
Hidden Relationships between N-Glycosylation and Disulfide Bonds in Individual Proteins


Abstract
1. Introduction
2. Molecular Foundations
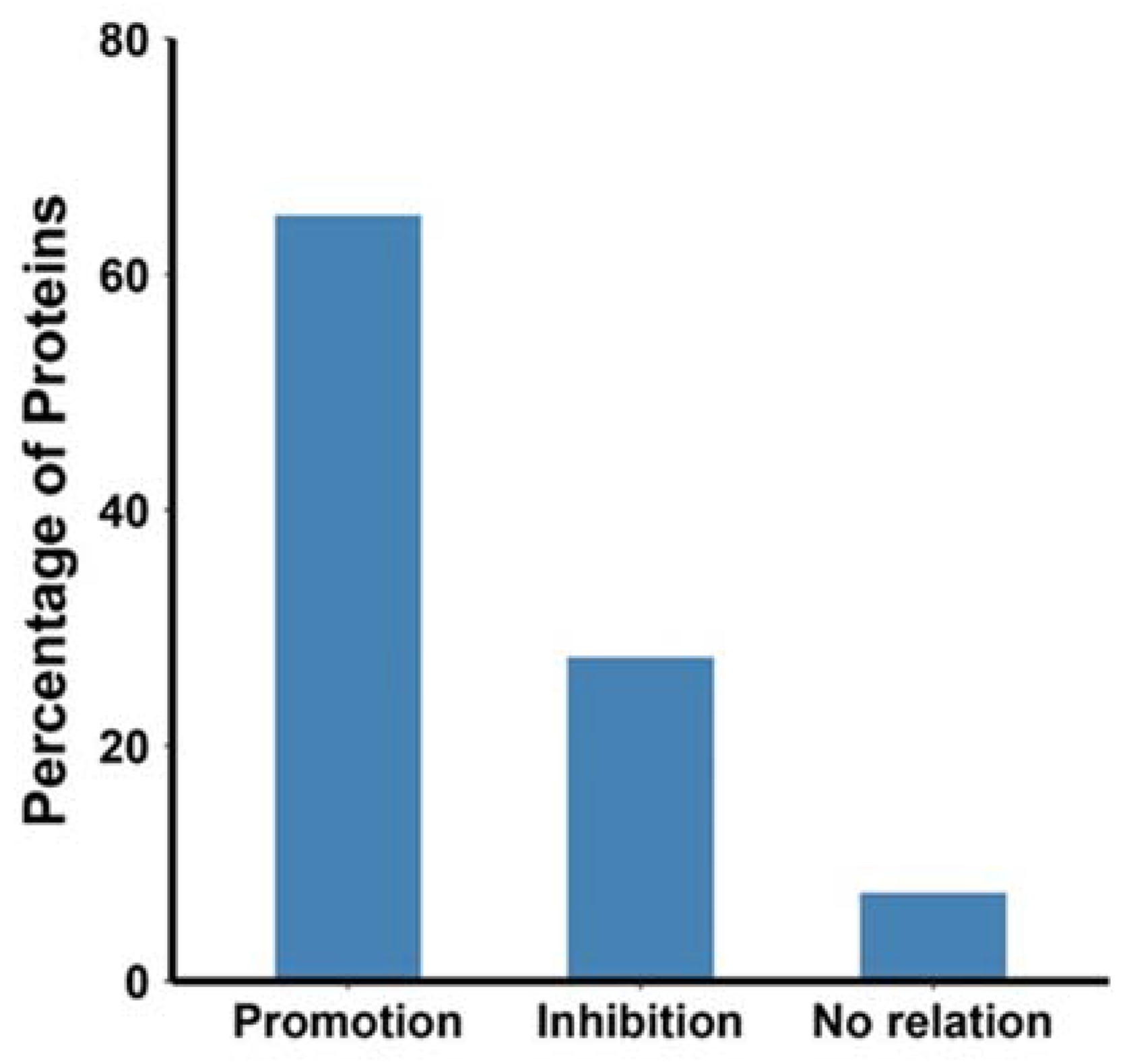
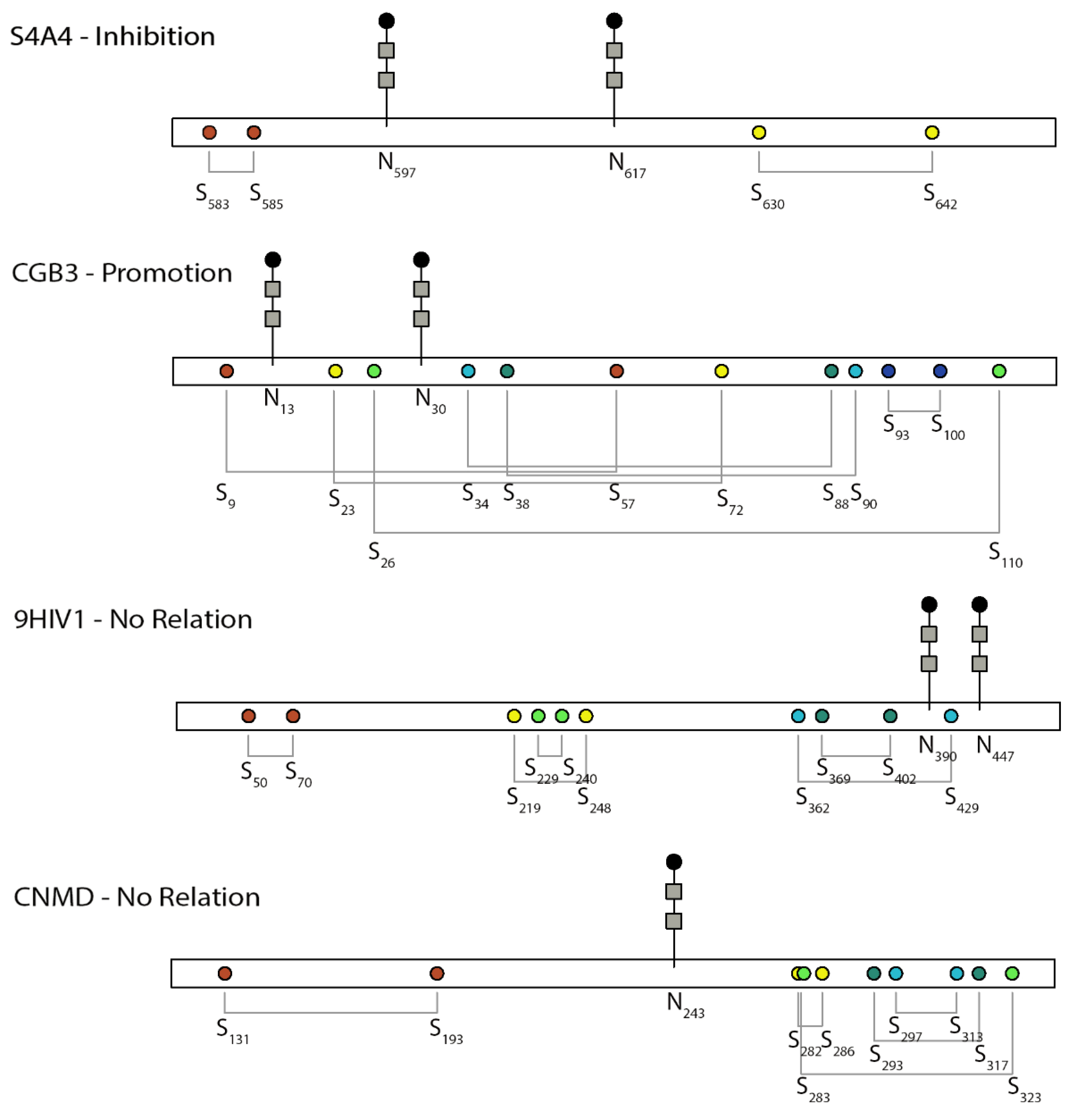
3. Overview of the Studies

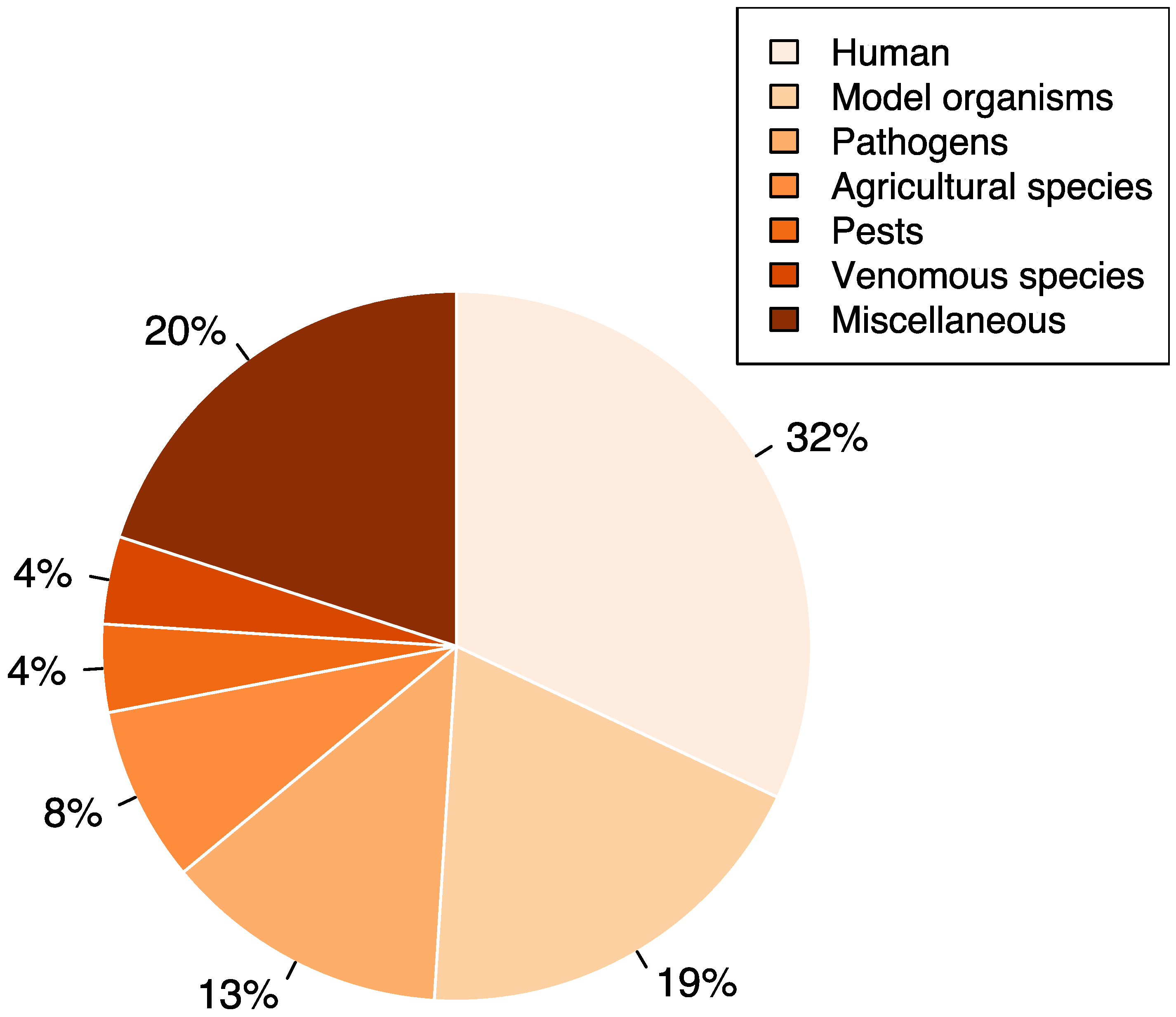{kind=link}
{kind=link}
{kind=link}
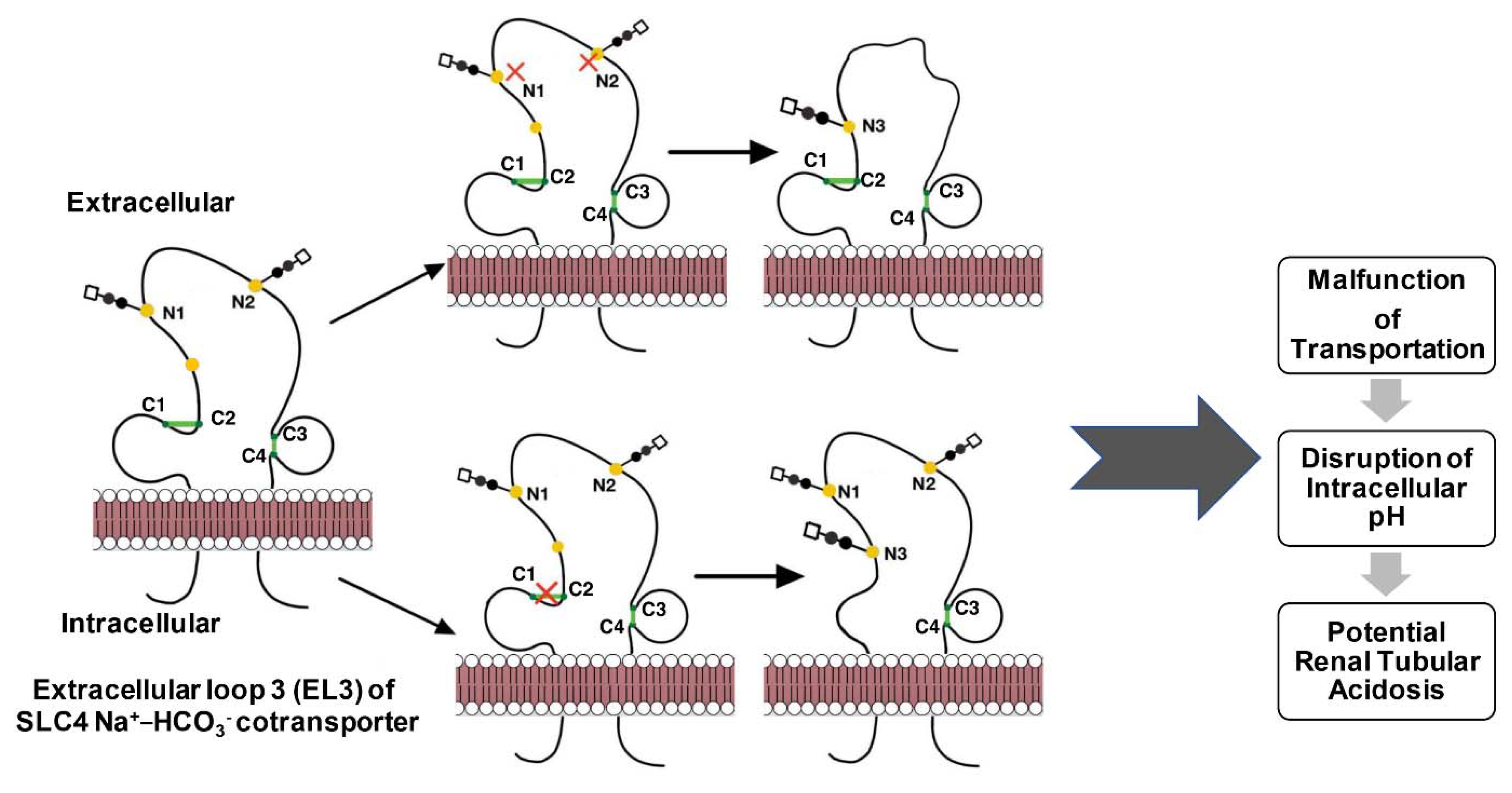{kind=link}
| UniProt Accession | Protein Name | Position of DBs | Position of N-Glycans | Class | Reference |
|---|---|---|---|---|---|
| A0A346IHA8 | CD4-binding region of the envelope glycoproteins | 50–70 a, 219–248 a, 229–240 a, 369–402 a, 362–429, 597–603 a | 390, 447 | No relation | [27] |
| Q9Y6R1 | SLC4 Na+-coupled transporter (NBCe1-A) | 583–585, 630–642 c | 597, 617 | Inhibition | [28] |
| Q99062 | Granulocyte colony-stimulating factor receptor | 107–118, 153–162, 224–271, 364–371 | 27, 69, 104, 110, 365, 450, 547, 555, 586 | Promotion | [29] |
| Q62635 | Gastric mucin (rat) | 160 a, 420 a, 667 a, 767 a, 837 a, 892 a, 1136 a, 1151 a, 1212 a, 1227 a, 1243 a, 1350 a, | Promotion | [30] | |
| P49018 | GPI-anchor transamidase (GPI8) yeast | 85 (interchain with C-194 in GPI16) b | 256 a, 346 a | Promotion | [31] |
| P00750 | Tissue-type plasminogen activator (t-PA) | 41–71, 69–78, 86–97, 91–108, 110–119, 127–208 b, 148–190 b, 179–203 b, 215–296, 236–278, 267–291, 299–430, 342–358 b, 350–419 b, 444–519 b, 476–492 b, 509–537 b | 117, 184, 448 | Inhibition | [32,33] |
| P04275 | Von Willebrand factor (VWF) | 35–162 d, 57–200 d, 388–524 d, 410–559 d, 432–440 d, 509–695, 767–808, 776–804, 810–821, 867–996 d, 889–1031 d, 898–993 d, 914–921 d, 1060–1084, 1071–1111, 1089–1091, 1126–1130, 1149–1169, 1153–1165, 1196–1199, 1234–1237, 1272–1458, 1669–1670, 1686–1872, 1879–1904, 1899–1940 d, 1927–2088, 1950–2085 d, 1972–2123 d, 1993–2001 d, 2724–2774 b, 2739–2788 b, 2750–2804 b, 2754–2806 b | 99 a, 156 a, 211 a, 666 a, 857 e, 1147 (atypical) e, 1231 e, 1515, 1574 e, 2223 e, 2290 e, 2357 e, 2400 e, 2546, 2585 e, 2790 e | Promotion | [34,35,36] |
| P0DN86 | Human chorionic gonadotropin beta-subunit | 9–57, 23–72, 26–110, 34–88, 38–90, 93–100 | 13, 30 | Promotion | [37,38,39] |
| Engineered heterodimeric knob-into-hole Fc fragments | 349–354 (interchain) | 297 | Promotion | [40] | |
| P32906 | Yeast-alpha1,2 mannosidase | 340–385, 468–471 | 96, 155, 224 | Promotion | [41] |
| O60896 | Receptor activity-modifying protein 3 (RAMP3) | 40–72 b, 57–104 b | 28, 57, 70, 102 | Inhibition | [42] |
| Q13087 | PDIA2 | 18 (interchain) | 127, 284, 516 | Inhibition | [43] |
| P04853 | Hemagglutinin-neuraminidase (Sendai virus) | 129 (interchain) a | 77, 499, 511 | Promotion | [21,44] |
| Q91UL0 | Hemagglutinin-neuraminidase (NDV) | 123 (interchain), 172–196, 186–247, 238–251, 344–461, 455–465, 531–542 | 119, 341, 433, 481 | Inhibition | [45,46] |
| Hemagglutinin (Influenza A virus) | 14–466, 52–277, 64–76, 97–139, 281–305, 473–477 | 8, 22, 38, 81, 165, 285, 483 | Promotion | [47,48] | |
| P01229 | Human lutropin subunit beta | 29–77 b, 43–92 b, 46–130 b, 54–108 b, 58–110 b, 113–120 b | 30 | Promotion | [49] |
| P35625 | Metalloproteinase inhibitor 3 (TIMP3) | 24–91, 26–118, 36–143, 145–192 d, 150–155 d, 163–184 d | Inhibition | [50] | |
| Q16820 | Meprin A | 103–255, 124–144, 265–427, 273 a (interchain), 305 (interchain), 492 (interchain) d, 608–619 d, 613–628 d, 630–643 d | 41, 152, 234, 270, 330, 426, 452, 546, 553 | Promotion | [51,52] |
| P01130 | Low-density lipoprotein (LDL) receptor | 27–39 e, 34–52 e, 46–63 e, 68–82 e, 75–95 e, 89–104 e, 109–121 b, 116–134 e, 128–143 e, 148–160 e, 155–173 e, 167–184 e, 197–209 e, 204–222 e, 216–231 e, 236–248 e, 243–261 e, 255–270 e, 276–289 e, 284–302 e, 296–313 e, 318–329 e, 325–338 e, 340–352 e, 358–368 e, 364–377 e, 379–392 e, 667–681 e, 677–696 e, 698–711 e | 97 a, 156, 272, 515 a, 657 | Inhibition | [53,54,55] |
| Q02817 | MUC2 mucin | 59–67 b, 37–169 d, 59–206 d, 391–528 d, 413–563 d, 435–443 d, 860–992 d, 882–1027 d, 891–989 d, 909–916 d, 4481–4622 d, 4503–4661 d, 4527–4535 d, 5075–5122 b, 5089–5136 b, 5098–5152 b, 5102–5154 b | 163 a, 423 a, 670 a, 770 a, 894 a, 1139 a, 1154 a, 1215 a, 1230 a, 1246 a, 1787 a, 1820 a, 4339 a, 4351 a, 4362 a, 4373 a, 4422 a, 4438 a, 4502 a, 4616 a, 4627 a, 4752 a, 4787 a, 4881 a, 4888 a, 4955 a, 4970 a, 5019 a, 5038 a, 5069 a | Promotion | [56,57] |
| P12476 | VP7 | 82–135, 165–249, 191–244, 196–207 | 69 | Promotion | [22,54] |
| P04156 | Major prion protein | 179–214 | 181, 197 | Promotion | [58] |
| P05026 | Sodium/potassium-transporting ATPase subunit beta-1 | 126–149, 159–175, 213–276 | 158, 193, 265 | Promotion | [59,60] |
| P05231 | Interleukin-6 (IL6) | 45–51, 74–84 | 46 | Inhibition | [61,62,63] |
| H2AM12 | Glycoprotein Gc | 523–550, 580–589, 591–598, 471–487 | 493, 686 | Promotion | [64] |
| P22146 | 1,3-beta-Glucanosyltransferase | 74–103, 216–348, 234–265, 370–421, 372–462 b, 379–445 b, 398–403 b | 40 a, 57, 95 a, 149 a, 165 a, 253, 283 a, 321 a, 409 a, 495 a | Promotion | [65,66] |
| P32623 | Probable glycosidase CRH2 | 28, 96 a, 190 a, 196 a, 233 a, 237 a, 261 a, 297 a, 310 a | Promotion | [66] | |
| Q9UMF0 | Intercellular adhesion molecule-5 (ICAM5) | 55–99, 59–103, 142–198, 249–302 d, 344–383 d, 415–470 d, 498–552 d, 580–645 d, 673–725 d, 769–814 d | 54,74,137,195,214,274,316,371,397,582,636,645,762,793,794 | Promotion | [67,68] |
| O75829 | Chondromodulin-I | 131–193 b, 282–286 b, 283–323 b, 293–317 b, 297–313 b | 243 a | No relation | [69] |
| P40225 | Thrombopoietin | 7–151, 29–85 | 197, 206, 234, 255, 340 a, 348 a | No relation | [70,71] |
| Q9UNQ0 | ABCG2 protein | 592–608, 603 (interchain) | 596 | Promotion | [8,72,73] |
| P56817 | beta-Site APP-cleaving enzyme (BACE) | 216–420, 278–443, 330–380 | 153 a, 172 a, 223 a, 354 a | Promotion | [74] |
| Q9H1U4 | Multiple epidermal growth factor-like domains protein 9 (MEGF9) | 204–217 a, 206–224 a, 226–235 a, 238–251 a, 254–266 a, 256–272 a, 274–283 a, 286–298 a, 301–310 a, 303–317 a, 320–329 a, 332–346 a, 349–360 a, 351–371 a, 374–383 a, 386–397 a, 400–415 a, 402–422 a, 425–434 a, 437–449 a | 40 a, 182 a, 205 a, 218 a, 245 a, 267 a, 305 a, 428 a, 468 a, 481 a, 500 a | Inhibition | [75] |
| P53634 | Cathepsin | 30–118, 54–136 e, 255–298 e, 291–331 e, 321–337 e | 29 | Inhibition | [76,77] |
| P08709 | Coagulation factor VII | 348–367 | 360 | Inhibition | [76] |
| Q07837 | rBAT | 242–273, 571–666, 673–685 | 214, 261, 332, 495, 513, 575 | Promotion | [78] |
| O75355 | Nucleoside triphosphate diphosphohydrolase 3 (NTPDase3) | 92–116, 261–308, 289–334, 347–353, 399–422 | 81 a, 149 a, 238 a, 381 a, 392 a, 402 a, 454 a | Inhibition | [79] |
| P08563 | E2 glycoprotein (rubella virus) | 53, 71, 115 | Promotion | [80] | |
| Q9H9S5 | Fukutin-related protein (FKRP) | 6 (interchain) | 172, 209 | No relation | [81] |
| O14773 | Tripeptidyl-peptidase I | 111–122, 365–526, 522–537 | 210, 222, 286, 313, 443 | Promotion | [82,83,84,85] |
| P15813 | CD1d | 120–184, 224–279 | 20, 42, 108, 163 | Promotion | [86,87,88,89] |
| O14656 | TorsinA | 44–162 a, 280–319 | 143, 158 | Promotion | [90,91] |
| P21825 | Translocation protein SEC62 | 153, 62 e | Promotion | [92] | |
| P04070 | Protein C | 331–345 | 97, 248, 313, 329 (atypical) | Promotion | [93] |
| K7WJ21 | PtrMAN6 | 448, 452, 456 | 23, 194, 227, 375, 392 | No relation | [94] |
| P01848/P01850 | TCR alpha and beta | 22–72, 94 (with C-130 in TRBC1 or TRBC2) and 30–95, 130 (with C-94 in TRAC) | 32, 66, 77 d, 113 d and 69 d | Inhibition | [95] |
| P01857 | Immunoglobulin G1 Fc | 27–83 d, 103 e, 109 e, 112 e, 144–204 d, 250–308 d | 180, 297 | Promotion | [84,96,97,98] |
| Nicotinic acetylcholine receptor fragment | 128–142 | 141 | Promotion | [12] | |
| Q07108 | CD69 | 68 (interchain) e, 85–96 e, 113–194 e, 173–186 e | 111 (atypical), 166 | Inhibition | [99,100] |
| P15813 | Antigen-presenting glycoprotein CD1d | 120–184 e, 224–279 e | 38 e, 60 e, 126 e, 181 e | Promotion | [101,102] |
| P04439 | MHC class I heavy chain | 125–188 e, 227–283 e | 110 e | Promotion | [103,104] |
4. Promoting Relationship
5. Inhibitory Relationship
6. Independent Relationship
7. Unknown Relation
8. Common Methods
| NG detection | Staining procedures [130] | |
| ||
| Affinity-based methods [130] | ||
| ||
| NMR spectroscopy [131,132] | ||
| X-ray crystallography [133] | ||
| Circular dichroism (CD) spectroscopy [64] | ||
| NG structural analysis | Requires N-glycan removal prior to further analysis which can be achieved by | Chromatography |
| ||
| NG functional analysis | Chemical tools: inhibitors of glycosyltransferases and glycosidases (in vivo) [150] | |
| ||
| Physical tools: adjust temperature, ATP, pH, etc. | ||
| Site-directed mutagenesis | ||
| Genetic glyco-engineering [152] | ||
| ||
| Quantify the amount of unbound cysteine residues | Chemical labeling and spectroscopic detection [153] | |
| ||
| Reduction methods | Reducing enzymes [153] | |
| ||
| Detection/Analysis of DB | Edman degradation sequencing [154] | |
| NMR spectroscopy [155,156] | ||
| X-ray crystallography [157] | ||
| 2-Nitro-5-thiosulfobenzoate (NTSB) assay [158,159] | ||
| ||
| Mass spectrometry approaches: Front-end separation [153,163,164,165,166,167,168,169] | Mass spectrometry approaches: Fragmentation types | |
| ||
| Detect structural changes | Partial proteolysis by enzymes such as trypsin and pepsin | |
| Functional assays | ||
| ||
| Site-directed mutagenesis | ||
9. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, B.M.; Oster, M.E.; Hebert, D.N. Protein Quality Control in the Endoplasmic Reticulum. Protein J. 2019, 38, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Grek, C.; Townsend, D.M. Protein Disulfide Isomerase Superfamily in Disease and the Regulation of Apoptosis. Endoplasmic Reticulum Stress Dis. 2014, 1, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef] [PubMed]
- UniProt Consortium, T. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2018, 46, 2699. [Google Scholar] [CrossRef] [PubMed]
- Braakman, I.; Hoover-Litty, H.; Wagner, K.R.; Helenius, A. Folding of influenza hemagglutinin in the endoplasmic reticulum. J. Cell Biol. 1991, 114, 401–411. [Google Scholar] [CrossRef]
- Chen, W.; Helenius, J.; Braakman, I.; Helenius, A. Cotranslational folding and calnexin binding during glycoprotein synthesis. Proc. Natl. Acad. Sci. USA 1995, 92, 6229–6233. [Google Scholar] [CrossRef]
- Hebert, D.N.; Zhang, J.X.; Chen, W.; Foellmer, B.; Helenius, A. The number and location of glycans on influenza hemagglutinin determine folding and association with calnexin and calreticulin. J. Cell Biol. 1997, 139, 613–623. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Nakagawa, H.; Tamura, A.; Koshiba, S.; Hoshijima, K.; Komada, M.; Ishikawa, T. Intramolecular disulfide bond is a critical check point determining degradative fates of ATP-binding cassette (ABC) transporter ABCG2 protein. J. Biol. Chem. 2007, 282, 27841–27846. [Google Scholar] [CrossRef]
- Patel, C.; Saad, H.; Shenkman, M.; Lederkremer, G.Z. Oxidoreductases in Glycoprotein Glycosylation, Folding, and ERAD. Cells 2020, 9, 2138. [Google Scholar] [CrossRef]
- Chang, V.T.; Crispin, M.; Aricescu, A.R.; Harvey, D.J.; Nettleship, J.E.; Fennelly, J.A.; Yu, C.; Boles, K.S.; Evans, E.J.; Stuart, D.I.; et al. Glycoprotein structural genomics: Solving the glycosylation problem. Structure 2007, 15, 267–273. [Google Scholar] [CrossRef]
- Hanson, S.R.; Culyba, E.K.; Hsu, T.L.; Wong, C.H.; Kelly, J.W.; Powers, E.T. The core trisaccharide of an N-linked glycoprotein intrinsically accelerates folding and enhances stability. Proc. Natl. Acad. Sci. USA 2009, 106, 3131–3136. [Google Scholar] [CrossRef]
- O’Connor, S.E.; Pohlmann, J.; Imperiali, B.; Saskiawan, I.; Yamamoto, K. Probing the effect of the outer saccharide residues of N-linked glycans on peptide conformation. J. Am. Chem. Soc. 2001, 123, 6187–6188. [Google Scholar] [CrossRef]
- Jitsuhara, Y.; Toyoda, T.; Itai, T.; Yamaguchi, H. Chaperone-like functions of high-mannose type and complex-type N-glycans and their molecular basis. J. Biochem. 2002, 132, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Kimura, N.; Uchida, M.; Nishimura, S.; Yamaguchi, H. Promotion of polypeptide folding by interactions with Asn-Glycans. J. Biochem. 1998, 124, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Rickert, K.W.; Imperiali, B. Analysis of the conserved glycosylation site in the nicotinic acetylcholine receptor: Potential roles in complex assembly. Chem. Biol. 1995, 2, 751–759. [Google Scholar] [CrossRef]
- Matsusaki, M.; Kanemura, S.; Kinoshita, M.; Lee, Y.H.; Inaba, K.; Okumura, M. The Protein Disulfide Isomerase Family: From proteostasis to pathogenesis. Biochim. Et Biophys. Acta. Gen. Subj. 2020, 1864, 129338. [Google Scholar] [CrossRef]
- Robinson, P.J.; Bulleid, N.J. Mechanisms of Disulfide Bond Formation in Nascent Polypeptides Entering the Secretory Pathway. Cells 2020, 9, 1994. [Google Scholar] [CrossRef]
- Shrimal, S.; Cherepanova, N.A.; Gilmore, R. Cotranslational and posttranslocational N-glycosylation of proteins in the endoplasmic reticulum. Semin. Cell Dev. Biol. 2015, 41, 71–78. [Google Scholar] [CrossRef]
- Fu, J.; Gao, J.; Liang, Z.; Yang, D. PDI-regulated disulfide bond formation in protein folding and biomolecular assembly. Molecules 2021, 26, 171. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. The Sep15 protein family: Roles in disulfide bond formation and quality control in the endoplasmic reticulum. IUBMB Life 2007, 59, 1–5. [Google Scholar] [CrossRef]
- Segawa, H.; Inakawa, A.; Yamashita, T.; Taira, H. Functional analysis of individual oligosaccharide chains of Sendai virus hemagglutinin-neuraminidase protein. Biosci. Biotechnol. Biochem. 2003, 67, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Z.; Settembre, E.C.; Aoki, S.T.; Zhang, X.; Bellamy, A.R.; Dormitzer, P.R.; Harrison, S.C.; Grigorieff, N. Molecular interactions in rotavirus assembly and uncoating seen by high-resolution cryo-EM. Proc. Natl. Acad. Sci. USA 2009, 106, 10644–10648. [Google Scholar] [CrossRef] [PubMed]
- Arigoni-Affolter, I.; Scibona, E.; Lin, C.W.; Bruhlmann, D.; Souquet, J.; Broly, H.; Aebi, M. Mechanistic reconstruction of glycoprotein secretion through monitoring of intracellular N-glycan processing. Sci. Adv. 2019, 5, eaax8930. [Google Scholar] [CrossRef]
- Pace, N.J.; Weerapana, E. Diverse functional roles of reactive cysteines. ACS Chem. Biol. 2013, 8, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, K.; Marth, J.D. Glycosylation in cellular mechanisms of health and disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef]
- Fewell, S.W.; Travers, K.J.; Weissman, J.S.; Brodsky, J.L. The action of molecular chaperones in the early secretory pathway. Annu. Rev. Genet. 2001, 35, 149–191. [Google Scholar] [CrossRef]
- Bolmstedt, A.; Hemming, A.; Flodby, P.; Berntsson, P.; Travis, B.; Lin, J.P.; Ledbetter, J.; Tsu, T.; Wigzell, H.; Hu, S.L.; et al. Effects of mutations in glycosylation sites and disulphide bonds on processing, CD4-binding and fusion activity of human immunodeficiency virus envelope glycoproteins. J. Gen. Virol. 1991, 72 Pt 6, 1269–1277. [Google Scholar] [CrossRef]
- Zhu, Q.; Kao, L.; Azimov, R.; Abuladze, N.; Newman, D.; Kurtz, I. Interplay between disulfide bonding and N-glycosylation defines SLC4 Na+-coupled transporter extracellular topography. J. Biol. Chem. 2015, 290, 5391–5404. [Google Scholar] [CrossRef]
- Haniu, M.; Horan, T.; Arakawa, T.; Le, J.; Katta, V.; Hara, S.; Rohde, M.F. Disulfide structure and N-glycosylation sites of an extracellular domain of granulocyte-colony stimulating factor receptor. Biochemistry 1996, 35, 13040–13046. [Google Scholar] [CrossRef]
- Dekker, J.; Strous, G.J. Covalent oligomerization of rat gastric mucin occurs in the rough endoplasmic reticulum, is N-glycosylation-dependent, and precedes initial O-glycosylation. J. Biol. Chem. 1990, 265, 18116–18122. [Google Scholar] [CrossRef]
- Yi, L.; Bozkurt, G.; Li, Q.; Lo, S.; Menon, A.K.; Wu, H. Disulfide Bond Formation and N-Glycosylation Modulate Protein-Protein Interactions in GPI-Transamidase (GPIT). Sci. Rep. 2017, 8, 45912. [Google Scholar] [CrossRef] [PubMed]
- Vlahos, C.J.; Wilhelm, O.G.; Hassell, T.; Jaskunas, S.R.; Bang, N.U. Disulfide pairing of the recombinant kringle-2 domain of tissue plasminogen activator produced in Escherichia coli. J. Biol. Chem. 1991, 266, 10070–10072. [Google Scholar] [CrossRef]
- Allen, S.; Naim, H.Y.; Bulleid, N.J. Intracellular folding of tissue-type plasminogen activator. Effects of disulfide bond formation on N-linked glycosylation and secretion. J. Biol. Chem. 1995, 270, 4797–4804. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.A.; Handin, R.I.; Wise, R.J. The interaction of the von Willebrand factor-A1 domain with platelet glycoprotein Ib/IX. The role of glycosylation and disulfide bonding in a monomeric recombinant A1 domain protein. J. Biol. Chem. 1993, 268, 21238–21245. [Google Scholar] [CrossRef]
- Marti, T.; Rosselet, S.J.; Titani, K.; Walsh, K.A. Identification of disulfide-bridged substructures within human von Willebrand factor. Biochemistry 1987, 26, 8099–8109. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, T.A.; Goode, E.C.; Birdsey, G.M.; Nowak, A.A.; Chan, A.C.; Lane, D.A.; Laffan, M.A. Specific N-linked glycosylation sites modulate synthesis and secretion of von Willebrand factor. Blood 2010, 116, 640–648. [Google Scholar] [CrossRef]
- Feng, W.; Huth, J.R.; Norton, S.E.; Ruddon, R.W. Asparagine-linked oligosaccharides facilitate human chorionic gonadotropin beta-subunit folding but not assembly of prefolded beta with alpha. Endocrinology 1995, 136, 52–61. [Google Scholar] [CrossRef]
- Feng, W.; Matzuk, M.M.; Mountjoy, K.; Bedows, E.; Ruddon, R.W.; Boime, I. The asparagine-linked oligosaccharides of the human chorionic gonadotropin beta subunit facilitate correct disulfide bond pairing. J. Biol. Chem. 1995, 270, 11851–11859. [Google Scholar] [CrossRef]
- Moriwaki, T.; Suganuma, N.; Furuhashi, M.; Kikkawa, F.; Tomoda, Y.; Boime, I.; Nakata, M.; Mizuochi, T. Alteration of N-linked oligosaccharide structures of human chorionic gonadotropin beta-subunit by disruption of disulfide bonds. Glycoconj. J. 1997, 14, 225–229. [Google Scholar] [CrossRef]
- Kuglstatter, A.; Stihle, M.; Neumann, C.; Muller, C.; Schaefer, W.; Klein, C.; Benz, J. Structural differences between glycosylated, disulfide-linked heterodimeric Knob-into-Hole Fc fragment and its homodimeric Knob-Knob and Hole-Hole side products. Protein Eng. Des. Sel. PEDS 2017, 30, 649–656. [Google Scholar] [CrossRef]
- Lipari, F.; Herscovics, A. Role of the cysteine residues in the alpha1,2-mannosidase involved in N-glycan biosynthesis in Saccharomyces cerevisiae. The conserved Cys340 and Cys385 residues form an essential disulfide bond. J. Biol. Chem. 1996, 271, 27615–27622. [Google Scholar] [CrossRef] [PubMed]
- Flahaut, M.; Pfister, C.; Rossier, B.C.; Firsov, D. N-Glycosylation and conserved cysteine residues in RAMP3 play a critical role for the functional expression of CRLR/RAMP3 adrenomedullin receptor. Biochemistry 2003, 42, 10333–10341. [Google Scholar] [CrossRef]
- Walker, A.K.; Soo, K.Y.; Levina, V.; Talbo, G.H.; Atkin, J.D. N-linked glycosylation modulates dimerization of protein disulfide isomerase family A member 2 (PDIA2). FEBS J. 2013, 280, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; Mottet, G.; Kolakofsky, D.; Roux, L. Addition of high-mannose sugars must precede disulfide bond formation for proper folding of Sendai virus glycoproteins. J. Virol. 1989, 63, 892–900. [Google Scholar] [CrossRef] [PubMed]
- McGinnes, L.W.; Morrison, T.G. Disulfide bond formation is a determinant of glycosylation site usage in the hemagglutinin-neuraminidase glycoprotein of Newcastle disease virus. J. Virol. 1997, 71, 3083–3089. [Google Scholar] [CrossRef]
- Pitt, J.J.; Da Silva, E.; Gorman, J.J. Determination of the disulfide bond arrangement of Newcastle disease virus hemagglutinin neuraminidase. Correlation with a beta-sheet propeller structural fold predicted for paramyxoviridae attachment proteins. J. Biol. Chem. 2000, 275, 6469–6478. [Google Scholar] [CrossRef]
- Daniels, R.; Kurowski, B.; Johnson, A.E.; Hebert, D.N. N-linked glycans direct the cotranslational folding pathway of influenza hemagglutinin. Mol. Cell 2003, 11, 79–90. [Google Scholar] [CrossRef]
- Thornlow, D.N.; Macintyre, A.N.; Oguin, T.H.; Karlsson, A.B.; Stover, E.L.; Lynch, H.E.; Sempowski, G.D.; Schmidt, A.G. Altering the Immunogenicity of Hemagglutinin Immunogens by Hyperglycosylation and Disulfide Stabilization. Front. Immunol. 2021, 12, 737973. [Google Scholar] [CrossRef]
- Suzuki, S.; Furuhashi, M.; Suganuma, N. Additional N-glycosylation at Asn(13) rescues the human LHbeta-subunit from disulfide-linked aggregation. Mol. Cell. Endocrinol. 2000, 160, 157–163. [Google Scholar] [CrossRef]
- Naessens, S.; De Zaeytijd, J.; Syx, D.; Vandenbroucke, R.E.; Smeets, F.; Van Cauwenbergh, C.; Leroy, B.P.; Peelman, F.; Coppieters, F. The N-terminal p.(Ser38Cys) TIMP3 mutation underlying Sorsby fundus dystrophy is a founder mutation disrupting an intramolecular disulfide bond. Hum. Mutat. 2019, 40, 539–551. [Google Scholar] [CrossRef]
- Arolas, J.L.; Broder, C.; Jefferson, T.; Guevara, T.; Sterchi, E.E.; Bode, W.; Stocker, W.; Becker-Pauly, C.; Gomis-Ruth, F.X. Structural basis for the sheddase function of human meprin beta metalloproteinase at the plasma membrane. Proc. Natl. Acad. Sci. USA 2012, 109, 16131–16136. [Google Scholar] [CrossRef] [PubMed]
- Ishmael, S.S.; Ishmael, F.T.; Jones, A.D.; Bond, J.S. Protease domain glycans affect oligomerization, disulfide bond formation, and stability of the meprin A metalloprotease homo-oligomer. J. Biol. Chem. 2006, 281, 37404–37415. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Qian, W.J.; Gritsenko, M.A.; Camp, D.G., 2nd; Monroe, M.E.; Moore, R.J.; Smith, R.D. Human plasma N-glycoproteome analysis by immunoaffinity subtraction, hydrazide chemistry, and mass spectrometry. J. Proteome Res. 2005, 4, 2070–2080. [Google Scholar] [CrossRef]
- McCormick, L.M.; Urade, R.; Arakaki, Y.; Schwartz, A.L.; Bu, G. Independent and cooperative roles of N-glycans and molecular chaperones in the folding and disulfide bond formation of the low-density lipoprotein (LDL) receptor-related protein. Biochemistry 2005, 44, 5794–5803. [Google Scholar] [CrossRef] [PubMed]
- Zelcer, N.; Hong, C.; Boyadjian, R.; Tontonoz, P. LXR regulates cholesterol uptake through Idol-dependent ubiquitination of the LDL receptor. Science 2009, 325, 100–104. [Google Scholar] [CrossRef]
- Asker, N.; Axelsson, M.A.; Olofsson, S.O.; Hansson, G.C. Dimerization of the human MUC2 mucin in the endoplasmic reticulum is followed by a N-glycosylation-dependent transfer of the mono- and dimers to the Golgi apparatus. J. Biol. Chem. 1998, 273, 18857–18863. [Google Scholar] [CrossRef]
- Bell, S.L.; Xu, G.; Khatri, I.A.; Wang, R.; Rahman, S.; Forstner, J.F. N-linked oligosaccharides play a role in disulphide-dependent dimerization of intestinal mucin Muc2. Biochem. J. 2003, 373, 893–900. [Google Scholar] [CrossRef]
- Bosques, C.J.; Imperiali, B. The interplay of glycosylation and disulfide formation influences fibrillization in a prion protein fragment. Proc. Natl. Acad. Sci. USA 2003, 100, 7593–7598. [Google Scholar] [CrossRef]
- Beggah, A.T.; Jaunin, P.; Geering, K. Role of glycosylation and disulfide bond formation in the beta subunit in the folding and functional expression of Na, K-ATPase. J. Biol. Chem. 1997, 272, 10318–10326. [Google Scholar] [CrossRef]
- Laughery, M.D.; Todd, M.L.; Kaplan, J.H. Mutational analysis of alpha-beta subunit interactions in the delivery of Na, K-ATPase heterodimers to the plasma membrane. J. Biol. Chem. 2003, 278, 34794–34803. [Google Scholar] [CrossRef]
- Clogston, C.L.; Boone, T.C.; Crandall, B.C.; Mendiaz, E.A.; Lu, H.S. Disulfide structures of human interleukin-6 are similar to those of human granulocyte colony stimulating factor. Arch. Biochem. Biophys. 1989, 272, 144–151. [Google Scholar] [CrossRef]
- May, L.T.; Shaw, J.E.; Khanna, A.K.; Zabriskie, J.B.; Sehgal, P.B. Marked cell-type-specific differences in glycosylation of human interleukin-6. Cytokine 1991, 3, 204–211. [Google Scholar] [CrossRef]
- Orita, T.; Oh-eda, M.; Hasegawa, M.; Kuboniwa, H.; Esaki, K.; Ochi, N. Polypeptide and carbohydrate structure of recombinant human interleukin-6 produced in Chinese hamster ovary cells. J. Biochem. 1994, 115, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Roman-Sosa, G.; Karger, A.; Kraatz, F.; Aebischer, A.; Wernike, K.; Maksimov, P.; Lillig, C.H.; Reimann, I.; Brocchi, E.; Keller, M.; et al. The amino terminal subdomain of glycoprotein Gc of Schmallenberg virus: Disulfide bonding and structural determinants of neutralization. J. Gen. Virol. 2017, 98, 1259–1273. [Google Scholar] [CrossRef]
- Mohd Yusuf, S.N.; Bailey, U.M.; Tan, N.Y.; Jamaluddin, M.F.; Schulz, B.L. Mixed disulfide formation in vitro between a glycoprotein substrate and yeast oligosaccharyltransferase subunits Ost3p and Ost6p. Biochem. Biophys. Res. Commun. 2013, 432, 438–443. [Google Scholar] [CrossRef]
- Schulz, B.L.; Stirnimann, C.U.; Grimshaw, J.P.; Brozzo, M.S.; Fritsch, F.; Mohorko, E.; Capitani, G.; Glockshuber, R.; Grutter, M.G.; Aebi, M. Oxidoreductase activity of oligosaccharyltransferase subunits Ost3p and Ost6p defines site-specific glycosylation efficiency. Proc. Natl. Acad. Sci. USA 2009, 106, 11061–11066. [Google Scholar] [CrossRef]
- Ohgomori, T.; Nanao, T.; Morita, A.; Ikekita, M. Asn54-linked glycan is critical for functional folding of intercellular adhesion molecule-5. Glycoconj. J. 2012, 29, 47–55. [Google Scholar] [CrossRef]
- Zhang, H.; Casasnovas, J.M.; Jin, M.; Liu, J.H.; Gahmberg, C.G.; Springer, T.A.; Wang, J.H. An unusual allosteric mobility of the C-terminal helix of a high-affinity alphaL integrin I domain variant bound to ICAM-5. Mol. Cell 2008, 31, 432–437. [Google Scholar] [CrossRef]
- Kondo, J.; Shibata, H.; Miura, S.; Yamakawa, A.; Sato, K.; Higuchi, Y.; Shukunami, C.; Hiraki, Y. A functional role of the glycosylated N-terminal domain of chondromodulin-I. J. Bone Miner. Metab. 2011, 29, 23–30. [Google Scholar] [CrossRef]
- Hoffman, R.C.; Andersen, H.; Walker, K.; Krakover, J.D.; Patel, S.; Stamm, M.R.; Osborn, S.G. Peptide, disulfide, and glycosylation mapping of recombinant human thrombopoietin from ser1 to Arg246. Biochemistry 1996, 35, 14849–14861. [Google Scholar] [CrossRef]
- Kato, T. Protein characteristics of thrombopoietin. Stem Cells 1996, 14 (Suppl. S1), 139–147. [Google Scholar] [CrossRef] [PubMed]
- Diop, N.K.; Hrycyna, C.A. N-Linked glycosylation of the human ABC transporter ABCG2 on asparagine 596 is not essential for expression, transport activity, or trafficking to the plasma membrane. Biochemistry 2005, 44, 5420–5429. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi-Nakao, K.; Tamura, A.; Furukawa, T.; Nakagawa, H.; Ishikawa, T. Quality control of human ABCG2 protein in the endoplasmic reticulum: Ubiquitination and proteasomal degradation. Adv. Drug Deliv. Rev. 2009, 61, 66–72. [Google Scholar] [CrossRef]
- Fischer, F.; Molinari, M.; Bodendorf, U.; Paganetti, P. The disulphide bonds in the catalytic domain of BACE are critical but not essential for amyloid precursor protein processing activity. J. Neurochem. 2002, 80, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Cherepanova, N.A.; Venev, S.V.; Leszyk, J.D.; Shaffer, S.A.; Gilmore, R. Quantitative glycoproteomics reveals new classes of STT3A- and STT3B-dependent N-glycosylation sites. J. Cell Biol. 2019, 218, 2782–2796. [Google Scholar] [CrossRef]
- Cherepanova, N.A.; Shrimal, S.; Gilmore, R. Oxidoreductase activity is necessary for N-glycosylation of cysteine-proximal acceptor sites in glycoproteins. J. Cell Biol. 2014, 206, 525–539. [Google Scholar] [CrossRef]
- Ruiz-Canada, C.; Kelleher, D.J.; Gilmore, R. Cotranslational and posttranslational N-glycosylation of polypeptides by distinct mammalian OST isoforms. Cell 2009, 136, 272–283. [Google Scholar] [CrossRef]
- Rius, M.; Sala, L.; Chillaron, J. The role of N-glycans and the C-terminal loop of the subunit rBAT in the biogenesis of the cystinuria-associated transporter. Biochem. J. 2016, 473, 233–244. [Google Scholar] [CrossRef]
- Ivanenkov, V.V.; Meller, J.; Kirley, T.L. Characterization of disulfide bonds in human nucleoside triphosphate diphosphohydrolase 3 (NTPDase3): Implications for NTPDase structural modeling. Biochemistry 2005, 44, 8998–9012. [Google Scholar] [CrossRef]
- Qiu, Z.; Hobman, T.C.; McDonald, H.L.; Seto, N.O.; Gillam, S. Role of N-linked oligosaccharides in processing and intracellular transport of E2 glycoprotein of rubella virus. J. Virol. 1992, 66, 3514–3521. [Google Scholar] [CrossRef]
- Alhamidi, M.; Kjeldsen Buvang, E.; Fagerheim, T.; Brox, V.; Lindal, S.; Van Ghelue, M.; Nilssen, O. Fukutin-related protein resides in the Golgi cisternae of skeletal muscle fibres and forms disulfide-linked homodimers via an N-terminal interaction. PLoS ONE 2011, 6, e22968. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Kraetzner, R.; Gruene, T.; Grapp, M.; Schreiber, K.; Gronborg, M.; Urlaub, H.; Becker, S.; Asif, A.R.; Gartner, J.; et al. Structure of tripeptidyl-peptidase I provides insight into the molecular basis of late infantile neuronal ceroid lipofuscinosis. J. Biol. Chem. 2009, 284, 3976–3984. [Google Scholar] [CrossRef] [PubMed]
- Wujek, P.; Kida, E.; Walus, M.; Wisniewski, K.E.; Golabek, A.A. N-glycosylation is crucial for folding, trafficking, and stability of human tripeptidyl-peptidase I. J. Biol. Chem. 2004, 279, 12827–12839. [Google Scholar] [CrossRef]
- Chen, R.; Jiang, X.; Sun, D.; Han, G.; Wang, F.; Ye, M.; Wang, L.; Zou, H. Glycoproteomics analysis of human liver tissue by combination of multiple enzyme digestion and hydrazide chemistry. J. Proteome Res. 2009, 8, 651–661. [Google Scholar] [CrossRef]
- Guhaniyogi, J.; Sohar, I.; Das, K.; Stock, A.M.; Lobel, P. Crystal structure and autoactivation pathway of the precursor form of human tripeptidyl-peptidase 1, the enzyme deficient in late infantile ceroid lipofuscinosis. J. Biol. Chem. 2009, 284, 3985–3997. [Google Scholar] [CrossRef] [PubMed]
- Wollscheid, B.; Bausch-Fluck, D.; Henderson, C.; O’Brien, R.; Bibel, M.; Schiess, R.; Aebersold, R.; Watts, J.D. Mass-spectrometric identification and relative quantification of N-linked cell surface glycoproteins. Nat. Biotechnol. 2009, 27, 378–386. [Google Scholar] [CrossRef]
- Koch, M.; Stronge, V.S.; Shepherd, D.; Gadola, S.D.; Mathew, B.; Ritter, G.; Fersht, A.R.; Besra, G.S.; Schmidt, R.R.; Jones, E.Y.; et al. The crystal structure of human CD1d with and without alpha-galactosylceramide. Nat Immunol 2005, 6, 819–826. [Google Scholar] [CrossRef]
- Vallee, F.; Lipari, F.; Yip, P.; Sleno, B.; Herscovics, A.; Howell, P.L. Crystal structure of a class I alpha1,2-mannosidase involved in N-glycan processing and endoplasmic reticulum quality control. EMBO J. 2000, 19, 581–588. [Google Scholar] [CrossRef]
- Borg, N.A.; Wun, K.S.; Kjer-Nielsen, L.; Wilce, M.C.; Pellicci, D.G.; Koh, R.; Besra, G.S.; Bharadwaj, M.; Godfrey, D.I.; McCluskey, J.; et al. CD1d-lipid-antigen recognition by the semi-invariant NKT T-cell receptor. Nature 2007, 448, 44–49. [Google Scholar] [CrossRef]
- Honer, J.; Niemeyer, K.M.; Fercher, C.; Diez Tissera, A.L.; Jaberolansar, N.; Jafrani, Y.M.A.; Zhou, C.; Caramelo, J.J.; Shewan, A.M.; Schulz, B.L.; et al. TorsinA folding and N-linked glycosylation are sensitive to redox homeostasis. Biochim. Et Biophys. Acta. Mol. Cell Res. 2021, 1868, 119073. [Google Scholar] [CrossRef]
- Kustedjo, K.; Bracey, M.H.; Cravatt, B.F. Torsin A and its torsion dystonia-associated mutant forms are lumenal glycoproteins that exhibit distinct subcellular localizations. J. Biol. Chem. 2000, 275, 27933–27939. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.C.; Schekman, R. Role of Sec61p in the ER-associated degradation of short-lived transmembrane proteins. J. Cell Biol. 2008, 181, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, M.; Koyama, T.; Nakazawa, F.; Nishio, M.; Shibamiya, A.; Hirosawa, S. Gradually glycosylated protein C mutants (Arg178Gln and Cys331Arg) are degraded by proteasome after mannose trimming. Thromb Haemost 2004, 92, 1284–1290. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, Q.; Yuan, L.; Zhang, R.; Li, L. N-glycosylation and dimerization regulate the PtrMAN6 enzyme activity that may modulate generation of oligosaccharide signals. Plant Signal Behav. 2013, 8, e26956. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gardner, T.G.; Kearse, K.P. Modification of the T cell antigen receptor (TCR) complex by UDP-glucose:glycoprotein glucosyltransferase. TCR folding is finalized convergent with formation of alpha beta delta epsilon gamma epsilon complexes. J. Biol. Chem. 1999, 274, 14094–14099. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Lu, Z.; Fu, Y.; Wang, H.P.; Wang, L.H.; Chi, H.; Yuan, Z.F.; Zheng, Z.B.; Song, L.N.; Han, H.H.; et al. A strategy for precise and large scale identification of core fucosylated glycoproteins. Mol Cell Proteom. 2009, 8, 913–923. [Google Scholar] [CrossRef]
- Thaysen-Andersen, M.; Mysling, S.; Hojrup, P. Site-specific glycoprofiling of N-linked glycopeptides using MALDI-TOF MS: Strong correlation between signal strength and glycoform quantities. Anal. Chem. 2009, 81, 3933–3943. [Google Scholar] [CrossRef]
- Frank, M.; Walker, R.C.; Lanzilotta, W.N.; Prestegard, J.H.; Barb, A.W. Immunoglobulin G1 Fc domain motions: Implications for Fc engineering. J. Mol. Biol. 2014, 426, 1799–1811. [Google Scholar] [CrossRef]
- Vance, B.A.; Bennett, M.J.; Ward, Y.; Gress, R.G.; Kearse, K.P. Distinct but dispensable N-glycosylation of human CD69 proteins. Arch. Biochem. Biophys. 1999, 368, 214–220. [Google Scholar] [CrossRef]
- Vance, B.A.; Wu, W.; Ribaudo, R.K.; Segal, D.M.; Kearse, K.P. Multiple dimeric forms of human CD69 result from differential addition of N-glycans to typical (Asn-X-Ser/Thr) and atypical (Asn-X-cys) glycosylation motifs. J. Biol. Chem. 1997, 272, 23117–23122. [Google Scholar] [CrossRef]
- Kang, S.J.; Cresswell, P. Calnexin, calreticulin, and ERp57 cooperate in disulfide bond formation in human CD1d heavy chain. J. Biol. Chem. 2002, 277, 44838–44844. [Google Scholar] [CrossRef] [PubMed]
- Paduraru, C.; Spiridon, L.; Yuan, W.; Bricard, G.; Valencia, X.; Porcelli, S.A.; Illarionov, P.A.; Besra, G.S.; Petrescu, S.M.; Petrescu, A.J.; et al. An N-linked glycan modulates the interaction between the CD1d heavy chain and beta 2-microglobulin. J. Biol. Chem. 2006, 281, 40369–40378. [Google Scholar] [CrossRef] [PubMed]
- Dick, T.P.; Bangia, N.; Peaper, D.R.; Cresswell, P. Disulfide bond isomerization and the assembly of MHC class I-peptide complexes. Immunity 2002, 16, 87–98. [Google Scholar] [CrossRef]
- Wearsch, P.A.; Peaper, D.R.; Cresswell, P. Essential glycan-dependent interactions optimize MHC class I peptide loading. Proc. Natl. Acad. Sci. USA 2011, 108, 4950–4955. [Google Scholar] [CrossRef]
- Mirazimi, A.; Svensson, L. Carbohydrates facilitate correct disulfide bond formation and folding of rotavirus VP7. J. Virol. 1998, 72, 3887–3892. [Google Scholar] [CrossRef]
- McGinnes, L.W.; Wilde, A.; Morrison, T.G. Nucleotide sequence of the gene encoding the Newcastle disease virus hemagglutinin-neuraminidase protein and comparisons of paramyxovirus hemagglutinin-neuraminidase protein sequences. Virus Res. 1987, 7, 187–202. [Google Scholar] [CrossRef]
- McGinnes, L.W.; Morrison, T.G. The role of the individual cysteine residues in the formation of the mature, antigenic HN protein of Newcastle disease virus. Virology 1994, 200, 470–483. [Google Scholar] [CrossRef]
- Mirza, A.M.; Sheehan, J.P.; Hardy, L.W.; Glickman, R.L.; Iorio, R.M. Structure and function of a membrane anchor-less form of the hemagglutinin-neuraminidase glycoprotein of Newcastle disease virus. J. Biol. Chem. 1993, 268, 21425–21431. [Google Scholar] [CrossRef]
- Kao, L.; Sassani, P.; Azimov, R.; Pushkin, A.; Abuladze, N.; Peti-Peterdi, J.; Liu, W.; Newman, D.; Kurtz, I. Oligomeric structure and minimal functional unit of the electrogenic sodium bicarbonate cotransporter NBCe1-A. J. Biol. Chem. 2008, 283, 26782–26794. [Google Scholar] [CrossRef]
- Hemming, A.; Bolmstedt, A.; Flodby, P.; Lundberg, L.; Gidlund, M.; Wigzell, H.; Olofsson, S.O. Cysteine 402 of HIV gp120 is essential for CD4-binding and resistance of gp120 to intracellular degradation. Arch. Virol. 1989, 109, 269–276. [Google Scholar] [CrossRef]
- Tschachler, E.; Buchow, H.; Gallo, R.C.; Reitz, M.S., Jr. Functional contribution of cysteine residues to the human immunodeficiency virus type 1 envelope. J. Virol. 1990, 64, 2250–2259. [Google Scholar] [CrossRef] [PubMed]
- Norskov-Lauritsen, L.; Jorgensen, S.; Brauner-Osborne, H. N-glycosylation and disulfide bonding affects GPRC6A receptor expression, function, and dimerization. FEBS Lett. 2015, 589, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Pi, M.; Nishimoto, S.K.; Quarles, L.D. GPRC6A: Jack of all metabolism (or master of none). Mol. Metab. 2017, 6, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Clemmensen, C.; Smajilovic, S.; Wellendorph, P.; Brauner-Osborne, H. The GPCR, class C, group 6, subtype A (GPRC6A) receptor: From cloning to physiological function. Br. J. Pharmacol. 2014, 171, 1129–1141. [Google Scholar] [CrossRef]
- Muto, T.; Tsuchiya, D.; Morikawa, K.; Jingami, H. Structures of the extracellular regions of the group II/III metabotropic glutamate receptors. Proc. Natl. Acad. Sci. USA 2007, 104, 3759–3764. [Google Scholar] [CrossRef]
- Rider, P.J.F.; Naderi, M.; Bergeron, S.; Chouljenko, V.N.; Brylinski, M.; Kousoulas, K.G. Cysteines and N-Glycosylation Sites Conserved among All Alphaherpesviruses Regulate Membrane Fusion in Herpes Simplex Virus 1 Infection. J. Virol. 2017, 91, e00873-17. [Google Scholar] [CrossRef]
- Neubauer, A.; Osterrieder, N. Equine herpesvirus type 1 (EHV-1) glycoprotein K is required for efficient cell-to-cell spread and virus egress. Virology 2004, 329, 18–32. [Google Scholar] [CrossRef]
- Zhang, S.; Go, E.P.; Ding, H.; Anang, S.; Kappes, J.C.; Desaire, H.; Sodroski, J. Analysis of glycosylation and disulfide bonding of wild-type SARS-CoV-2 spike glycoprotein. J. Virol. 2022, 96, e01626-21. [Google Scholar] [CrossRef]
- Brun, J.; Vasiljevic, S.; Gangadharan, B.; Hensen, M.; Chandran, A.V.; Hill, M.L.; Kiappes, J.L.; Dwek, R.A.; Alonzi, D.S.; Struwe, W.B.; et al. Analysis of SARS-CoV-2 spike glycosylation reveals shedding of a vaccine candidate. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wang, D.; Baudys, J.; Bundy, J.L.; Solano, M.; Keppel, T.; Barr, J.R. Comprehensive Analysis of the Glycan Complement of SARS-CoV-2 Spike Proteins Using Signature Ions-Triggered Electron-Transfer/Higher-Energy Collisional Dissociation (EThcD) Mass Spectrometry. Anal. Chem. 2020, 92, 14730–14739. [Google Scholar] [CrossRef]
- Yao, H.; Song, Y.; Chen, Y.; Wu, N.; Xu, J.; Sun, C.; Zhang, J.; Weng, T.; Zhang, Z.; Wu, Z.; et al. Molecular Architecture of the SARS-CoV-2 Virus. Cell 2020, 183, 730–738.e13. [Google Scholar] [CrossRef]
- Sanda, M.; Morrison, L.; Goldman, R. N and O glycosylation of the SARS-CoV-2 spike protein. Anal. Chem. 2021, 93, 2003–2009. [Google Scholar] [CrossRef]
- Watanabe, Y.; Allen, J.D.; Wrapp, D.; McLellan, J.S.; Crispin, M. Site-specific glycan analysis of the SARS-CoV-2 spike. Science 2020, 369, 330–333. [Google Scholar] [CrossRef]
- Bangaru, S.; Ozorowski, G.; Turner, H.L.; Antanasijevic, A.; Huang, D.; Wang, X.; Torres, J.L.; Diedrich, J.K.; Tian, J.H.; Portnoff, A.D.; et al. Structural analysis of full-length SARS-CoV-2 spike protein from an advanced vaccine candidate. Science 2020, 370, 1089–1094. [Google Scholar] [CrossRef]
- Zhao, P.; Praissman, J.L.; Grant, O.C.; Cai, Y.; Xiao, T.; Rosenbalm, K.E.; Aoki, K.; Kellman, B.P.; Bridger, R.; Barouch, D.H.; et al. Virus-Receptor Interactions of Glycosylated SARS-CoV-2 Spike and Human ACE2 Receptor. Cell Host Microbe 2020, 28, 586–601 e586. [Google Scholar] [CrossRef]
- Hati, S.; Bhattacharyya, S. Impact of Thiol-Disulfide Balance on the Binding of Covid-19 Spike Protein with Angiotensin-Converting Enzyme 2 Receptor. ACS Omega 2020, 5, 16292–16298. [Google Scholar] [CrossRef]
- Singh, J.; Dhindsa, R.S.; Misra, V.; Singh, B. SARS-CoV2 infectivity is potentially modulated by host redox status. Comput. Struct. Biotechnol. J. 2020, 18, 3705–3711. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Mehdipour, A.R.; Hummer, G. Dual nature of human ACE2 glycosylation in binding to SARS-CoV-2 spike. Proc. Natl. Acad. Sci. USA 2021, 118, e2100425118. [Google Scholar] [CrossRef]
- Roth, Z.; Yehezkel, G.; Khalaila, I. Identification and Quantification of Protein Glycosylation. Int. J. Carbohydr. Chem. 2012, 2012, 640923. [Google Scholar] [CrossRef]
- Gimeno, A.; Valverde, P.; Arda, A.; Jimenez-Barbero, J. Glycan structures and their interactions with proteins. A NMR view. Curr. Opin. Struct. Biol. 2020, 62, 22–30. [Google Scholar] [CrossRef]
- Imperiali, B.; O’Connor, S.E. Effect of N-linked glycosylation on glycopeptide and glycoprotein structure. Curr. Opin. Chem. Biol. 1999, 3, 643–649. [Google Scholar] [CrossRef]
- Kozak, S.; Bloch, Y.; De Munck, S.; Mikula, A.; Bento, I.; Savvides, S.N.; Meijers, R. Homogeneously N-glycosylated proteins derived from the GlycoDelete HEK293 cell line enable diffraction-quality crystallogenesis. Acta Crystallogr. Sect. D Struct. Biol. 2020, 76, 1244–1255. [Google Scholar] [CrossRef]
- Freeze, H.H.; Kranz, C. Endoglycosidase and glycoamidase release of N-linked glycans. Curr. Protoc. Mol. Biol. 2010, 89, 17.13A.1–17.13A.25. [Google Scholar] [CrossRef]
- Kinsella, M.G.; Wight, T.N. Isolation and characterization of dermatan sulfate proteoglycans synthesized by cultured bovine aortic endothelial cells. J. Biol. Chem. 1988, 263, 19222–19231. [Google Scholar] [CrossRef]
- Guile, G.R.; Wong, S.Y.; Dwek, R.A. Analytical and preparative separation of anionic oligosaccharides by weak anion-exchange high-performance liquid chromatography on an inert polymer column. Anal. Biochem. 1994, 222, 231–235. [Google Scholar] [CrossRef]
- Townsend, R.R.; Hardy, M.R. Analysis of glycoprotein oligosaccharides using high-pH anion exchange chromatography. Glycobiology 1991, 1, 139–147. [Google Scholar] [CrossRef]
- Guile, G.R.; Rudd, P.M.; Wing, D.R.; Prime, S.B.; Dwek, R.A. A rapid high-resolution high-performance liquid chromatographic method for separating glycan mixtures and analyzing oligosaccharide profiles. Anal. Biochem. 1996, 240, 210–226. [Google Scholar] [CrossRef]
- Guile, G.R.; Harvey, D.J.; O’Donnell, N.; Powell, A.K.; Hunter, A.P.; Zamze, S.; Fernandes, D.L.; Dwek, R.A.; Wing, D.R. Identification of highly fucosylated N-linked oligosaccharides from the human parotid gland. Eur. J. Biochem. 1998, 258, 623–656. [Google Scholar] [CrossRef]
- Han, L.; Costello, C.E. Mass spectrometry of glycans. Biochem. Biokhimiia 2013, 78, 710–720. [Google Scholar] [CrossRef]
- Harvey, D.J. Matrix-assisted laser desorption/ionisation mass spectrometry of oligosaccharides and glycoconjugates. J. Chromatogr. A 1996, 720, 429–446. [Google Scholar] [CrossRef]
- Illiano, A.; Pinto, G.; Melchiorre, C.; Carpentieri, A.; Faraco, V.; Amoresano, A. Protein Glycosylation Investigated by Mass Spectrometry: An Overview. Cells 2020, 9, 1986. [Google Scholar] [CrossRef] [PubMed]
- Mechref, Y.; Novotny, M.V. Structural investigations of glycoconjugates at high sensitivity. Chem. Rev. 2002, 102, 321–369. [Google Scholar] [CrossRef]
- Sagi, D.; Kienz, P.; Denecke, J.; Marquardt, T.; Peter-Katalinic, J. Glycoproteomics of N-glycosylation by in-gel deglycosylation and matrix-assisted laser desorption/ionisation-time of flight mass spectrometry mapping: Application to congenital disorders of glycosylation. Proteomics 2005, 5, 2689–2701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, X.J.; Martin, D.B.; Aebersold, R. Identification and quantification of N-linked glycoproteins using hydrazide chemistry, stable isotope labeling and mass spectrometry. Nat. Biotechnol. 2003, 21, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Trinidad, J.C.; Clemmer, D.E. Glycopeptide Site Heterogeneity and Structural Diversity Determined by Combined Lectin Affinity Chromatography/IMS/CID/MS Techniques. J. Am. Soc. Mass Spectrom. 2015, 26, 1092–1102. [Google Scholar] [CrossRef] [PubMed]
- Bloom, J.W.; Madanat, M.S.; Ray, M.K. Cell line and site specific comparative analysis of the N-linked oligosaccharides on human ICAM-1des454-532 by electrospray ionization mass spectrometry. Biochemistry 1996, 35, 1856–1864. [Google Scholar] [CrossRef]
- Bunkenborg, J.; Pilch, B.J.; Podtelejnikov, A.V.; Wisniewski, J.R. Screening for N-glycosylated proteins by liquid chromatography mass spectrometry. Proteomics 2004, 4, 454–465. [Google Scholar] [CrossRef]
- Ford, K.L.; Zeng, W.; Heazlewood, J.L.; Bacic, A. Characterization of protein N-glycosylation by tandem mass spectrometry using complementary fragmentation techniques. Front Plant Sci 2015, 6, 674. [Google Scholar] [CrossRef]
- Esko, J.D.; Bertozzi, C.; Schnaar, R.L. Chemical Tools for Inhibiting Glycosylation. In Essentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor: Huntington, NY, USA, 2015; pp. 701–712. [Google Scholar] [CrossRef]
- Wainwright, L.J.; Field, M.C. Quality control of glycosylphosphatidylinositol anchor attachment in mammalian cells: A biochemical study. Biochem. J. 1997, 321 Pt 3, 655–664. [Google Scholar] [CrossRef]
- Barolo, L.; Abbriano, R.M.; Commault, A.S.; George, J.; Kahlke, T.; Fabris, M.; Padula, M.P.; Lopez, A.; Ralph, P.J.; Pernice, M. Perspectives for Glyco-Engineering of Recombinant Biopharmaceuticals from Microalgae. Cells 2020, 9, 633. [Google Scholar] [CrossRef] [PubMed]
- Weinfurtner, D. Analysis of Disulfide Bond Formation in Therapeutic Proteins. In Oxidative Folding of Proteins: Basic Principles, Cellular Regulation and Engineerining; Royal Society of Chemistry: London, UK, 2018; pp. 81–98. [Google Scholar] [CrossRef]
- Haniu, M.; Acklin, C.; Kenney, W.C.; Rohde, M.F. Direct assignment of disulfide bonds by Edman degradation of selected peptide fragments. Int. J. Pept. Protein Res. 1994, 43, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Klaus, W.; Broger, C.; Gerber, P.; Senn, H. Determination of the disulphide bonding pattern in proteins by local and global analysis of nuclear magnetic resonance data. Application to flavoridin. J. Mol. Biol. 1993, 232, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Rajarathnam, K. 13C NMR chemical shifts can predict disulfide bond formation. J. Biomol. NMR 2000, 18, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.A.; Kjeldgaard, M. Electron-density map interpretation. Methods Enzymol. 1997, 277, 173–208. [Google Scholar] [CrossRef]
- Damodaran, S. Estimation of disulfide bonds using 2-nitro-5-thiosulfobenzoic acid: Limitations. Anal. Biochem. 1985, 145, 200–204. [Google Scholar] [CrossRef]
- Thannhauser, T.W.; Konishi, Y.; Scheraga, H.A. Analysis for disulfide bonds in peptides and proteins. Methods Enzymol. 1987, 143, 115–119. [Google Scholar] [CrossRef]
- Aitken, A.; Learmonth, M. Quantitation of cysteine residues and disulfide bonds by electrophoresis. In The Protein Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 1057–1062. [Google Scholar] [CrossRef]
- Hirose, M.; Takahashi, N.; Oe, H.; Doi, E. Analyses of intramolecular disulfide bonds in proteins by polyacrylamide gel electrophoresis following two-step alkylation. Anal. Biochem. 1988, 168, 193–201. [Google Scholar] [CrossRef]
- Saraswat, R.; McDonagh, B. Diagonal Electrophoresis for the Detection of Proteins Involved in Disulfide Bonds. Methods Mol. Biol. 2019, 1855, 279–286. [Google Scholar] [CrossRef]
- Chrisman, P.A.; Pitteri, S.J.; Hogan, J.M.; McLuckey, S.A. SO2-* electron transfer ion/ion reactions with disulfide linked polypeptide ions. J. Am. Soc. Mass Spectrom. 2005, 16, 1020–1030. [Google Scholar] [CrossRef]
- Cramer, C.N.; Haselmann, K.F.; Olsen, J.V.; Nielsen, P.K. Disulfide Linkage Characterization of Disulfide Bond-Containing Proteins and Peptides by Reducing Electrochemistry and Mass Spectrometry. Anal. Chem. 2016, 88, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Gorman, J.J.; Wallis, T.P.; Pitt, J.J. Protein disulfide bond determination by mass spectrometry. Mass Spectrom. Rev. 2002, 21, 183–216. [Google Scholar] [CrossRef] [PubMed]
- Lakbub, J.C.; Shipman, J.T.; Desaire, H. Recent mass spectrometry-based techniques and considerations for disulfide bond characterization in proteins. Anal. Bioanal. Chem. 2018, 410, 2467–2484. [Google Scholar] [CrossRef]
- Tsai, P.; Chen, S.; Huang, S.Y. Mass spectrometry-based strategies for protein disulfide bond identification. Rev. Anal. Chem. 2013, 32, 257–268. [Google Scholar] [CrossRef]
- Wiesner, J.; Resemann, A.; Evans, C.; Suckau, D.; Jabs, W. Advanced mass spectrometry workflows for analyzing disulfide bonds in biologics. Expert Rev. Proteom. 2015, 12, 115–123. [Google Scholar] [CrossRef]
- Zhou, Y.; Dong, J.; Vachet, R.W. Electron transfer dissociation of modified peptides and proteins. Curr. Pharm. Biotechnol. 2011, 12, 1558–1567. [Google Scholar] [CrossRef]
- Sarbu, M.; Ghiulai, R.M.; Zamfir, A.D. Recent developments and applications of electron transfer dissociation mass spectrometry in proteomics. Amino Acids 2014, 46, 1625–1634. [Google Scholar] [CrossRef]
- Wu, S.L.; Jiang, H.; Hancock, W.S.; Karger, B.L. Identification of the unpaired cysteine status and complete mapping of the 17 disulfides of recombinant tissue plasminogen activator using LC-MS with electron transfer dissociation/collision induced dissociation. Anal. Chem. 2010, 82, 5296–5303. [Google Scholar] [CrossRef][Green Version]
- Lakbub, J.C.; Clark, D.F.; Shah, I.S.; Zhu, Z.; Go, E.P.; Tolbert, T.J.; Desaire, H. Disulfide Bond Characterization of Endogenous IgG3 Monoclonal Antibodies Using LC-MS: An Investigation of IgG3 Disulfide-mediated Isoforms. Anal. Methods 2016, 8, 6046–6055. [Google Scholar] [CrossRef]
- Clark, D.F.; Go, E.P.; Toumi, M.L.; Desaire, H. Collision induced dissociation products of disulfide-bonded peptides: Ions result from the cleavage of more than one bond. J. Am. Soc. Mass Spectrom. 2011, 22, 492–498. [Google Scholar] [CrossRef][Green Version]
- Olsen, J.V.; Macek, B.; Lange, O.; Makarov, A.; Horning, S.; Mann, M. Higher-energy C-trap dissociation for peptide modification analysis. Nat. Methods 2007, 4, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Frese, C.K.; Altelaar, A.F.; van den Toorn, H.; Nolting, D.; Griep-Raming, J.; Heck, A.J.; Mohammed, S. Toward full peptide sequence coverage by dual fragmentation combining electron-transfer and higher-energy collision dissociation tandem mass spectrometry. Anal. Chem. 2012, 84, 9668–9673. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; van Breukelen, B.; Heck, A.J. Facilitating protein disulfide mapping by a combination of pepsin digestion, electron transfer higher energy dissociation (EThcD), and a dedicated search algorithm SlinkS. Mol. Cell Proteom. 2014, 13, 2776–2786. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Diedrich, J.K.; Julian, R.R. Direct elucidation of disulfide bond partners using ultraviolet photodissociation mass spectrometry. Anal. Chem. 2011, 83, 6455–6458. [Google Scholar] [CrossRef]
- Suga, A.; Nagae, M.; Yamaguchi, Y. Analysis of protein landscapes around N-glycosylation sites from the PDB repository for understanding the structural basis of N-glycoprotein processing and maturation. Glycobiology 2018, 28, 774–785. [Google Scholar] [CrossRef]
- Wu, D.; Struwe, W.B.; Harvey, D.J.; Ferguson, M.A.J.; Robinson, C.V. N-glycan microheterogeneity regulates interactions of plasma proteins. Proc. Natl. Acad. Sci. USA 2018, 115, 8763–8768. [Google Scholar] [CrossRef]
- Bendiak, B.; Ward, L.D.; Simpson, R.J. Proteins of the Golgi apparatus. Purification to homogeneity, N-terminal sequence, and unusually large Stokes radius of the membrane-bound form of UDP-galactose:N-acetylglucosamine beta 1-4galactosyltransferase from rat liver. Eur. J. Biochem. 1993, 216, 405–417. [Google Scholar] [CrossRef]
- Krambeck, F.J.; Betenbaugh, M.J. A mathematical model of N-linked glycosylation. Biotechnol. Bioeng. 2005, 92, 711–728. [Google Scholar] [CrossRef]
- Krambeck, F.J.; Bennun, S.V.; Andersen, M.R.; Betenbaugh, M.J. Model-based analysis of N-glycosylation in Chinese hamster ovary cells. PLoS ONE 2017, 12, e0175376. [Google Scholar] [CrossRef]
- Nakajima, K.; Ito, E.; Ohtsubo, K.; Shirato, K.; Takamiya, R.; Kitazume, S.; Angata, T.; Taniguchi, N. Mass isotopomer analysis of metabolically labeled nucleotide sugars and N- and O-glycans for tracing nucleotide sugar metabolisms. Mol. Cell Proteom. 2013, 12, 2468–2480. [Google Scholar] [CrossRef]
- North, S.J.; Huang, H.H.; Sundaram, S.; Jang-Lee, J.; Etienne, A.T.; Trollope, A.; Chalabi, S.; Dell, A.; Stanley, P.; Haslam, S.M. Glycomics profiling of Chinese hamster ovary cell glycosylation mutants reveals N-glycans of a novel size and complexity. J. Biol. Chem. 2010, 285, 5759–5775. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Franc, V.; Heck, A.J.R. Glycoproteomics: A Balance between High-Throughput and In-Depth Analysis. Trends Biotechnol. 2017, 35, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Thaysen-Andersen, M.; Packer, N.H. Advances in LC-MS/MS-based glycoproteomics: Getting closer to system-wide site-specific mapping of the N- and O-glycoproteome. Biochim. Biophys. Acta 2014, 1844, 1437–1452. [Google Scholar] [CrossRef]
- Sun, B.; Hood, L. Protein-centric proteomics analysis of membrane and plasma membrane proteins. J. Proteome Res. 2014, 13, 2705–2714. [Google Scholar] [CrossRef] [PubMed]
- Lennicke, C.; Rahn, J.; Heimer, N.; Lichtenfels, R.; Wessjohann, L.A.; Seliger, B. Redox proteomics: Methods for the identification and enrichment of redox-modified proteins and their applications. Proteomics 2016, 16, 197–213. [Google Scholar] [CrossRef]
- Solda, T.; Garbi, N.; Hammerling, G.J.; Molinari, M. Consequences of ERp57 deletion on oxidative folding of obligate and facultative clients of the calnexin cycle. J. Biol. Chem. 2006, 281, 6219–6226. [Google Scholar] [CrossRef]
- Watt, G.M.; Lund, J.; Levens, M.; Kolli, V.S.; Jefferis, R.; Boons, G.J. Site-specific glycosylation of an aglycosylated human IgG1-Fc antibody protein generates neoglycoproteins with enhanced function. Chem. Biol. 2003, 10, 807–814. [Google Scholar] [CrossRef][Green Version]
- Mathys, L.; Balzarini, J. Several N-Glycans on the HIV Envelope Glycoprotein gp120 Preferentially Locate Near Disulphide Bridges and Are Required for Efficient Infectivity and Virus Transmission. PLoS ONE 2015, 10, e0130621. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakshi, T.; Pham, D.; Kaur, R.; Sun, B. Hidden Relationships between N-Glycosylation and Disulfide Bonds in Individual Proteins. Int. J. Mol. Sci. 2022, 23, 3742. https://doi.org/10.3390/ijms23073742
Bakshi T, Pham D, Kaur R, Sun B. Hidden Relationships between N-Glycosylation and Disulfide Bonds in Individual Proteins. International Journal of Molecular Sciences. 2022; 23(7):3742. https://doi.org/10.3390/ijms23073742
Chicago/Turabian StyleBakshi, Tania, David Pham, Raminderjeet Kaur, and Bingyun Sun. 2022. "Hidden Relationships between N-Glycosylation and Disulfide Bonds in Individual Proteins" International Journal of Molecular Sciences 23, no. 7: 3742. https://doi.org/10.3390/ijms23073742
APA StyleBakshi, T., Pham, D., Kaur, R., & Sun, B. (2022). Hidden Relationships between N-Glycosylation and Disulfide Bonds in Individual Proteins. International Journal of Molecular Sciences, 23(7), 3742. https://doi.org/10.3390/ijms23073742