
A Novel Selenium Polysaccharide Alleviates the Manganese (Mn)-Induced Toxicity in Hep G2 Cells and Caenorhabditis elegans


 ,
, 

Abstract
1. Introduction
2. Results
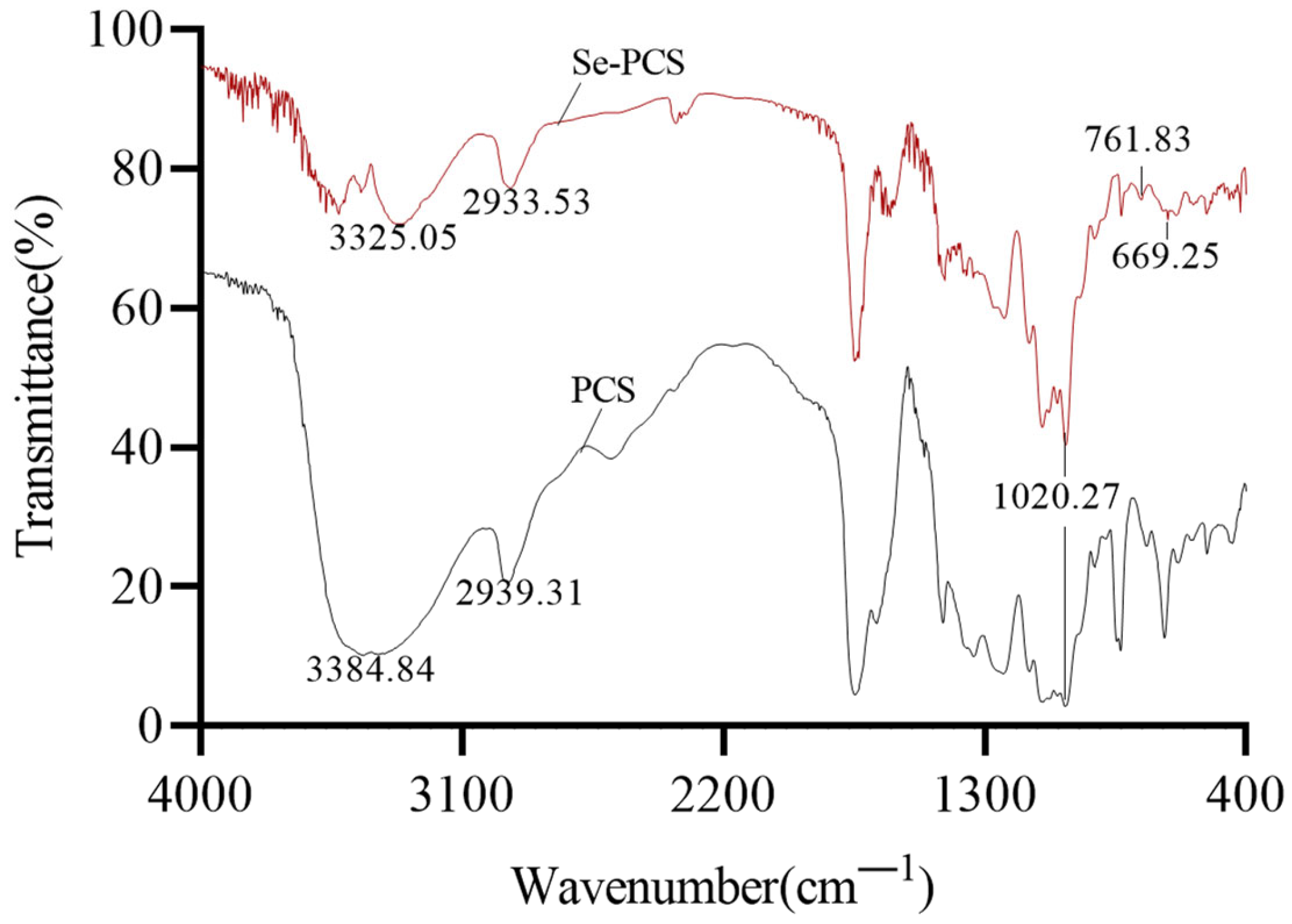
2.1. Chemical Properties of Se-PCS
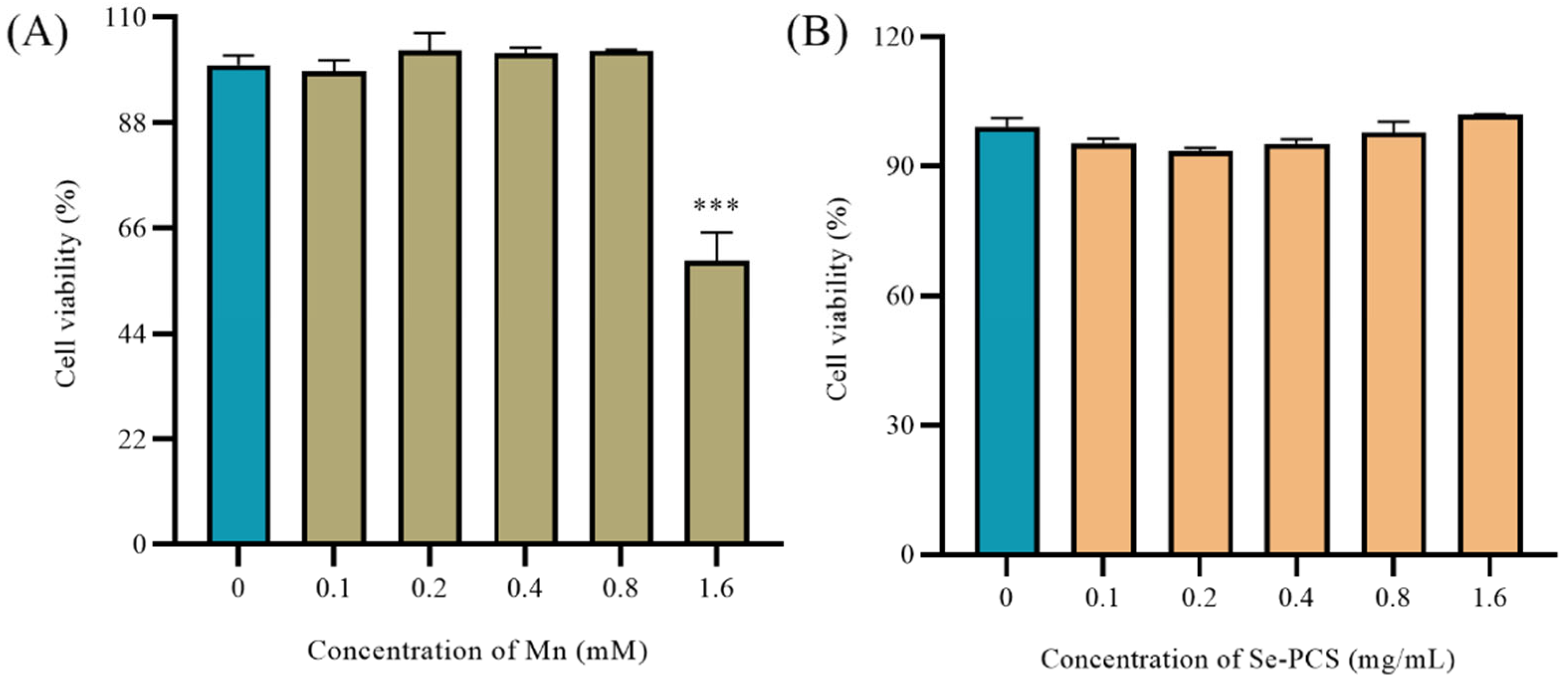
2.2. The Toxicity of Se-PCS on Hep G2 Cells
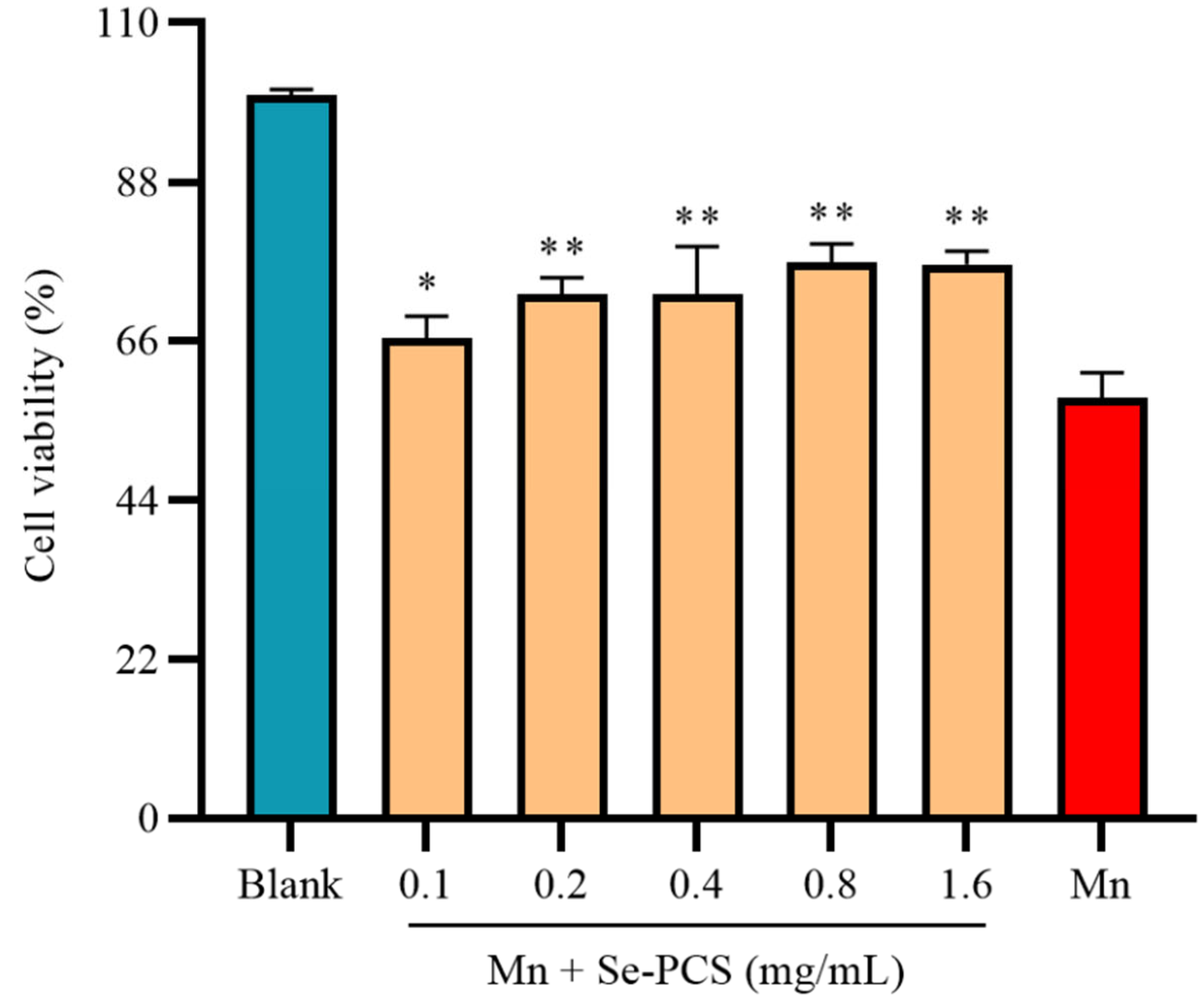
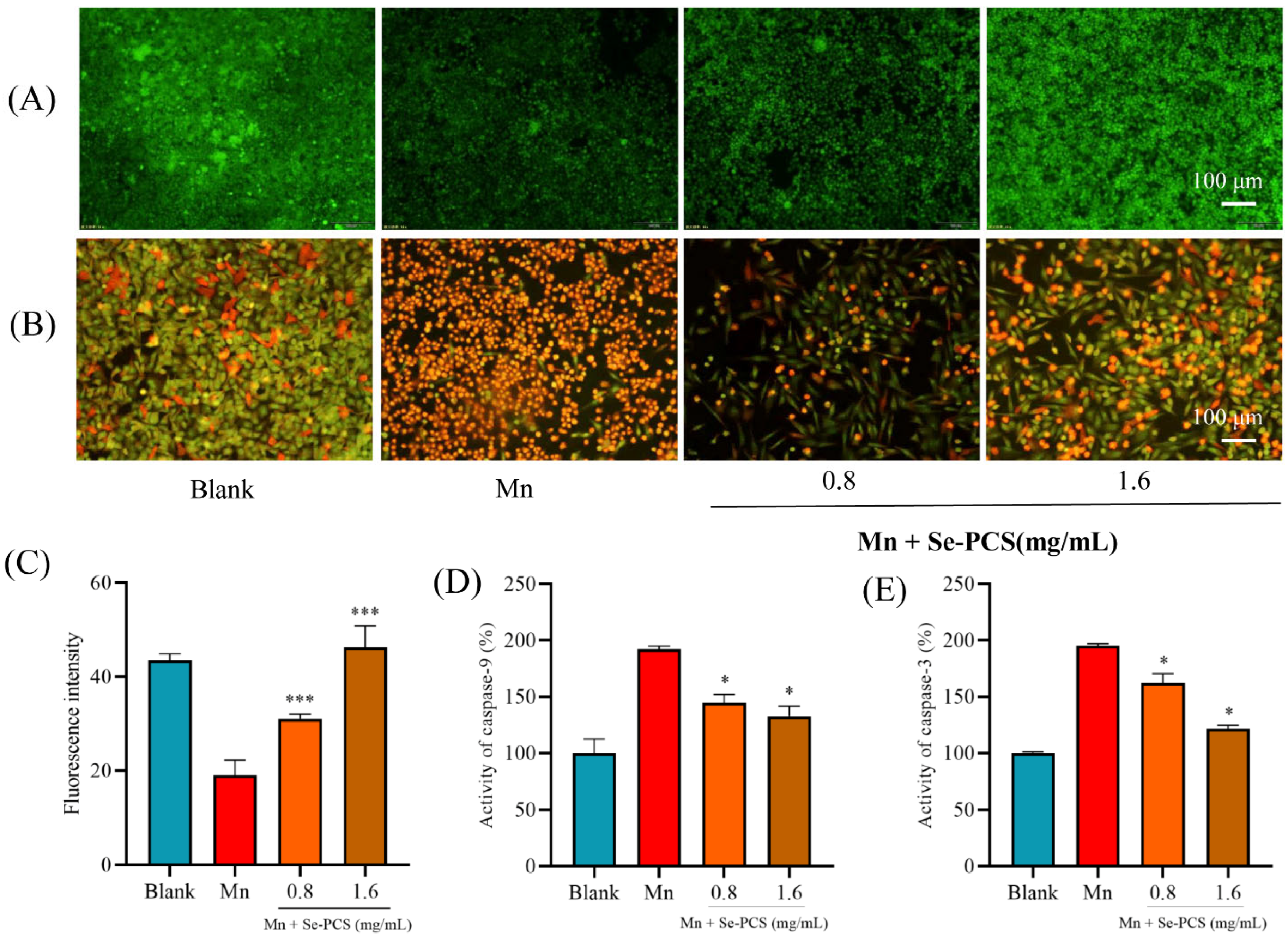
2.3. Se-PCS Protected Hep G2 Cells under Mn Stress
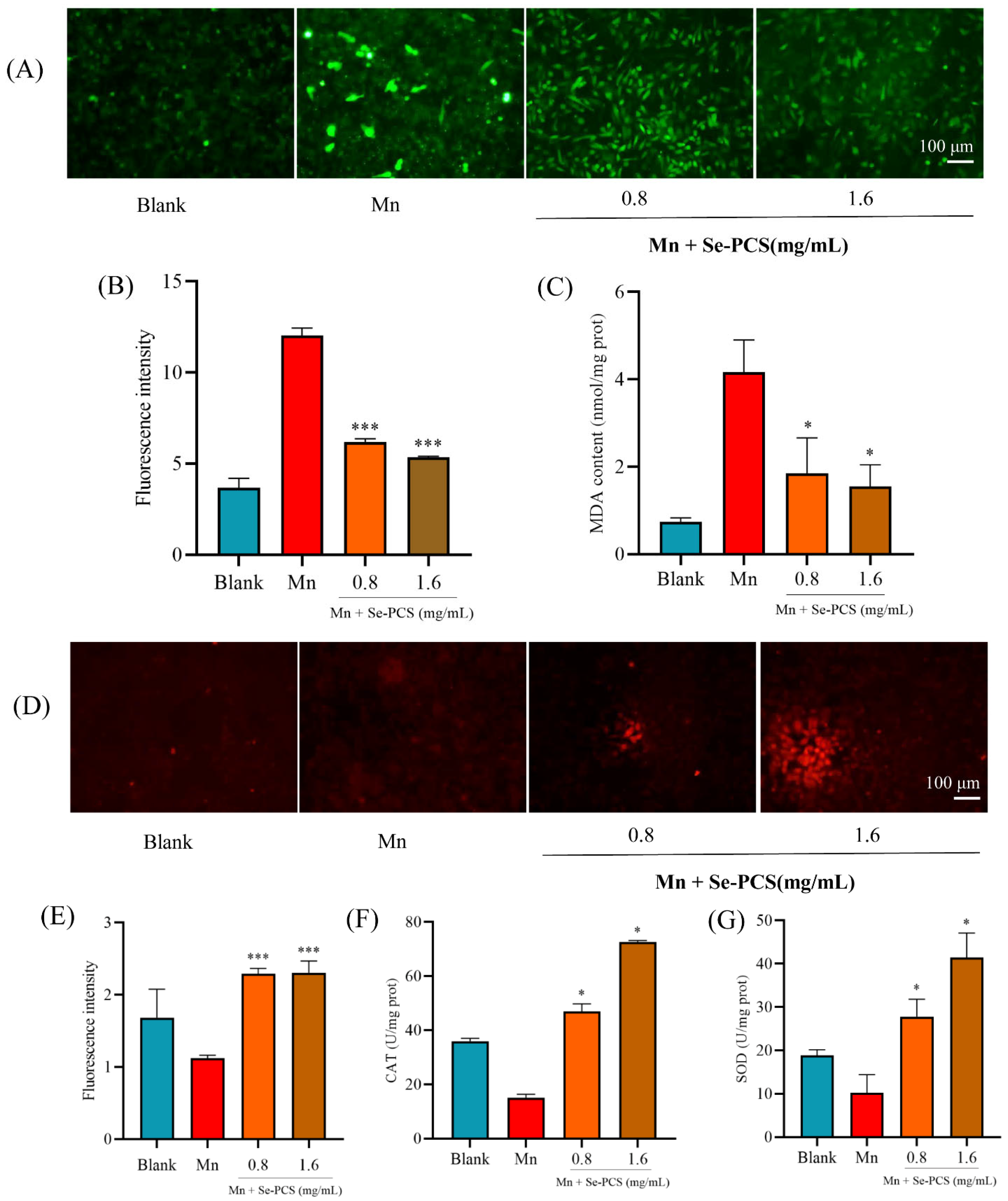
2.4. Se-PCS Reduced Oxidative Damage in Hep G2 Cells
2.5. Se-PCS Enhanced the Antioxidant System in Hep G2 Cells
2.6. Se-PCS Reduced Cell Apoptosis under Mn Stress
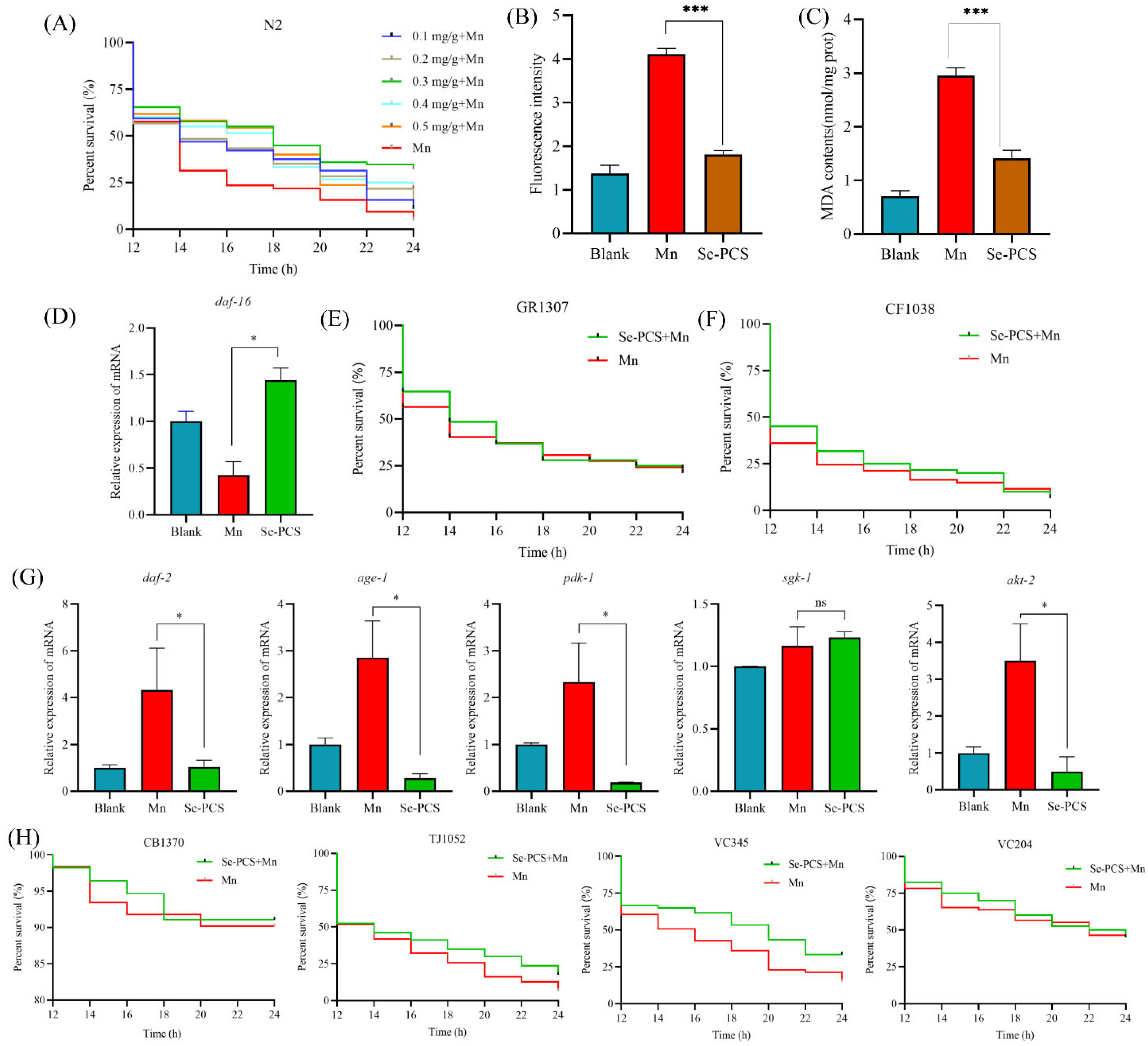
2.7. Se-PCS Protected C. elegans under Mn Stress
2.8. Se-PCS Decreased the ROS Level and MDA Content in C. elegans
2.9. Se-PCS Activated daf-16 Gene in C. elegans
2.10. Se-PCS Regulated IIS Pathway in C. elegans
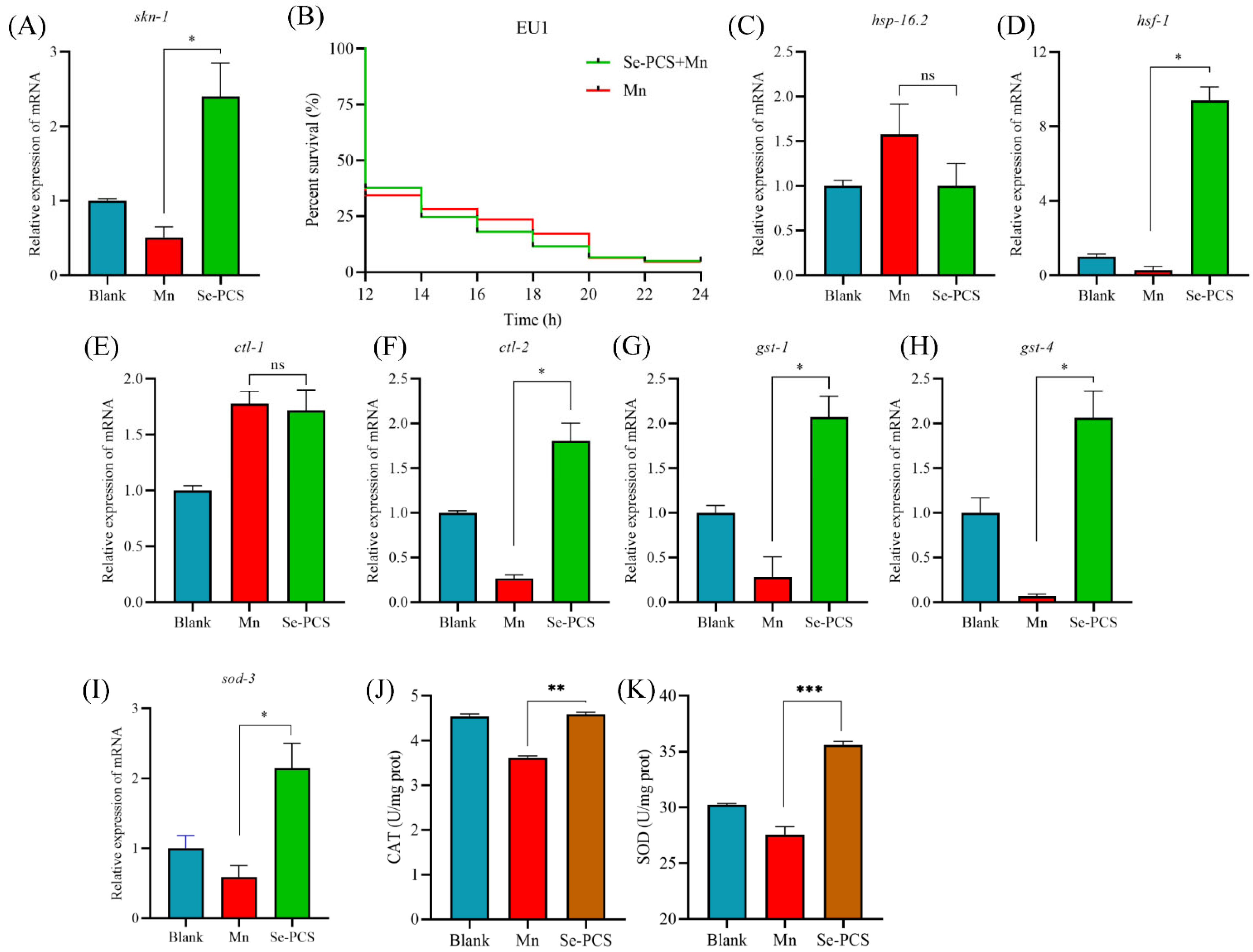
2.11. Se-PCS Activated skn-1 Gene in C. elegans
2.12. Se-PCS Increased the Defense System in C. elegans
3. Discussion
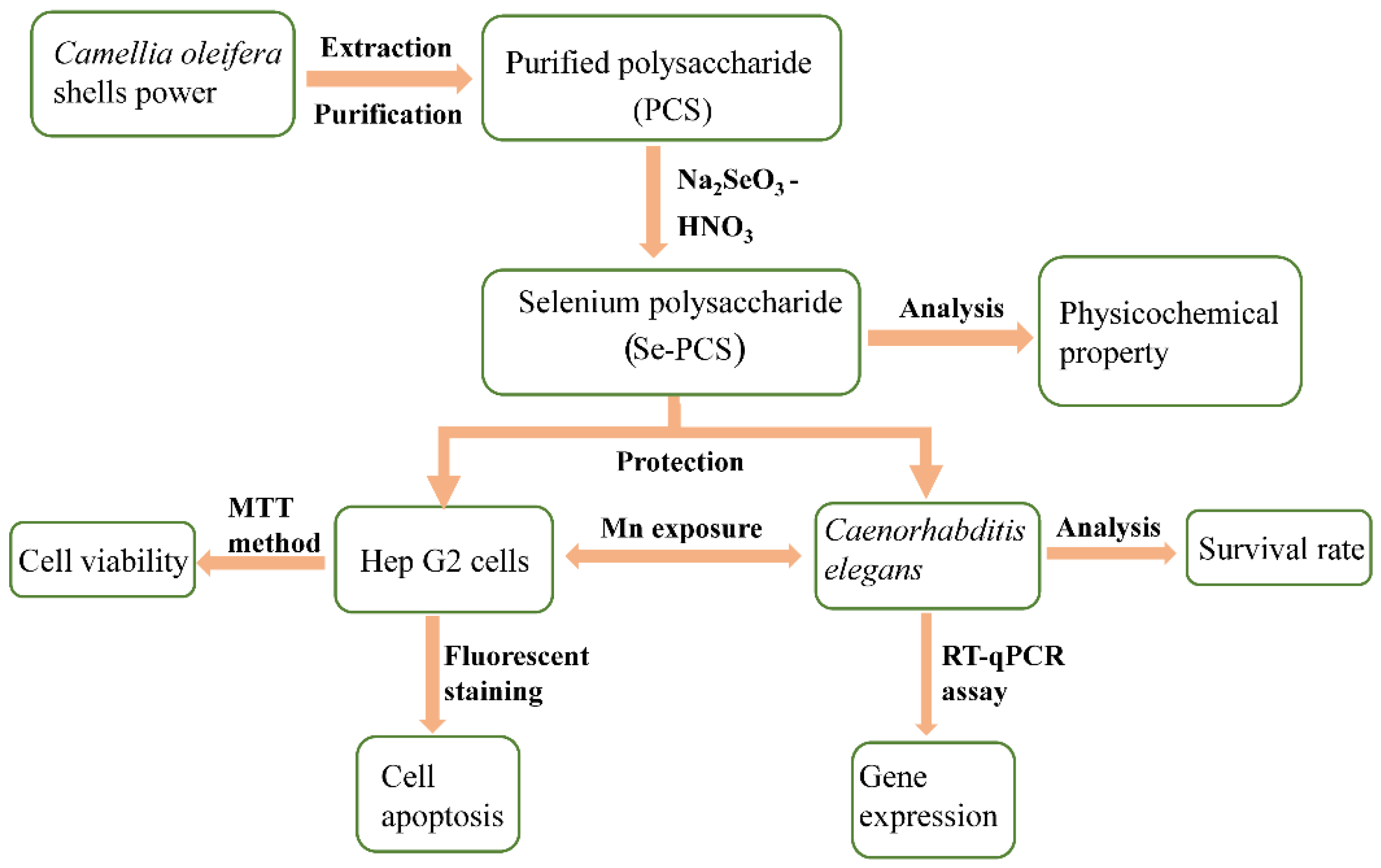
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation of Se-PCS
4.3. Structural Characterization and Chemical Properties of Se-PCS
4.4. Determination of Monosaccharide Composition of Se-PCS
4.5. Determination of Protection of Se-PCS on Hep G2 Cells
4.5.1. Maintenance of Cells
4.5.2. Determination of the Toxicity of Se-PCS
4.5.3. Determination of Protection of Se-PCS on Cells under Mn Stress
4.5.4. Fluorescent Staining
4.5.5. Determination of the Activities of SOD, CAT, Caspase-3, and Caspase-9 and the Content of MDA
4.6. Determination of Protection of Se-PCS on C. elegans
4.6.1. Maintenance of C. elegans
4.6.2. Determination of the Protection of Se-PCS in C. elegans under Mn Stress
4.6.3. Determination of the ROS Levels in C. elegans
4.6.4. Determination of the Activities of SOD and CAT and the Content of MDA
4.6.5. RT-qPCR Assay
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takeda, A. Manganese action in brain function. Brain Res. Rev. 2003, 41, 79–87. [Google Scholar] [CrossRef]
- Li, S.J.; Li, Y.; Chen, J.W.; Yuan, Z.X.; Mo, Y.H.; Lu, G.D.; Jiang, Y.M.; Ou, C.Y.; Wang, F.; Huang, X.W.; et al. Sodium para-aminosalicylic acid protected primary cultured basal ganglia neurons of rat from manganese-induced oxidative impairment and changes of amino acid neurotransmitters. Biol. Trace Elem. Res. 2015, 170, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Alaimo, A.; Gorojod, R.M.; Beauquis, J.; Muñoz, M.J.; Saravia, F.; Kotler, M.L. Deregulation of mitochondria-shaping proteins opa-1 and drp-1 in manganese-induced apoptosis. PLoS ONE 2014, 9, e91848. [Google Scholar] [CrossRef] [PubMed]
- Sfriso, A.; Marcomini, A. Macrophyte production in a shallow coastal lagoon. Part II: Coupling with sediment, SPM and tissue carbon, nitrogen and phosphorus concentrations. Mar. Environ. Res. 1999, 47, 285–309. [Google Scholar] [CrossRef]
- Criswell, S.R.; Perlmutter, J.S.; Huang, J.L.; Golchin, N.; Flores, H.P.; Hobson, A.; Aschner, M.; Erikson, K.M.; Checkoway, H.; Racette, B.A. Basal ganglia intensity indices and diffusion weighted imaging in manganese-exposed welders. Occup. Environ. Med. 2012, 69, 437–443. [Google Scholar] [CrossRef]
- Jiang, Y.; Zheng, W.; Long, L.; Zhao, W.; Li, X.; Mo, X.; Lu, J.; Fu, X.; Li, W.; Liu, S.; et al. Brain magnetic resonance imaging and manganese concentrations in red blood cells of smelting workers: Search for biomarkers of manganese exposure. Neurotoxicology 2007, 28, 126–135. [Google Scholar] [CrossRef]
- O’Neal, S.L.; Zheng, W. Manganese toxicity upon overexposure: A decade in review. Curr. Environ. Health Rep. 2015, 2, 315–328. [Google Scholar] [CrossRef]
- Li, Q.; Liu, H.; Alattar, M.; Jiang, S.; Han, J.; Ma, Y.; Jiang, C. The preferential accumulation of heavy metals in different tissues following frequent respiratory exposure to PM2.5 in rats. Sci. Rep. 2015, 5, 16936. [Google Scholar] [CrossRef]
- Maffeo, E.; Montuschi, A.; Stura, G.; Giordana, M.T. Chronic acquired hepatocerebral degeneration, pallidal T1 MRI hyperintensity and manganese in a series of cirrhotic patients. Neurol. Sci. 2014, 35, 523–530. [Google Scholar] [CrossRef]
- Beckett, G.J.; Arthur, J.R. Selenium and endocrine systems. J. Endocrinol. 2005, 184, 455–465. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, Y.; He, X.; Wei, X. Preparation, structural characterization and bioactivities of Se-containing polysaccharide: A review. Int. J. Biol. Macromol. 2018, 120, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shen, B.; Nie, S.; Duan, Z.; Chen, K. A combination of selenium and polysaccharides: Promising therapeutic potential. Carbohydr. Polym. 2019, 206, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Chen, C.; Li, C.; Huang, Q.; Fu, X. Physicochemical characterization, antioxidant and hypoglycemic activities of selenized polysaccharides from Sargassum pallidum. Int. J. Biol. Macromol. 2019, 132, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liu, J.; Liu, A.; Wang, Y. Cytoprotective effect of selenium polysaccharide from Pleurotus ostreatus against H2O2-induced oxidative stress and apoptosis in PC12 cells. Arab. J. Chem. 2022, 15, 103686. [Google Scholar] [CrossRef]
- Zhang, S.; Li, X.-Z. Inhibition of α-glucosidase by polysaccharides from the fruit hull of Camellia oleifera Abel. Carbohydr. Polym. 2015, 115, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Jin, X. Bioactivities of water-soluble polysaccharides from fruit shell of Camellia oleifera Abel: Antitumor and antioxidant activities. Carbohydr. Polym. 2012, 87, 2198–2201. [Google Scholar] [CrossRef]
- Zhou, L.; Luo, S.; Li, J.; Zhou, Y.; Wang, X.; Kong, Q.; Chen, T.; Feng, S.; Yuan, M.; Ding, C. Optimization of the extraction of polysaccharides from the shells of Camellia oleifera and evaluation on the antioxidant potential in vitro and in vivo. J. Funct. Foods 2021, 86, 104678. [Google Scholar] [CrossRef]
- Zhou, N.; Long, H.; Wang, C.; Zhu, Z.; Yu, L.; Yang, W.; Ren, X.; Liu, X. Characterization of selenium-containing polysaccharide from Spirulina platensis and its protective role against Cd-induced toxicity. Int. J. Biol. Macromol. 2020, 164, 2465–2476. [Google Scholar] [CrossRef]
- Huang, R.; Shen, M.; Yu, Y.; Liu, X.; Xie, J. Physicochemical characterization and immunomodulatory activity of sulfated Chinese yam polysaccharide. Int. J. Biol. Macromol. 2020, 165, 635–644. [Google Scholar] [CrossRef]
- Feng, S.; Cheng, H.; Fu, L.; Ding, C.; Zhang, L.; Yang, R.; Zhou, Y. Ultrasonic-assisted extraction and antioxidant activities of polysaccharides from Camellia oleifera leaves. Int. J. Biol. Macromol. 2014, 68, 7–12. [Google Scholar] [CrossRef]
- Li, Q.; Wang, W.; Zhu, Y.; Chen, Y.; Zhang, W.; Yu, P.; Mao, G.; Zhao, T.; Feng, W.; Yang, L.; et al. Structural elucidation and antioxidant activity a novel Se-polysaccharide from Se-enriched Grifola frondosa. Carbohydr. Polym. 2017, 161, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Adeline, S.I.; Victor, C.; Mihai, P.S.; Alina, M.; Mircea, N.; Stefan, B.E.; Adrian, S.S.; Caterina, F. Bioconcentration of essential and nonessential elements in Black Sea Turbot (psetta maxima maeotica linnaeus, 1758) in relation to fish gender. J. Mar. Sci. Eng. 2019, 7, 466. [Google Scholar] [CrossRef]
- Fu, X.; Chen, S.; Wang, X.; Shen, Y.; Zeng, R.; Wu, Q.; Lu, Y.; Shi, J.; Zhou, S. Dendrobium nobile Lindl. alkaloids alleviate Mn-induced neurotoxicity via PINK1/Parkin-mediated mitophagy in PC12 cells. Biochem. Biophys. Rep. 2021, 26, 100877. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, R.; Schaffitzel, E.; Hertweck, M. Endocrine signaling in Caenorhabditis elegans controls stress response and longevity. J. Endocrinol. 2006, 190, 191–202. [Google Scholar] [CrossRef]
- Samuelson, A.V.; Carr, C.E.; Ruvkun, G. Gene activities that mediate increased life span of C. elegans insulin-like signaling mutants. Genes Dev. 2007, 21, 2976–2994. [Google Scholar] [CrossRef]
- Zhou, L.; Luo, S.; Wang, X.; Zhou, Y.; Zhang, Y.; Zhu, S.; Chen, T.; Feng, S.; Yuan, M.; Ding, C. Blumea laciniata protected Hep G2 cells and Caenorhabditis elegans against acrylamide-induced toxicity via insulin/IGF-1 signaling pathway. Food Chem. Toxicol. 2021, 158, 112667. [Google Scholar] [CrossRef]
- Tullet, J.M.A.; Hertweck, M.; An, J.H.; Baker, J.; Hwang, J.Y.; Liu, S.; Oliveira, R.P.; Baumeister, R.; Blackwell, T.K. Direct inhibition of the longevity-promoting factor SKN-1 by insulin-like signaling in C. elegans. Cell 2008, 132, 1025–1038. [Google Scholar] [CrossRef]
- Bouabid, S.; Delaville, C.; Deurwaerdère, P.D.; Lakhdar-Ghazal, N.; Benazzouz, A. Manganese-induced atypical parkinsonism is associated with altered basal ganglia activity and changes in tissue levels of monoamines in the rat. PLoS ONE 2014, 9, e98952. [Google Scholar] [CrossRef]
- Xu, B.; Wu, S.W.; Lu, C.W.; Deng, Y.; Liu, W.; Wei, Y.G.; Yang, T.Y.; Xu, Z.F. Oxidative stress involvement in manganese-induced alpha-synuclein oligomerization in organotypic brain slice cultures. Toxicology 2013, 305, 71–78. [Google Scholar] [CrossRef]
- Ali, Z.; Yousafzai, A.M.; Sher, N.; Muhammad, I.; Nayab, G.E.; Aqeel, S.A.M.; Shah, S.T.; Aschner, M.; Khan, I.; Khan, H. Toxicity and bioaccumulation of manganese and chromium in different organs of common carp (Cyprinus carpio) fish. Toxicol. Rep. 2021, 8, 343–348. [Google Scholar] [CrossRef]
- Wua, F.; Yang, H.; Liu, Y.; Yang, X.; Xu, B.; Liu, W.; Xu, Z.; Deng, Y. Manganese exposure caused reproductive toxicity of male mice involving activation of GnRH secretion in the hypothalamus by prostaglandin E2 receptors EP1 and EP2. Ecotoxicol. Environ. Saf. 2020, 201, 110712. [Google Scholar] [CrossRef] [PubMed]
- Chtourou, Y.; Fetoui, H.; Sefi, M.; Trabelsi, K.; Barkallah, M.; Boudawara, T.; Kallel, H.; Zeghal, N. Silymarin, a natural antioxidant, protects cerebral cortex against manganese-induced neurotoxicity in adult rats. Biometals 2010, 23, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Nkpaa, K.W.; Onyeso, G.I.; Kponee, K.Z. Rutin abrogates manganese-Induced striatal and hippocampal toxicity via inhibition of iron depletion, oxidative stress, inflammation and suppressing the NF-κB signaling pathway. J. Trace Elem. Med. Biol. 2019, 53, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Neumann, C.; Baesler, J.; Steffen, G.; Nicolai, M.M.; Zubel, T.; Aschner, M.; Bürkle, A.; Mangerich, A.; Schwerdtle, T.; Bornhorst, J. The role of poly(ADP-ribose) polymerases in manganese exposed Caenorhabditis elegans. J. Trace Elem. Med. Biol. 2019, 57, 21–27. [Google Scholar] [CrossRef]
- Wollenhaupt, S.G.N.; Soares, A.T.; Salgueiro, W.G.; Noremberg, S.; Reis, G.; Viana, C.; Gubert, P.; Soares, F.A.; Affeldt, R.F.; Lüdtke, D.S.; et al. Seleno- and Telluro-xylofuranosides attenuate Mn-induced toxicity in C. elegans via the DAF-16/FOXO pathway. Food Chem. Toxicol. 2014, 64, 192–199. [Google Scholar] [CrossRef]
- Milatovic, D.; Zaja-Milatovic, S.; Gupta, R.C.; Yu, Y.; Aschner, M. Oxidative damage and neurodegeneration in manganese-induced neurotoxicity. Toxicol. Appl. Pharmacol. 2009, 240, 219–225. [Google Scholar] [CrossRef]
- Gunter, T.E.; Gavin, C.E.; Aschner, M.; Gunter, K.K. Speciation of manganese in cells and mitochondria: A search for the proximal cause of manganese neurotoxicity. Neurotoxicology 2006, 27, 765–776. [Google Scholar] [CrossRef]
- Facchinetti, F.; Dawson, V.L.; Dawson, T.M. Free radicals as mediators of neuronal injury. Cell. Mol. Neurobiol. 1998, 18, 667–682. [Google Scholar] [CrossRef]
- Latronico, T.; Branà, M.T.; Merra, E.; Fasano, A.; Bari, G.D.; Casalino, E.; Liuzzi, G.M. Impact of manganese neurotoxicity on MMP-9 production and superoxide dismutase activity in rat primary astrocytes. Effect of resveratrol and therapeutical implications for the treatment of CNS diseases. Toxicol. Sci. 2013, 135, 218–228. [Google Scholar] [CrossRef]
- Roth, J.A.; Eichhorn, M. Down-regulation of LRRK2 in control and DAT transfected HEK cells increases manganese-induced oxidative stress and cell toxicity. Neurotoxicology 2013, 37, 100–107. [Google Scholar] [CrossRef][Green Version]
- Yang, X.; Yang, H.; Wu, F.; Qi, Z.; Li, J.; Xu, B.; Liu, W.; Xu, Z.; Deng, Y. Mn inhibits GSH dynthesis via downregulation of neuronal EAAC1 and astrocytic xCT to cause oxidative damage in the striatum of mice. Oxid. Med. Cell. Longevity 2018, 2018, 4235695. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Shen, M.; Wen, P.; Hong, Y.; Liu, X.; Xie, J. Preparation, characterization, antioxidant activity and protective effect against cellular oxidative stress of phosphorylated polysaccharide from Cyclocarya paliurus. Food Chem. Toxicol. 2020, 145, 111754. [Google Scholar] [CrossRef] [PubMed]
- Bratton, S.B.; Salvesen, G.S. Regulation of the Apaf-1-caspase-9 apoptosome. J. Cell Sci. 2010, 123, 3209–3214. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, A.S.; Guarda, G.; D’Ombrain, M.C.; Drexler, S.K. Inflammatory caspases in innate immunity and inflammation. J. Innate Immun. 2010, 2, 228–237. [Google Scholar] [CrossRef]
- Scorrano, L. Opening the doors to cytochrome c: Changes in mitochondrial shape and apoptosis. Int. J. Biochem. Cell Biol. 2009, 41, 16. [Google Scholar] [CrossRef]
- Yassine, C.; Khaled, T.; Hamadi, F.; Ghada, M.; Kallel, H.; Najiba, Z. Manganese induces oxidative stress, redox state unbalance and disrupts membrane bound ATPases on murine neuroblastoma cells in vitro: Protective role of Silymarin. Neurochem. Res. 2011, 36, 1546–1557. [Google Scholar] [CrossRef]
- Maddirala, Y.; Tobwala, S.; Ercal, N. N-acetylcysteineamide protects against manganese-induced toxicity in SHSY5Y cell line. Brain Res. 2015, 1608, 157–166. [Google Scholar] [CrossRef]
- Evren, V.; Apaydin, M.; Khalilnezhad, A.; Erbas, O.; Taskiran, D. Protective effect of edaravone against manganese-induced toxicity in cultured rat astrocytes. Environ. Toxicol. Pharmacol. 2015, 40, 563–567. [Google Scholar] [CrossRef]
- Benedetto, A.; Au, C.; Aschner, M. Manganese-induced dopaminergic neurodegeneration: Insights into mechanisms and genetics shared with Parkinson’s disease. Cheminform 2010, 109, 4862–4884. [Google Scholar] [CrossRef]
- Vila, D.S.A.; Puntel, R.L.; Folmer, V.; Rocha, J.o.B.T.; Santos, A.P.M.d.; Aschner, M. Manganese Neurotoxicity; Springer Science+Business Media: New York, NY, USA, 2014. [Google Scholar] [CrossRef]
- Alexandre, B.; Catherine, A.; Silva, A.D.; Dejan, M.; Michael, A.; Kaveh, A. Extracellular dopamine potentiates Mn-induced oxidative stress, lifespan reduction, and dopaminergic neurodegeneration in a BLI-3–dependent manner in Caenorhabditis elegans. PLoS Genet. 2010, 6, e1001084. [Google Scholar] [CrossRef]
- Zheng, S.Q.; Huang, X.B.; Xing, T.K.; Ding, A.J.; Wu, G.S.; Luo, H.R. Chlorogenic acid extends the lifespan of Caenorhabditis elegans via Insulin/IGF-1 signaling pathway. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2017, 72, 464–472. [Google Scholar] [CrossRef]
- Boehm, M.; Slack, F. A developmental timing microRNA and its target regulate life span in C. elegans. Science 2005, 310, 1954–1957. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, V.N. Insulin/IGF-1 signaling pathway driving aging and cancer as a target for pharmacological intervention. Exp. Gerontol. 2003, 38, 1041–1049. [Google Scholar] [CrossRef]
- Cornes, E.; Porta-De-La-Riva, M.; Aristizábal-Corrales, D.; Brokate-Llanos, A.M.; García-Rodríguez, F.J.; Ertl, I.; Díaz, M.; Fontrodona, L.; Reis, K.; Johnsen, R.; et al. Cytoplasmic LSM-1 protein regulates stress responses through the insulin/IGF-1 signaling pathway in Caenorhabditis elegans. RNA 2015, 21, 1544–1553. [Google Scholar] [CrossRef]
- Li, H.; Yu, X.; Meng, F.; Zhao, Z.; Guan, S.; Wang, L. Ferulic acid supplementation increases lifespan and stress resistance via Insulin/IGF-1 signaling pathway in C. elegans. Int. J. Mol. Sci. 2021, 22, 4279. [Google Scholar] [CrossRef] [PubMed]
- Hertweck, M.; Gobel, C.; Baumeister, R.C. elegans SGK-1 is the critical component in the Akt/PKB Kinase complex to control stress response and life span. Dev. Cell 2004, 6, 577–588. [Google Scholar] [CrossRef]
- Tullet, J.M.A. DAF-16 target identification in C. elegans: Past, present and future. Biogerontology 2015, 16, 221–234. [Google Scholar] [CrossRef]
- Wolff, S.; Dillin, A. The trifecta of aging in Caenorhabditis elegans. Exp. Gerontol. 2006, 41, 894–903. [Google Scholar] [CrossRef]
- Lee, S.S.; Kennedy, S.; Tolonen, A.C.; Ruvkun, G. DAF-16 target genes that control C. elegans life-span and metabolism. Science 2003, 300, 644–647. [Google Scholar] [CrossRef]
- Strayer, A.; Wu, Z.; Christen, Y.; Link, C.D.; Luo, Y. Expression of the small heat-shock protein Hsp16-2 in Caenorhabditis elegans is suppressed by Ginkgo biloba extract EGb 761. FASEB J. 2003, 17, 2305–2307. [Google Scholar] [CrossRef]
- Soerensen, M.; Dato, S.; Tan, Q.; Thinggaard, M.; Kleindorp, R.; Beekman, M.; Jacobsen, R.; Suchiman, H.E.D.; Craen, A.J.M.d.; Westendorp, R.G.J.; et al. Human longevity and variation in GH/IGF-1/insulin signaling, DNA damage signaling and repair and pro/antioxidant pathway genes: Cross sectional and longitudinal studies. Exp. Gerontol. 2012, 47, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.C.; Yu, H.; Ma, T.H.; Tjong, W.Y.; Stern, A.; Chiu, D.T.Y. tert-Butyl Hydroperoxide (tBHP)-induced lipid peroxidation and embryonic defects resemble glucose-6-phosphate dehydrogenase (G6PD) deficiency in C. elegans. Int. J. Mol. Sci. 2020, 21, 8688. [Google Scholar] [CrossRef] [PubMed]
- Lü, H.; Gao, Y.; Shan, H.; Lin, Y. Preparation and antibacterial activity studies of degraded polysaccharide selenide from Enteromorpha prolifera. Carbohydr. Polym. 2014, 107, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Xi, X.; Wei, X.; Wang, Y.; Chu, Q.; Xiao, J. Determination of tea polysaccharides in Camellia sinensis by a modified phenol-sulfuric acid method. Arch. Biol. Sci. 2010, 62, 669–676. [Google Scholar] [CrossRef]
- Pierce, J.; Suelter, C.H. An evaluation of the Coomassie brilliant blue G-250 dye-binding method for quantitative protein determination. Anal. Biochem. 1977, 81, 478–480. [Google Scholar] [CrossRef]
- He, J.; Wu, Z.; Pan, D.; Guo, Y.; Zeng, X. Effect of selenylation modification on antitumor activity of peptidoglycan from Lactobacillus acidophilus. Carbohydr. Polym. 2017, 165, 344–350. [Google Scholar] [CrossRef]
- Porta-de-la-Riva, M.; Fontrodona, L.; Villanueva, A.; Cerón, J. Basic Caenorhabditis elegans methods: Synchronization and observation. Jove-J. Vis. Exp. 2012, 64, e4019. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Se-PCS |
|---|---|
| Molecular weight (kDa) | 62.23 |
| Carbohydrate (%) | 86.15 ± 0.37 |
| Protein (%) | 1.01 ± 0.04 |
| Se (μg/g) | 444.9 ± 1.029 |
| Monosaccharide composition | |
| Xylose (%) | 45.17 |
| Glucose (%) | 5.27 |
| Mannose (%) | 36.03 |
| Galactose (%) | 13.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Wang, X.; Yan, X.; Dai, Y.; Liang, T.; Zhou, L.; Feng, S.; Yuan, M.; Yang, H.; Ding, C. A Novel Selenium Polysaccharide Alleviates the Manganese (Mn)-Induced Toxicity in Hep G2 Cells and Caenorhabditis elegans. Int. J. Mol. Sci. 2022, 23, 4097. https://doi.org/10.3390/ijms23084097
Chen T, Wang X, Yan X, Dai Y, Liang T, Zhou L, Feng S, Yuan M, Yang H, Ding C. A Novel Selenium Polysaccharide Alleviates the Manganese (Mn)-Induced Toxicity in Hep G2 Cells and Caenorhabditis elegans. International Journal of Molecular Sciences. 2022; 23(8):4097. https://doi.org/10.3390/ijms23084097
Chicago/Turabian StyleChen, Tao, Xiaoju Wang, Xinchen Yan, Yali Dai, Tao Liang, Lijun Zhou, Shiling Feng, Ming Yuan, Hongyu Yang, and Chunbang Ding. 2022. "A Novel Selenium Polysaccharide Alleviates the Manganese (Mn)-Induced Toxicity in Hep G2 Cells and Caenorhabditis elegans" International Journal of Molecular Sciences 23, no. 8: 4097. https://doi.org/10.3390/ijms23084097
APA StyleChen, T., Wang, X., Yan, X., Dai, Y., Liang, T., Zhou, L., Feng, S., Yuan, M., Yang, H., & Ding, C. (2022). A Novel Selenium Polysaccharide Alleviates the Manganese (Mn)-Induced Toxicity in Hep G2 Cells and Caenorhabditis elegans. International Journal of Molecular Sciences, 23(8), 4097. https://doi.org/10.3390/ijms23084097