
TaKLU Plays as a Time Regulator of Leaf Growth via Auxin Signaling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
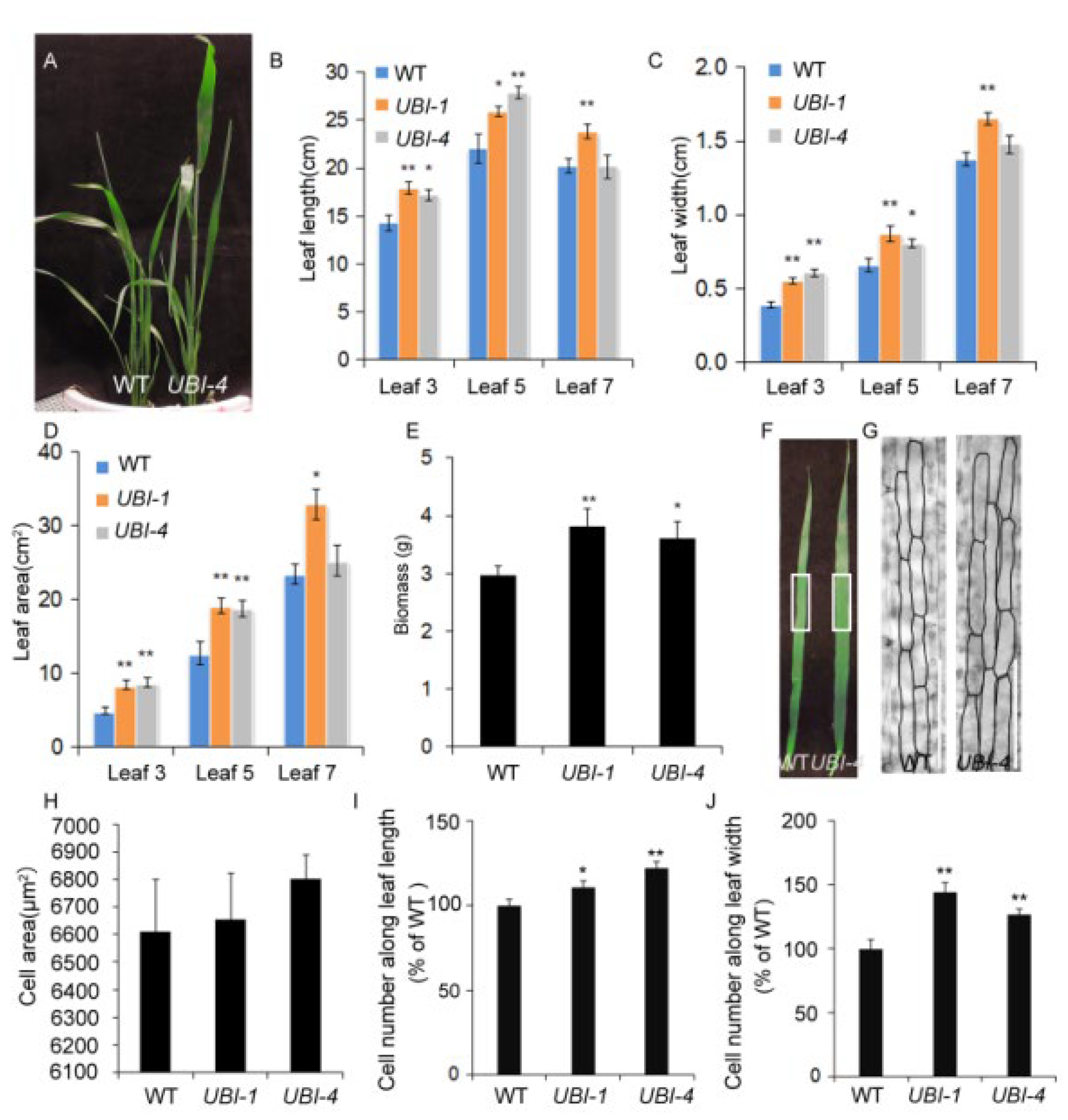
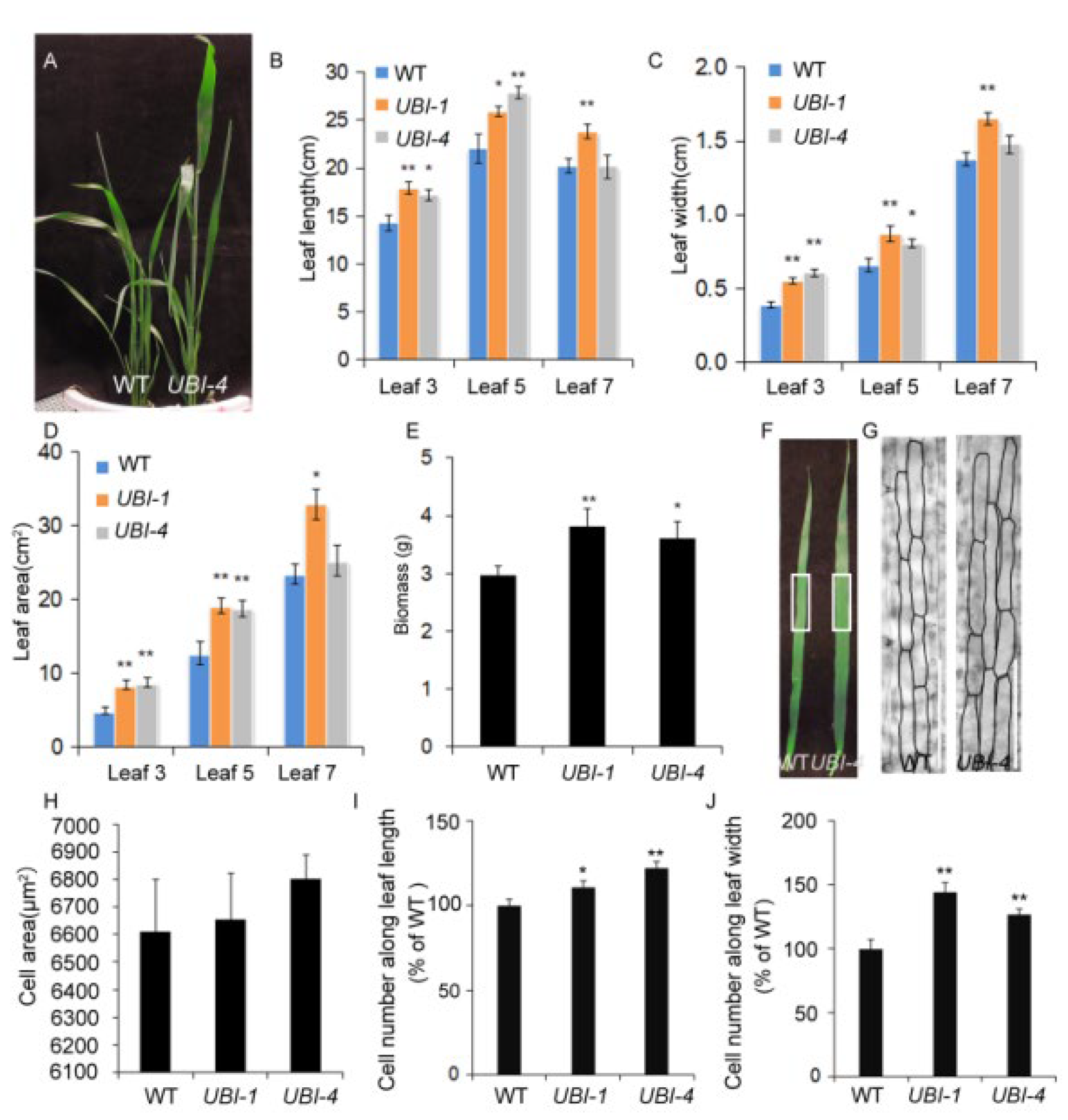
2.1. Overexpression of TaKLU Increased Leaf Size and Biomass by Promoting Cell Division
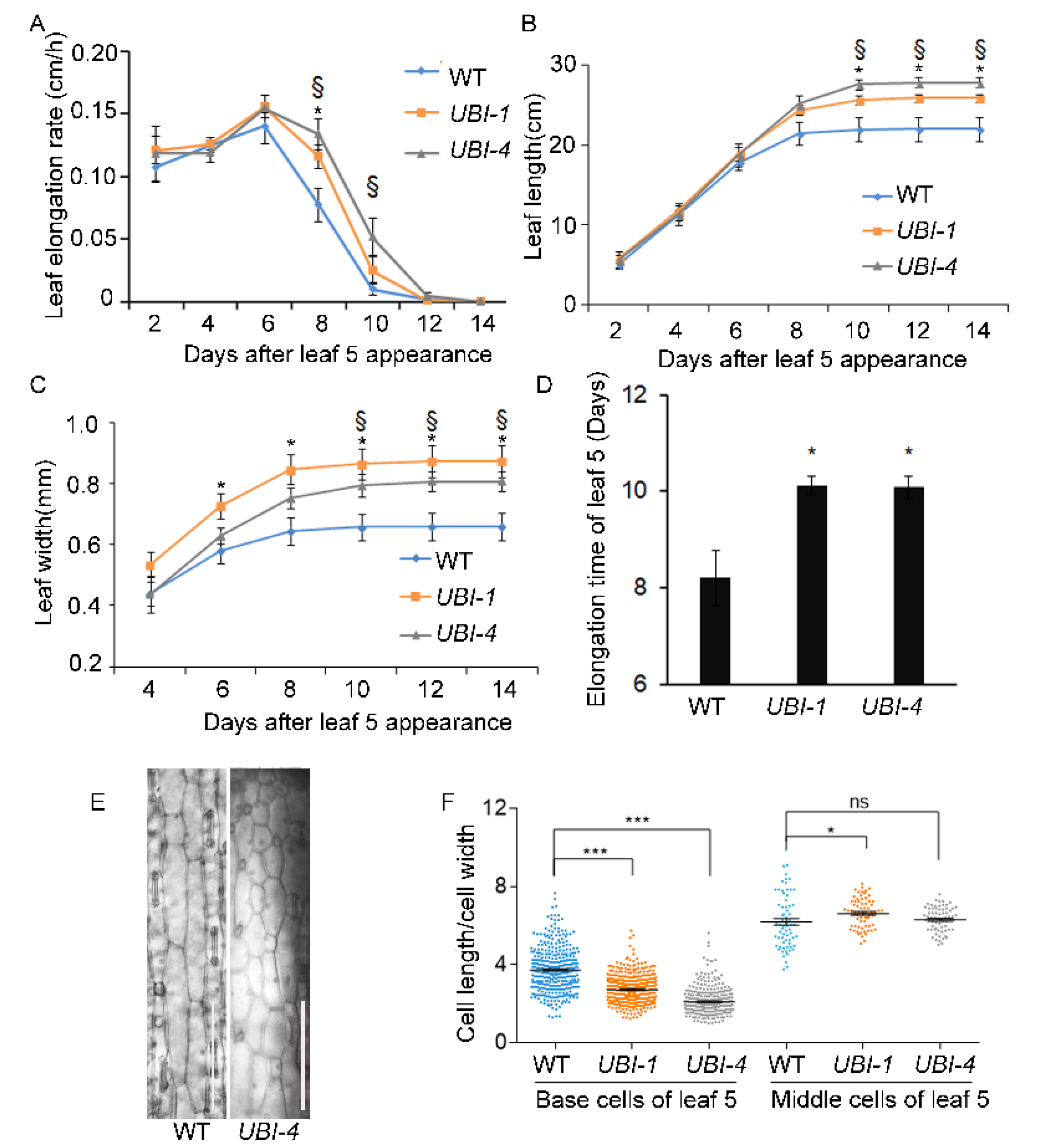
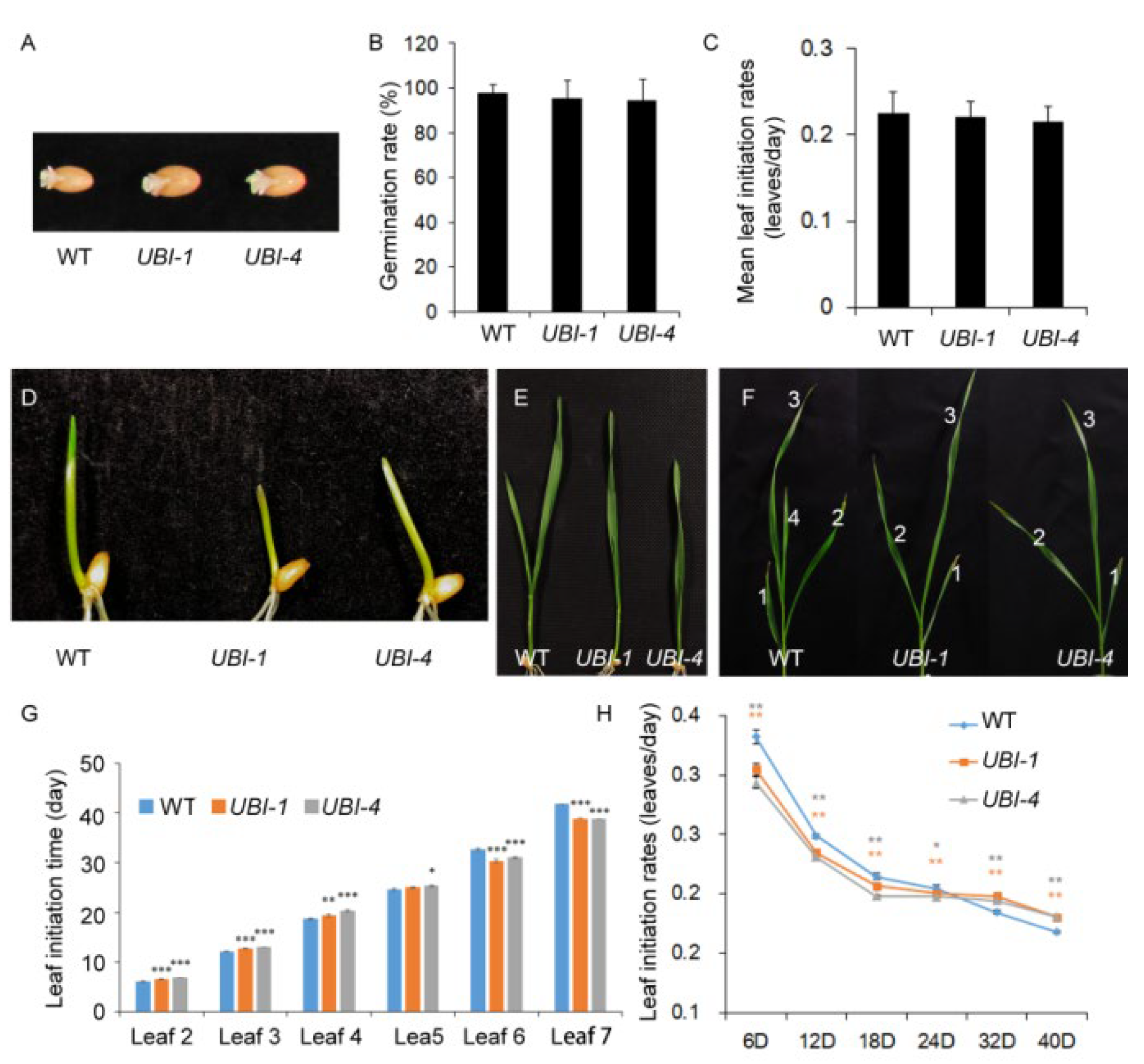
2.2. TaKLU Overexpression Extended the Time of Leaf Elongation
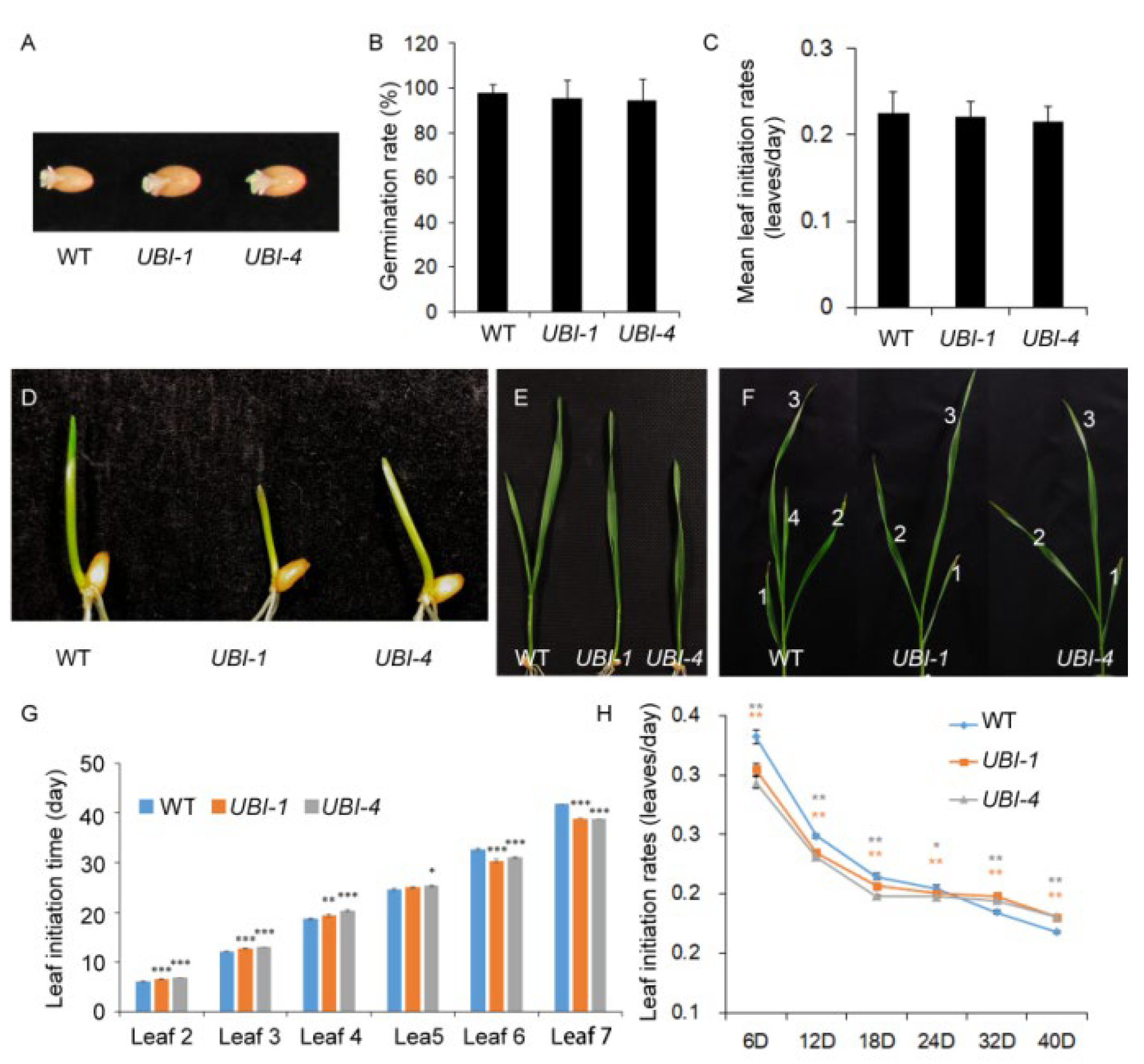
2.3. Overexpression of TaKLU Smoothed the Fluctuations of Leaf Initiation Time in Wheat
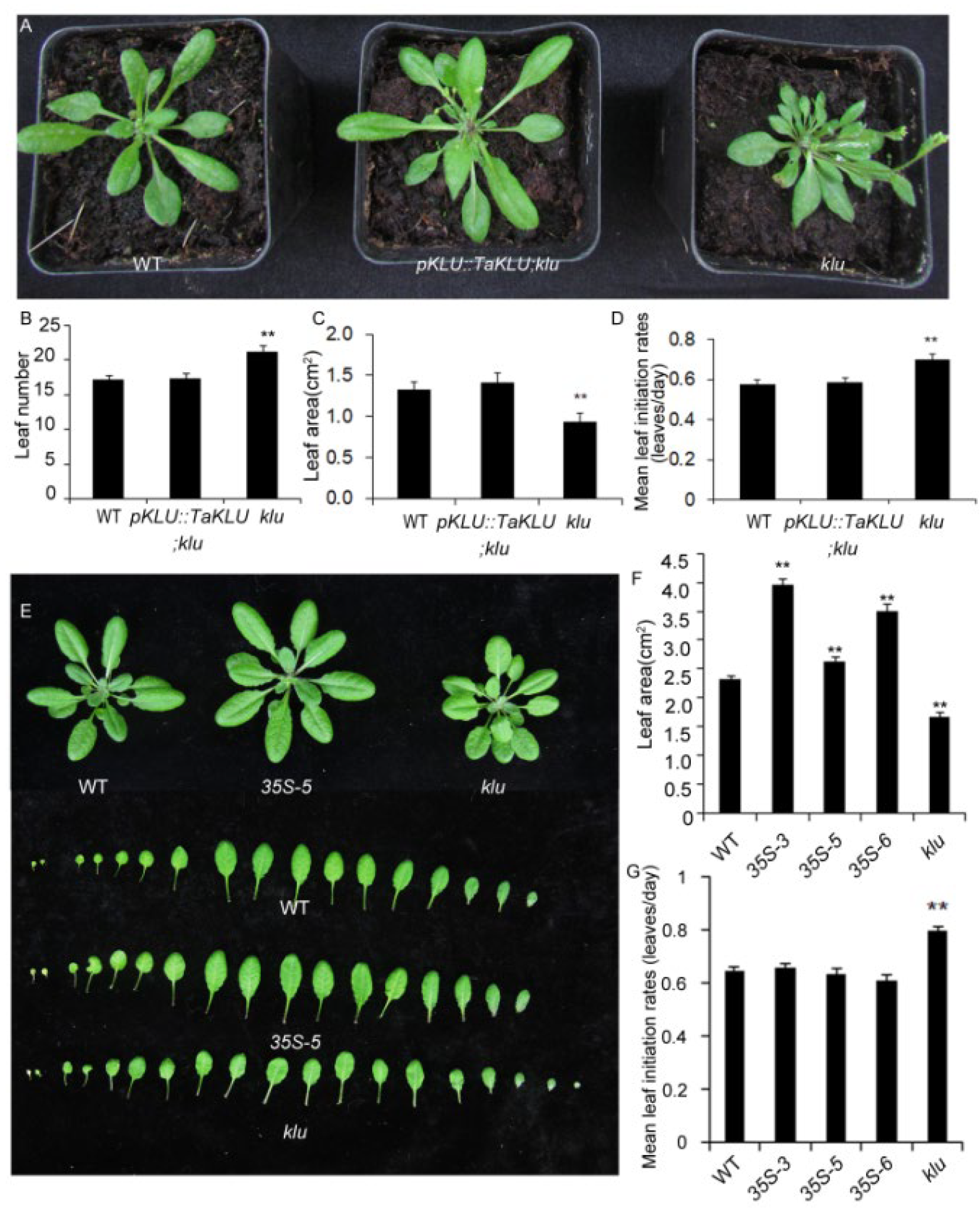
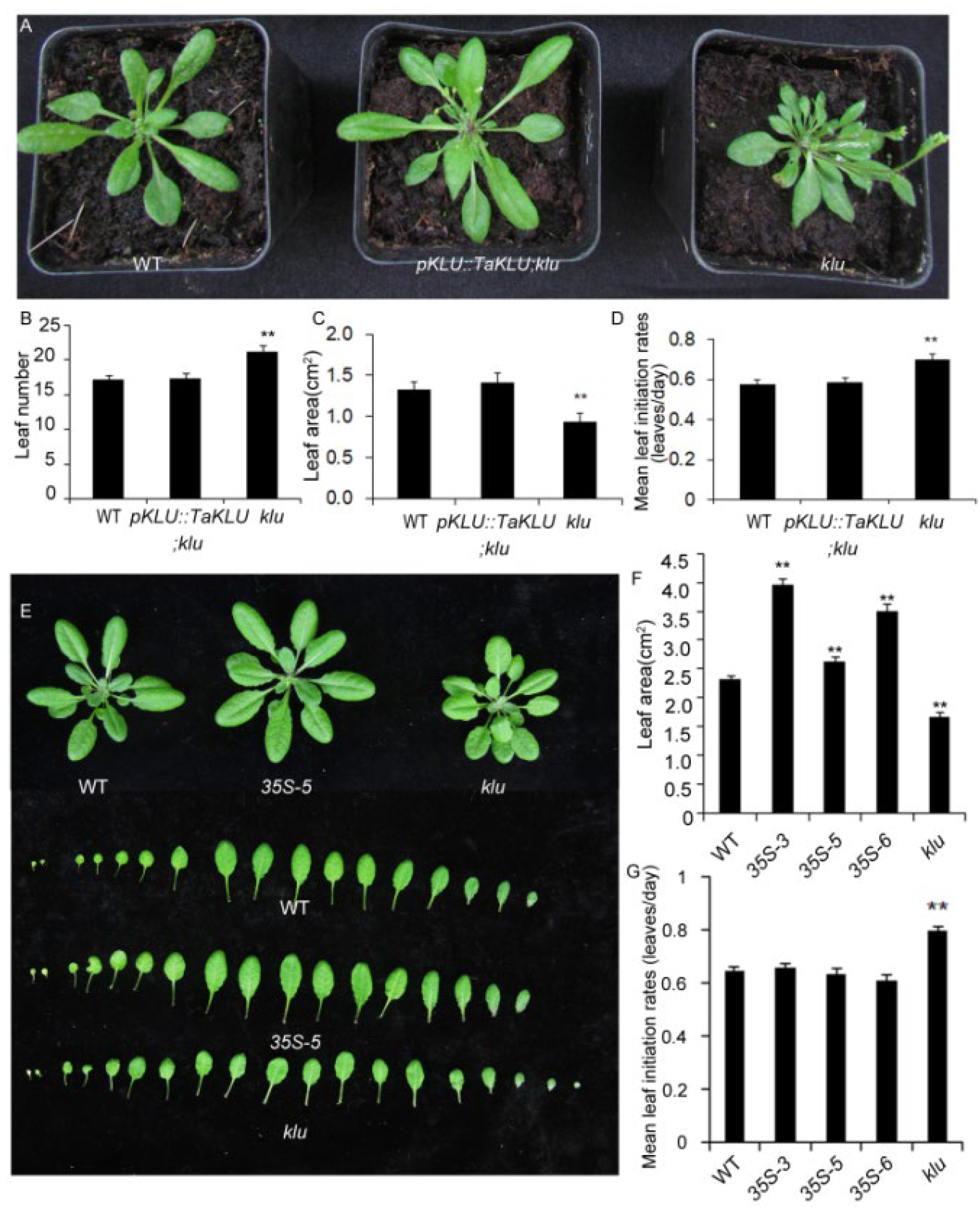
2.4. TaKLU Was Functionally Conserved in Both Monocots and Dicots
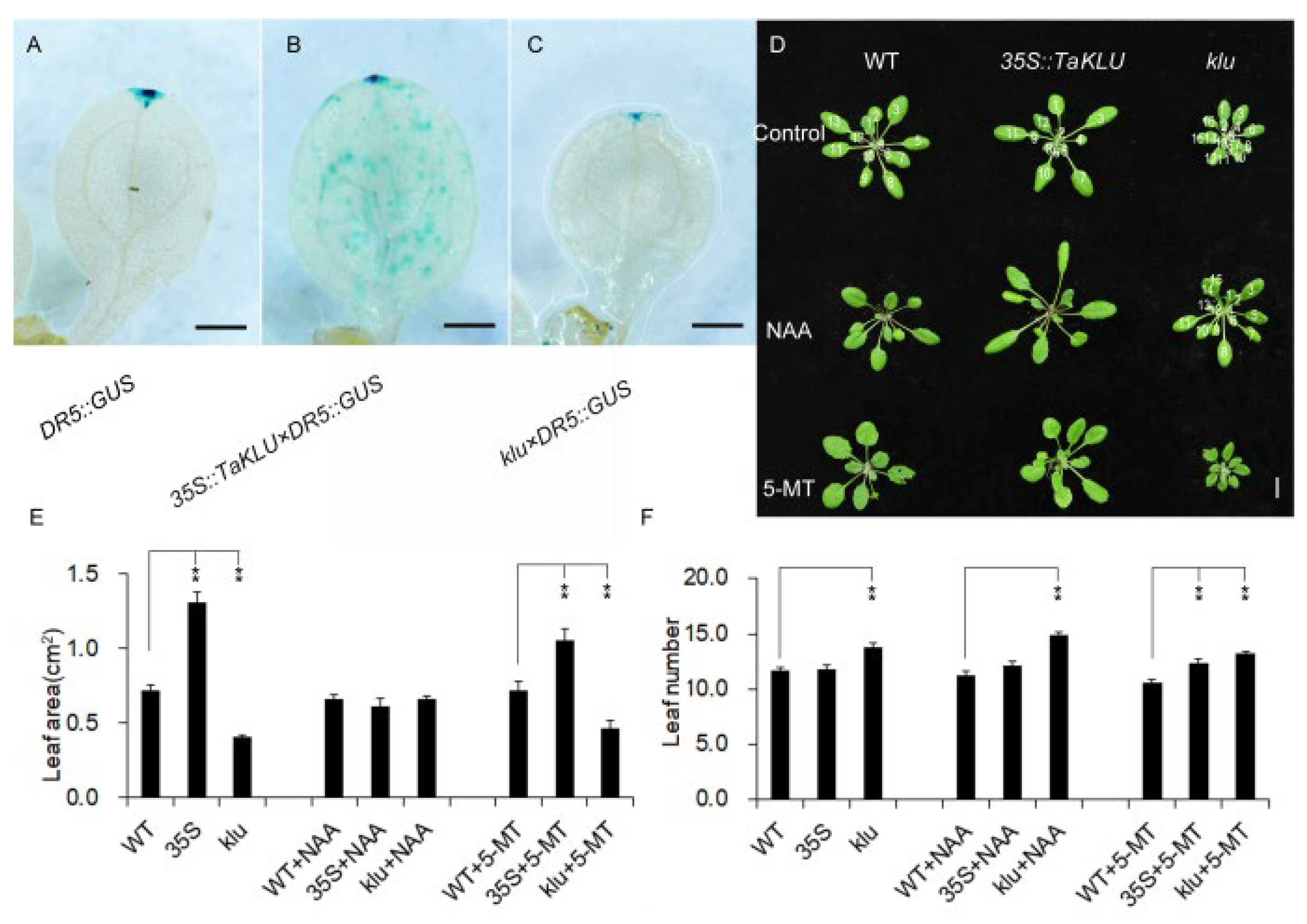
2.5. TaKLU Promoted Leaf Growth via Auxin Signaling
3. Discussion
3.1. KLU Plays as a Time Regulator of Leaf Growth
3.2. Relationship between KLU and Plant Hormones
3.3. Strategies of TaKLU Applied in Wheat Breeding
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Vector Construction and Transformation
4.3. RNA Isolation and QRT-PCR Analysis
4.4. Germination Experiment
4.5. Morphological Analysis
4.6. Chemical Treatments
4.7. Cytological Analysis
4.8. Histochemical Staining
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xiong, Y.; Jiao, Y. The Diverse Roles of Auxin in Regulating Leaf Development. Plants 2019, 8, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Y.; Fang, Y.; Hu, P.; Tan, Y.; Wang, Y.; Hou, L.; Deng, X.; Wu, H.; Zhu, L.; Zhu, L.; et al. Construction of a High-Density Genetic Map Based on Slaf Markers and Qtl Analysis of Leaf Size in Rice. Front. Plant Sci. 2020, 11, 1143. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Tu, Y.; Zhu, J.; Luo, W.; Liu, H.; Li, C.; Li, S.; Liu, J.; Ding, P.; Habib, A.; et al. Flag Leaf Size and Posture of Bread Wheat: Genetic Dissection, Qtl Validation and Their Relationships with Yield-Related Traits. Theor. Appl. Genet. 2020, 133, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Sizani, B.L.; Kalve, S.; Markakis, M.N.; Domagalska, M.A.; Stelmaszewska, J.; AbdElgawad, H.; Zhao, X.; De Veylder, L.; De Vos, D.; Broeckhove, J.; et al. Multiple Mechanisms Explain How Reduced Krp Expression Increases Leaf Size of Arabidopsis Thaliana. New Phytol. 2019, 221, 1345–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.K.; Rhee, J.Y.; Lee, S.H.; Chung, G.C.; Park, S.J.; Segami, S.; Maeshima, M.; Choi, G. Functionally Redundant Lng3 and Lng4 Genes Regulate Turgor-Driven Polar Cell Elongation through Activation of Xth17 and Xth24. Plant Mol. Biol. 2018, 97, 23–36. [Google Scholar] [CrossRef]
- Vanhaeren, H.; Nam, Y.J.; De Milde, L.; Chae, E.; Storme, V.; Weigel, D.; Gonzalez, N.; Inzé, D. Forever Young: The Role of Ubiquitin Receptor Da1 and E3 Ligase Big Brother in Controlling Leaf Growth and Development. Plant Physiol. 2017, 173, 1269–1282. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.; Lowndes, L.; Regan, S.; Beardmore, T. Overexpression of Cycd1;2 in Activation-Tagged Populus Tremula X Populus Alba Results in Decreased Cell Size and Altered Leaf Morphology. Tree Genet. Genomes 2015, 11, 66. [Google Scholar] [CrossRef]
- Donnelly, P.M.; Bonetta, D.; Tsukaya, H.; Dengler, R.E.; Dengler, N.G. Cell Cycling and Cell Enlargement in Developing Leaves of Arabidopsis. Dev. Biol. 1999, 215, 407–419. [Google Scholar] [CrossRef] [Green Version]
- Mizutani, M.; Ohta, D. Diversification of P450 Genes During Land Plant Evolution. Annu. Rev. Plant Biol. 2010, 61, 291–315. [Google Scholar] [CrossRef]
- Adamski, N.M.; Anastasiou, E.; Eriksson, S.; O’Neill, C.M.; Lenhard, M. Local Maternal Control of Seed Size by Kluh/Cyp78a5-Dependent Growth Signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 20115–20120. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Wang, Z.; Cui, R.; Li, J.; Li, Y. Maternal Control of Seed Size by Eod3/Cyp78a6 in Arabidopsis Thaliana. Plant J. Cell Mol. Biol. 2012, 70, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Sotelo-Silveira, M.; Cucinotta, M.; Chauvin, A.L.; Chávez Montes, R.A.; Colombo, L.; Marsch-Martínez, N.; de Folter, S. Cytochrome P450 Cyp78a9 Is Involved in Arabidopsis Reproductive Development. Plant Physiol. 2013, 162, 779–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Fang, J.; Ou, S.; Gao, S.; Zhang, F.; Du, L.; Xiao, Y.; Wang, H.; Sun, X.; Chu, J.; et al. Variations in Cyp78a13 Coding Region Influence Grain Size and Yield in Rice. Plant Cell Environ. 2015, 38, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Liu, C.; Song, L.; Li, Y.; Li, M. Pacyp78a9, a Cytochrome P450, Regulates Fruit Size in Sweet Cherry (Prunus Avium L.). Front. Plant Sci. 2017, 8, 2076. [Google Scholar] [CrossRef] [Green Version]
- Zondlo, S.C.; Irish, V.F. Cyp78a5 Encodes a Cytochrome P450 That Marks the Shoot Apical Meristem Boundary in Arabidopsis. Plant J. 1999, 19, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Anastasiou, E.; Kenz, S.; Gerstung, M.; MacLean, D.; Timmer, J.; Fleck, C.; Lenhard, M. Control of Plant Organ Size by Kluh/Cyp78a5-Dependent Intercellular Signaling. Dev. Cell 2007, 13, 843–856. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.W.; Schwab, R.; Czech, B.; Mica, E.; Weigel, D. Dual Effects of Mir156-Targeted Spl Genes and Cyp78a5/Kluh on Plastochron Length and Organ Size in Arabidopsis Thaliana. Plant Cell 2008, 20, 1231–1243. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Cai, H.; Su, Z.; Wang, L.; Huang, X.; Zhang, M.; Chen, P.; Dai, X.; Zhao, H.; Palanivelu, R.; et al. Klu Suppresses Megasporocyte Cell Fate through Swr1-Mediated Activation of Wrky28 Expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E526–E535. [Google Scholar] [CrossRef] [Green Version]
- Poretska, O.; Yang, S.; Pitorre, D.; Poppenberger, B.; Sieberer, T. Amp1 and Cyp78a5/7 Act through a Common Pathway to Govern Cell Fate Maintenance Inarabidopsis Thaliana. PLoS Genet. 2020, 16, e1009043. [Google Scholar] [CrossRef]
- Nobusawa, T.; Kamei, M.; Ueda, H.; Matsushima, N.; Yamatani, H.; Kusaba, M. Highly Pleiotropic Functions of Cyp78as and Amp1 Are Regulated in Non-Cell-Autonomous/Organ-Specific Manners. Plant Physiol. 2021, 186, 767–781. [Google Scholar] [CrossRef]
- Jiang, L.; Yoshida, T.; Stiegert, S.; Jing, Y.; Alseekh, S.; Lenhard, M.; Pérez-Alfocea, F.; Fernie, A.R. Multi-Omics Approach Reveals the Contribution of Klu to Leaf Longevity and Drought Tolerance. Plant Physiol. 2021, 185, 352–368. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Ma, M.; Wu, L.; Zhou, M.; Li, M.; Wu, B.; Li, L.; Liu, X.; Jing, R.; Chen, W.; et al. Modified Expression of Tacyp78a5 Enhances Grain Weight with Yield Potential by Accumulating Auxin in Wheat (Triticum Aestivum L.). Plant Biotechnol. J. 2021, 20, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.; Stransfeld, L.; Adamski, N.M.; Breuninger, H.; Lenhard, M. Kluh/Cyp78a5-Dependent Growth Signaling Coordinates Floral Organ Growth in Arabidopsis. Curr. Biol. 2010, 20, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalve, S.; De Vos, D.; Beemster, G.T. Leaf Development: A Cellular Perspective. Front. Plant Sci. 2014, 5, 362. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Kong, H.; Sun, Y.; Zhang, X.; Zhang, W.; Altman, N.; De Pamphilis, C.W.; Ma, H. Genome-Wide Analysis of the Cyclin Family in Arabidopsis and Comparative Phylogenetic Analysis of Plant Cyclin-Like Proteins. Plant Physiol. 2004, 135, 1084–1099. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.J.; Zou, W.S.; Wu, G.; Lin, H.H.; Xi, D.H. Tobacco Alpha-Expansin Expa4 Plays a Role in Nicotiana Benthamiana Defence against Tobacco Mosaic Virus. Planta 2018, 247, 355–368. [Google Scholar] [CrossRef]
- De Souza, A.; Hull, P.A.; Gille, S.; Pauly, M. Identification and Functional Characterization of the Distinct Plant Pectin Esterases Pae8 and Pae9 and Their Deletion Mutants. Planta 2014, 240, 1123–1138. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Wang, Q.; Li, Z.; Cheng, H.; Li, Z.; Liu, X.; Song, W.; Appels, R.; Zhao, H. Expression of Tacyp78a3, a Gene Encoding Cytochrome P450 Cyp78a3 Protein in Wheat (Triticum Aestivum L.), Affects Seed Size. Plant J. 2015, 83, 312–325. [Google Scholar]
- Kim, J.I.; Murphy, A.S.; Baek, D.; Lee, S.W.; Yun, D.J.; Bressan, R.A.; Narasimhan, M.L. Yucca6 over-expression Demonstrates Auxin Function in Delaying Leaf Senescence in Arabidopsis Thaliana. J. Exp. Bot. 2011, 62, 3981–3992. [Google Scholar] [CrossRef]
- Zhao, Y.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A Role for Flavin Monooxygenase-Like Enzymes in Auxin Biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef]
- Chandler, J.W. The Hormonal Regulation of Flower Development. J. Plant Growth Regul. 2010, 30, 242–254. [Google Scholar] [CrossRef]
- Vernoux, T.; Brunoud, G.; Farcot, E.; Morin, V.; Van den Daele, H.; Legrand, J.; Oliva, M.; Das, P.; Larrieu, A.; Wells, D.; et al. The Auxin Signalling Network Translates Dynamic Input into Robust Patterning at the Shoot Apex. Mol. Syst. Biol. 2011, 7, 508. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Du, Q.; Zhuo, C.; Qi, L.; Wang, H. Lbd29-Involved Auxin Signaling Represses Nac Master Regulators and Fiber Wall Biosynthesis. Plant Physiol. 2019, 181, 595–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelissen, H.; Gonzalez, N.; Inze, D. Leaf Growth in Dicots and Monocots: So Different yet So Alike. Curr. Opin. Plant Biol. 2016, 33, 72–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, K.; Ahn, B.O.; Kawakatsu, T.; Ito, Y.; Itoh, J.; Nagato, Y.; Kurata, N. Plastochron1, a Timekeeper of Leaf Initiation in Rice, Encodes Cytochrome P450. Proc. Natl. Acad. Sci. USA 2004, 101, 875–880. [Google Scholar] [CrossRef] [Green Version]
- Imaishi, H.; Matsuo, S.; Swai, E.; Ohkawa, H. Cyp78a1 Preferentially Expressed in Developing Inflorescences of Zea Mays Encoded a Cytochrome P450-Dependent Lauric Acid 12-Monooxygenase. Biosci. Biotechnol. Biochem. 2000, 64, 1696–1701. [Google Scholar] [CrossRef]
- Yang, W.; Gao, M.; Yin, X.; Liu, J.; Xu, Y.; Zeng, L.; Li, Q.; Zhang, S.; Wang, J.; Zhang, X.; et al. Control of Rice Embryo Development, Shoot Apical Meristem Maintenance, and Grain Yield by a Novel Cytochrome P450. Mol. Plant 2013, 6, 1945–1960. [Google Scholar] [CrossRef] [Green Version]
- Santner, A.; Calderon-Villalobos, L.I.; Estelle, M. Plant Hormones Are Versatile Chemical Regulators of Plant Growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef]
- Mimura, M.; Nagato, Y.; Itoh, J. Rice Plastochron Genes Regulate Leaf Maturation Downstream of the Gibberellin Signal Transduction Pathway. Planta 2012, 235, 1081–1089. [Google Scholar] [CrossRef]
- Sun, X.; Cahill, J.; Van Hautegem, T.; Feys, K.; Whipple, C.; Novák, O.; Delbare, S.; Versteele, C.; Demuynck, K.; De Block, J.; et al. Altered Expression of Maize Plastochron1 Enhances Biomass and Seed Yield by Extending Cell Division Duration. Nat. Commun. 2017, 8, 14752. [Google Scholar] [CrossRef] [Green Version]
- Calderini, D.F.; Castillo, F.M.; Arenas-M, A.; Molero, G.; Reynolds, M.P.; Craze, M.; Bowden, S.; Milner, M.J.; Wallington, E.J.; Dowle, A.; et al. Overcoming the Trade-Off between Grain Weight and Number in Wheat by the Ectopic Expression of Expansin in Developing Seeds Leads to Increased Yield Potential. New Phytol. 2021, 230, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-Mediated Transformation of Arabidopsis Thaliana Using the Floral Dip Method. Nat. Protocols 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative Expression Software Tool (Rest) for Group-Wise Comparison and Statistical Analysis of Relative Expression Results in Real-Time Pcr. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Cui, F.; Zhao, C.; Zhang, W.; Yang, L.; Zhao, X.; Han, J.; Su, Q.; Ji, J.; Zhao, Z.; et al. Qtls for Flag Leaf Size and Their Influence on Yield-Related Traits in Wheat (Triticum Aestivum L.). Mol. Breed. 2015, 35, 24. [Google Scholar] [CrossRef]
- Disch, S.; Anastasiou, E.; Sharma, V.K.; Laux, T.; Fletcher, J.C.; Lenhard, M. The E3 Ubiquitin Ligase Big Brother Controls Arabidopsis Organ Size in a Dosage-Dependent Manner. Curr. Biol. 2006, 16, 272–279. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Peng, H.; Wu, L.; Li, M.; Guo, L.; Chen, H.; Wu, B.; Liu, X.; Zhao, H.; Li, W.; et al. TaKLU Plays as a Time Regulator of Leaf Growth via Auxin Signaling. Int. J. Mol. Sci. 2022, 23, 4219. https://doi.org/10.3390/ijms23084219
Zhou M, Peng H, Wu L, Li M, Guo L, Chen H, Wu B, Liu X, Zhao H, Li W, et al. TaKLU Plays as a Time Regulator of Leaf Growth via Auxin Signaling. International Journal of Molecular Sciences. 2022; 23(8):4219. https://doi.org/10.3390/ijms23084219
Chicago/Turabian StyleZhou, Mengdie, Haixia Peng, Linnan Wu, Mengyao Li, Lijian Guo, Haichao Chen, Baowei Wu, Xiangli Liu, Huixian Zhao, Wenqiang Li, and et al. 2022. "TaKLU Plays as a Time Regulator of Leaf Growth via Auxin Signaling" International Journal of Molecular Sciences 23, no. 8: 4219. https://doi.org/10.3390/ijms23084219
APA StyleZhou, M., Peng, H., Wu, L., Li, M., Guo, L., Chen, H., Wu, B., Liu, X., Zhao, H., Li, W., & Ma, M. (2022). TaKLU Plays as a Time Regulator of Leaf Growth via Auxin Signaling. International Journal of Molecular Sciences, 23(8), 4219. https://doi.org/10.3390/ijms23084219