- Nelson, W.C.; Stegen, J.C. The reduced genomes of Parcubacteria (OD1) contain signatures of a symbiotic lifestyle. Front. Microbiol. 2015, 6, 713. [Google Scholar] [CrossRef] [PubMed]
- Memon, D.; Bi, J.; Miller, C.J. In silico prediction of housekeeping long intergenic non-coding RNAs reveals HKlincR1 as an essential player in lung cancer cell survival. Sci. Rep. 2019, 9, 7372. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Lin, Y.; Wu, J. Long non-coding RNA expression profiling of mouse testis during postnatal development. PLoS ONE 2013, 8, e75750. [Google Scholar] [CrossRef] [PubMed]
- Guglas, K.; Bogaczyńska, M.; Kolenda, T.; Ryś, M.; Teresiak, A.; Bliźniak, R.; Łasińska, I.; Mackiewicz, J.; Lamperska, K. lncRNA in HNSCC: Challenges and potential. Contemp. Oncol. 2017, 21, 259. [Google Scholar] [CrossRef]
- Zhang, B.; Li, W.; Liu, K.; Jia, Q.; Zhang, H.; Bie, Q. The roles of host noncoding RNAs in Mycobacterium tuberculosis infection. Front. Immunol. 2021, 12, 1689. [Google Scholar]
- Kuhn, D.E.; Martin, M.M.; Feldman, D.S.; Terry, A.V., Jr.; Nuovo, G.J.; Elton, T.S. Experimental validation of miRNA targets. Methods 2008, 44, 47–54. [Google Scholar] [CrossRef]
- Bavelloni, A.; Ramazzotti, G.; Poli, A.; Piazzi, M.; Focaccia, E.; Blalock, W.; Faenza, I. MiRNA-210: A current overview. Anticancer. Res. 2017, 37, 6511–6521. [Google Scholar]
- Miotto, P.; Mwangoka, G.; Valente, I.C.; Norbis, L.; Sotgiu, G.; Bosu, R.; Ambrosi, A.; Codecasa, L.R.; Goletti, D.; Matteelli, A. miRNA signatures in sera of patients with active pulmonary tuberculosis. PLoS ONE 2013, 8, e80149. [Google Scholar] [CrossRef]
- Gan, E.-S.; Huang, J.; Ito, T. Functional roles of histone modification, chromatin remodeling and microRNAs in Arabidopsis flower development. Int. Rev. Cell Mol. Biol. 2013, 305, 115–161. [Google Scholar]
- Beermann, J.; Piccoli, M.-T.; Viereck, J.; Thum, T. Non-coding RNAs in development and disease: Background, mechanisms, and therapeutic approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: microRNA biogenesis pathways and their regulation. Nat. Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Curtis, H.J.; Sibley, C.R.; Wood, M.J. Mirtrons, an emerging class of atypical miRNA. Wiley Interdiscip. Rev. RNA 2012, 3, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, C.; Lv, M.; Hu, Q.; Guo, L.; Xiong, D. Correlation between alterations of gut microbiota and miR-122-5p expression in patients with type 2 diabetes mellitus. Ann. Transl. Med. 2020, 8, 1481. [Google Scholar] [CrossRef] [PubMed]
- Banada, P.P.; Sivasubramani, S.K.; Blakemore, R.; Boehme, C.; Perkins, M.D.; Fennelly, K.; Alland, D. Containment of bioaerosol infection risk by the Xpert MTB/RIF assay and its applicability to point-of-care settings. J. Clin. Microbiol. 2010, 48, 3551–3557. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Y.; Lee, M.-C.; Shu, C.-C.; Lee, C.-H.; Lee, L.-N.; Chao, K.-M.; Chang, F.-Y. Optimal duration of anti-TB treatment in patients with diabetes: Nine or six months? Chest 2015, 147, 520–528. [Google Scholar] [CrossRef]
- Naing, N.N.; D’Este, C.; Isa, A.R.; Salleh, R.; Bakar, N.; Mahmod, M.R. Factors contributing to poor compliance with anti-TB treatment among tuberculosis patients. Southeast Asian J. Trop. Med. Public Health 2001, 32, 369–382. [Google Scholar]
- Yang, G.; Lu, X.; Yuan, L. LncRNA: A link between RNA and cancer. Biochim. Biophys. Acta BBA-Gene Regul. Mech. 2014, 1839, 1097–1109. [Google Scholar] [CrossRef]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing miRNA–lncRNA interactions. In Long Non-Coding RNAs; Springer: Berlin, Germany, 2016; pp. 271–286. [Google Scholar]
- Chen, X.; Yan, G.-Y. Novel human lncRNA–disease association inference based on lncRNA expression profiles. Bioinformatics 2013, 29, 2617–2624. [Google Scholar] [CrossRef]
- Ferre, F.; Colantoni, A.; Helmer-Citterich, M. Revealing protein–lncRNA interaction. Brief. Bioinform. 2016, 17, 106–116. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Tanigawa, A.; Naganuma, T.; Ohkawa, Y.; Souquere, S.; Pierron, G.; Hirose, T. SWI/SNF chromatin-remodeling complexes function in noncoding RNA-dependent assembly of nuclear bodies. Proc. Natl. Acad. Sci. USA 2015, 112, 4304–4309. [Google Scholar] [CrossRef]
- Quinodoz, S.; Guttman, M. Long noncoding RNAs: An emerging link between gene regulation and nuclear organization. Trends Cell Biol. 2014, 24, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Khan, F.B.; Safdari, H.A.; Almatroudi, A.; Alzohairy, M.A.; Safdari, M.; Amirizadeh, M.; Rehman, S.; Equbal, M.J.; Hoque, M. Prospective therapeutic potential of Tanshinone IIA: An updated overview. Pharmacol. Res. 2021, 164, 105364. [Google Scholar] [CrossRef] [PubMed]
- Parasramka, M.A.; Maji, S.; Matsuda, A.; Yan, I.K.; Patel, T. Long non-coding RNAs as novel targets for therapy in hepatocellular carcinoma. Pharmacol. Ther. 2016, 161, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Koch, L. Screening for lncRNA function. Nat. Rev. Genet. 2017, 18, 70. [Google Scholar] [CrossRef] [PubMed]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E. Translation of circRNAs. Mol. Cell 2017, 66, 9–21.e27. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.; Harries, L.W. Circular RNAs (circRNAs) in health and disease. Genes 2017, 8, 353. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Gao, K.; Li, R.; Fu, Y. Dysregulated circRNAs in plasma from active tuberculosis patients. J. Cell. Mol. Med. 2018, 22, 4076–4084. [Google Scholar] [CrossRef]
- Fan, X.; Weng, X.; Zhao, Y.; Chen, W.; Gan, T.; Xu, D. Circular RNAs in cardiovascular disease: An overview. BioMed Res. Int. 2017, 2017, 5135781. [Google Scholar] [CrossRef]
- Ebbesen, K.K.; Kjems, J.; Hansen, T.B. Circular RNAs: Identification, biogenesis and function. Biochim. Biophys. Acta BBA-Gene Regul. Mech. 2016, 1859, 163–168. [Google Scholar] [CrossRef]
- Liu, Y.; Tahir ul Qamar, M.; Feng, J.-W.; Ding, Y.; Wang, S.; Wu, G.; Ke, L.; Xu, Q.; Chen, L.-L. Comparative analysis of miniature inverted–repeat transposable elements (MITEs) and long terminal repeat (LTR) retrotransposons in six Citrus species. BMC Plant Biol. 2019, 19, 140. [Google Scholar] [CrossRef]
- Kumarswamy, R.; Volkmann, I.; Thum, T. Regulation and function of miRNA-21 in health and disease. RNA Biol. 2011, 8, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-C.; Du, L.-Q.; Tian, L.-L.; Wu, H.-L.; Jiang, X.-Y.; Zhang, H.; Li, D.-G.; Wang, Y.-Y.; Wu, H.-Y.; She, Y. Expression and function of miRNA in postoperative radiotherapy sensitive and resistant patients of non-small cell lung cancer. Lung Cancer 2011, 72, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Kadkhoda, S.; Ghafouri-Fard, S. Function of miRNA-145-5p in the pathogenesis of human disorders. Pathol. Res. Pract. 2022, 231, 153780. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.E.; Shao, W.-H. Epigenetic alterations in immune cells of systemic Lupus erythematosus and therapeutic implications. Cells 2022, 11, 506. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Huang, M.; Li, T.; Li, L.; Sussman, H.; Dai, Y.; Siemann, D.; Xie, M.; Tang, X. Towards an integrative understanding of cancer mechanobiology: Calcium, YAP, and microRNA under biophysical forces. Soft Matter 2022, 18, 1112–1148. [Google Scholar] [CrossRef] [PubMed]
- Doxakis, E. Insights into the multifaceted role of circular RNAs: Implications for Parkinson’s disease pathogenesis and diagnosis. NPJ Parkinsons Dis. 2022, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Ganguly, N.; Banerjee, S. Exploring clinical implications and role of non-coding RNAs in lung carcinogenesis. Mol. Biol. Rep. 2022, 49, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Xu, S.; Liu, X.; Zhang, Q.; Xu, X.; Liu, M.; Hua, M.; Li, N.; Yao, H.; Cao, X. The microRNA miR-29 controls innate and adaptive immune responses to intracellular bacterial infection by targeting interferon-γ. Nat. Immunol. 2011, 12, 861–869. [Google Scholar] [CrossRef]
- Laws, M.; Jin, P.; Rahman, K.M. Efflux pumps in Mycobacterium tuberculosis and their inhibition to tackle antimicrobial resistance. Trends Microbiol. 2022, 30, 57–68. [Google Scholar] [CrossRef]
- Pauley, K.M.; Satoh, M.; Pauley, B.A.; Dominguez-Gutierrez, P.R.; Wallet, S.M.; Holliday, L.S.; Cha, S.; Reeves, W.H.; Chan, E.K. Formation of GW/P bodies as marker for microRNA-mediated regulation of innate immune signaling in THP-1 cells. Immunol. Cell Biol. 2010, 88, 205–212. [Google Scholar] [CrossRef]
- Savan, R. Post-transcriptional regulation of interferons and their signaling pathways. J. Interferon Cytokine Res. 2014, 34, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.; Joshi, R.; Bandyopadhyay, M.; Narang, P.; Dogra, S.; Taksande, B.; Kalantri, S. Sensitivity of a whole-blood interferon-gamma assay among patients with pulmonary tuberculosis and variations in T-cell responses during anti-tuberculosis treatment. Infection 2007, 35, 98–103. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ferrand, R.; Bothamley, G.; Whelan, A.; Dockrell, H. Interferon-gamma responses to ESAT-6 in tuberculosis patients early into and after anti-tuberculosis treatment. Int. J. Tuberc. Lung Dis. 2005, 9, 1034–1039. [Google Scholar] [PubMed]
- Cartwright, N.; Demaine, A.; Jahromi, M.; Sanders, H.; Kaminski, E.R. A study of cytokine protein secretion, frequencies of cytokine expressing cells and IFN-G gene polymorphisms in normal individuals. Transplantation 1999, 68, 1546–1552. [Google Scholar] [CrossRef]
- Cavalla, F.; Biguetti, C.C.; Colavite, P.M.; Silveira, E.V.; Martins Jr, W.; Letra, A.; Trombone, A.P.F.; Silva, R.M.; Garlet, G.P. TBX21-1993T/C (rs4794067) polymorphism is associated with increased risk of chronic periodontitis and increased T-bet expression in periodontal lesions, but does not significantly impact the IFN-g transcriptional level or the pattern of periodontophatic bacterial infection. Virulence 2015, 6, 293–304. [Google Scholar]
- Chen, S.; Crabill, G.A.; Pritchard, T.S.; McMiller, T.L.; Wei, P.; Pardoll, D.M.; Pan, F.; Topalian, S.L. Mechanisms regulating PD-L1 expression on tumor and immune cells. J. Immunother. Cancer 2019, 7, 1–12. [Google Scholar] [CrossRef]
- Galvin, J.; Tiberi, S.; Akkerman, O.; Kerstjens, H.; Kunst, H.; Kurhasani, X.; Ambrosino, N.; Migliori, G. Pulmonary tuberculosis in intensive care setting, with a focus on the use of severity scores, a multinational collaborative systematic review. Pulmonology 2022, 28. in press. [Google Scholar] [CrossRef]
- Sun, X.; Liu, K.; Zhao, Y.; Zhang, T. High miRNA-378 expression has high diagnostic values for pulmonary tuberculosis and predicts adverse outcomes. BMC Mol. Cell Biol. 2022, 23, 14. [Google Scholar] [CrossRef]
- Murdaca, G.; Paladin, F.; Tonacci, A.; Borro, M.; Greco, M.; Gerosa, A.; Isola, S.; Allegra, A.; Gangemi, S. Involvement of Il-33 in the pathogenesis and prognosis of major respiratory viral infections: Future perspectives for personalized therapy. Biomedicines 2022, 10, 715. [Google Scholar] [CrossRef]
- Sailliet, N.; Ullah, M.; Dupuy, A.; Silva, A.K.; Gazeau, F.; Le Mai, H.; Brouard, S. Extracellular vesicles in transplantation. Front. Immunol. 2022, 13, 1521–1531. [Google Scholar] [CrossRef]
- Adewumi, A.T.; Elrashedy, A.; Soremekun, O.S.; Ajadi, M.B.; Soliman, M.E. Weak spots inhibition in the Mycobacterium tuberculosis antigen 85C target for antitubercular drug design through selective irreversible covalent inhibitor-SER124. J. Biomol. Struct. Dyn. 2022, 40, 2934–2954. [Google Scholar] [CrossRef] [PubMed]
- Arish, M.; Naz, F. Macrophage plasticity as a therapeutic target in tuberculosis. Eur. J. Immunol. 2022, 1–9. [Google Scholar] [CrossRef]
- Birhanu, A.G.; Gómez-Muńoz, M.; Kalayou, S.; Riaz, T.; Lutter, T.; Yimer, S.A.; Abebe, M.; Tønjum, T. Proteome profiling of Mycobacterium tuberculosis cells exposed to nitrosative stress. ACS Omega 2022, 7, 3470–3482. [Google Scholar] [CrossRef] [PubMed]
- Lim, Z.L.; Drever, K.; Dhar, N.; Cole, S.T.; Chen, J.M. Mycobacterium tuberculosis EspK has active but distinct roles in the secretion of EsxA and EspB. J. Bacteriol. 2022, 10, e00060-22. [Google Scholar] [CrossRef]
- Clarke, C.; Cooper, D.V.; Miller, M.A.; Goosen, W.J. Detection of Mycobacterium tuberculosis complex DNA in oronasal swabs from infected African buffaloes (Syncerus caffer). Sci. Rep. 2022, 12, 1834. [Google Scholar] [CrossRef]
- Pérez, I.; Campos-Pardos, E.; Díaz, C.; Uranga, S.; Sayes, F.; Vicente, F.; Aguiló, N.; Brosch, R.; Martín, C.; Gonzalo-Asensio, J. The Mycobacterium tuberculosis PhoPR virulence system regulates expression of the universal second messenger c-di-AMP and impacts vaccine safety and efficacy. Mol. Ther. Nucleic Acids 2022, 27, 1235–1248. [Google Scholar] [CrossRef]
- Lata, S.; Mahatha, A.C.; Mal, S.; Gupta, U.D.; Kundu, M.; Basu, J. Unravelling novel roles of the Mycobacterium tuberculosis transcription factor Rv0081 in regulation of the nucleoid-associated proteins Lsr2 and EspR, cholesterol utilization and subversion of lysosomal trafficking in macrophages. Mol. Microbiol. 2022. [Google Scholar] [CrossRef]
- Petruccioli, E.; Farroni, C.; Cuzzi, G.; Vanini, V.; Palmieri, F.; Vittozzi, P.; Goletti, D. VIDAS® TB-IGRA reagents induce a CD4+ and CD8+ T-cell IFN-γ response for both TB infection and active TB. Int. J. Tuberc. Lung Dis. Off. J. Int. Union Against Tuberc. Lung Dis. 2022, 26, 65–68. [Google Scholar] [CrossRef]
- Khabibullina, N.F.; Kutuzova, D.M.; Burmistrova, I.A.; Lyadova, I.V. The biological and clinical aspects of a latent tuberculosis infection. Trop. Med. Infect. Dis. 2022, 7, 48. [Google Scholar] [CrossRef]
- Al Tbeishat, H. Novel in silico mRNA vaccine design exploiting proteins of M. tuberculosis that modulates host immune responses by inducing epigenetic modifications. Sci. Rep. 2022, 12, 4645. [Google Scholar] [CrossRef]
- Junqueira-Kipnis, A.P.; de Castro Souza, C.; de Oliveira Carvalho, A.C.; de Oliveira, F.M.; Almeida, V.P.; de Paula, A.R.; Celes, M.R.; Kipnis, A. Protease-based subunit vaccine in mice boosts BCG protection against Mycobacterium tuberculosis. Vaccines 2022, 10, 306. [Google Scholar] [CrossRef] [PubMed]
- Dockrell, H.M.; Ramachandran, V.; Bothamley, G.H. Tuberculosis risk stratification of psoriatic patients before anti-TNF-a treatment. Immunol. Biomark. Tuberc. 2022, 12, 672894. [Google Scholar]
- Mao, L.; Xu, L.; Wang, X.; Xing, Y.; Wang, J.; Zhang, Y.; Yuan, W.; Du, J.; Shi, Z.; Ma, J. Enhanced immunogenicity of the tuberculosis subunit Rv0572c vaccine delivered in DMT liposome adjuvant as a BCG-booster. Tuberculosis 2022, 134, 102186. [Google Scholar] [CrossRef] [PubMed]
- Linnstaedt, S.D.; Gottwein, E.; Skalsky, R.L.; Luftig, M.A.; Cullen, B.R. Virally induced cellular microRNA miR-155 plays a key role in B-cell immortalization by Epstein-Barr virus. J. Virol. 2010, 84, 11670–11678. [Google Scholar] [CrossRef] [PubMed]
- Faraoni, I.; Antonetti, F.R.; Cardone, J.; Bonmassar, E. miR-155 gene: A typical multifunctional microRNA. Biochim. Biophys. Acta BBA-Mol. Basis Dis. 2009, 1792, 497–505. [Google Scholar] [CrossRef]
- Hou, L.; Chen, J.; Zheng, Y.; Wu, C. Critical role of miR-155/FoxO1/ROS axis in the regulation of non-small cell lung carcinomas. Tumor Biol. 2016, 37, 5185–5192. [Google Scholar] [CrossRef]
- Zhuang, G.; Sun, A.; Teng, M.; Luo, J. A tiny RNA that packs a big punch: The critical role of a viral miR-155 ortholog in lymphomagenesis in Marek’s disease. Front. Microbiol. 2017, 8, 1169. [Google Scholar] [CrossRef]
- Tili, E.; Croce, C.M.; Michaille, J.-J. miR-155: On the crosstalk between inflammation and cancer. Int. Rev. Immunol. 2009, 28, 264–284. [Google Scholar] [CrossRef]
- Xiao, R.; Li, C.; Chai, B. miRNA-144 suppresses proliferation and migration of colorectal cancer cells through GSPT1. Biomed. Pharmacother. 2015, 74, 138–144. [Google Scholar] [CrossRef]
- Yu, A.; Zhang, T.; Zhong, W.; Duan, H.; Wang, S.; Ye, P.; Wang, J.; Zhong, S.; Yang, Z. miRNA-144 induces microglial autophagy and inflammation following intracerebral hemorrhage. Immunol. Lett. 2017, 182, 18–23. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, C.; Gu, C.; Cui, X.; Wu, J. MiRNA-144-3p inhibits high glucose induced cell proliferation through suppressing FGF16. Biosci. Rep. 2019, 39, BSR20181788. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-X.; Rajeev, B.W.; Stromberg, A.J.; Ren, N.; Tang, G.; Huang, Q.; Rigoutsos, I.; Nelson, P.T. The expression of microRNA miR-107 decreases early in Alzheimer’s disease and may accelerate disease progression through regulation of β-site amyloid precursor protein-cleaving enzyme 1. J. Neurosci. 2008, 28, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Fix, L.N.; Shah, M.; Efferth, T.; Farwell, M.A.; Zhang, B. MicroRNA expression profile of MCF-7 human breast cancer cells and the effect of green tea polyphenon-60. Cancer Genom. Proteom. 2010, 7, 261–277. [Google Scholar]
- Carr, T.J. Expanding the Avian Micro RNA Repertoire of Domesticated Poultry and Investigating the NLRP3 Inflammasome Messenger RNA and Micro RNA Transcriptomes in Anas Platyrhynchos (Ducks). Ph.D. Thesis, University of Delaware, Newark, DE, USA, 2017. [Google Scholar]
- Wang, C.; Yang, S.; Sun, G.; Tang, X.; Lu, S.; Neyrolles, O.; Gao, Q. Comparative miRNA expression profiles in individuals with latent and active tuberculosis. PLoS ONE 2011, 6, e25832. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lu, H.; Liu, J.; Wu, S.; Kim, P.; Zhou, X. lncRNAfunc: A knowledgebase of lncRNA function in human cancer. Nucleic Acids Res. 2022, 50, D1295–D1306. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Huang, G.; Gao, M.; Huang, J.; Li, H.; Xia, H.; Xiang, X.; Wu, S.; Ruan, Y. Identification of atrial fibrillation-related lncRNA based on bioinformatic analysis. Dis. Mark. 2022, 2022, 8307975. [Google Scholar] [CrossRef]
- Chen, Q.; Lin, G.; Lin, L.; Huang, J.; Chen, L.; Lian, N.; Chen, M.; Zeng, A.; Lin, Q. LncRNA XR_596701 protects H9c2 cells against intermittent hypoxia-induced injury through regulation of the miR-344b-5p/FAIM3 axis. Cell Death Discov. 2022, 8, 42. [Google Scholar] [CrossRef]
- Yan, Q.; Tian, Y.; Hao, F. Downregulation of lncRNA UCA1 inhibits proliferation and invasion of cervical cancer cells through miR-206 expression. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2022, 28, 7–8. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Liang, L.; Fan, M.; Li, X.; Feng, N.; Pan, Y.; Tan, Q.; Xu, Q.; Xie, Y. Effect of XBP1 deficiency in cartilage on the regulatory network Of LncRNA/circRNA-miRNA-mRNA. Int. J. Biol. Sci. 2022, 18, 315. [Google Scholar] [CrossRef]
- Ansari, M.A.; Thiruvengadam, M.; Farooqui, Z.; Rajakumar, G.; Jamal, Q.M.S.; Alzohairy, M.A.; Almatroudi, A.; Alomary, M.N.; Chung, I.-M.; Al-Suhaimi, E.A. Nanotechnology, in silico and endocrine-based strategy for delivering paclitaxel and miRNA: Prospects for the therapeutic management of breast cancer. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2021; Volume 69, pp. 109–128. [Google Scholar]
- Tao, S.; Chen, Y.; Hu, M.; Xu, L.; Fu, C.-B.; Hao, X.-B. LncRNA PVT1 facilitates DLBCL development via miR-34b-5p/Foxp1 pathway. Mol. Cell. Biochem. 2022, 477, 951–963. [Google Scholar] [CrossRef]
- Zhu, X.; Wu, Y.-B.; Zhou, J.; Kang, D.-M. Upregulation of lncRNA MEG3 promotes hepatic insulin resistance via increasing FoxO1 expression. Biochem. Biophys. Res. Commun. 2016, 469, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H. Mechanism associated with aberrant lncRNA MEG3 expression in gestational diabetes mellitus. Exp. Ther. Med. 2019, 18, 3699–3706. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhong, H.; Xie, X.; Chen, C.Y.; Huang, D.; Shen, L.; Zhang, H.; Chen, Z.W.; Zeng, G. Long noncoding RNA derived from CD244 signaling epigenetically controls CD8+ T-cell immune responses in tuberculosis infection. Proc. Natl. Acad. Sci. USA 2015, 112, E3883–E3892. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.; Xu, J.; Yang, W.; You, X.; Zhong, Q.; Wang, X. The research progress of LncRNA involved in the regulation of inflammatory diseases. Mol. Immunol. 2018, 101, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Fathizadeh, H.; Hayat, S.M.G.; Dao, S.; Ganbarov, K.; Tanomand, A.; Asgharzadeh, M.; Kafil, H.S. Long non-coding RNA molecules in tuberculosis. Int. J. Biol. Macromol. 2020, 156, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wu, W.; Wang, Z.; Wang, X. lncRNA NEAT1 is closely related with progression of breast cancer via promoting proliferation and EMT. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1020–1026. [Google Scholar]
- Chakravarty, D.; Sboner, A.; Nair, S.S.; Giannopoulou, E.; Li, R.; Hennig, S.; Mosquera, J.M.; Pauwels, J.; Park, K.; Kossai, M. The oestrogen receptor alpha-regulated lncRNA NEAT1 is a critical modulator of prostate cancer. Nat. Commun. 2014, 5, 5383. [Google Scholar] [CrossRef]
- Yu, H.M.; Wang, C.; Yuan, Z.; Chen, G.L.; Ye, T.; Yang, B.W. LncRNA NEAT1 promotes the tumorigenesis of colorectal cancer by sponging miR-193a-3p. Cell Prolif. 2019, 52, e12526. [Google Scholar] [CrossRef]
- Yao, Z.; Zhang, Q.; Guo, F.; Guo, S.; Yang, B.; Liu, B.; Li, P.; Li, J.; Guan, S.; Liu, X. Long noncoding RNA PCED1B-AS1 promotes the Warburg effect and tumorigenesis by upregulating HIF-1α in glioblastoma. Cell Transplant. 2020, 29, 0963689720906777. [Google Scholar] [CrossRef]
- Li, Z.; Yu, X.; Shen, J. Long non-coding RNAs: Emerging players in osteosarcoma. Tumor Biol. 2016, 37, 2811–2816. [Google Scholar] [CrossRef]
- Li, C.; Chen, J.; Zhang, K.; Feng, B.; Wang, R.; Chen, L. Progress and prospects of long noncoding RNAs (lncRNAs) in hepatocellular carcinoma. Cell. Physiol. Biochem. 2015, 36, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Gao, J.; Wang, T.; Wang, J.; Li, H.; Wang, Z. Long non-coding RNA BANCR regulates growth and metastasis and is associated with poor prognosis in retinoblastoma. Tumor Biol. 2015, 36, 7205–7211. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, H.; Wei, Z.; Jia, Y.; Wu, Y.; Qi, X.; Li, Y.; Gao, X. The role of non-coding RNA on macrophage modification in tuberculosis infection. Microb. Pathog. 2020, 149, 104592. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Zhou, R.; Ji, C.; Liu, D.; Wu, T.; Xu, H.; Lan, D.; Yao, C.; Xu, Y.; Fang, L. The function of circRNA-0047604 in regulating the tumor suppressor gene DACH1 in breast cancer. BioMed Res. Int. 2022, 2022, 6589651. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Zhang, J.; Du, X.; Li, Q.; Pan, Z. circSLC41A1 resists porcine granulosa cell apoptosis and follicular atresia by promoting SRSF1 through miR-9820-5p sponging. Int. J. Mol. Sci. 2022, 23, 1509. [Google Scholar] [CrossRef]
- Feng, K.; Jiang, Z.; Liu, P.; Liu, J.; Wen, X.; He, L. Circular RNA, circ1-3p, is involved in cyflumetofen resistance by acting as a competitive RNA against miR-1-3p in Tetranychus cinnabarinus. J. Agric. Food Chem. 2022, 70, 1068–1078. [Google Scholar] [CrossRef]
- Zheng, W.; Su, H.; Lv, X.; Xin, S.; Xu, T. Exon–intron circular RNA circRNF217 promotes innate immunity and antibacterial activity in teleost fish by reducing miR-130-3p function. J. Immunol. 2022, 208, 1099–1114. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Li, J.; Ke, S. CircRNA CORO1C regulates miR-654-3p/USP7 axis to mediate laryngeal squamous cell carcinoma progression. Biochem. Genet. 2022, 60, 1–15. [Google Scholar] [CrossRef]
- Xia, R.-P.; Zhao, F.; Ma, T.-D.; Zou, C.-J.; Xu, G.; Zhou, C.-G. Circ-ITCH overexpression promoted cell proliferation and migration in Hirschsprung disease through miR-146b-5p/RET axis. Pediatric Res. 2022, 91, 1–9. [Google Scholar] [CrossRef]
- Foruzandeh, Z.; Zeinali-Sehrig, F.; Nejati, K.; Rahmanpour, D.; Pashazadeh, F.; Seif, F.; Alivand, M.R. CircRNAs as potent biomarkers in ovarian cancer: A systematic scoping review. Cell. Mol. Biol. Lett. 2021, 26, 41. [Google Scholar] [CrossRef]
- Yang, W.; Bai, C.; Zhang, L.; Li, Z.; Tian, Y.; Yang, Z.; Wang, L.; Wu, W. Correlation between serum circRNA and thyroid micropapillary carcinoma with cervical lymph node metastasis. Medicine 2020, 99, e23255. [Google Scholar] [CrossRef] [PubMed]
- Clerbaux, L.A.; Schultz, H.; Roman-Holba, S.; Ruan, D.F.; Yu, R.; Lamb, A.M.; Bommer, G.T.; Kennell, J.A. The microRNA miR-33 is a pleiotropic regulator of metabolic and developmental processes in Drosophila melanogaster. Dev. Dyn. 2021, 250, 1634–1650. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Xing, L.; Zheng, X.; Sun, Y.; Wang, X.; Chen, J. The circRNA circAGFG1 acts as a sponge of miR-195-5p to promote triple-negative breast cancer progression through regulating CCNE1 expression. Mol. Cancer 2019, 18, 4. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, J. CircAGFG1 promotes cervical cancer progression via miR-370-3p/RAF1 signaling. BMC Cancer 2019, 19, 1067. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.-L.; Pi, J.; Zhang, J.-A.; Yang, E.-Z.; Xu, H.; Luo, H.; Shen, L.; Peng, Y.; Liu, G.B.; Song, C.-M. Circular RNA TRAPPC6B inhibits intracellular Mycobacterium tuberculosis growth while inducing autophagy in macrophages by targeting microRNA-874-3p. Clin. Transl. Immunol. 2021, 10, e1254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, Y.; Min, Z.; Liang, G.; Mo, J.; Ju, Z.; Zeng, B.; Guan, W.; Zhang, Y.; Chen, J. circMine: A comprehensive database to integrate, analyze and visualize human disease–related circRNA transcriptome. Nucleic Acids Res. 2022, 50, D83–D92. [Google Scholar] [CrossRef]
- Di Agostino, S.; Vahabi, M.; Turco, C.; Fontemaggi, G. Secreted non-coding RNAs: Functional impact on the tumor microenvironment and clinical relevance in triple-negative breast cancer. Non-Coding RNA 2022, 8, 5. [Google Scholar] [CrossRef]
- Chen, Z.; Lin, W.; Zhao, S.; Mo, X.; Wen, Z.; Cheung, W.H.; Fu, D.; Chen, B. Identification of circRNA expression profiles in BMSCs from glucocorticoid-induced osteoporosis model. Stem Cells Int. 2022, 2022, 3249737. [Google Scholar] [CrossRef]
- Grabowska, M.; Misiorek, J.O.; Zarębska, Ż.; Rolle, K. Applications of noncoding RNAs in brain cancer patients. In Clinical Applications of Non-Coding RNAs in Cancer; Elsevier: Amsterdam, The Netherlands, 2022; pp. 17–64. [Google Scholar]
- He, S.; Dou, L.; Li, X.; Zhang, Y. Review of bioinformatics in Azheimer’s disease research. Comput. Biol. Med. 2022, 143, 105269. [Google Scholar] [CrossRef]
- Kumar, S.; Vishvakarma, N.K.; Kumar, A. Clinical applications of noncoding RNAs in lung cancer patients. In Clinical Applications of Non-Coding RNAs in Cancer; Elsevier: Amsterdam, The Netherlands, 2022; pp. 141–175. [Google Scholar]
- Burzynski, J.; Mangan, J.M.; Lam, C.K.; Macaraig, M.; Salerno, M.M.; deCastro, B.R.; Goswami, N.D.; Lin, C.Y.; Schluger, N.W.; Vernon, A. In-person vs electronic directly observed therapy for tuberculosis treatment adherence: A randomized noninferiority trial. JAMA Netw. Open 2022, 5, e2144210. [Google Scholar] [CrossRef]
- Ramakrishnan, M.; Ng, W.L.; Corbett, M.; Durcan, L.; Fitzpatrick, F. Latent tuberculosis infection amongst patients with rheumatic diseases in an Irish tertiary referral centre–A five-year review. J. Infect. 2022, 84, e18–e20. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chopra, N.; Nho, K.; Maloney, B.; Obukhov, A.G.; Nelson, P.T.; Counts, S.E.; Lahiri, D.K. Human microRNA (miR-20b-5p) modulates Alzheimer’s disease pathways and neuronal function, and a specific polymorphism close to the MIR20B gene influences Alzheimer’s biomarkers. Mol. Psychiatry 2022, 27, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kuskov, A.; Selina, O.; Kulikov, P.; Imatdinov, I.; Balysheva, V.; Kryukov, A.; Shtilman, M.; Markvicheva, E. Amphiphilic poly (N-vinylpyrrolidone) nanoparticles loaded with DNA plasmids encoding Gn and Gc glycoproteins of the Rift valley fever virus: Preparation and in vivo evaluation. ACS Appl. Bio Mater. 2021, 4, 6084–6092. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Liu, H.; Su, L.; Dawood, A.; Hu, C.; Chen, X.; Chen, H.; Chen, Y.; Guo, A. Identification of unique key miRNAs, TFs, and mRNAs in virulent MTB infection macrophages by network analysis. Int. J. Mol. Sci. 2022, 23, 382. [Google Scholar] [CrossRef]
- Chandra, P.; Coullon, H.; Agarwal, M.; Goss, C.W.; Philips, J.A. Macrophage global metabolomics identifies cholestenone as host/pathogen cometabolite present in human Mycobacterium tuberculosis infection. J. Clin. Investig. 2022, 132, e152509. [Google Scholar] [CrossRef]
- Di Marsico, M.; Paytuvi Gallart, A.; Sanseverino, W.; Aiese Cigliano, R. GreeNC 2.0: A comprehensive database of plant long non-coding RNAs. Nucleic Acids Res. 2022, 50, D1442–D1447. [Google Scholar] [CrossRef]
- Montico, B.; Giurato, G.; Pecoraro, G.; Salvati, A.; Covre, A.; Colizzi, F.; Steffan, A.; Weisz, A.; Maio, M.; Sigalotti, L. The pleiotropic roles of circular and long noncoding RNAs in cutaneous melanoma. Mol. Oncol. 2022, 16, 565. [Google Scholar] [CrossRef]
- Ross, C.J.; Ulitsky, I. Discovering functional motifs in long noncoding RNAs. Wiley Interdiscip. Rev. RNA 2022, 13, e1708. [Google Scholar] [CrossRef]
- Lan, X.; Han, J.; Wang, B.; Sun, M. Integrated analysis of transcriptome profiling of lncRNAs and mRNAs in livers of type 2 diabetes mellitus. Physiol. Genom. 2022, 54, 86–97. [Google Scholar] [CrossRef]
- Valenzuela-Muñoz, V.; Gallardo-Escárate, C.; Benavente, B.P.; Valenzuela-Miranda, D.; Núñez-Acuña, G.; Escobar-Sepulveda, H.; Váldes, J.A. Whole-genome transcript expression profiling reveals novel insights into transposon genes and non-coding RNAs during Atlantic salmon seawater adaptation. Biology 2022, 11, 1. [Google Scholar] [CrossRef]
- Anwar, S.; Almatroudi, A.; Alsahli, M.A.; Khan, M.A.; Khan, A.A.; Rahmani, A.H. Natural products: Implication in cancer prevention and treatment through modulating various biological activities. Anti-Cancer Agents Med. Chem. Former. Curr. Med. Chem. -Anti-Cancer Agents 2020, 20, 2025–2040. [Google Scholar] [CrossRef] [PubMed]
- Sharma, Y.; Sharma, A.; Singh, K.; Upadhyay, S.K. Long non-coding RNAs as emerging regulators of pathogen response in plants. Non-Coding RNA 2022, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, L.; Guo, C.; Zhang, H.; Xu, P.; Liu, S.; Hu, X.; Gao, Q. Polymorphisms of SLC11A1 (NRAMP1) rs17235409 associated with and susceptibility to spinal tuberculosis in a southern Han Chinese population. Infect. Genet. Evol. 2022, 98, 105202. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Tan, Y.; Zhang, X.; Wang, Y.; Su, B.; Xue, Z.; Wang, J.; Pang, Y. Effect of mixed infections with Mycobacterium tuberculosis and nontuberculous Mycobacteria on diagnosis of multidrug-resistant tuberculosis: A retrospective multicentre study in China. Infect. Drug Resist. 2022, 15, 157. [Google Scholar] [CrossRef] [PubMed]
- Kleinwaks, G.; Schmit, V.; Morrison, J. Considering human challenge trials for tuberculosis vaccine development. Vaccine 2022, 40, 173–174. [Google Scholar] [CrossRef]
- Wang, Z.; Posey, D.L.; Brostrom, R.J.; Morris, S.B.; Marano, N.; Phares, C.R. Post-arrival evaluation of immigrant and refugee children in the USA with latent tuberculosis infection diagnosed overseas, 2007–2019. J. Pediatrics 2022, S0022-3476(22)00080-4. in press. [Google Scholar] [CrossRef]
- Phuoc Long, N.; Heo, D.Y.; Park, S.; Thi Hai Yen, N.; Cho, Y.-S.; Shin, J.-G.; Oh, J.Y.; Kim, D.-H. Molecular perturbations in pulmonary tuberculosis patients identified by pathway-level analysis of plasma metabolic features. PLoS ONE 2022, 17, e0262545. [Google Scholar] [CrossRef]
- De Araujo, L.S.; Ribeiro-Alves, M.; Leal-Calvo, T.; Leung, J.; Durán, V.; Samir, M.; Talbot, S.; Tallam, A.; Mello, F.C.d.Q.; Geffers, R. Reprogramming of small noncoding RNA populations in peripheral blood reveals host biomarkers for latent and active Mycobacterium tuberculosis infection. MBio 2019, 10, e01037-19. [Google Scholar] [CrossRef]
- Lyu, L.; Zhang, X.; Li, C.; Yang, T.; Wang, J.; Pan, L.; Jia, H.; Li, Z.; Sun, Q.; Yue, L. Small RNA profiles of serum exosomes derived from individuals with latent and active tuberculosis. Front. Microbiol. 2019, 10, 1174. [Google Scholar] [CrossRef]
- Ndzi, E.N.; Nkenfou, C.N.; Mekue, L.M.; Zentilin, L.; Tamgue, O.; Pefura, E.W.Y.; Kuiaté, J.-R.; Giacca, M.; Ndjolo, A. MicroRNA hsa-miR-29a-3p is a plasma biomarker for the differential diagnosis and monitoring of tuberculosis. Tuberculosis 2019, 114, 69–76. [Google Scholar] [CrossRef]
- Chakrabarty, S.; Kumar, A.; Raviprasad, K.; Mallya, S.; Satyamoorthy, K.; Chawla, K. Host and MTB genome encoded miRNA markers for diagnosis of tuberculosis. Tuberculosis 2019, 116, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Alipoor, S.D.; Tabarsi, P.; Varahram, M.; Movassaghi, M.; Dizaji, M.K.; Folkerts, G.; Garssen, J.; Adcock, I.M.; Mortaz, E. Serum exosomal miRNAs are associated with active pulmonary tuberculosis. Dis. Markers 2019, 2019, 1907426. [Google Scholar] [CrossRef] [PubMed]
- Barry, S.E.; Ellis, M.; Yang, Y.; Guan, G.; Wang, X.; Britton, W.J.; Saunders, B.M. Identification of a plasma microRNA profile in untreated pulmonary tuberculosis patients that is modulated by anti-mycobacterial therapy. J. Infect. 2018, 77, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Duffy, F.J.; Thompson, E.; Downing, K.; Suliman, S.; Mayanja-Kizza, H.; Boom, W.H.; Thiel, B.; Weiner Iii, J.; Kaufmann, S.H.; Dover, D. A serum circulating miRNA signature for short-term risk of progression to active tuberculosis among household contacts. Front. Immunol. 2018, 9, 661. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, S.; Liu, C.-M.; Jiang, T.-T.; Chen, Z.-L.; Tu, H.-H.; Mao, L.-G.; Li, Z.-J.; Li, J.-C. Screening and identification of four serum miRNAs as novel potential biomarkers for cured pulmonary tuberculosis. Tuberculosis 2018, 108, 26–34. [Google Scholar] [CrossRef]
- Cui, J.-Y.; Liang, H.-W.; Pan, X.-L.; Li, D.; Jiao, N.; Liu, Y.-H.; Fu, J.; He, X.-Y.; Sun, G.-X.; Zhang, C.-L. Characterization of a novel panel of plasma microRNAs that discriminates between Mycobacterium tuberculosis infection and healthy individuals. PLoS ONE 2017, 12, e0184113. [Google Scholar] [CrossRef]
- Wang, C.; Liu, C.-M.; Wei, L.-L.; Shi, L.-Y.; Pan, Z.-F.; Mao, L.-G.; Wan, X.-C.; Ping, Z.-P.; Jiang, T.-T.; Chen, Z.-L. A group of novel serum diagnostic biomarkers for multidrug-resistant tuberculosis by iTRAQ-2D LC-MS/MS and solexa sequencing. Int. J. Biol. Sci. 2016, 12, 246. [Google Scholar] [CrossRef]
- Wagh, V.; Urhekar, A.; Modi, D. Levels of microRNA miR-16 and miR-155 are altered in serum of patients with tuberculosis and associate with responses to therapy. Tuberculosis 2017, 102, 24–30. [Google Scholar] [CrossRef]
- Zheng, M.-L.; Zhou, N.-K.; Luo, C.-H. MiRNA-155 and miRNA-132 as potential diagnostic biomarkers for pulmonary tuberculosis: A preliminary study. Microb. Pathog. 2016, 100, 78–83. [Google Scholar] [CrossRef]
- Xin, H.; Yang, Y.; Liu, J.; Li, X.; Li, M.; Feng, B.; Li, Z.; Zhang, H.; Li, H.; Shen, F. Association between tuberculosis and circulating microRNA hsa-let-7b and hsa-miR-30b: A pilot study in a Chinese population. Tuberculosis 2016, 99, 63–69. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, A.; Ni, J.; Zhang, Q.; Wang, Y.; Lu, J.; Wu, W.; Karakousis, P.C.; Lu, S.; Yao, Y. Differential expression of miRNAs and their relation to active tuberculosis. Tuberculosis 2015, 95, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Latorre, I.; Leidinger, P.; Backes, C.; Domínguez, J.; de Souza-Galvão, M.L.; Maldonado, J.; Prat, C.; Ruiz-Manzano, J.; Sánchez, F.; Casas, I. A novel whole-blood miRNA signature for a rapid diagnosis of pulmonary tuberculosis. Eur. Respir. J. 2015, 45, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, Q.; Xi, X.; Jiao, J.; Xu, W.; Huang, J.; Lai, Z. High serum miR-183 level is associated with the bioactivity of macrophage derived from tuberculosis patients. Int. J. Clin. Exp. Pathol. 2015, 8, 655. [Google Scholar] [PubMed]
- Zhang, H.; Sun, Z.; Wei, W.; Liu, Z.; Fleming, J.; Zhang, S.; Lin, N.; Wang, M.; Chen, M.; Xu, Y. Identification of serum microRNA biomarkers for tuberculosis using RNA-seq. PLoS ONE 2014, 9, e88909. [Google Scholar] [CrossRef]
- Abd-El-Fattah, A.A.; Sadik, N.A.H.; Shaker, O.G.; Aboulftouh, M.L. Differential microRNAs expression in serum of patients with lung cancer, pulmonary tuberculosis, and pneumonia. Cell Biochem. Biophys. 2013, 67, 875–884. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, J.; Fan, S.; Li, Y.; Wei, L.; Yang, X.; Jiang, T.; Chen, Z.; Wang, C.; Liu, J. Screening and identification of six serum microRNAs as novel potential combination biomarkers for pulmonary tuberculosis diagnosis. PLoS ONE 2013, 8, e81076. [Google Scholar] [CrossRef]
- Maertzdorf, J.; Weiner, J.; Mollenkopf, H.-J.; Network, T.; Bauer, T.; Prasse, A.; Müller-Quernheim, J.; Kaufmann, S.H. Common patterns and disease-related signatures in tuberculosis and sarcoidosis. Proc. Natl. Acad. Sci. USA 2012, 109, 7853–7858. [Google Scholar] [CrossRef]
- Qi, Y.; Cui, L.; Ge, Y.; Shi, Z.; Zhao, K.; Guo, X.; Yang, D.; Yu, H.; Cui, L.; Shan, Y. Altered serum microRNAs as biomarkers for the early diagnosis of pulmonary tuberculosis infection. BMC Infect. Dis. 2012, 12, 384. [Google Scholar] [CrossRef]
- Fu, Y.; Yi, Z.; Wu, X.; Li, J.; Xu, F. Circulating microRNAs in patients with active pulmonary tuberculosis. J. Clin. Microbiol. 2011, 49, 4246–4251. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, Y.; Yu, H.; Tian, R.; Wang, G.; Li, F. Identification of unique key genes and miRNAs in latent tuberculosis infection by network analysis. Mol. Immunol. 2019, 112, 103–114. [Google Scholar] [CrossRef]
- Zhuang, X.; Li, Z.; Lin, H.; Gu, L.; Lin, Q.; Lu, Z.; Tzeng, C.-M. Integrated miRNA and mRNA expression profiling to identify mRNA targets of dysregulated miRNAs in non-obstructive azoospermia. Sci. Rep. 2015, 5, 7922. [Google Scholar] [CrossRef] [PubMed]
- Van Rensburg, I.; Du Toit, L.; Walzl, G.; Du Plessis, N.; Loxton, A. Decreased neutrophil–associated miRNA and increased B-cell associated miRNA expression during tuberculosis. Gene 2018, 655, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Corral-Fernández, N.E.; Cortes-García, J.D.; Bruno, R.-S.; Romano-Moreno, S.; Medellín-Garibay, S.E.; Magaña-Aquino, M.; Salazar-González, R.A.; González-Amaro, R.; Portales-Pérez, D.P. Analysis of transcription factors, microRNAs and cytokines involved in T lymphocyte differentiation in patients with tuberculosis after directly observed treatment short-course. Tuberculosis 2017, 105, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.V.; Fernández, R.d.V.; Zoff, L.; Bongiovanni, B.; Díaz, A.; D’Attilio, L.; Santucci, N.; Alvarez, T.; Marchesini, M.M.; Bogue, C. miR-30c is specifically repressed in patients with active pulmonary tuberculosis. Tuberculosis 2017, 105, 73–79. [Google Scholar] [CrossRef]
- Pan, D.; Pan, M.; Xu, Y.-M. Mir-29a expressions in peripheral blood mononuclear cell and cerebrospinal fluid: Diagnostic value in patients with pediatric tuberculous meningitis. Brain Res. Bull. 2017, 130, 231–235. [Google Scholar] [CrossRef]
- Zhou, M.; Yu, G.; Yang, X.; Zhu, C.; Zhang, Z.; Zhan, X. Circulating microRNAs as biomarkers for the early diagnosis of childhood tuberculosis infection. Mol. Med. Rep. 2016, 13, 4620–4626. [Google Scholar] [CrossRef]
- Zheng, L.; Leung, E.; Lee, N.; Lui, G.; To, K.-F.; Chan, R.C.; Ip, M. Differential microRNA expression in human macrophages with Mycobacterium tuberculosis infection of Beijing/W and non-Beijing/W strain types. PLoS ONE 2015, 10, e0126018. [Google Scholar] [CrossRef]
- Wang, J.; Xu, J.; Han, Y.; Zhu, Y.; Zhang, W. Diagnostic values of microRNA-31 in peripheral blood mononuclear cells for pediatric pulmonary tuberculosis in Chinese patients. Genet. Mol. Res. 2015, 14, 17235–17243. [Google Scholar] [CrossRef]
- Fu, Y.; Yi, Z.; Li, J.; Li, R. Deregulated micro RNA s in CD 4+ T cells from individuals with latent tuberculosis versus active tuberculosis. J. Cell. Mol. Med. 2014, 18, 503–513. [Google Scholar] [CrossRef]
- Yi, Z.; Gao, K.; Li, R.; Fu, Y. Changed immune and miRNA response in RAW264. 7 cells infected with cell wall deficient Mycobacterium tuberculosis. Int. J. Mol. Med. 2018, 41, 2885–2892. [Google Scholar]
- Spinelli, S.V.; Diaz, A.; D’Attilio, L.; Marchesini, M.M.; Bogue, C.; Bay, M.L.; Bottasso, O.A. Altered microRNA expression levels in mononuclear cells of patients with pulmonary and pleural tuberculosis and their relation with components of the immune response. Mol. Immunol. 2013, 53, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Tamgue, O.; Mezajou, C.F.; Ngongang, N.N.; Kameni, C.; Ngum, J.A.; Simo, U.S.F.; Tatang, F.J.; Akami, M.; Ngono, A.N. Non-coding RNAs in the etiology and control of major and neglected human tropical diseases. Front. Immunol. 2021, 12, 703936. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Yan, C.; Zhang, L.; Li, Y.; Wan, Q. LncRNA ENST00113 promotes proliferation, survival, and migration by activating PI3K/Akt/mTOR signaling pathway in atherosclerosis. Medicine 2018, 97, e0473. [Google Scholar] [CrossRef] [PubMed]
- Lungu, P.; Njelesani, E.; Sukwa, T.; Ngalamika, O.; Munsaka, S.; Kilembe, W.; Lakhi, S.; Mwaba, P. Immune correlates of Mycobacterium tuberculosis patients in Zambia stratified by HIV serostatus and level of immunity-a cross-sectional analytical laboratory based study. PLoS ONE 2022, 17, e0262454. [Google Scholar] [CrossRef]
- Swain, S.S.; Pati, S.; Hussain, T. Quinoline heterocyclic containing plant and marine candidates against drug-resistant Mycobacterium tuberculosis: A systematic drug-ability investigation. Eur. J. Med. Chem. 2022, 232, 114173. [Google Scholar] [CrossRef]
- Zonghai, C.; Tao, L.; Pengjiao, M.; Liang, G.; Rongchuan, Z.; Xinyan, W.; Wenyi, N.; Wei, L.; Yi, W.; Lang, B. Mycobacterium tuberculosis ESAT6 modulates host innate immunity by downregulating miR-222-3p target PTEN. Biochim. Biophys. Acta BBA-Mol. Basis Dis. 2022, 1868, 166292. [Google Scholar] [CrossRef]
- Zhuang, Z.-G.; Zhang, J.-A.; Luo, H.-L.; Liu, G.-B.; Lu, Y.-B.; Ge, N.-H.; Zheng, B.-Y.; Li, R.X.; Chen, C.; Wang, X. The circular RNA of peripheral blood mononuclear cells: Hsa_circ_0005836 as a new diagnostic biomarker and therapeutic target of active pulmonary tuberculosis. Mol. Immunol. 2017, 90, 264–272. [Google Scholar] [CrossRef]
- Qian, Z.; Liu, H.; Li, M.; Shi, J.; Li, N.; Zhang, Y.; Zhang, X.; Lv, J.; Xie, X.; Bai, Y. Potential diagnostic power of blood circular RNA expression in active pulmonary tuberculosis. EBioMedicine 2018, 27, 18–26. [Google Scholar] [CrossRef]
- Sun, Y.; Jiang, X.; Lv, Y.; Liang, X.; Zhao, B.; Bian, W.; Zhang, D.; Jiang, J.; Zhang, C. Circular RNA expression profiles in plasma from patients with heart failure related to platelet activity. Biomolecules 2020, 10, 187. [Google Scholar] [CrossRef]
- Micheni, L.N.; Kassaza, K.; Kinyi, H.; Ntulume, I.; Bazira, J. Detection of Mycobacterium tuberculosis multiple strains in sputum samples from patients with pulmonary tuberculosis in south western Uganda using MIRU-VNTR. Sci. Rep. 2022, 12, 1656. [Google Scholar] [CrossRef]
- Verma, A.K.; Bharti, P.S.; Rafat, S.; Bhatt, D.; Goyal, Y.; Pandey, K.K.; Ranjan, S.; Almatroodi, S.A.; Alsahli, M.A.; Rahmani, A.H. Autophagy paradox of cancer: Role, regulation, and duality. Oxidative Med. Cell. Longev. 2021, 2021, 8832541. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zhang, L.; Liang, J.; Li, X.; Yang, Y.; Sun, W.; Hou, J. Comparison of clinical and imaging features between pulmonary tuberculosis complicated with lung cancer and simple pulmonary tuberculosis: A systematic review and meta-analysis. Epidemiol. Infect. 2022, 150, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Singer, S.N.; Ndumnego, O.C.; Kim, R.S.; Ndung’u, T.; Anastos, K.; French, A.; Churchyard, G.; Paramithiothis, E.; Kasprowicz, V.O.; Achkar, J.M. Plasma host protein biomarkers correlating with increasing Mycobacterium tuberculosis infection activity prior to tuberculosis diagnosis in people living with HIV. EBioMedicine 2022, 75, 103787. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, K.; Ranjbar, M.; Pirpour Tazehkand, A.; Asgharian, P.; Montazersaheb, S.; Tarhriz, V.; Ghasemnejad, T. Evaluation of exosomal non-coding RNAs in cancer using high-throughput sequencing. J. Transl. Med. 2022, 20, 30. [Google Scholar] [CrossRef] [PubMed]
- Toden, S.; Goel, A. Non-coding RNAs as liquid biopsy biomarkers in cancer. Br. J. Cancer 2022, 126, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Diamantopoulos, M.A.; Georgoulia, K.K.; Scorilas, A. Identification and expression analysis of ten novel small non-coding RNAs (sncRNAs) in cancer cells using a high-throughput sequencing approach. Gene 2022, 809, 146025. [Google Scholar] [CrossRef] [PubMed]
- Khamina, K.; Diendorfer, A.B.; Skalicky, S.; Weigl, M.; Pultar, M.; Krammer, T.L.; Fournier, C.A.; Schofield, A.L.; Otto, C.; Smith, A.T. A MicroRNA next-generation-sequencing discovery assay (miND) for genome-scale analysis and absolute quantitation of circulating microRNA biomarkers. Int. J. Mol. Sci. 2022, 23, 1226. [Google Scholar] [CrossRef]
- Lee, J.; Kang, H. Role of microRNAs and long non-coding RNAs in sarcopenia. Cells 2022, 11, 187. [Google Scholar] [CrossRef]
- Shah, P.; Chen, C.-S. Systematic screening of penetratin’s protein targets by yeast proteome microarrays. Int. J. Mol. Sci. 2022, 23, 712. [Google Scholar] [CrossRef]
- Han, X.; Cai, L.; Shi, Y.; Hua, Z.; Lu, Y.; Li, D.; Yang, J. Integrated analysis of long non-coding RNA-mRNA profile and validation in diabetic cataract. Curr. Eye Res. 2022, 47, 382–390. [Google Scholar] [CrossRef]
- Wang, H.; Gao, Y.; Vafaei, S.; Yu, Q.; Zhang, J.; Wang, L. A chemoresistance lncRNA signature for recurrence risk stratification of colon cancer patients with chemotherapy. Mol. Ther. Nucleic Acids 2022, 27, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Baaklini, I.; Hraiky, C.; Rallu, F.; Tse-Dinh, Y.C.; Drolet, M. RNase HI overproduction is required for efficient full-length RNA synthesis in the absence of topoisomerase I in Escherichia coli. Mol. Microbiol. 2004, 54, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Piredda, R.; Mottola, A.; Cipriano, G.; Carlucci, R.; Ciccarese, G.; Di Pinto, A. Next generation sequencing (NGS) approach applied to species identification in mixed processed seafood products. Food Control 2022, 133, 108590. [Google Scholar] [CrossRef]
- Harris, C.M.; Foley, S.E.; Goedken, E.R.; Michalak, M.; Murdock, S.; Wilson, N.S. Merits and pitfalls in the characterization of covalent inhibitors of Bruton’s tyrosine kinase. Slas Discov. Adv. Life Sci. R&D 2018, 23, 1040–1050. [Google Scholar]
- Gutiérrez-García, C.; Ahmed, S.S.; Ramalingam, S.; Selvaraj, D.; Srivastava, A.; Paul, S.; Sharma, A. Identification of microRNAs from medicinal plant Murraya koenigii by high-throughput sequencing and their functional implications in secondary metabolite biosynthesis. Plants 2022, 11, 46. [Google Scholar] [CrossRef]
- Cheng, S.; Zhu, B.; Luo, F.; Lin, X.; Sun, C.; You, Y.; Yi, C.; Xu, B.; Wang, J.; Lu, Y. Comparative transcriptome profiles of Schistosoma japonicum larval stages: Implications for parasite biology and host invasion. PLoS Negl. Trop. Dis. 2022, 16, e0009889. [Google Scholar] [CrossRef]
- Gibb, E.A.; Vucic, E.A.; Enfield, K.S.; Stewart, G.L.; Lonergan, K.M.; Kennett, J.Y.; Becker-Santos, D.D.; MacAulay, C.E.; Lam, S.; Brown, C.J. Human cancer long non-coding RNA transcriptomes. PLoS ONE 2011, 6, e25915. [Google Scholar] [CrossRef]
- Xu, W.; Yang, W.; Zhang, Y.; Chen, Y.; Zhang, Q.; Wang, X.; Song, K.; Jin, W.; Chen, X. ISSAAC-seq enables sensitive and flexible multimodal profiling of chromatin accessibility and gene expression in single cells. bioRxiv 2022. [Google Scholar] [CrossRef]
- Withanage, M.H.H.; Liang, H.; Zeng, E. RNA-Seq experiment and data analysis. Methods Mol. Biol. 2022, 2418, 405–424. [Google Scholar]
- De Paoli-Iseppi, R.; Joshi, S.; Wrzesinski, T.; Harrison, P.J.; Haerty, W.; Tunbridge, E.M.; Clark, M.B. Using long-read RNA sequencing to decipher the role of RNA isoforms in disease. Pathology 2022, 54, S17. [Google Scholar] [CrossRef]
- Arnvig, K.; Young, D. Non-coding RNA and its potential role in Mycobacterium tuberculosis pathogenesis. RNA Biol. 2012, 9, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Park, H.; Kim, S.J.; Lee, K.W.; Son, J.K.; Hong, J.H.; Kim, S.H.; Cho, H.J.; Park, J.B.; Kim, T.M. Circulating RNA profiling in postreperfusion plasma from kidney transplant recipients. In Transplantation Proceedings; Elsevier: Amsterdam, The Netherlands, 2021; Volume 53, pp. 2853–2865. [Google Scholar]
- Cong, L.; Yang, Q.; Hu, C.; Yu, Q.; Hao, S.; Li, D. Current status of functional studies on circular RNAs in bladder cancer and their potential role as diagnostic and prognostic biomarkers: A review. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 3425. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Lin, L.; Chen, J. scDIOR: Single cell RNA-seq data IO software. BMC Bioinform. 2022, 23, 16. [Google Scholar] [CrossRef] [PubMed]
- Dwight, Z. Data Innovation Provides a Smooth Road to Production: Bioinformatics Needs to Accelerate; Oxford University Press: Oxford, UK, 2022; Volume 68, pp. 264–265. [Google Scholar]
- Yu, C.; Qi, X.; Yan, W.; Wu, W.; Shen, B. Next-generation sequencing markup language (NGSML): A medium for the representation and exchange of NGS data. IEEE/ACM Trans. Comput. Biol. Bioinform. 2022, 19, 1. [Google Scholar] [CrossRef]
- Cheng, J.; Lin, Y.; Xu, L.; Chen, K.; Li, Q.; Xu, K.; Ning, L.; Kang, J.; Cui, T.; Huang, Y. ViRBase v3. 0: A virus and host ncRNA-associated interaction repository with increased coverage and annotation. Nucleic Acids Res. 2022, 50, D928–D933. [Google Scholar] [CrossRef]
- Federico, A.; Saarimäki, L.A.; Serra, A.; Giudice, G.d.; Kinaret, P.A.S.; Scala, G.; Greco, D. Microarray data preprocessing: From experimental design to differential analysis. In Microarray Data Analysis; Springer: Berlin, Germany, 2022; pp. 79–100. [Google Scholar]
- Kerachian, M.A. Noncoding RNAs in patients with colorectal cancer. In Clinical Applications of Non-Coding RNAs in Cancer; Elsevier: Amsterdam, The Netherlands, 2022; pp. 65–95. [Google Scholar]
- Schon, M.A.; Lutzmayer, S.; Hofmann, F.; Nodine, M.D. Precise transcript reconstruction with end-guided assembly. bioRxiv 2022. [Google Scholar] [CrossRef]
- Bahonar, E.; Chahardowli, M.; Ghalenoei, Y.; Simjoo, M. New correlations to predict oil viscosity using data mining techniques. J. Pet. Sci. Eng. 2022, 208, 109736. [Google Scholar] [CrossRef]
- Naphade, S.; Bhatnagar, R.; Hanson-Smith, V.; Choi, I.; Zhang, A. Systematic comparative analysis of strand-specific RNA-seq library preparation methods for low input samples. Sci. Rep. 2022, 12, 1789. [Google Scholar] [CrossRef]
- Taban, P.T.M.A.Q.; Bhat, B.; Ahmad, S.M.; Dar, M.A.; Kashoo, Z.A.; Ganie, N.A.; Shah, R.A. Expression of lncRNAs in response to bacterial infections of goat mammary epithelial cells reveals insights into mammary gland diseases. Microb. Pathog. 2022, 162, 105367. [Google Scholar]
- Pucci, P. Combination therapy and noncoding RNAs: A new era of cancer personalized medicine. Future Med. 2022, 14, 117–120. [Google Scholar] [CrossRef]
- Sridhar, J.; Gunasekaran, P. Computational small RNA prediction in bacteria. Bioinform. Biol. Insights 2013, 7, BBI-S11213. [Google Scholar] [CrossRef] [PubMed]
- Ngabonziza, J.C.S.; Rigouts, L.; Torrea, G.; Decroo, T.; Kamanzi, E.; Lempens, P.; Rucogoza, A.; Habimana, Y.M.; Laenen, L.; Niyigena, B.E. Multidrug-resistant tuberculosis control in Rwanda overcomes a successful clone that causes most disease over a quarter century. J. Clin. Tuberc. Other Mycobact. Dis. 2022, 27, 100299. [Google Scholar] [CrossRef] [PubMed]
- Hackenberg, M.; Sturm, M.; Langenberger, D.; Falcon-Perez, J.M.; Aransay, A.M. miRanalyzer: A microRNA detection and analysis tool for next-generation sequencing experiments. Nucleic Acids Res. 2009, 37, W68–W76. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, Q.; Wang, X.; Zheng, J.; Wang, T.; You, M.; Sheng Sun, Z.; Shi, Q. mirTools 2.0 for non-coding RNA discovery, profiling, and functional annotation based on high-throughput sequencing. RNA Biol. 2013, 10, 1087–1092. [Google Scholar] [CrossRef]
- Bonnal, R.J.; Rossi, R.L.; Carpi, D.; Ranzani, V.; Abrignani, S.; Pagani, M. miRiadne: A web tool for consistent integration of miRNA nomenclature. Nucleic Acids Res. 2015, 43, W487–W492. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Georgakilas, G.; Vlachos, I.S.; Zagganas, K.; Vergoulis, T.; Paraskevopoulou, M.D.; Kanellos, I.; Tsanakas, P.; Dellis, D.; Fevgas, A.; Dalamagas, T. DIANA-miRGen v3. 0: Accurate characterization of microRNA promoters and their regulators. Nucleic Acids Res. 2016, 44, D190–D195. [Google Scholar] [CrossRef]
- Manual, I. miRStar™ Human Cancer Focus miRNA & Target mRNA PCR Array; Instruction Manual Version 1.0; Arrastay, Inc.: Rockville, MD, USA, 2020. [Google Scholar]
- Gkirtzou, K.; Tsamardinos, I.; Tsakalides, P.; Poirazi, P. MatureBayes: A probabilistic algorithm for identifying the mature miRNA within novel precursors. PLoS ONE 2010, 5, e11843. [Google Scholar] [CrossRef]
- Schulhofer, S.J. Reconsidering Miranda. Univ. Chic. Law Rev. 1987, 54, 435–461. [Google Scholar] [CrossRef]
- Volders, P.-J.; Helsens, K.; Wang, X.; Menten, B.; Martens, L.; Gevaert, K.; Vandesompele, J.; Mestdagh, P. LNCipedia: A database for annotated human lncRNA transcript sequences and structures. Nucleic Acids Res. 2013, 41, D246–D251. [Google Scholar] [CrossRef]
- Ma, L.; Cao, J.; Liu, L.; Du, Q.; Li, Z.; Zou, D.; Bajic, V.B.; Zhang, Z. LncBook: A curated knowledgebase of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, D128–D134. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ma, R.; Ma, W.; Chen, J.; Yang, J.; Xi, Y.; Cui, Q. LncDisease: A sequence based bioinformatics tool for predicting lncRNA-disease associations. Nucleic Acids Res. 2016, 44, e90. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, H.; Fang, S.; Kang, Y.; Wu, W.; Hao, Y.; Li, Z.; Bu, D.; Sun, N.; Zhang, M.Q. NONCODE 2016: An informative and valuable data source of long non-coding RNAs. Nucleic Acids Res. 2016, 44, D203–D208. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-H.; Liu, S.; Zhou, H.; Qu, L.-H.; Yang, J.-H. starBase v2. 0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein–RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef]
- Dinger, M.E.; Pang, K.C.; Mercer, T.R.; Crowe, M.L.; Grimmond, S.M.; Mattick, J.S. NRED: A database of long noncoding RNA expression. Nucleic Acids Res. 2009, 37, D122–D126. [Google Scholar] [CrossRef]
- Ghosal, S.; Das, S.; Sen, R.; Basak, P.; Chakrabarti, J. Circ2Traits: A comprehensive database for circular RNA potentially associated with disease and traits. Front. Genet. 2013, 4, 283. [Google Scholar] [CrossRef]
- Chen, X.; Han, P.; Zhou, T.; Guo, X.; Song, X.; Li, Y. circRNADb: A comprehensive database for human circular RNAs with protein-coding annotations. Sci. Rep. 2016, 6, 34985. [Google Scholar] [CrossRef]
- Glažar, P.; Papavasileiou, P.; Rajewsky, N. circBase: A database for circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef]
- Jakobi, T.; Uvarovskii, A.; Dieterich, C. Circtools—A one-stop software solution for circular RNA research. Bioinformatics 2019, 35, 2326–2328. [Google Scholar] [CrossRef]
- Li, X.; Chu, C.; Pei, J.; Măndoiu, I.; Wu, Y. CircMarker: A fast and accurate algorithm for circular RNA detection. BMC Genom. 2018, 19, 79–87. [Google Scholar] [CrossRef]
- Monteiro, S.; Rente, D.; Cunha, M.V.; Marques, T.A.; Cardoso, E.; Vilaça, J.; Coelho, N.; Brôco, N.; Carvalho, M.; Santos, R. Discrimination and surveillance of infectious severe acute respiratory syndrome Coronavirus 2 in wastewater using cell culture and RT-qPCR. Sci. Total Environ. 2022, 815, 152914. [Google Scholar] [CrossRef]
- Lee, M.S.; Hyun, H.; Park, I.; Kim, S.; Jang, D.H.; Kim, S.; Im, J.K.; Kim, H.; Lee, J.H.; Kwon, T. Quantitative fluorescence in situ hybridization (FISH) of magnetically confined bacteria enables early detection of human bacteremia. Small Methods 2022, 6, 2101239. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Analysis of RNA by northern blotting. Cold Spring Harb. Protoc. 2022, 2022, 101741. [Google Scholar] [CrossRef] [PubMed]
- Stiens, J.; Arnvig, K.B.; Kendall, S.L.; Nobeli, I. Challenges in defining the functional, non-coding, expressed genome of members of the Mycobacterium tuberculosis complex. Mol. Microbiol. 2022, 117, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Duarte, I.; Carraco, G.; de Azevedo, N.T.; Benes, V.; Andrade, R.P. gga-miRNOME, a microRNA-sequencing dataset from chick embryonic tissues. Sci. Data 2022, 9, 29. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Preparation of labeled DNA, RNA, and oligonucleotide probes. Cold Spring Harb. Protoc. 2022, 2022, 100578. [Google Scholar] [CrossRef]
- Torii, S.; Oishi, W.; Zhu, Y.; Thakali, O.; Malla, B.; Yu, Z.; Zhao, B.; Arakawa, C.; Kitajima, M.; Hata, A. Comparison of five polyethylene glycol precipitation procedures for the RT-qPCR based recovery of murine hepatitis virus, bacteriophage phi6, and pepper mild mottle virus as a surrogate for SARS-CoV-2 from wastewater. Sci. Total Environ. 2022, 807, 150722. [Google Scholar] [CrossRef]
- Taylor, S.; Wakem, M.; Dijkman, G.; Alsarraj, M.; Nguyen, M. A practical approach to RT-qPCR—Publishing data that conform to the MIQE guidelines. Methods 2010, 50, S1–S5. [Google Scholar] [CrossRef]
- Mestdagh, P.; Van Vlierberghe, P.; De Weer, A.; Muth, D.; Westermann, F.; Speleman, F.; Vandesompele, J. A novel and universal method for microRNA RT-qPCR data normalization. Genome Biol. 2009, 10, R64. [Google Scholar] [CrossRef]
- Levsky, J.M.; Singer, R.H. Fluorescence in situ hybridization: Past, present and future. J. Cell Sci. 2003, 116, 2833–2838. [Google Scholar] [CrossRef]
- Allemailem, K.S.; Almatroudi, A.; Alrumaihi, F.; Almansour, N.M.; Aldakheel, F.M.; Rather, R.A.; Afroze, D.; Rah, B. Single nucleotide polymorphisms (SNPs) in prostate cancer: Its implications in diagnostics and therapeutics. Am. J. Transl. Res. 2021, 13, 3868. [Google Scholar] [PubMed]
- Nederlof, P.; Van der Flier, S.; Wiegant, J.; Raap, A.; Tanke, H.; Ploem, J.; Van der Ploeg, M. Multiple fluorescence in situ hybridization. Cytom. J. Int. Soc. Anal. Cytol. 1990, 11, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Liehr, T. Fluorescence In Situ Hybridization (FISH); Springer: Berlin, Germany, 2017. [Google Scholar]
- Trask, B.J. Fluorescence in situ hybridization: Applications in cytogenetics and gene mapping. Trends Genet. 1991, 7, 149–154. [Google Scholar] [CrossRef]
- Gonzalez Plaza, J.J. Current roles of microRNAs in infectious diseases—Advancing into healthcare. Infektološki Glas. 2016, 36, 5–15. [Google Scholar]
- Lalevée, S.; Feil, R. Long noncoding RNAs in human disease: Emerging mechanisms and therapeutic strategies. Epigenomics 2015, 7, 877–879. [Google Scholar] [CrossRef]
- Grijalvo, S.; Alagia, A.; Jorge, A.F.; Eritja, R. Covalent strategies for targeting messenger and non-coding RNAs: An updated review on siRNA, miRNA and antimiR conjugates. Genes 2018, 9, 74. [Google Scholar] [CrossRef]
- Allemailem, K.S.; Almatroudi, A.; Alsahli, M.A.; Basfar, G.T.; Alrumaihi, F.; Rahmani, A.H.; Khan, A.A. Recent advances in understanding oligonucleotide aptamers and their applications as therapeutic agents. 3Biotech 2020, 10, 551. [Google Scholar] [CrossRef]
- Matsui, M.; Corey, D.R. Non-coding RNAs as drug targets. Nat. Rev. Drug Discov. 2017, 16, 167–179. [Google Scholar] [CrossRef]
- Corrêa, R.L.; Steiner, F.A.; Berezikov, E.; Ketting, R.F. MicroRNA–directed siRNA biogenesis in Caenorhabditis elegans. PLoS Genet. 2010, 6, e1000903. [Google Scholar] [CrossRef]
- Dykxhoorn, D.M.; Lieberman, J. Running interference: Prospects and obstacles to using small interfering RNAs as small molecule drugs. Annu. Rev. Biomed. Eng. 2006, 8, 377–402. [Google Scholar] [CrossRef]
- Castel, S.E.; Martienssen, R.A. RNA interference in the nucleus: Roles for small RNAs in transcription, epigenetics and beyond. Nat. Rev. Genet. 2013, 14, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Wang, Y.; Li, H.; Chen, L.; Liu, Q. Regulatory networks of LncRNA MALAT-1 in cancer. Cancer Manag. Res. 2020, 12, 10181. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Doudna, J.A. CRISPR–Cas9 structures and mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef]
- Tang, L.; Zeng, Y.; Du, H.; Gong, M.; Peng, J.; Zhang, B.; Lei, M.; Zhao, F.; Wang, W.; Li, X. CRISPR/Cas9-mediated gene editing in human zygotes using Cas9 protein. Mol. Genet. Genom. 2017, 292, 525–533. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Peng, R.; Lin, G.; Li, J. Potential pitfalls of CRISPR/Cas9-mediated genome editing. FEBS J. 2016, 283, 1218–1231. [Google Scholar] [CrossRef]
- Liu, L.; Zhan, Y.; Huang, Y.; Huang, L. LncRNA FGD5-AS1 can be predicted as therapeutic target in oral cancer. J. Oral Pathol. Med. 2020, 49, 243–252. [Google Scholar] [CrossRef]
- Mei, Y.; Wang, Y.; Chen, H.; Sun, Z.S.; Ju, X.-D. Recent progress in CRISPR/Cas9 technology. J. Genet. Genom. 2016, 43, 63–75. [Google Scholar] [CrossRef]
- Zhou, J.; Yuan, M.; Zhao, Y.; Quan, Q.; Yu, D.; Yang, H.; Tang, X.; Xin, X.; Cai, G.; Qian, Q. Efficient deletion of multiple circle RNA loci by CRISPR-Cas9 reveals Os06circ02797 as a putative sponge for OsMIR408 in rice. Plant Biotechnol. J. 2021, 19, 1240–1252. [Google Scholar] [CrossRef]
- Zhang, Y.; Nguyen, T.M.; Zhang, X.-O.; Wang, L.; Phan, T.; Clohessy, J.G.; Pandolfi, P.P. Optimized RNA-targeting CRISPR/Cas13d technology outperforms shRNA in identifying functional circRNAs. Genome Biol. 2021, 22, 41. [Google Scholar] [CrossRef] [PubMed]




| Tool Name | Description | References |
|---|---|---|
| miRanalyzer | miRNA detection tool for NGS experiments | [224] |
| miRTools | Toolbox for miRNA discovery and profiling | [225] |
| miRiadne | Tool for integrating the miRNA nomenclature | [226] |
| miRBase | Database of miRNA sequence and annotation | [227] |
| DIANA-mirGen | A tool to index promoters and regulator for miRNA | [228] |
| miRStart | miRNA’s Transcription Start Sites database | [229] |
| miRWalk 2.0 | Tool to predict miRNA-target interaction using Artificial intelligence algorithm | [229] |
| MatureBayes | miRNA-target prediction tool | [230] |
| miRanda | Tool to predict miRNA target using free energy feature | [231] |
| LNCipedia | lncRNA database | [232] |
| LNCBook | lncRNA database | [233] |
| LncDisease | lncRNA-disease associations predicting tool | [234] |
| NONCODE | Integrated knowledge database for ncRNA research | [235] |
| StarBase v2.0 | Tool to predict miRNA-ceRNA interaction | [236] |
| NRED | Database for expression information of lncRNAs | [237] |
| Circ2Traits | Toolbox for circularRNA discovery and analysis | [238] |
| CircRNABase | Tool to predict miRNA-circRNA interaction | [239] |
| circBase | circularRNA database | [240] |
| Circtools | Toolbox for circularRNA research | [241] |
| CircMarker | Tool for circularRNA detection | [242] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almatroudi, A. Non-Coding RNAs in Tuberculosis Epidemiology: Platforms and Approaches for Investigating the Genome’s Dark Matter. Int. J. Mol. Sci. 2022, 23, 4430. https://doi.org/10.3390/ijms23084430
Almatroudi A. Non-Coding RNAs in Tuberculosis Epidemiology: Platforms and Approaches for Investigating the Genome’s Dark Matter. International Journal of Molecular Sciences. 2022; 23(8):4430. https://doi.org/10.3390/ijms23084430
Chicago/Turabian StyleAlmatroudi, Ahmad. 2022. "Non-Coding RNAs in Tuberculosis Epidemiology: Platforms and Approaches for Investigating the Genome’s Dark Matter" International Journal of Molecular Sciences 23, no. 8: 4430. https://doi.org/10.3390/ijms23084430
APA StyleAlmatroudi, A. (2022). Non-Coding RNAs in Tuberculosis Epidemiology: Platforms and Approaches for Investigating the Genome’s Dark Matter. International Journal of Molecular Sciences, 23(8), 4430. https://doi.org/10.3390/ijms23084430