
Novel Biomolecules in the Pathogenesis of Gestational Diabetes Mellitus 2.0

 , ,
, ,  ,
,
Abstract
:
1. Introduction
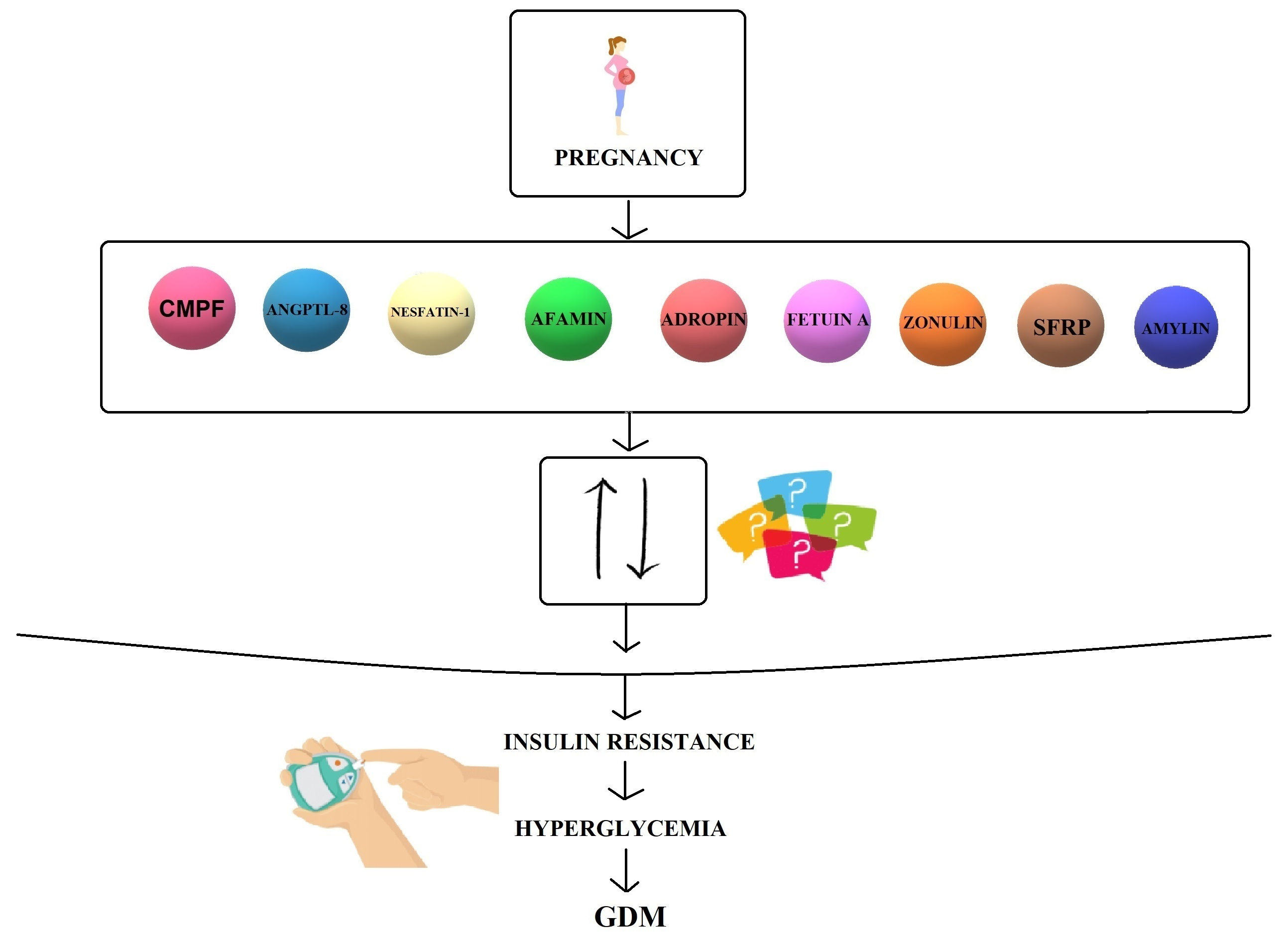
2. Biomolecules
2.1. CMPF
2.2. ANGPTL-8
2.3. Nesfatin-1
2.4. Afamin
2.5. Adropin
2.6. Fetuin-A
2.7. Zonulin
2.8. SFRPs
2.9. Amylin
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vince, K.; Perković, P.; Matijević, R. What is known and what remains unresolved regarding gestational diabetes mellitus (GDM). J. Perinat. Med. 2020, 48, 757–763. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, H.D.; Kapur, A.; Divakar, H.; Hod, M. Gestational Diabetes Mellitus—Innovative Approach to Prediction, Diagnosis, Management, and Prevention of Future NCD—Mother and Offspring. Front. Endocrinol. 2020, 11, 614533. [Google Scholar] [CrossRef] [PubMed]
- Kimber-Trojnar, Ż.; Marciniak, B.; Patro-Małysza, J.; Skórzyńska-Dziduszko, K.; Poniedziałek-Czajkowska, E.; Mierzyński, R.; Gałczynski, K.; Trojnar, M.; Leszczyńska-Gorzelak, B.; Oleszczuk, J. Is glyburide safe in pregnancy? Curr. Pharm. Biotechnol. 2014, 15, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Thanawala, U.; Divakar, H.; Jain, R.; Agarwal, M.M. Negotiating Gestational Diabetes Mellitus in India: A National Approach. Medicina 2021, 57, 942. [Google Scholar] [CrossRef]
- Alejandro, E.U.; Mamerto, T.P.; Chung, G.; Villavieja, A.; Gaus, N.L.; Morgan, E.; Pineda-Cortel, M.R.B. Gestational Diabetes Mellitus: A Harbinger of the Vicious Cycle of Diabetes. Int. J. Mol. Sci. 2020, 21, 5003. [Google Scholar] [CrossRef]
- Skórzyńska-Dziduszko, K.E.; Kimber-Trojnar, Ż.; Patro-Małysza, J.; Stenzel-Bembenek, A.; Oleszczuk, J.; Leszczyńska-Gorzelak, B. Heat Shock Proteins as a Potential Therapeutic Target in the Treatment of Gestational Diabetes Mellitus: What We Know so Far. Int. J. Mol. Sci. 2018, 19, 3205. [Google Scholar] [CrossRef] [Green Version]
- de Mendonça, E.L.S.S.; Fragoso, M.B.T.; de Oliveira, J.M.; Xavier, J.A.; Goulart, M.O.F.; de Oliveira, A.C.M. Gestational Diabetes Mellitus: The Crosslink among Inflammation, Nitroxidative Stress, Intestinal Microbiota and Alternative Therapies. Antioxidants 2022, 11, 129. [Google Scholar] [CrossRef]
- Plows, J.F.; Stanley, J.L.; Baker, P.N.; Reynolds, C.M.; Vickers, M.H. The Pathophysiology of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 3342. [Google Scholar] [CrossRef] [Green Version]
- Ramezani Tehrani, F.; Naz, M.S.G.; Yarandi, R.B.; Behboudi-Gandevani, S. The Impact of Diagnostic Criteria for Gestational Diabetes Mellitus on Adverse Maternal Outcomes: A Systematic Review and Meta-Analysis. J. Clin. Med. 2021, 10, 666. [Google Scholar] [CrossRef]
- Filardi, T.; Tavaglione, F.; Di Stasio, M.; Fazio, V.; Lenzi, A.; Morano, S. Impact of risk factors for gestational diabetes (GDM) on pregnancy outcomes in women with GDM. J. Endocrinol. Investig. 2018, 41, 671–676. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, C.M.; Zhang, Y.; Chen, Q.; Zhang, X.Q.; Li, X.F.; Shao, R.Y.; Gao, Y.M. Factors Associated with Gestational Diabetes Mellitus: A Meta-Analysis. J. Diabetes Res. 2021, 2021, 6692695. [Google Scholar] [CrossRef]
- US Preventive Services Task Force; Davidson, K.W.; Barry, M.J.; Mangione, C.M.; Cabana, M.; Caughey, A.B.; Davis, E.M.; Donahue, K.E.; Doubeni, C.A.; Kubik, M.; et al. Screening for Gestational Diabetes: US Preventive Services Task Force Recommendation Statement. JAMA 2021, 326, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, A.; Patro-Małysza, J.; Kimber-Trojnar, Ż.; Marciniak, B.; Oleszczuk, J.; Leszczyńska-Gorzelak, B. Fetal programming of the metabolic syndrome. Taiwan. J. Obstet. Gynecol. 2017, 56, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Wojczakowski, W.; Kimber-Trojnar, Ż.; Dziwisz, F.; Słodzińska, M.; Słodziński, H.; Leszczyńska-Gorzelak, B. Preeclampsia and Cardiovascular Risk for Offspring. J. Clin. Med. 2021, 10, 3154. [Google Scholar] [CrossRef] [PubMed]
- Patro-Małysza, J.; Trojnar, M.; Skórzyńska-Dziduszko, K.E.; Kimber-Trojnar, Ż.; Darmochwał-Kolarz, D.; Czuba, M.; Leszczyńska-Gorzelak, B. Leptin and Ghrelin in Excessive Gestational Weight Gain-Association between Mothers and Offspring. Int. J. Mol. Sci. 2019, 20, 2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ornoy, A.; Becker, M.; Weinstein-Fudim, L.; Ergaz, Z. Diabetes during Pregnancy: A Maternal Disease Complicating the Course of Pregnancy with Long-Term Deleterious Effects on the Offspring. A Clinical Review. Int. J. Mol. Sci. 2021, 22, 2965. [Google Scholar] [CrossRef]
- Skórzyńska-Dziduszko, K.E.; Kimber-Trojnar, Ż.; Patro-Małysza, J.; Olszewska, A.; Zaborowski, T.; Małecka-Massalska, T. An Interplay between Obesity and Inflammation in Gestational Diabetes Mellitus. Curr. Pharm. Biotechnol. 2016, 17, 603–613. [Google Scholar] [CrossRef]
- Patro-Małysza, J.; Kimber-Trojnar, Ż.; Skorzyńska-Dziduszko, K.; Marciniak, B.; Darmochwał-Kolarz, D.; Bartosiewicz, J.; Leszczyńska-Gorzelak, B.; Oleszczuk, J. The impact of substance P on the pathogenesis of insulin resistance leading to gestational diabetes. Curr. Pharm. Biotechnol. 2014, 15, 32–37. [Google Scholar] [CrossRef]
- Spaight, C.; Gross, J.; Horsch, A.; Puder, J.J. Gestational Diabetes Mellitus. Endocr. Dev. 2016, 31, 163–178. [Google Scholar]
- Trojnar, M.; Patro-Małysza, J.; Kimber-Trojnar, Ż.; Czuba, M.; Mosiewicz, J.; Leszczyńska-Gorzelak, B. Vaspin in Serum and Urine of Post-Partum Women with Excessive Gestational Weight Gain. Medicina 2019, 55, 76. [Google Scholar] [CrossRef] [Green Version]
- Chiefari, E.; Arcidiacono, B.; Foti, D.; Brunetti, A. Gestational diabetes mellitus: An updated overview. J. Endocrinol. Investig. 2017, 40, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Gabbay-Benziv, R.; Baschat, A.A. Gestational diabetes as one of the “great obstetrical syndromes”—The maternal, placental, and fetal dialog. Best Pract. Res. Clin. Obstet. Gynaecol. 2015, 29, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Hiersch, L.; Berger, H.; Okby, R.; Ray, J.G.; Geary, M.; McDonald, S.D.; Murray-Davis, B.; Riddell, C.; Halperin, I.; Hasan, H.; et al. DOH-NET (Diabetes, Obesity and Hypertension in Pregnancy Research Network); SOON (Southern Ontario Obstetrical Network) Investigators. Gestational diabetes mellitus is associated with adverse outcomes in twin pregnancies. Am. J. Obstet. Gynecol. 2019, 220, 102.e1–102.e8. [Google Scholar] [PubMed] [Green Version]
- Zile, I.; Ebela, E.; Rozenfelde, I.R. Maternal Risk Factors for Stillbirth: A Registry–Based Study. Medicina 2019, 55, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepin, M.E.; Schiano, C.; Miceli, M.; Benincasa, G.; Mansueto, G.; Grimaldi, V.; Soricelli, A.; Wende, A.R.; Napoli, C. The human aortic endothelium undergoes dose-dependent DNA methylation in response to transient hyperglycemia. Exp. Cell Res. 2021, 400, 112485. [Google Scholar] [CrossRef] [PubMed]
- Strutz, J.; Baumann, K.; Weiss, E.; Hidden, U. Transient Hyperglycemia and Hypoxia Induce Memory Effects in AngiomiR Expression Profiles of Feto-Placental Endothelial Cells. Int. J. Mol. Sci. 2021, 22, 13378. [Google Scholar] [CrossRef] [PubMed]
- Liu, O.H.; Kiema, M.; Beter, M.; Yla-Herttuala, S.; Laakkonen, J.P.; Kaikkonen, M.U. Hypoxia-Mediated Regulation of Histone Demethylases Affects Angiogenesis-Associated Functions in Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2665–2677. [Google Scholar] [CrossRef]
- Wacker-Gussmann, A.; Schopen, J.; Engelhard, J.; Sitzberger, C.; Lienert, N.; Ewert, P.; Müller, A.; Schmidt, G.; Oberhoffer-Fritz, R.; Lobmaier, S.M. The Impact of Gestational Diabetes in Pregnancy on the Cardiovascular System of Children at One Year of Age. J. Clin. Med. 2021, 10, 5839. [Google Scholar] [CrossRef]
- Nangaku, M.; Hirakawa, Y.; Mimura, I.; Inagi, R.; Tanaka, T. Epigenetic Changes in the Acute Kidney Injury-to-Chronic Kidney Disease Transition. Nephron 2017, 137, 256–259. [Google Scholar] [CrossRef]
- Saxena, K.; Jolly, M.K. Acute vs. Chronic vs. Cyclic Hypoxia: Their Differential Dynamics, Molecular Mechanisms, and Effects on Tumor Progression. Biomolecules 2019, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Hanna, F.W.; Duff, C.J.; Shelley-Hitchen, A.; Hodgson, E.; Fryer, A.A. Diagnosing gestational diabetes mellitus: Implications of recent changes in diagnostic criteria and role of glycated haemoglobin (HbA1c). Clin. Med. 2017, 17, 108–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keating, N.; Carpenter, K.; McCarthy, K.; Coveney, C.; McAuliffe, F.; Mahony, R.; Walsh, J.; Hatunic, M.; Higgins, M. Clinical Outcomes Following a Change in Gestational Diabetes Mellitus Diagnostic Criteria Due to the COVID-19 Pandemic: A Case-Control Study. Int. J. Environ. Res. Public Health 2022, 19, 1884. [Google Scholar] [CrossRef] [PubMed]
- Fedullo, A.L.; Schiattarella, A.; Morlando, M.; Raguzzini, A.; Toti, E.; De Franciscis, P.; Peluso, I. Mediterranean Diet for the Prevention of Gestational Diabetes in the Covid-19 Era: Implications of Il-6 In Diabesity. Int. J. Mol. Sci. 2021, 22, 1213. [Google Scholar] [CrossRef] [PubMed]
- Zang, B.Y.; He, L.X.; Xue, L. Intermittent Fasting: Potential Bridge of Obesity and Diabetes to Health? Nutrients 2022, 14, 981. [Google Scholar] [CrossRef]
- Pérez-Pérez, A.; Vilariño-García, T.; Guadix, P.; Dueñas, J.L.; Sánchez-Margalet, V. Leptin and Nutrition in Gestational Diabetes. Nutrients 2020, 12, 1970. [Google Scholar] [CrossRef]
- Pheiffer, C.; Dias, S.; Jack, B.; Malaza, N.; Adam, S. Adiponectin as a Potential Biomarker for Pregnancy Disorders. Int. J. Mol. Sci. 2021, 22, 1326. [Google Scholar] [CrossRef]
- Trojnar, M.; Patro-Małysza, J.; Kimber-Trojnar, Ż.; Leszczyńska-Gorzelak, B.; Mosiewicz, J. Associations between Fatty Acid-Binding Protein 4-A Proinflammatory Adipokine and Insulin Resistance, Gestational and Type 2 Diabetes Mellitus. Cells 2019, 8, 227. [Google Scholar] [CrossRef] [Green Version]
- Kimber-Trojnar, Ż.; Patro-Małysza, J.; Trojnar, M.; Skórzyńska-Dziduszko, K.E.; Bartosiewicz, J.; Oleszczuk, J.; Leszczyńska-Gorzelak, B. Fatty Acid-Binding Protein 4-An “Inauspicious” Adipokine-In Serum and Urine of Post-Partum Women with Excessive Gestational Weight Gain and Gestational Diabetes Mellitus. J. Clin. Med. 2018, 7, 505. [Google Scholar] [CrossRef] [Green Version]
- Patro-Małysza, J.; Trojnar, M.; Kimber-Trojnar, Ż.; Mierzyński, R.; Bartosiewicz, J.; Oleszczuk, J.; Leszczyńska-Gorzelak, B. FABP4 in Gestational Diabetes-Association between Mothers and Offspring. J. Clin. Med. 2019, 8, 285. [Google Scholar] [CrossRef] [Green Version]
- Ruszała, M.; Niebrzydowska, M.; Pilszyk, A.; Kimber-Trojnar, Ż.; Trojnar, M.; Leszczyńska-Gorzelak, B. Novel Biomolecules in the Pathogenesis of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2021, 22, 11578. [Google Scholar] [CrossRef]
- Florian, A.R.; Cruciat, G.; Pop, R.M.; Staicu, A.; Daniel, M.; Florin, S. Predictive role of altered leptin, adiponectin and 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid secretion in gestational diabetes mellitus. Exp. Ther. Med. 2021, 21, 520. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, Y.; Deguchi, T.; Takano, M.; Takadate, A.; Lindup, W.E.; Otagiri, M. Renal disposition of a furan dicarboxylic acid and other uremic toxins in the rat. J. Pharmacol. Exp. Ther. 2002, 303, 880–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.G.; Rao, S.; Weir, T.L.; O’Malia, J.; Bazan, M.; Brown, R.J.; Ryan, E.P. Metabolomics and metabolic pathway networks from human colorectal cancers, adjacent mucosa, and stool. Cancer Metab. 2016, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, C.J. Lipotoxicity, β cell dysfunction, and gestational diabetes. Cell Metab. 2014, 19, 553–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetter, W.; Wendlinger, C. Furan fatty acids—Valuable minor fatty acids in food. Lipid Technol. 2013, 25, 7–10. [Google Scholar] [CrossRef]
- Hanhineva, K.; Lankinen, M.A.; Pedret, A.; Schwab, U.; Kolehmainen, M.; Paananen, J.; de Mello, V.; Sola, R.; Lehtonen, M.; Poutanen, K.; et al. Nontargeted metabolite profiling discriminates diet-specific biomarkers for consumption of whole grains, fatty fish, and bilberries in a randomized controlled trial. J. Nutr. 2014, 145, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Koppe, L.; Poitout, V. CMPF: A Biomarker for Type 2 Diabetes Mellitus Progression? Trends Endocrinol. Metab. 2016, 27, 439–440. [Google Scholar] [CrossRef]
- Xu, L.; Sinclair, A.J.; Faiza, M.; Li, D.; Han, X.; Yin, H.; Wang, Y. Furan fatty acids—Beneficial or harmful to health? Prog. Lipid Res. 2017, 68, 119–137. [Google Scholar] [CrossRef]
- Prentice, K.J.; Luu, L.; Allister, E.M.; Liu, Y.; Jun, L.S.; Sloop, K.W.; Hardy, A.B.; Wei, L.; Jia, W.; Fantus, I.G.; et al. The furan fatty acid metabolite CMPF is elevated in diabetes and induces beta cell dysfunction. Cell Metab. 2014, 19, 653–666. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Prentice, K.J.; Eversley, J.A.; Hu, C.; Batchuluun, B.; Leavey, K.; Hansen, J.B.; Wei, D.W.; Cox, B.; Dai, F.F.; et al. Rapid elevation in CMPF may act as a tipping point in diabetes development. Cell Rep. 2016, 14, 2889–2900. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, Y.; Iwao, Y.; Mera, K.; Watanabe, H.; Kadowaki, D.; Ishima, Y.; Chuang, V.T.; Sato, K.; Otagiri, M.; Maruyama, T. A uremic toxin, 3-carboxy-4-methyl-5-propyl-2-furanpropionate induces cell damage to proximal tubular cells via the generation of a radical intermediate. Biochem. Pharmacol. 2012, 84, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Jin, H.; Zhang, R.; Zhang, S.; Chen, P.; Yu, X.; Zhang, X. Increased serum 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) levels are associated with glucose metabolism in Chinese pregnant women. J. Endocrinol. Investig. 2018, 41, 663–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Li, G.; Zhang, X.; Xing, G.; Hu, X.; Yang, L.; Li, D. Lipid extract from hard-shelled mussel (Mytilus coruscus) improves clinical conditions of patients with rheumatoid arthritis: A randomized controlled trial. Nutrients 2015, 7, 625–645. [Google Scholar] [CrossRef] [Green Version]
- Lankinen, M.A.; Hanhineva, K.; Kolehmainen, M.; Lehtonen, M.; Auriola, S.; Mykkänen, H.; Poutanen, K.; Schwab, U.; Uusitupa, M. CMPF does not associate with impaired glucose metabolism in individuals with features of metabolic syndrome. PLoS ONE 2015, 10, e124379. [Google Scholar] [CrossRef] [PubMed]
- Retnakaran, R.; Ye, C.; Kramer, C.K.; Connelly, P.W.; Hanley, A.J.; Sermer, M.; Zinman, B. Evaluation of Circulating Determinants of Beta-Cell Function in Women with and Without Gestational Diabetes. J. Clin. Endocrinol. Metab. 2016, 101, 2683–2691. [Google Scholar] [CrossRef] [PubMed]
- Retnakaran, R.; Ye, C.; Kramer, C.K.; Connelly, P.W.; Hanley, A.J.; Sermer, M.; Zinman, B. Maternal serum prolactin and prediction of postpartum β-cell function and risk of prediabetes/diabetes. Diabetes Care 2016, 39, 1250–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.S.; Lin, M.; Imamura, F.; Cai, W.; Wang, L.; Feng, J.P.; Ruan, Y.; Tang, J.; Wang, F.; Yang, H.; et al. Serum metabolomics profiles in response to n-3 fatty acids in Chinese patients with type 2 diabetes: A double-blind randomised controlled trial. Sci. Rep. 2016, 6, 29522. [Google Scholar] [CrossRef] [Green Version]
- Savolainen, O.; Lind, M.V.; Bergström, G.; Fagerberg, B.; Sandberg, A.-S.; Ross, A. Biomarkers of food intake and nutrient status are associated with glucose tolerance status and development of type 2 diabetes in older Swedish women. Am. J. Clin. Nutr. 2017, 106, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Fardipour, S.; Moein, S.; Teshnizi, S.H.; Khaytian, M.; Qujeq, D. Evaluation of MASP1, CMPF and U.A serum levels in pre-diabetic subjects in comparison to Normal individuals for early diagnosis of subjects with pre-diabetes, a case-control study. J. Diabetes Metab. Disord. 2020, 19, 1593–1598. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, P.; Jin, H.; Yi, J.; Xie, X.; Yang, M.; Gao, T.; Yang, L.; Hu, C.; Zhang, X.; et al. Circulating 3-carboxy-4-methyl-5-propyl-2-furanpropanoic acid (CMPF) levels are associated with hyperglycemia and β cell dysfunction in a Chinese population. Sci. Rep. 2017, 7, 3114. [Google Scholar] [CrossRef] [Green Version]
- Abdeltawab, A.; Zaki, M.E.; Abdeldayem, Y.; Mohamed, A.A.; Zaied, S.M. Circulating micro RNA-223 and angiopoietin-like protein 8 as biomarkers of gestational diabetes mellitus. Br. J. Biomed. Sci. 2021, 78, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Ebert, T.; Kralisch, S.; Wurst, U.; Lössner, U.; Kratzsch, J.; Blüher, M.; Stumvoll, M.; Tönjes, A.; Fasshauer, M. Betatrophin levels are increased in women with gestational diabetes mellitus compared to healthy pregnant controls. Eur. J. Endocrinol. 2015, 173, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulmuş, F.G.; Melekoğlu, R.; Gürsu, M.F.; Bağcı, H.; Celik Kavak, E.; Akyol, A. Evaluation of second-trimester maternal serum betatrophin levels and lipid and carbohydrate metabolism parameters in patients with gestational diabetes mellitus. Turk. J. Obstet. Gynecol. 2020, 17, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Seyhanli, Z.; Seyhanli, A.; Aksun, S.; Pamuk, B.O. Evaluation of serum Angiopoietin-like protein 2 (ANGPTL-2), Angiopoietin-like protein 8 (ANGPTL-8), and high-sensitivity C-reactive protein (hs-CRP) levels in patients with gestational diabetes mellitus and normoglycemic pregnant women. J. Matern. Fetal Neonatal Med. 2021, 21, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.J.; Ma, L.L.; Li, G.; Chen, Y.X.; Zhou, J.Q. Circulating Betatrophin Levels and Gestational Diabetes Mellitus: A Systematic Review and Meta-Analysis. PLoS ONE 2017, 12, e0169941. [Google Scholar] [CrossRef] [PubMed]
- Wawrusiewicz-Kurylonek, N.; Telejko, B.; Kuzmicki, M.; Sobota, A.; Lipinska, D.; Pliszka, J.; Raczkowska, B.; Kuc, P.; Urban, R.; Szamatowicz, J.; et al. Increased Maternal and Cord Blood Betatrophin in Gestational Diabetes. PLoS ONE 2015, 10, e0131171. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Fang, C.; Ma, Z.; Guo, H.; Wang, R.; Hu, J. Betatrophin Levels were Increased in Pregnant Women with or without Gestational Diabetes Mellitus and Associated with Beta Cell Function. Rev. Bras. Ginecol. Obstet. 2016, 38, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Le Lay, J.; Yu, M.; Naji, A.; Kaestner, K.H. Elevated mouse hepatic betatrophin expression does not increase human β-cell replication in the transplant setting. Diabetes 2014, 63, 1283–1288. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Chen, X.; Chen, X.; Feng, Y.; Guo, H.; Li, S.; Dai, T.; Jiang, R.; Zhang, X.; Fang, C.; et al. Angiopoietin-like protein 8 in early pregnancy improves the prediction of gestational diabetes. Diabetologia 2018, 61, 574–580. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Lu, P.; He, W.; Zhang, J.; Liu, L.; Yang, Y.; Liu, Z.; Xie, J.; Shao, S.; Du, T.; et al. Circulating betatrophin levels are increased in patients with type 2 diabetes and associated with insulin resistance. J. Clin. Endocrinol. Metab. 2015, 100, 96–100. [Google Scholar] [CrossRef]
- Martinez-Perez, B.; Ejarque, M.; Gutierrez, C.; Nuñez-Roa, C.; Roche, K.; Vila-Bedmar, R.; Ballesteros, M.; Redondo-Angulo, I.; Planavila, A.; Villarroya, F.; et al. Angiopoietin-like protein 8 (ANGPTL8) in pregnancy: A brown adipose tissue-derived endocrine factor with a potential role in fetal growth. Transl. Res. 2016, 178, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Zhang, H.; Yu, S.; Deng, J.; Ma, S.; Li, Y.; Yuan, G.; Wang, J. Betatrophin for diagnosis and prognosis of mothers with gestational diabetes mellitus. J. Int. Med. Res. 2019, 47, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Yang, W.; Wang, G.; Liu, Y.; Jin, J. Association of betatrophin amounts with 25-(OH)D levels in patients with gestational diabetes mellitus. Medicine 2021, 100, e25646. [Google Scholar] [CrossRef] [PubMed]
- Boutsikou, T.; Briana, D.D.; Boutsikou, M.; Kafalidis, G.; Piatopoulou, D.; Baka, S.; Hassiakos, D.; Gourgiotis, D.; Malamitsi-Puchner, A. Cord blood nesfatin-1 in large for gestational age pregnancies. Cytokine 2013, 61, 591–594. [Google Scholar] [CrossRef]
- Kucukler, F.K.; Gorkem, U.; Simsek, Y.; Kocabas, R.; Gulen, S.; Guler, S. Low level of Nesfatin-1 is associated with gestational diabetes mellitus. Gynecol. Endocrinol. 2016, 32, 759–761. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, D.; Xu, J.; Chen, C.; Deng, D.; Pan, F.; Dong, L.; Li, S.; Ye, S. Circulating FABP4, nesfatin-1, and osteocalcin concentrations in women with gestational diabetes mellitus: A meta-analysis. Lipids Health Dis. 2020, 19, 199. [Google Scholar] [CrossRef]
- Mierzyński, R.; Poniedziałek-Czajkowska, E.; Dłuski, D.; Patro-Małysza, J.; Kimber-Trojnar, Ż.; Majsterek, M.; Leszczyńska-Gorzelak, B. Nesfatin-1 and Vaspin as Potential Novel Biomarkers for the Prediction and Early Diagnosis of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2019, 20, 159. [Google Scholar] [CrossRef] [Green Version]
- Stengel, A.; Mori, M.; Taché, Y. The role of nesfatin-1 in the regulation of food intake and body weight: Recent developments and future endeavors. Obes. Rev. 2013, 14, 859–870. [Google Scholar] [CrossRef] [Green Version]
- Aslan, M.; Celik, O.; Celik, N.; Turkcuoglu, I.; Yilmaz, E.; Karaer, A.; Simsek, Y.; Celik, E.; Aydin, S. Cord blood nesfatin-1 and apelin-36 levels in gestational diabetes mellitus. Endocrine 2012, 41, 424–429. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, J.H.; Zheng, S.Y.; Yan, J.H.; Chen, L.; Liu, X.; Wu, W.Z.; Wang, F. Serum levels of nesfatin-1 are increased in gestational diabetes mellitus. Gynecol. Endocrinol. 2017, 33, 621–624. [Google Scholar] [CrossRef]
- Su, Y.; Zhang, J.; Tang, Y.; Bi, F.; Liu, J.N. The novel function of nesfatin-1: Anti-hyperglycemia. Biochem. Biophys. Res. Commun. 2010, 391, 1039–1042. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S. Multi-functional peptide hormone NUCB2/nesfatin-1. Endocrine 2013, 44, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Ademoglu, E.N.; Gorar, S.; Keskin, M.; Carlioglu, A.; Ucler, R.; Erdamar, H.; Culha, C.; Aral, Y. Serum nesfatin-1 levels are decreased in pregnant women newly diagnosed with gestational diabetes. Arch. Endocrinol. 2017, 61, 455–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çaltekin, M.D.; Caniklioğlu, A. Maternal Serum Delta-Like 1 and Nesfatin-1 Levels in Gestational Diabetes Mellitus: A Prospective Case-Control Study. Cureus 2021, 13, e17001. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S. The presence of the peptides apelin, ghrelin and nesfatin-1 in the human breast milk, and the lowering of their levels in patients with gestational diabetes mellitus. Peptides 2010, 31, 2236–2240. [Google Scholar] [CrossRef]
- Atakul, N.; Atamer, Y.; Selek, Ş.; Kılıç, B.S.; Unal, F. Novel metabolic marker Afamin: A predictive factor for Large-for-Gestational-Age (LGA) fetus estimation in pregnancies with gestational diabetes mellitus? J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 102201. [Google Scholar] [CrossRef]
- Köninger, A.; Mathan, A.; Mach, P.; Frank, M.; Schmidt, B.; Schleussner, E.; Kimmig, R.; Gellhaus, A.; Dieplinger, H. Is Afamin a novel biomarker for gestational diabetes mellitus? A pilot study. Reprod. Biol. Endocrinol. 2018, 16, 30. [Google Scholar] [CrossRef]
- Lorenzo-Almorós, A.; Hang, T.; Peiró, C.; Soriano-Guillén, L.; Egido, J.; Tuñón, J.; Lorenzo, Ó. Predictive and diagnostic biomarkers for gestational diabetes and its associated metabolic and cardiovascular diseases. Cardiovasc. Diabetol. 2019, 18, 140. [Google Scholar] [CrossRef]
- Cai, Z.; Yang, Y.; Zhang, J. Hepatokine levels during the first or early second trimester of pregnancy and the subsequent risk of gestational diabetes mellitus: A systematic review and meta-analysis. Biomarkers 2021, 26, 517–531. [Google Scholar] [CrossRef]
- Köninger, A.; Iannaccone, A.; Hajder, E.; Frank, M.; Schmidt, B.; Schleussner, E.; Kimmig, R.; Gellhaus, A.; Dieplinger, H. Afamin predicts gestational diabetes in polycystic ovary syndrome patients preconceptionally. Endocr. Connect. 2019, 8, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Eroğlu, H.; Örgül, G.; Tonyalı, N.V.; Biriken, D.; Polat, N.; Yücel, A.; Yazihan, N.; Şahin, D. The Role of Afamin and Other Trace Elements in the Prediction of GDM: A Tertiary Center Experience. Biol. Trace Elem. Res. 2021, 199, 4418–4422. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg, F.; Kollerits, B.; Kiechl, S.; Lamina, C.; Kedenko, L.; Meisinger, C.; Willeit, J.; Huth, C.; Wietzorrek, G.; Altmann, M.E.; et al. Plasma concentrations of afamin are associated with the prevalence and development of metabolic syndrome. Circ. Cardiovasc. Genet. 2014, 7, 822–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubalek, M.; Buchner, H.; Mörtl, M.G.; Schlembach, D.; Huppertz, B.; Firulovic, B.; Köhler, W.; Hafner, E.; Dieplinger, B.; Wildt, L.; et al. The vitamin E-binding protein afamin increases in maternal serum during pregnancy. Clin. Chim. Acta 2014, 434, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, K.G.; Trevaskis, J.L.; Lam, D.D.; Sutton, G.M.; Koza, R.A.; Chouljenko, V.N.; Kousoulas, K.G.; Rogers, P.M.; Kesterson, R.A.; Thearle, M. Identification of adropin as a secreted factor linking dietary macronutrient intake with energy homeostasis and lipid metabolism. Cell. Metab. 2008, 8, 468–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topuz, M.; Celik, A.; Aslantas, T.; Demir, A.K.; Aydin, S.; Aydin, S. Plasma adropin levels predict endothelial dysfunction like flow-mediated dilatation in patients with type 2 diabetes mellitus. J. Investig. Med. 2013, 61, 1161–1164. [Google Scholar] [CrossRef] [PubMed]
- Beigi, A.; Shirzad, N.; Nikpour, F.; Nasli-Esfahani, E.; Emamgholipour, S.; Bandarian, F. Association between serum adropin levels and gestational diabetes mellitus; a case-control study. Gynecol. Endocrinol. 2015, 31, 939–941. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowski, F.A.; Jarmużek, P.; Gondek, A.; Cudnoch-Jędrzejewska, A.; Bomba-Opoń, D.; Wielgoś, M. First and third trimester serum concentrations of adropin and copeptin in gestational diabetes mellitus and normal pregnancy. Ginekol. Pol. 2016, 87, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Vivek, K.; Reddy, E.P.; Thangappazham, B.; Raj, H.; Pérez-López, F.R.; Varikasuvu, S.R. Maternal adropin levels in patients with gestational diabetes mellitus: A systematic review and meta-analysis. Gynecol. Endocrinol. 2021, 11, 1–5. [Google Scholar] [CrossRef]
- Mori, K.; Emoto, M.; Inaba, M. Fetuin-A: A multifunctional protein. Recent Pat. Endocr. Metab. Immune Drug Discov. 2011, 5, 124–146. [Google Scholar] [CrossRef]
- Ou, H.Y.; Yang, Y.C.; Wu, H.T.; Wu, J.S.; Lu, F.H.; Chang, C.J. Serum fetuin-A concentrations are elevated in subjects with impaired glucose tolerance and newly diagnosed type 2 diabetes. Clin. Endocrinol. 2011, 75, 450–455. [Google Scholar] [CrossRef]
- Pal, D.; Dasgupta, S.; Kundu, R.; Maitra, S.; Das, G.; Mukhopadhyay, S.; Ray, S.; Majumdar, S.S.; Bhattacharya, S. Fetuin-A acts as an endogenous ligand of TLR4 to promote lipid-induced insulin resistance. Nat. Med. 2012, 18, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Iyidir, O.T.; Degertekin, C.K.; Yilmaz, B.A.; Altinova, A.E.; Toruner, F.B.; Bozkurt, N.; Ayvaz, G.; Akturk, M. Serum levels of fetuin A are increased in women with gestational diabetes mellitus. Arch. Gynecol. Obstet. 2015, 291, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Šimják, P.; Cinkajzlová, A.; Anderlová, K.; Kloučková, J.; Kratochvílová, H.; Lacinová, Z.; Kaválková, P.; Krejčí, H.; Mráz, M.; Pařízek, A.; et al. Changes in plasma concentrations and mRNA expression of hepatokines fetuin A, fetuin B and FGF21 in physiological pregnancy and gestational diabetes mellitus. Physiol. Res. 2018, 28, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Lin, L.; Han, N.; Zhao, Z.; Liu, Z.; Luo, S.; Xu, X.; Liu, J.; Wang, H. Effects of dynamic change in fetuin-A levels from the first to the second trimester on insulin resistance and gestational diabetes mellitus: A nested case-control study. BMJ Open Diabetes Res. Care 2020, 8, e000802. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.J.; Zhang, L.; Zheng, T.; Zhang, G.H.; Du, K.; Yang, M.N.; He, H.; Wang, S.; Wang, W.; Zhang, J.; et al. Fetuin-A and fetal growth in gestational diabetes mellitus. BMJ Open Diabetes Res. Care 2020, 8, 000864. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Uzzau, S.; Goldblum, S.E.; Fasano, A. Human zonulin, a potential modulator of intestinal tight junctions. J. Cell Sci. 2000, 113, 4435–4440. [Google Scholar] [CrossRef]
- Vanuytsel, T.; Vermeire, S.; Cleynen, I. The role of haptoglobin and its related protein, zonulin, in inflammatory bowel disease. Tissue Barriers 2013, 1, e27321. [Google Scholar] [CrossRef] [Green Version]
- Fasano, A. Zonulin, regulation of tight junctions, and autoimmune diseases. Ann. N. Y. Acad. Sci. 2012, 258, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Sturgeon, C.; Fasano, A. Zonulin, a regulator of epithelial and endothelial barrier functions, and its involvement in chronic inflammatory diseases. Tissue Barriers 2016, 4, e1251384. [Google Scholar] [CrossRef] [Green Version]
- Mokkala, K.; Tertti, K.; Rönnemaa, T.; Vahlberg, T.; Laitinen, K. Evaluation of serum zonulin for use as an early predictor for gestational diabetes. Nutr. Diabetes 2017, 7, e253. [Google Scholar] [CrossRef] [Green Version]
- Bawah, A.T.; Seini, M.M.; Yakubu, Y.A.; Ussher, F.A.; Amoah, B.Y.; Alidu, H. First trimester zonulin levels and adiposity as predictive indices of gestational diabetes mellitus. Int. J. Diabetes Dev. Ctries 2019, 39, 451–457. [Google Scholar] [CrossRef]
- Demir, E.; Ozkan, H.; Seckin, K.D.; Sahtiyancı, B.; Demir, B.; Tabak, O.; Kumbasar, A.; Uzun, H. Plasma Zonulin Levels as a Non-Invasive Biomarker of Intestinal Permeability in Women with Gestational Diabetes Mellitus. Biomolecules 2019, 9, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, H.; Zhang, J.; Luan, J.; Xu, H.; Huang, Z.; Yu, Q.; Gou, X.; Xu, L. Secreted Frizzled Related Proteins in Cardiovascular and Metabolic Diseases. Front. Endocrinol. 2021, 12, 712217. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, S.; Kabekkodu, S.P.; Pai, K.S.R. Aberrant Canonical Wnt Signaling: Phytochemical Based Modulation. Phytomedicine 2020, 76, 153243. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Agostino, M.; Pohl, S.O. Wnt Binding Affinity Prediction for Putative Frizzled-Type Cysteine-Rich Domains. Int. J. Mol. Sci. 2019, 20, 4168. [Google Scholar] [CrossRef] [Green Version]
- Park, J.R.; Jung, J.W.; Lee, Y.S.; Kang, K.S. The roles of Wnt antagonists Dkk1 and sFRP4 during adipogenesis of human adipose tissue-derived mesenchymal stem cells. Cell Prolif. 2008, 41, 859–874. [Google Scholar] [CrossRef]
- Ouchi, N.; Higuchi, A.; Ohashi, K.; Oshima, Y.; Gokce, N.; Shibata, R.; Akasaki, Y.; Shimono, A.; Walsh, K. Sfrp5 is an anti-inflammatory adipokine that modulates metabolic dysfunction in obesity. Science 2010, 329, 454–457. [Google Scholar] [CrossRef] [Green Version]
- Ehrlund, A.; Mejhert, N.; Lorente-Cebrian, S.; Astrom, G.; Dahlman, I.; Laurencikiene, J.; Ryden, M. Characterization of the Wnt inhibitors secreted frizzled-related proteins (SFRPs) in human adipose tissue. J. Clin. Endocrinol. Metab. 2013, 98, E503–E508. [Google Scholar] [CrossRef]
- Taciak, B.; Pruszynska, I.; Kiraga, L.; Bialasek, M.; Krol, M. Wnt Signaling Pathway in Development and Cancer. J. Physiol. Pharmacol. 2018, 69, 185–196. [Google Scholar]
- Huang, A.; Huang, Y. Role of Sfrps in cardiovascular disease. Ther. Adv. Chronic Dis. 2020, 11, 2040622320901990. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.M.; Werner, C.; Böhm, M.; Laufs, U.; Winkler, K. Association of secreted frizzled-related protein 4 (SFRP4) with type 2 diabetes in patients with stable coronary artery disease. Cardiovasc. Diabetol. 2014, 13, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schinner, S.; Ulgen, F.; Papewalis, C.; Schott, M.; Woelk, A.; Vidal-Puig, A.; Scherbaum, W.A. Regulation of insulin secretion, glucokinase gene transcription and beta cell proliferation by adipocyte-derived Wnt signalling molecules. Diabetologia 2007, 51, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Qu, H.; Li, Y.; Tang, Q.; Yang, Z.; Wang, H.; Deng, H. Relationship between serum secreted frizzled-related protein 4 levels and the first-phase of glucose-stimulated insulin secretion in individuals with different glucose tolerance. Endocr. J. 2015, 62, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Brix, J.M.; Krzizek, E.C.; Hoebaus, C.; Ludvik, B.; Schernthaner, G.; Schernthaner, G.H. Secreted Frizzled-Related Protein 4 (SFRP4) Is Elevated in Patients with Diabetes Mellitus. Horm. Metab. Res. 2016, 48, 345–348. [Google Scholar] [CrossRef]
- Baldane, S.; Ipekci, S.H.; Ekin, A.; Abusoglu, S.; Unlu, A.; Kebapcilar, L. Evaluation of Fractalkine (Fkn) and Secreted Frizzled-Related Protein 4 (Sfrp-4) Serum Levels in Patients with Prediabetes and Type 2 Diabetes. Bratisl. Lek. Listy 2018, 119, 112–115. [Google Scholar] [CrossRef]
- Mahdi, T.; Hänzelmann, S.; Salehi, A.; Muhammed, S.J.; Reinbothe, T.M.; Tang, Y.; Axelsson, A.S.; Zhou, Y.; Jing, X.; Almgren, P.; et al. Secreted frizzled-related protein 4 reduces insulin secretion and is overexpressed in type 2 diabetes. Cell Metab. 2012, 16, 625–633. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Xu, C.; Luo, Y.; Wang, G.; Wu, J.; Wan, Q. Circulating Mir-103 Family as Potential Biomarkers for Type 2 Diabetes Through Targeting Cav-1 and Sfrp4. Acta Diabetol. 2019, 57, 309–322. [Google Scholar] [CrossRef]
- Bukhari, S.A.; Yasmin, A.; Zahoor, M.A.; Mustafa, G.; Sarfraz, I.; Rasul, A. Secreted frizzled-related protein 4 and its implication in obesity and type-2 diabetes. IUBMB Life 2019, 71, 1701–1710. [Google Scholar] [CrossRef]
- Oh, K.J.; Lee, D.S.; Kim, W.K.; Han, B.S.; Lee, S.C.; Bae, K.H. Metabolic Adaptation in Obesity and Type II Diabetes: Myokines, Adipokines and Hepatokines. Int. J. Mol. Sci. 2016, 18, 8. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.B.; Chen, X.D.; Zhou, X.Y.; Zhu, Q. The Wnt antagonist and secreted frizzled-related protein 5: Implications on lipid metabolism, inflammation, and type 2 diabetes mellitus. Biosci. Rep. 2018, 38, BSR20180011. [Google Scholar] [CrossRef] [PubMed]
- Wang, C. Obesity, inflammation, and lung injury (OILI): The good. Mediators Inflamm. 2014, 2014, 978463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimber-Trojnar, Ż.; Patro-Małysza, J.; Skórzyńska-Dziduszko, K.E.; Oleszczuk, J.; Trojnar, M.; Mierzyński, R.; Leszczyńska-Gorzelak, B. Ghrelin in Serum and Urine of Post-Partum Women with Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2018, 19, 3001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhang, S.; Liu, H.; Wang, L.; Zhang, C.; Leng, J.; Yu, Z.; Yang, X.; Tian, H.; Hu, G. Different associations of diabetes with β-cell dysfunction and insulin resistance among obese and nonobese Chinese women with prior gestational diabetes mellitus. Diabetes Care 2014, 37, 2533–2539. [Google Scholar] [CrossRef] [Green Version]
- Trojnar, M.; Kimber-Trojnar, Ż.; Leszczyńska-Gorzelak, B. Secreted Frizzled-Related Protein 5 in Serum and Urine of Post-Partum Women with Gestational Diabetes Mellitus. J. Educ. Health Sport 2019, 9, 140–152. [Google Scholar]
- Oztas, E.; Ozler, S.; Ersoy, E.; Ersoy, A.O.; Tokmak, A.; Ergin, M.; Uygur, D.; Danisman, N. Prediction of gestational diabetes mellitus by first trimester serum secreted frizzle-related protein-5 levels. J. Matern. Fetal Neonatal Med. 2016, 29, 1515–1519. [Google Scholar] [CrossRef]
- Hu, Z.; Deng, H.; Qu, H. Plasma Sfrp5 Levels Are Decreased in Chinese Subjects with Obesity and Type 2 Diabetes and Negatively Correlated with Parameters of Insulin Resistance. Diabetes Res. Clin. Pract. 2013, 99, 391–395. [Google Scholar] [CrossRef]
- Lu, Y.C.; Wang, C.P.; Hsu, C.C.; Chiu, C.A.; Yu, T.H.; Hung, W.C.; Lu, L.F.; Chung, F.M.; Tsai, I.T.; Lin, H.C.; et al. Circulating secreted frizzled-related protein 5 (Sfrp5) and wingless-type MMTV integration site family member 5a (Wnt5a) levels in patients with type 2 diabetes mellitus. Diabetes Metab. Res. Rev. 2013, 29, 551–556. [Google Scholar]
- Toan, N.L.; Van Hoan, N.; Cuong, D.V.; Dung, N.V.; Dung, P.T.; Hang, N.T.; Dieu, D.T.H.; Chung, D.T.; Son, H.A.; Phong, P.X.; et al. Adipose tissue-derived cytokines and their correlations with clinical characteristics in Vietnamese patients with type 2 diabetes mellitus. Diabetol. Metab. Syndr. 2018, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Guan, B.; Li, W.; Li, F.; Xie, Y.; Ni, Q.; Gu, Y.; Li, X.; Wang, Q.; Zhang, H.; Ning, G. Sfrp5 mediates glucose-induced proliferation in rat pancreatic β-cells. J. Endocrinol. 2016, 229, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Christodoulides, C.; Lagathu, C.; Sethi, J.K.; Vidal-Puig, A. Adipogenesis and WNT signalling. Trends Endocrinol. Metab. 2009, 20, 16–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, H.; Prestwich, T.C.; Reid, M.A.; Longo, K.A.; Gerin, I.; Cawthorn, W.P.; Susulic, V.S.; Krishnan, V.; Greenfield, A.; Macdougald, O.A. Secreted frizzled-related protein 5 suppresses adipocyte mitochondrial metabolism through WNT inhibition. J. Clin. Investig. 2012, 122, 2405–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rulifson, I.C.; Majeti, J.Z.; Xiong, Y.; Hamburger, A.; Lee, K.J.; Miao, L.; Lu, M.; Gardner, J.; Gong, Y.; Wu, H.; et al. Inhibition of secreted frizzled-related protein 5 improves glucose metabolism. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E1144–E1152. [Google Scholar] [CrossRef]
- Van Camp, J.K.; Beckers, S.; Zegers, D.; Verrijken, A.; Van Gaal, L.F.; Van Hul, W. Common genetic variationin sFRP5 is associated with fat distribution in men. Endocrine 2014, 46, 477–484. [Google Scholar] [CrossRef]
- Prats-Puig, A.; Soriano-Rodríguez, P.; Carreras-Badosa, G.; Riera-Pérez, E.; Ros-Miquel, M.; Gomila-Borja, A.; de Zegher, F.; Ibáñez, L.; Bassols, J.; López-Bermejo, A. Balanced duo of anti-inflammatory SFRP5 and proinflammatory WNT5A in children. Pediatr. Res. 2014, 75, 793–797. [Google Scholar] [CrossRef] [Green Version]
- Kimber-Trojnar, Ż.; Patro-Małysza, J.; Trojnar, M.; Darmochwał-Kolarz, D.; Oleszczuk, J.; Leszczyńska-Gorzelak, B. Umbilical Cord SFRP5 Levels of Term Newborns in Relation to Normal and Excessive Gestational Weight Gain. Int. J. Mol. Sci. 2019, 20, 595. [Google Scholar] [CrossRef] [Green Version]
- Gurlo, T.; Ryazantsev, S.; Huang, C.J.; Yeh, M.W.; Reber, H.A.; Hines, O.J.; O’Brien, T.D.; Glabe, C.G.; Butler, P.C. Evidence for proteotoxicity in β cells in type 2 diabetes: Toxic islet amyloid polypeptide oligomers form intracellularly in the secretory pathway. Am. J. Pathol. 2010, 176, 861–869. [Google Scholar] [CrossRef]
- Akter, R.; Cao, P.; Noor, H.; Ridgway, Z.; Tu, L.H.; Wang, H.; Wong, A.G.; Zhang, X.; Abedini, A.; Schmidt, A.M.; et al. Islet Amyloid Polypeptide: Structure, Function, and Pathophysiology. J. Diabetes Res. 2016, 2016, 2798269. [Google Scholar] [CrossRef] [Green Version]
- Gurlo, T.; Kim, S.; Butler, A.E.; Liu, C.; Pei, L.; Rosenberger, M.; Butler, P.C. Pregnancy in human IAPP transgenic mice recapitulates beta cell stress in type 2 diabetes. Diabetologia 2019, 62, 1000–1010. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Westermark, G.T. The Amyloid Forming Peptides Islet Amyloid Polypeptide and Amyloid β Interact at the Molecular Level. Int. J. Mol. Sci. 2021, 22, 11153. [Google Scholar] [CrossRef]
- Abioye, R.O.; Okagu, O.D.; Udenigwe, C.C. Disaggregation of Islet Amyloid Polypeptide Fibrils as a Potential Anti-Fibrillation Mechanism of Tetrapeptide TNGQ. Int. J. Mol. Sci. 2022, 23, 1972. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, P.I.; Martín-Montalvo, A.; Cobo Vuilleumier, N.; Gauthier, B.R. Molecular Modelling of Islet β-Cell Adaptation to Inflammation in Pregnancy and Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2019, 20, 6171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigorakis, S.I.; Alevizaki, M.; Rapti, E.; Anastasiou, E.; Alevizaki, C.C.; Souvatzoglou, A. Islet amyloid polypeptide (amylin) does not seem to be directly involved in the development of gestational diabetes mellitus. Gynecol. Obstet. Investig. 1997, 44, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Wareham, N.J.; Swinn, R.; Fineman, M.S.; Koda, J.E.; Taylor, K.; Williams, D.E.; Rahilly, S.O. Gestational diabetes mellitus is associated with an increase in the total concentration of amylin molecules. Diabetes Care 1998, 21, 668–669. [Google Scholar] [CrossRef] [PubMed]
- Kinalski, M.; Sledziewski, A.; Telejko, B.; Straczkowski, M.; Kretowski, A.; Kinalska, I. Post-partum evaluation of amylin in lean patients with gestational diabetes mellitus. Acta Diabetol. 2004, 41, 1–4. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Biomolecules | Localization | Mechanism of the Action |
|---|---|---|
| CMPF | Liver, pancreas | stimulates oxidative stress, acts on mitochondrial functions and dysregulates glucose-induced ATP accumulation [49] |
| ANGPTL-8 | Adipose tissue, liver, vascular and hematopoietic systems | supporting role in stimulating proliferation and increasing pancreatic beta cell mass, improves glucose tolerance in insulin resistance, likely by increasing insulin secretion [64,67], |
| Nesfatin-1 | Pancreas, duodenum, adipose tissue, peripheral and central nervous system (arcuate, paraventricular nuclei and nucleus of the solitary tract), skeletal muscles, heart, kidneys, liver, skin, lungs, articular cartilage | stimulates pre-proinsulin mRNA expression and increases glucose-induced insulin release [78], inhibits glucagon secretion [78], antihyperglycemic effects in animal studies [81] |
| Afamin | Liver, placenta, ovarian follicular, seminal fluids, cerebrospinal fluids, plasma | antiapoptotic and antioxidant qualities [90,91], upregulation of oxidative stress increases concentrations of afamin [86,90] |
| Adropin | Lungs, liver, cardiovascular system, adipose tissue, kidneys, pancreas, brain, cerebellum, small intestine, endothelial cells | regulates cellular energy metabolism and anti-inflammatory processes (IL-10↑, TGFβ↑, IL-12↓, TNFα↓), takes part in anti-oxidative stress [95,96] |
| Fetuin-A | Skeletal muscles, adipose tissue, cardiovascular system, liver | a ligand for toll-like receptor 4 (TLR-4), through which lipids induce insulin resistance, induce inflammatory signaling [101] |
| Zonulin | Digestive system, liver, heart, brain, adipose tissue, lungs, kidneys, skin, immune cells | physiological modulator of intercellular tight junctions (TJs) between intestinal epithelial cells [106] |
| SFRPs | Heart, adipose tissue, pancreas, skeletal muscles, liver, aorta, endometrium, gallbladder, kidneys, prostate, testis, urinary bladder, ovary, esophagus, skin, small intestine, colon, appendix, spleen, bone marrow, duodenum, adrenal | extracellular signaling ligands and Wnt signaling pathway antagonistic inhibitors [113,114,115], take part in the adipogenesis [117,118,119], adipocyte differentiation [120,121,122], may worsen insulin secretion by acting on islet cells through β-catenin, TCF4, CyclinD [123], sensitizing skeletal muscle cells to insulin. By the Wnt/Ca2+ signaling pathway, enhance intracellular Ca2+ and protein kinase C, calmodulin kinase II is activated [122,124,125,126], steer hypothalamic insulin signaling pathway, inhibits N-methyl-D-aspartate receptor and inhibits the secretion of hepatic glucose [140] |
| Amylin | Pancreatic β-cells | plays a significant role in regulation of glucose metabolism [147], controls gastric emptying, suppression of glucagon release and regulation of satiety [148,149], penetrates cell membranes [152], resulting in an imbalance of intracellular ions, formation of reactive oxidant species, membrane damage and loss of β-cells [150,151] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruszała, M.; Pilszyk, A.; Niebrzydowska, M.; Kimber-Trojnar, Ż.; Trojnar, M.; Leszczyńska-Gorzelak, B. Novel Biomolecules in the Pathogenesis of Gestational Diabetes Mellitus 2.0. Int. J. Mol. Sci. 2022, 23, 4364. https://doi.org/10.3390/ijms23084364
Ruszała M, Pilszyk A, Niebrzydowska M, Kimber-Trojnar Ż, Trojnar M, Leszczyńska-Gorzelak B. Novel Biomolecules in the Pathogenesis of Gestational Diabetes Mellitus 2.0. International Journal of Molecular Sciences. 2022; 23(8):4364. https://doi.org/10.3390/ijms23084364
Chicago/Turabian StyleRuszała, Monika, Aleksandra Pilszyk, Magdalena Niebrzydowska, Żaneta Kimber-Trojnar, Marcin Trojnar, and Bożena Leszczyńska-Gorzelak. 2022. "Novel Biomolecules in the Pathogenesis of Gestational Diabetes Mellitus 2.0" International Journal of Molecular Sciences 23, no. 8: 4364. https://doi.org/10.3390/ijms23084364
APA StyleRuszała, M., Pilszyk, A., Niebrzydowska, M., Kimber-Trojnar, Ż., Trojnar, M., & Leszczyńska-Gorzelak, B. (2022). Novel Biomolecules in the Pathogenesis of Gestational Diabetes Mellitus 2.0. International Journal of Molecular Sciences, 23(8), 4364. https://doi.org/10.3390/ijms23084364