
First Trimester Prediction of Preterm Delivery in the Absence of Other Pregnancy-Related Complications Using Cardiovascular-Disease Associated MicroRNA Biomarkers



Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics of Patients and Controls
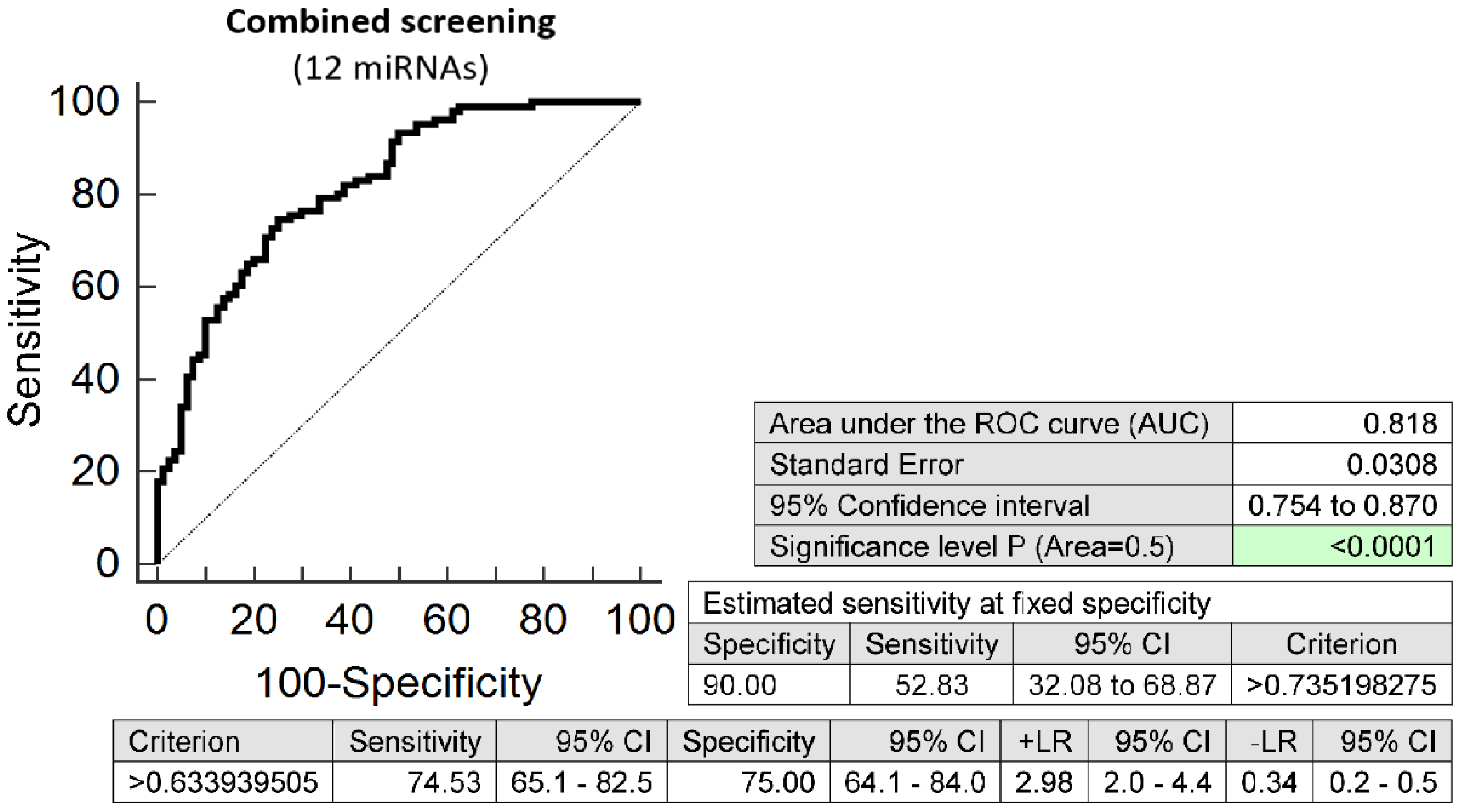
2.2. Dysregulated Expression Profile of Cardiovascular Disease Associated MicroRNAs in Early Stages of Gestation Is Able to Predict Preterm Delivery
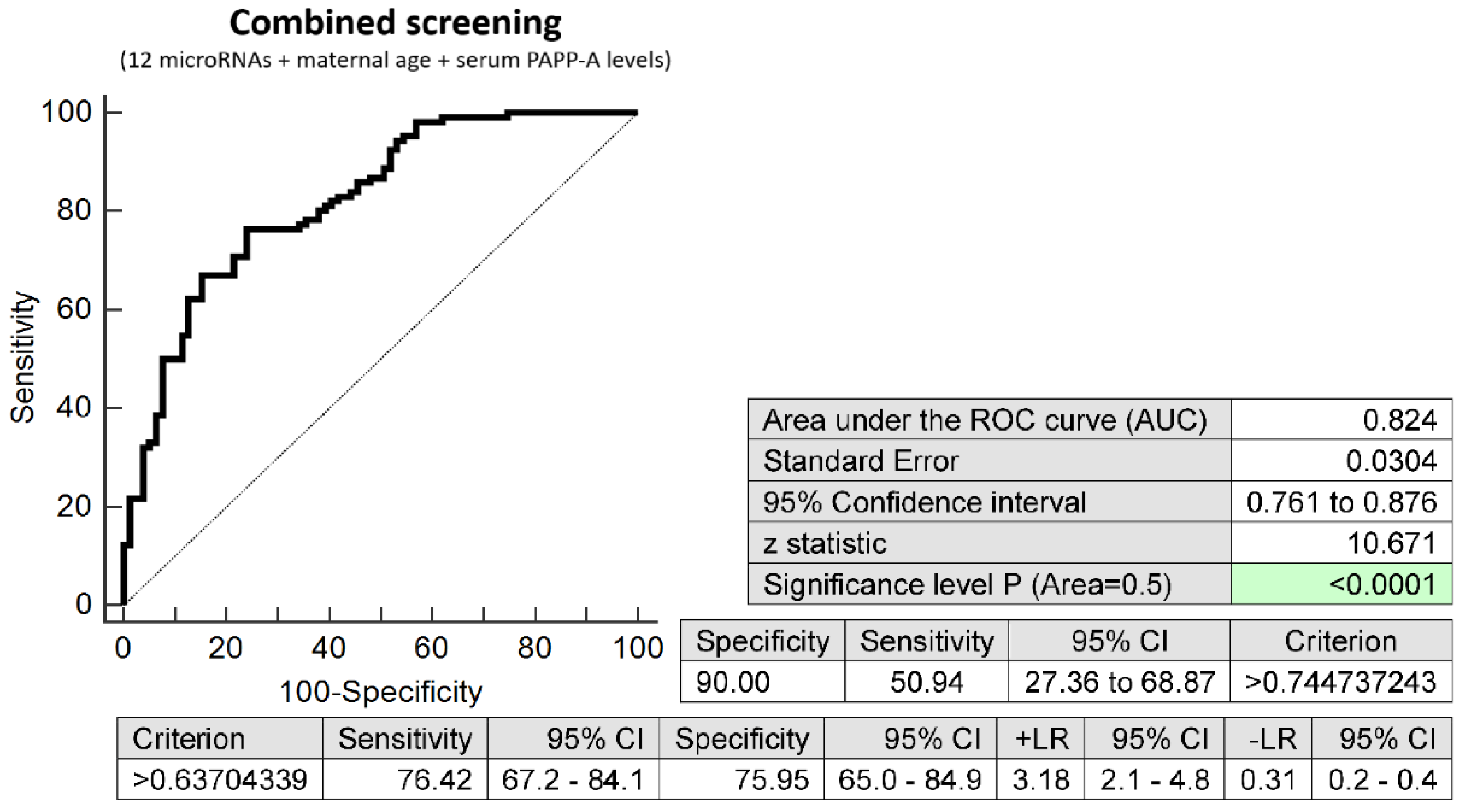
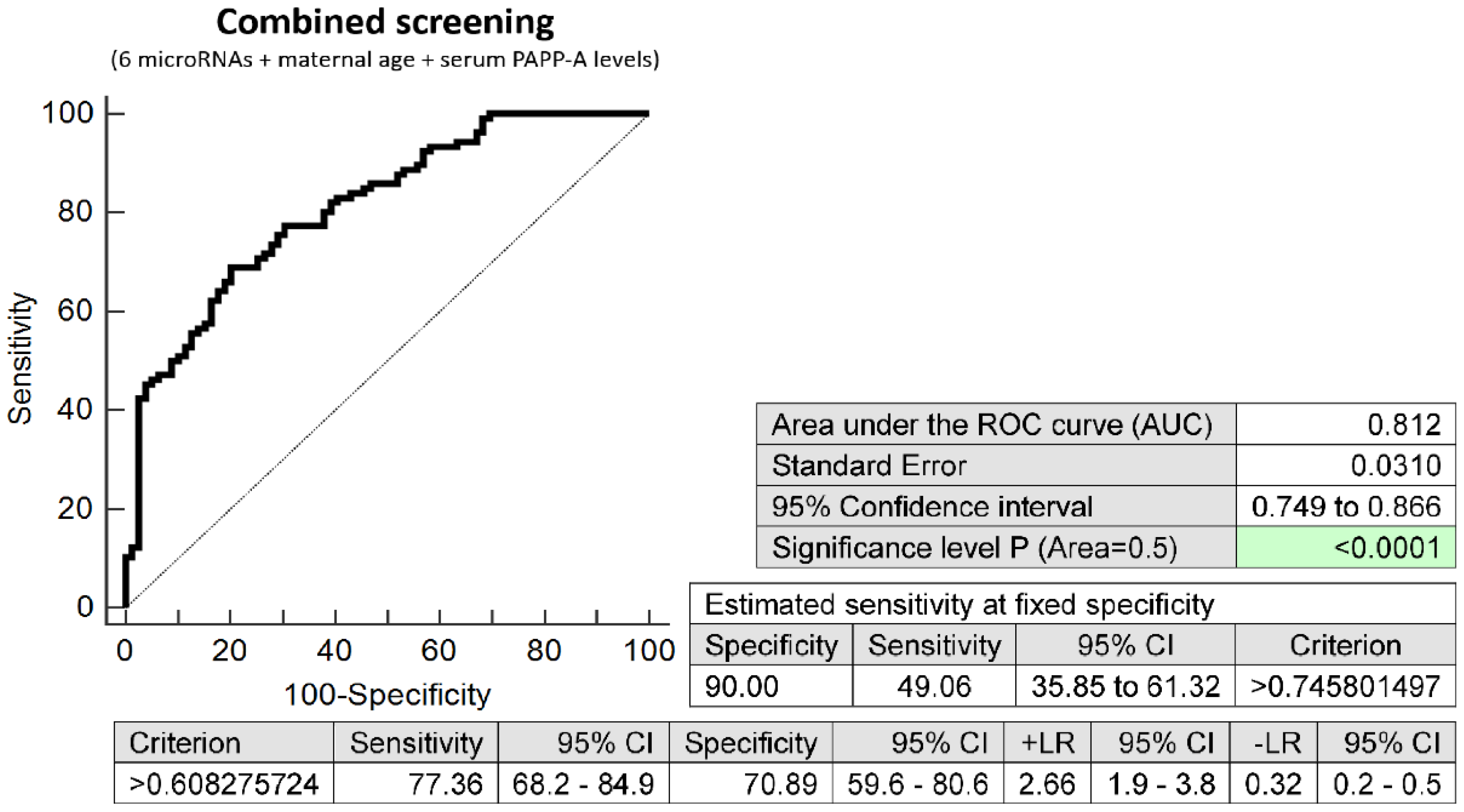
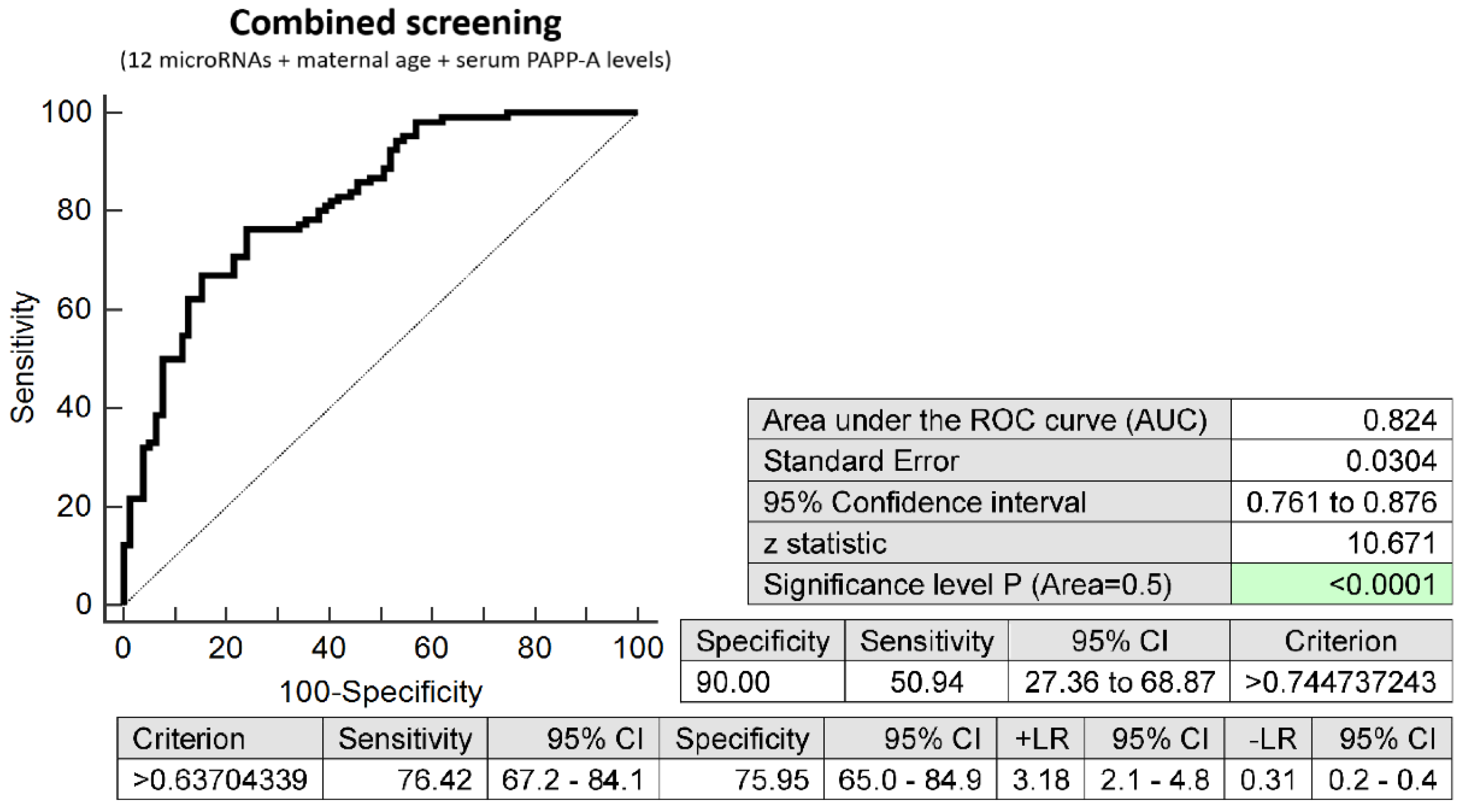
2.3. Prediction Model of Preterm Delivery Based on the Combination of MicroRNA Biomarkers, Maternal Age at Sampling and Serum PAPP-A Levels (IU/L)
2.4. Dysregulated Expression Profile of Cardiovascular Disease Associated MicroRNAs in Early Stages of Gestation with Respect to the Gestational Age of the Delivery
2.5. Dysregulated Expression Profile of Cardiovascular Disease Associated MicroRNAs in Early Stages of Gestation with Respect to the Type of Preterm Delivery
2.6. Information on MicroRNA-Gene-Biological Pathways Interactions
3. Discussion
4. Materials and Methods
4.1. Patients Cohort
4.2. Combined First Trimester Risk Analysis
4.3. Processing of Samples
4.4. Statistical Analysis
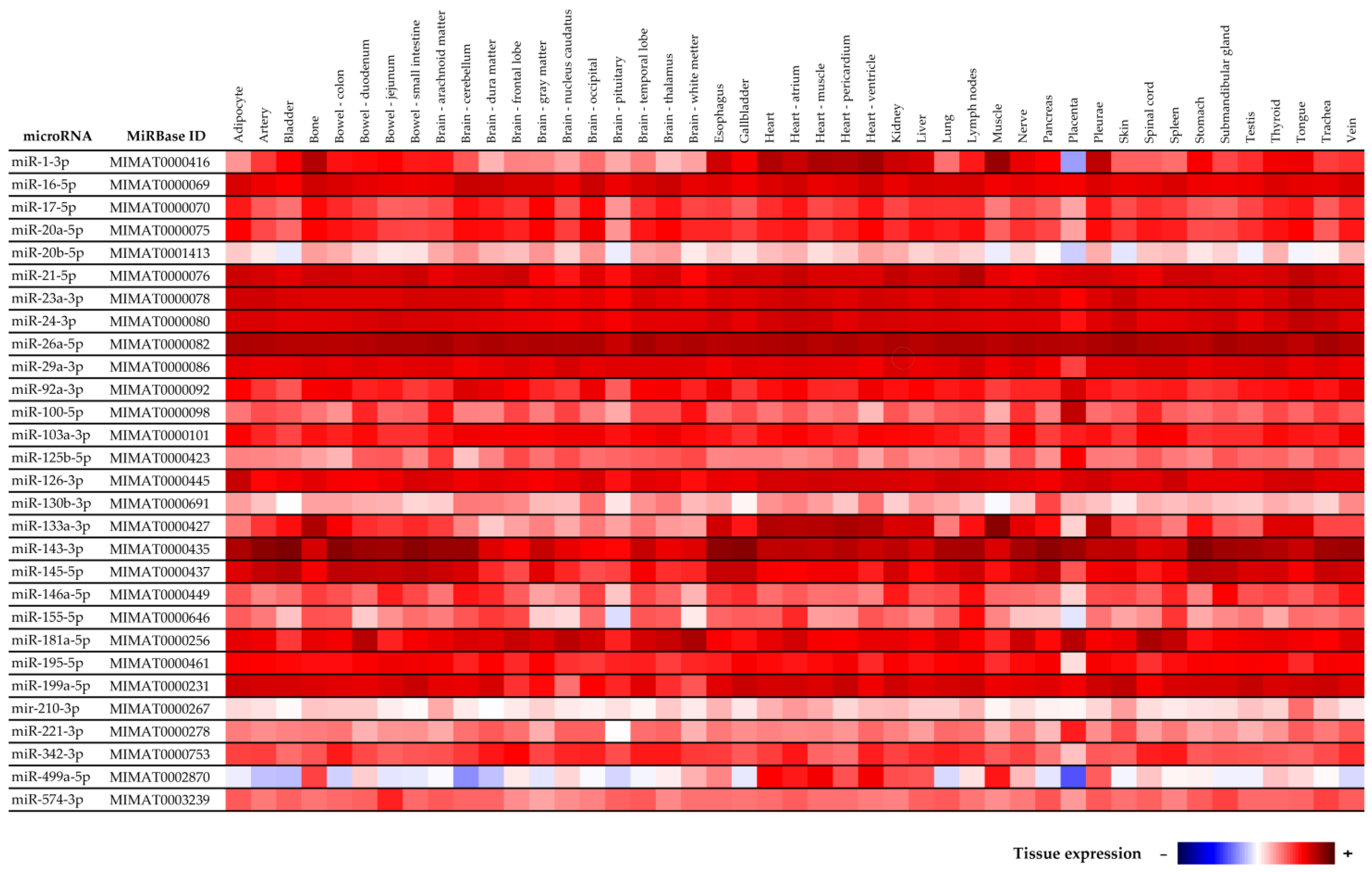
4.5. Tissue Specificity of MicroRNAs
4.6. Information on MicroRNA-Gene-Biological Pathways Interactions
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- Moutquin, J.M.; Milot Roy, V.; Irion, O. Preterm prevention: Effectivenss of current strategies. J. Soc. Obstet. Gynaecol. Can. 1996, 18, 571–588. [Google Scholar] [CrossRef]
- Romero, R.; Espinoza, J.; Kusanovic, J.P.; Gotsch, F.; Hassan, S.; Erez, O.; Chaiworapongsa, T.; Mazor, M. The preterm parturition syndrome. BJOG 2006, 113, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Hromadnikova, I.; Kotlabova, K.; Krofta, L. A History of Preterm Delivery Is Associated with Aberrant Postpartal MicroRNA Expression Profiles in Mothers with an Absence of Other Pregnancy-Related Complications. Int. J. Mol. Sci. 2021, 22, 4033. [Google Scholar] [CrossRef]
- Tanz, L.J.; Stuart, J.J.; Williams, P.L.; Rimm, E.B.; Missmer, S.A.; Rexrode, K.M.; Mukamal, K.J.; Rich-Edwards, J.W. Preterm Delivery and Maternal Cardiovascular Disease in Young and Middle-Aged Adult Women. Circulation 2017, 135, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Catov, J.M.; Dodge, R.; Barinas-Mitchell, E.; Sutton-Tyrrell, K.; Yamal, J.M.; Piller, L.B.; Ness, R.B. Prior preterm birth and maternal subclinical cardiovascular disease 4 to 12 years after pregnancy. J. Womens Health 2013, 22, 835–843. [Google Scholar] [CrossRef] [Green Version]
- Catov, J.M.; Lewis, C.E.; Lee, M.; Wellons, M.F.; Gunderson, E.P. Preterm birth and future maternal blood pressure, inflam-mation, and intimal-medial thickness: The CARDIA study. Hypertension 2013, 61, 641–646. [Google Scholar] [CrossRef] [Green Version]
- Perng, W.; Stuart, J.; Rifas-Shiman, S.L.; Rich-Edwards, J.W.; Stuebe, A.; Oken, E. Preterm birth and long-term maternal car-diovascular health. Ann. Epidemiol. 2015, 25, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; An, S.; Niu, J.; Zhao, H.; Wang, Y.; Wu, S.; Yang, X. Effect of premature birth on long-term systolic blood pressure variability in women. Anatol. J. Cardiol. 2018, 20, 347–353. [Google Scholar] [CrossRef]
- Haas, D.M.; Parker, C.B.; Marsh, D.J.; Grobman, W.A.; Ehrenthal, D.B.; Greenland, P.; Bairey Merz, C.N.; Pemberton, V.L.; Silver, R.M.; Barnes, S.; et al. NHLBI nuMoM2b Heart Health Study. Association of Adverse Pregnancy Outcomes with Hypertension 2 to 7 Years Postpartum. J. Am. Heart Assoc. 2019, 8, e013092. [Google Scholar] [CrossRef]
- Tanz, L.J.; Stuart, J.J.; Williams, P.L.; Missmer, S.A.; Rimm, E.B.; James-Todd, T.M.; Rich-Edwards, J.W. Preterm Delivery and Maternal Cardiovascular Disease Risk Factors: The Nurses’ Health Study II. J. Womens Health 2019, 28, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Catov, J.M.; Snyder, G.G.; Fraser, A.; Lewis, C.E.; Liu, K.; Althouse, A.D.; Bertolet, M.; Gunderson, E.P. Blood Pressure Patterns and Subsequent Coronary Artery Calcification in Women Who Delivered Preterm Births. Hypertension 2018, 72, 159–166. [Google Scholar] [CrossRef] [PubMed]
- James-Todd, T.M.; Karumanchi, S.A.; Hibert, E.L.; Mason, S.M.; Vadnais, M.A.; Hu, F.B.; Rich-Edwards, J.W. Gestational age, infant birth weight, and subsequent risk of type 2 diabetes in mothers: Nurses’ Health Study II. Prev. Chronic Dis. 2013, 10, E156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lykke, J.A.; Paidas, M.J.; Damm, P.; Triche, E.W.; Kuczynski, E.; Langhoff-Roos, J. Preterm delivery and risk of subsequent cardiovascular morbidity and type-II diabetes in the mother. BJOG 2010, 117, 274–281. [Google Scholar] [CrossRef]
- James-Todd, T.; Wise, L.; Boggs, D.; Rich-Edwards, J.; Rosenberg, L.; Palmer, J. Preterm birth and subsequent risk of type 2 diabetes in black women. Epidemiology 2014, 25, 805–810. [Google Scholar] [CrossRef] [Green Version]
- Catov, J.M.; Snyder, G.G.; Bullen, B.L.; Barinas-Mitchell, E.J.M.; Holzman, C. Women with Preterm Birth Have Evidence of Subclinical Atherosclerosis a Decade after Delivery. J. Womens Health 2019, 28, 621–627. [Google Scholar] [CrossRef]
- Bonamy, A.K.; Parikh, N.I.; Cnattingius, S.; Ludvigsson, J.F.; Ingelsson, E. Birth characteristics and subsequent risks of maternal cardiovascular disease: Effects of gestational age and fetal growth. Circulation 2011, 124, 2839–2846. [Google Scholar] [CrossRef] [Green Version]
- Ngo, A.D.; Chen, J.S.; Figtree, G.; Morris, J.M.; Roberts, C.L. Preterm birth and future risk of maternal cardiovascular disease—Is the association independent of smoking during pregnancy? BMC Pregnancy Childbirth 2015, 15, 144. [Google Scholar] [CrossRef] [Green Version]
- Nardi, O.; Zureik, M.; Courbon, D.; Ducimetière, P.; Clavel-Chapelon, F. Preterm delivery of a first child and subsequent mothers’ risk of ischaemic heart disease: A nested case-control study. Eur. J. Cardiovasc. Prev. Rehabil. 2006, 13, 281–283. [Google Scholar] [CrossRef]
- Crump, C.; Sundquist, J.; Howell, E.A.; McLaughlin, M.A.; Stroustrup, A.; Sundquist, K. Pre-Term Delivery and Risk of Is-chemic Heart Disease in Women. J. Am. Coll. Cardiol. 2020, 76, 57–67. [Google Scholar] [CrossRef]
- Smith, G.C.; Pell, J.P.; Walsh, D. Pregnancy complications and maternal risk of ischaemic heart disease: A retrospective cohort study of 129,290 births. Lancet 2001, 357, 2002–2006. [Google Scholar] [CrossRef]
- Catov, J.M.; Wu, C.S.; Olsen, J.; Sutton-Tyrrell, K.; Li, J.; Nohr, E.A. Early or recurrent preterm birth and maternal cardio-vascular disease risk. Ann. Epidemiol. 2010, 20, 604–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lykke, J.A.; Langhoff-Roos, J.; Lockwood, C.J.; Triche, E.W.; Paidas, M.J. Mortality of mothers from cardiovascular and non-cardiovascular causes following pregnancy complications in first delivery. Paediatr. Perinat. Epidemiol. 2010, 24, 323–330. [Google Scholar] [CrossRef]
- Rich-Edwards, J.W.; Klungsoyr, K.; Wilcox, A.J.; Skjaerven, R. Duration of pregnancy, even at term, predicts long-term risk of coronary heart disease and stroke mortality in women: A population-based study. Am. J. Obstet. Gynecol. 2015, 213, e1–e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crump, C.; Sundquist, J.; Sundquist, K. Preterm delivery and long term mortality in women: National cohort and co-sibling study. BMJ 2020, 370, m2533. [Google Scholar] [CrossRef] [PubMed]
- Hastie, C.E.; Smith, G.C.; Mackay, D.F.; Pell, J.P. Maternal risk of ischaemic heart disease following elective and spontaneous pre-term delivery: Retrospective cohort study of 750 350 singleton pregnancies. Int. J. Epidemiol. 2011, 40, 914–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.; Gulati, M.; Kwok, C.S.; Wong, C.W.; Narain, A.; O’Brien, S.; Chew-Graham, C.A.; Verma, G.; Kadam, U.T.; Mamas, M.A. Preterm Delivery and Future Risk of Maternal Cardiovascular Disease: A Systematic Review and Meta-Analysis. J. Am. Heart Assoc. 2018, 7, e007809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winger, E.E.; Reed, J.L.; Ji, X. Early first trimester peripheral blood cell microRNA predicts risk of preterm delivery in pregnant women: Proof of concept. PLoS ONE 2017, 12, e0180124. [Google Scholar] [CrossRef] [Green Version]
- Winger, E.E.; Reed, J.L.; Ji, X.; Gomez-Lopez, N.; Pacora, P.; Romero, R. MicroRNAs isolated from peripheral blood in the first trimester predict spontaneous preterm birth. PLoS ONE 2020, 15, e0236805. [Google Scholar] [CrossRef]
- Beta, J.; Akolekar, R.; Ventura, W.; Syngelaki, A.; Nicolaides, K.H. Prediction of spontaneous preterm delivery from maternal factors, obstetric history and placental perfusion and function at 11–13 weeks. Prenat. Diagn. 2011, 31, 75–83. [Google Scholar] [CrossRef]
- O’Gorman, N.; Wright, D.; Poon, L.C.; Rolnik, D.L.; Syngelaki, A.; de Alvarado, M.; Carbone, I.F.; Dutemeyer, V.; Fiolna, M.; Frick, A.; et al. Multicenter screening for pre-eclampsia by maternal factors and biomarkers at 11–13 weeks’ gestation: Comparison with NI-CE guidelines and ACOG recommendations. Ultrasound Obstet. Gynecol. 2017, 49, 756–760. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, N.; Wright, D.; Syngelaki, A.; Akolekar, R.; Wright, A.; Poon, L.C.; Nicolaides, K.H. Competing risks model in screening for preeclampsia by maternal factors and biomarkers at 11–13 weeks gestation. Am. J. Obstet. Gynecol. 2016, 214, e1–e103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Fetal Medicine Foundation. Stratification of Pregnancy Management 11–13 Weeks’ Gestation. Available online: www.courses.fetalmedicine.com/fmf/show/861?locale=en (accessed on 6 January 2022).
- Mazer Zumaeta, A.; Wright, A.; Syngelaki, A.; Maritsa, V.A.; Da Silva, A.B.; Nicolaides, K.H. Screening for pre-eclampsia at 11–13 weeks’ gestation: Use of pregnancy-associated plasma protein-A, placental growth factor or both. Ultrasound Obstet. Gynecol. 2020, 56, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.Y.; Syngelaki, A.; Poon, L.C.; Rolnik, D.L.; O’Gorman, N.; Delgado, J.L.; Akolekar, R.; Konstantinidou, L.; Tsavdari-dou, M.; Galeva, S.; et al. Screening for pre-eclampsia by maternal factors and biomarkers at 11–13 weeks’ gestation. Ultrasound Obstet. Gynecol. 2018, 52, 186–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vora, B.; Wang, A.; Kosti, I.; Huang, H.; Paranjpe, I.; Woodruff, T.J.; MacKenzie, T.; Sirota, M. Meta-Analysis of Maternal and Fetal Transcriptomic Data Elucidates the Role of Adaptive and Innate Immunity in Preterm Birth. Front. Immunol. 2018, 9, 993. [Google Scholar] [CrossRef]
- Hromadnikova, I.; Kotlabova, K.; Krofta, L. Cardiovascular-Disease Associated MicroRNA Dysregulation during the First Trimester of Gestation in Women with Chronic Hypertension and Normotensive Women Subsequently Developing Gestational Hypertension or Preeclampsia with or without Fetal Growth Restriction. Biomedicines 2022, 10, 256. [Google Scholar]
- Hromadnikova, I.; Kotlabova, K.; Krofta, L. First Trimester Screening for Fetal Growth Restriction and Small-for-Gestational-Age Pregnancies without Preeclampsia using Cardiovascular-Disease Associated MicroRNA Biomarkers. Biomedicines 2022, 10, 718. [Google Scholar] [CrossRef]
- Guinn, D.A.; Goldenberg, R.L.; Hauth, J.C.; Andrews, W.W.; Thom, E.; Romero, R. Risk factors for the development of pre-term premature rupture of the membranes after arrest of preterm labor. Am. J. Obstet. Gynecol. 1995, 173, 1310–1315. [Google Scholar] [CrossRef]
- Challis, J.R.; Lockwood, C.J.; Myatt, L.; Norman, J.E.; Strauss, J.F., 3rd; Petraglia, F. Inflammation and pregnancy. Reprod. Sci. 2009, 16, 206–215. [Google Scholar] [CrossRef]
- Keelan, J.A. Pharmacological inhibition of inflammatory pathways for the prevention of preterm birth. J. Reprod. Immunol. 2011, 88, 176–184. [Google Scholar] [CrossRef]
- Ekwo, E.E.; Gosselink, C.A.; Woolson, R.; Moawad, A. Risks for premature rupture of amniotic membranes. Int. J. Epidemiol. 1993, 22, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Hadley, C.B.; Main, D.M.; Gabbe, S.G. Risk factors for preterm premature rupture of the fetal membranes. Am. J. Perinatol. 1990, 7, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Naeye, R.L.; Peters, E.C. Causes and consequences of premature rupture of fetal membranes. Lancet 1980, 1, 192–197. [Google Scholar] [CrossRef]
- Menon, R.; Boldogh, I.; Hawkins, H.K.; Woodson, M.; Polettini, J.; Syed, T.A.; Fortunato, S.J.; Saade, G.R.; Papaconstantinou, J.; Taylor, R.N. Histological evidence of oxidative stress and premature senescence in preterm premature rupture of the human fetal membranes recapitulated in vitro. Am. J. Pathol. 2014, 184, 1740–1751. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Polettini, J.; Syed, T.A.; Saade, G.R.; Boldogh, I. Expression of 8-oxoguanine glycosylase in human fetal membranes. Am. J. Reprod. Immunol 2014, 72, 75–84. [Google Scholar] [CrossRef]
- Menon, R.; Yu, J.; Basanta-Henry, P.; Brou, L.; Berga, S.L.; Fortunato, S.J.; Taylor, R.N. Short fetal leukocyte telomere length and preterm prelabor rupture of the membranes. PLoS ONE 2013, 7, e31136. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Sun, J.; Groome, L.J.; Wang, Y. Differential miRNA expression profiles between the first and third trimester human placentas. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E836–E843. [Google Scholar] [CrossRef]
- Addo, K.A.; Palakodety, N.; Hartwell, H.J.; Tingare, A.; Fry, R.C. Placental microRNAs: Responders to environmental chemicals and mediators of pathophysiology of the human placenta. Toxicol. Rep. 2020, 7, 1046–1056. [Google Scholar] [CrossRef]
- Committee on Obstetric Practice; Society for Maternal-Fetal Medicine. ACOG Committee Opinion No. 743. Low-Dose Aspirin Use during Pregnancy. Obstet. Gynecol. 2018, 132, e44–e52. [Google Scholar]
- National Institute for Health and Care Excellence. Hypertension in Pregnancy: Diagnosis and Management. Available online: www.nice.org.uk/guidance/ng133 (accessed on 6 January 2022).
- Hromadnikova, I.; Kotlabova, K.; Krofta, L. Association Analysis in Young and Middle-Aged Mothers-Relation between Expression of Cardiovascular Disease Associated MicroRNAs and Abnormal Clinical Findings. J. Pers. Med. 2021, 11, 39. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hromadnikova, I.; Kotlabova, K.; Hympanova, L.; Krofta, L. Gestational hypertension, preeclampsia and intrauterine growth restriction induce dysregulation of cardiovascular and cerebrovascular disease associated microRNAs in maternal whole peripheral blood. Thromb. Res. 2016, 137, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Hromadnikova, I.; Kotlabova, K.; Hympanova, L.; Krofta, L. Cardiovascular and Cerebrovascular Disease Associated microRNAs Are Dysregulated in Placental Tissues Affected with Gestational Hypertension, Preeclampsia and Intrauterine Growth Restriction. PLoS ONE 2015, 10, e0138383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dweep, H.; Sticht, C.; Pandey, P.; Gretz, N. miRWalk—Database: Prediction of possible miRNA binding sites by “walking” the genes of three genomes. J. Biomed. Inform. 2011, 44, 839–847. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term Normal Pregnancies (n = 80) | PTB (n = 41) | PPROM (n = 65) | p-Value 1 | p-Value 2 | |
|---|---|---|---|---|---|
| Maternal Characteristics | |||||
| Maternal age (years) | 32 (25–42) | 33 (21–43) | 32 (25–42) | 0.735 | 1.0 |
| Advanced maternal age (≥35 years old) | 20 (25%) | 15 (36.58%) | 20 (30.77%) | 0.183 | 0.439 |
| Caucasian ethnic group | 80 (100%) | 41 (100%) | 65 (100%) | 1.0 | 1.0 |
| Prepregnancy BMI (kg/m2) | 21.28 (17.16–29.76) | 22.04 (17.96–31.83) | 22.14 (16.51–33.5) | 1.0 | 1.0 |
| Diabetes mellitus (T1DM, T2DM) | 0 (0%) | 3 (7.32%) | 2 (3.08%) | - | - |
| Autoimmune diseases (SLE/APS/RA) | 0 (0%) | 0 (0%) | 2 (3.08%) | - | - |
| Parity | |||||
| Nulliparous—no previous pregnancy | 32 (40.0%) | 11 (26.83%) | 30 (46.15%) | 0.152 | 0.456 |
| Nulliparous—previous abortion(s) (including miscarriages) | 9 (11.25%) | 3 (7.32%) | 12 (18.46%) | 0.493 | 0.220 |
| Parous—previous abortion(s) (including miscarriages) | 12 (15.0%) | 12 (29.27%) | 11 (16.92%) | 0.062 | 0.752 |
| Parous—previous preterm delivery(ies) | 0 (0%) | 11 (26.83%) | 6 (9.23%) | - | - |
| Parous—previous term delivery(ies) | 39 (48.75%) | 16 (39.02%) | 17 (26.15%) | 0.309 | 0.005 |
| ART (IVF/ICSI/other) | 2 (2.5%) | 1 (2.44%) | 7 (10.77%) | 0.984 | 0.040 |
| Smoking during pregnancy | 2 (2.5%) | 3 (7.32%) | 2 (3.08%) | 0.208 | 0.833 |
| Pregnancy Details (First Trimester of Gestation) | |||||
| Gestational age at sampling (weeks) | 10.29 (9.57–13.71) | 10.14 (9.43–12.86) | 10.14 (9.86–14.57) | 0.477 | 0.291 |
| MAP (mmHg) | 88.75 (67.67–103.83) | 88.58 (75.0–108.0) | 89.58 (69.75–106.75) | 1.0 | 1.0 |
| MAP (MoM) | 1.05 (0.84–1.25) | 1.04 (0.89–1.26) | 1.07 (0.83–1.26) | 1.0 | 1.0 |
| Mean UtA-PI | 1.39 (0.56–2.43) | 1.40 (0.66–2.42) | 1.46 (0.69–2.95) | 1.0 | 0.377 |
| Mean UtA-PI (MoM) | 0.90 (0.37–1.55) | 0.89 (0.44–1.64) | 0.91 (0.43–1.88) | 1.0 | 0.665 |
| PIGF serum levels (pg/mL) | 27.1 (8.1–137.0) | 23.65 (5.6–67.0) | 24.45 (10.2–65.7) | 0.327 | 0.245 |
| PIGF serum levels (MoM) | 1.04 (0.38–2.61) | 0.96 (0.26–1.68) | 0.97 (0.44–1.80) | 0.347 | 0.468 |
| PAPP-A serum levels (IU/L) | 1.49 (0.48–15.69) | 1.34 (0.34–15.28) | 1.10 (0.31–8.36) | 1.0 | 0.009 |
| PAPP-A serum levels (MoM) | 1.17 (0.37–3.18) | 0.96 (0.33–3.34) | 0.85 (0.21–3.24) | 1.0 | 0.038 |
| Free β-hCG serum levels (μg/L) | 60.21 (9.9–200.6) | 71.27 (17.1–234.4) | 49.89 (0.96–179.78) | 0.515 | 0.672 |
| Free β-hCG serum levels (MoM) | 1.02 (0.31–3.57) | 1.19 (0.40–3.01) | 0.89 (0.20–2.50) | 0.850 | 0.248 |
| Screen-positive for preterm delivery (<34 weeks) by FMF algorithm | 5 (6.25%) | 12 (29.27%) | 13 (20.0%) | <0.001 | 0.039 |
| Screen-positive for PE and/or FGR by FMF algorithm | 0 (0%) | 2 (4.88%) | 8 (12.31%) | - | - |
| Aspirin intake during pregnancy | 0 (0%) | 1 (2.44%) | 6 (10.77%) | - | - |
| Pregnancy Details (At Delivery) | |||||
| Gestational age at delivery (weeks) | 40.07 (37.57–42.0) | 34.57 (24.0–36.86) | 35.29 (26.43–36.86) | <0.001 | <0.001 |
| Delivery at gestational age <34 weeks | 0 (0%) | 16 (39.02%) | 13 (20.0%) | - | - |
| BMI (kg/m2) | 26.66 (21.71–34.82) | 25.95 (18.14–34.08) | 25.86 (20.08–37.18) | 0.259 | 0.338 |
| Weight gain during pregnancy (kg) | 14 (3–25) | 10 (3–20) | 11 (3–20) | <0.001 | <0.001 |
| Administration of corticosteroids | 0 (0%) | 23 (56.10%) | 26 (40.0%) | - | - |
| Administration of antibiotics | 0 (0%) | 19 (46.34%) | 52 (80.0%) | - | - |
| Tocolytic therapy | 0 (0%) | 27 (65.85%) | 20 (30.77%) | - | - |
| Fetal birth weight (grams) | 3470 (2920–4240) | 2290 (680–3800) | 2530 (770–3370) | < 0.001 | < 0.001 |
| Fetal sex | |||||
| Boy | 40 (50.0%) | 21 (51.22%) | 33 (50.77%) | 0.899 | 0.926 |
| Girl | 40 (50.0%) | 20 (48.78%) | 32 (49.23%) | ||
| Mode of delivery | |||||
| Vaginal | 69 (86.25%) | 20 (48.78%) | 46 (70.77%) | <0.001 | 0.022 |
| CS | 11 (13.75%) | 21 (51.22%) | 19 (29.23%) | ||
| Apgar score <7, 5 min | 0 (0%) | 2 (4.88%) | 0 (0%) | - | - |
| Apgar score <7, 10 min | 0 (0%) | 0 (0%) | 0 (0%) | - | - |
| Umbilical blood pH | 7.3 (7.29–7.38) | 7.3 (6.9–7.3) | 7.3 (6.9–7.31) | 0.701 | 1.0 |
| Predicted Targets | |||
|---|---|---|---|
| MicroRNA | KEGG Pathways | Wiki Pathways | Panther Pathways |
| miR-16-5p | BCL2, IKBKB, IRAK2, PRKAR1A, PRKAR2A | BCL2, IKBKB | BCL2, CRADD, IKBKB |
| miR-20b-5p | ATM, CASP6, CASP7, CASP8, CASP10, CFLAR, CYCS, DFFA, EXOG, FASLG, IL1 R1, IRAK1, IRAK4, MAP3K14, PIK3R2, PPP3CA, PRKAR2A, PRKX, TNFRSF10A, TNFRSF10D, XIAP | BNIP3L, CASP6, CASP7, CASP8, CASP10, CFLAR, CYCS, DFFA, FASLG, IRF1, TNFRSF1B, TNFRSF21 | ATF6, BAG1, CASP7, CASP8, CASP10, CFLAR, CREB1, CREM, CYCS, EIF2S1, FASLG, HSPA5, MAP3K14, MAPK9, PRKCQ, REL, TNFRSF10D, TNFRSF1B, XIAP |
| miR-21-5p | APAF1, CFLAR, FASLG | APAF1, CFLAR, FASLG, MAP3K1 | APAF1, CFLAR, DAXX, EIF2S1, FASLG, MAP2K3 |
| miR-24-3p | BCL2L1, EXOG, FASLG, IKBKB, IL1B, IRAK4, MYD88, PIK3CB, RIPK1 | BBC3, BCL2L2, BCL2L11, BNIP3L, FASLG, IKBKB, MYC, NFKBIE, RIPK1, TRAF1, TRAF3 | BCL2L1, BCL2L2, BCL2L11, EIF2AK2, FASLG, FOS, IKBKB, PIK3CB, PRKCA, PRKCH, RIPK1 |
| miR-26a-5p | APAF1, BID, BIRC2, CASP6, DFFB, PPP3CB, PPP3CC | APAF1, BAK1, BID, BIRC2, CASP6, CRADD, DFFB, MDM2, PMAIP1 | APAF1, ATF2, BAG4, BAK1, BID, BIRC2, CRADD, CREB1, EIF2AK2, FOS, HSPA8, PRKCD, PRKCQ, RELB |
| miR-92a-3p | APAF1, ATM, BCL2L1, BCL2L11, BIRC3, CASP10, CASP8, CASP9, CFLAR, CSF2RB, CTSB, CTSS, CTSV, DAB2IP, DFFA, DFFB, FOS, GZMB, HRK, KRAS, MAP3K14, MAPK10, NTRK1, RIPK1, TNFRSF10B, TP53, XIAP | AKT1, APAF1, BCL2L1, BCL2L11, BCL2L2, BIRC3, CASP10, CASP8, CASP9, CFLAR, DFFA, DFFB, GZMB, IGF1, IGF2, IRF1, IRF4, MAPK10, MDM2, RIPK1, TNFRSF10B, TNFRSF25, TP53, TP63, TP73, TRAF3 | APAF1, APPL1, BCL2L1, BCL2L11, BMF, CARD8, CASP8, CASP9, CDH1, CFLAR, DAPK2, DAPK3, DFFA, DFFB, GZMB, KPNB1, MAPT, NMT1, PAK2, PSMB2, PSMD5, PSMD8, PSMD9, PSMF1, PTK2, RIPK1, STAT3, STK26, TFDP2, TNFRSF10B, TP53, TP63, TP73, UNC5B, XIAP, YWHAZ |
| miR-126-3p | TNFRSF10B | TNFRSF10B | - |
| miR-133a-3p | ENDOD1, IRAK3, MAP3K14, TNFRSF10B | BCL2L2, BNIP3L, TNFRSF10B | BCL2L2, MAP3K14, TNFRSF10B |
| miR-145-5p | AIFM1, PIK3R5, TNFRSF10B | TNFRSF10B, TNFRSF25 | AIFM1, MAP4K2, TMBIM6, TNFRSF10B |
| miR-146a-5p | CASP7, CASP9, DFFA, IL3, IRAK1, IRAK4, PPP3R2, PRKACA | CASP2, CASP7, CASP9, DFFA, PMAIP1, PRF1 | BAG1, CASP7, CASP9, HSPA1A, JDP2, PRKCE |
| miR-155-5p | PIK3R1, LMNB2 | AKT1, IGF1, PIK3R1 | PSMD8, TJP1 |
| miR-210-3p | AKT1, ATM, BAK1, BBC3, BCL2L11, BIRC3, CASP10, CSF2RB, CTSB, EIF2S1, ERN1, MAPK10, MCL1, PDPK1, RAF1, RIPK1, TP53AIP1 | AKT1, BAK1, BBC3, BCL2L11, BIRC3, CASP10, IGF1, IGF1R, IRF2, MAPK10, MCL1, RIPK1, TP63, TP73 | AKT1, APPL1, BAK1, BBC3, BCAP31, BCL2L11, CLSPN, DAPK1, GSDME, HMGB1, MAPT, PAK2, PSMA5, PSMB2, PSMB5, PSMC1, PSMD9, PSME3, PTK2, RIPK1, ROCK1, STAT3, TFDP2, TP63, UBA52 |
| miR-221-3p | AKT3, APAF1, CASP10, IKBKG, IL1RAP, PIK3CD, PPP3R1, TNFSF10 | APAF1, BNIP3L, CASP10, IKBKG, IRF4, MAPK10, MDM2, TNFSF10 | AKT3, APAF1, ATF2, ATF4, CASP10, CREB1, MAPK10, PIK3CD, PRKCB, TNFSF10 |
| miR-342-3p | ACTG1, CFLAR, EIF2S1, MAPK1, MAPK9, PMAIP1 | BCL2L2, CFLAR, PMAIP1 | CFLAR, DAPK2, DYNLL2, E2F1, MAPK1, OCLN, PMAIP1, PSME3, ROCK1, SFN, TFDP2, TICAM2, UNC5B |
| Predicted Targets (Wiki Pathways) | ||
|---|---|---|
| MicroRNA | Senescence and Autophagy Pathways | Inflammatory Response Pathway |
| miR-16-5p | BCL2, CREG1, HMGA1, LAMP2, MAP2K1, RAF1, SMAD4 | IL2RA |
| miR-20b-5p | ATG10, ATG12, CD44, CDKN1A, E2F1, IL6R, IRF1, LAMP2, RNASEL, RSL1D1, SERPINE1, SH3GLB1 | CD28, IL5, LAMC1, LAMC2, TNFRSF1B |
| miR-21-5p | MAP2K3 | THBS3 |
| miR-24-3p | ATG13, CDKN1B, FN1, IFNG, IGFBP5, IL1B, IL6R, MAP1LC3A, MAP1LC3C, MMP14 | CD28, CD86, FN1, IFNG, IL2RB, LAMC1 |
| miR-26a-5p | ATG13, COL10A1, HMGA1, IFNG, IL6, MDM2, PCNA, PTEN, RB1, ULK1 | COL1A2, IFNG |
| miR-92a-3p | CXCR2, HMGA1, IGF1, IL24, IL6R, IL6ST, ING1, IRF1, LAMP2, MAP2K6, MDM2, MMP14,PIK3C3, PLAT, SLC39A1, TP53 | CD28, CD40, IL2RA, IL5RA |
| miR-126-3p | - | - |
| miR-133a-3p | ATG14, FN1, GABARAPL1, MAPK14, MMP14, RB1CC1, SLC39A1 | CD28, FN1 |
| miR-145-5p | AMBRA1, CD44, HMGA1, IFNB1, MAP1LC3B, SLC39A2 | IL2RA |
| miR-146a-5p | ATG12, IL3, KMT2A, RNASEL, SERPINB2, TNFSF15 | CD80, CD86 |
| miR-155-5p | ATG7, BRAF, CDK6, IGF1 | CD28 |
| miR-210-3p | CDC25B, IGF1R, INHBA, MAPK14, PTEN, RAF1, SMAD3, SMAD4, IGF1, MAP2K6, PIK3C3 | CD28, IL5RA |
| miR-221-3p | - | THBS1, VTN |
| miR-342-3p | ATG14, ATG16L1, E2F1, GSK3B, MAPK1, ULK1, ING1 | - |
| K | i | Alpha = 0.05 | Alpha = 0.01 | Alpha = 0.001 |
|---|---|---|---|---|
| 2 | 0.05 | 0.01 | 0.001 | |
| 1 | 0.025 | 0.005 | 0.001 |
| K | i | Alpha = 0.05 | Alpha = 0.01 | Alpha = 0.001 |
|---|---|---|---|---|
| 3 | 0.05 | 0.01 | 0.001 | |
| 1 | 0.017 | 0.003 | 0.000 | |
| 2 | 0.033 | 0.007 | 0.001 | |
| 3 | 0.050 | 0.010 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hromadnikova, I.; Kotlabova, K.; Krofta, L. First Trimester Prediction of Preterm Delivery in the Absence of Other Pregnancy-Related Complications Using Cardiovascular-Disease Associated MicroRNA Biomarkers. Int. J. Mol. Sci. 2022, 23, 3951. https://doi.org/10.3390/ijms23073951
Hromadnikova I, Kotlabova K, Krofta L. First Trimester Prediction of Preterm Delivery in the Absence of Other Pregnancy-Related Complications Using Cardiovascular-Disease Associated MicroRNA Biomarkers. International Journal of Molecular Sciences. 2022; 23(7):3951. https://doi.org/10.3390/ijms23073951
Chicago/Turabian StyleHromadnikova, Ilona, Katerina Kotlabova, and Ladislav Krofta. 2022. "First Trimester Prediction of Preterm Delivery in the Absence of Other Pregnancy-Related Complications Using Cardiovascular-Disease Associated MicroRNA Biomarkers" International Journal of Molecular Sciences 23, no. 7: 3951. https://doi.org/10.3390/ijms23073951
APA StyleHromadnikova, I., Kotlabova, K., & Krofta, L. (2022). First Trimester Prediction of Preterm Delivery in the Absence of Other Pregnancy-Related Complications Using Cardiovascular-Disease Associated MicroRNA Biomarkers. International Journal of Molecular Sciences, 23(7), 3951. https://doi.org/10.3390/ijms23073951