
ELK3 Controls Gastric Cancer Cell Migration and Invasion by Regulating ECM Remodeling-Related Genes


Abstract
1. Introduction
2. Results
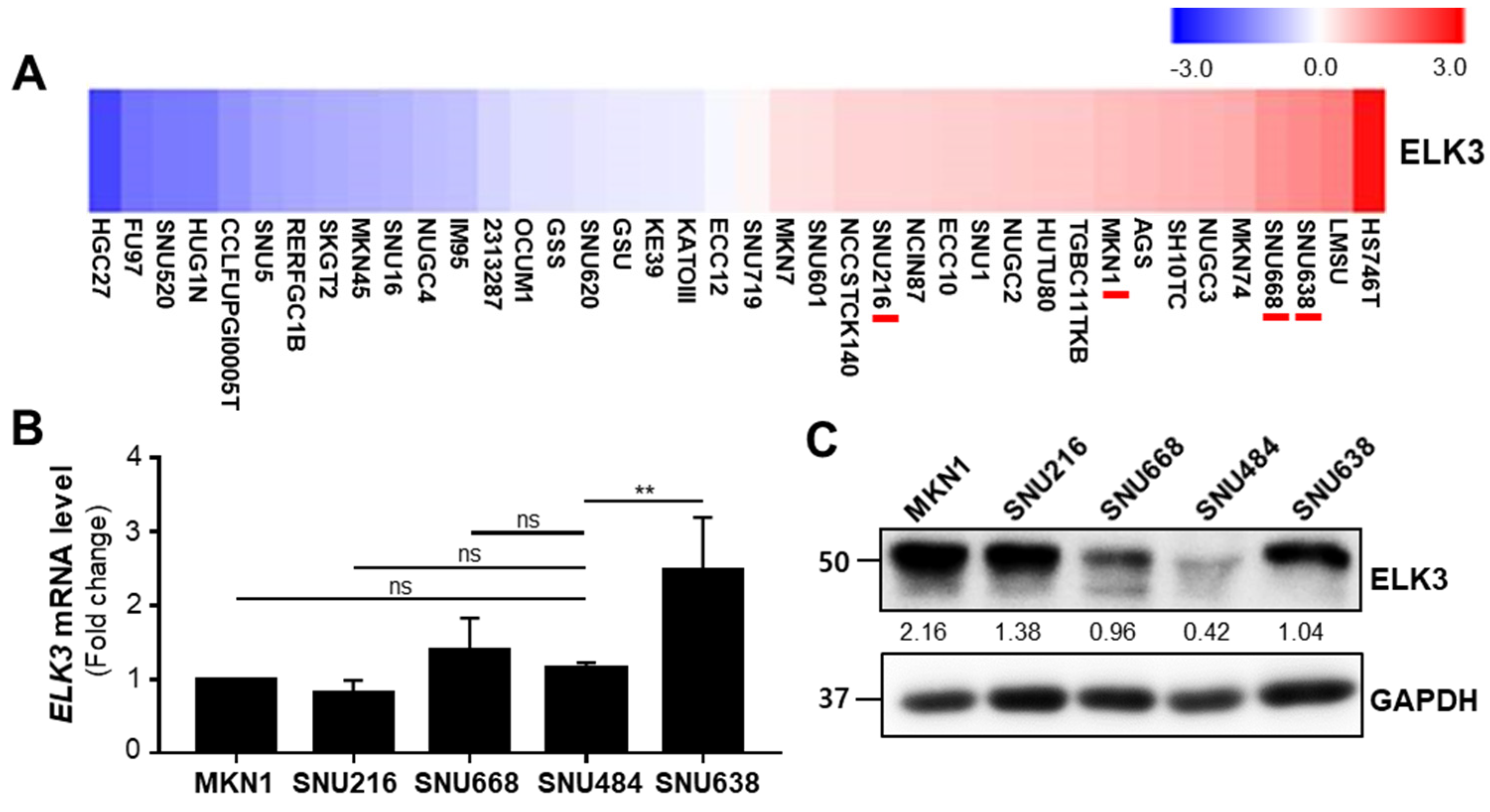
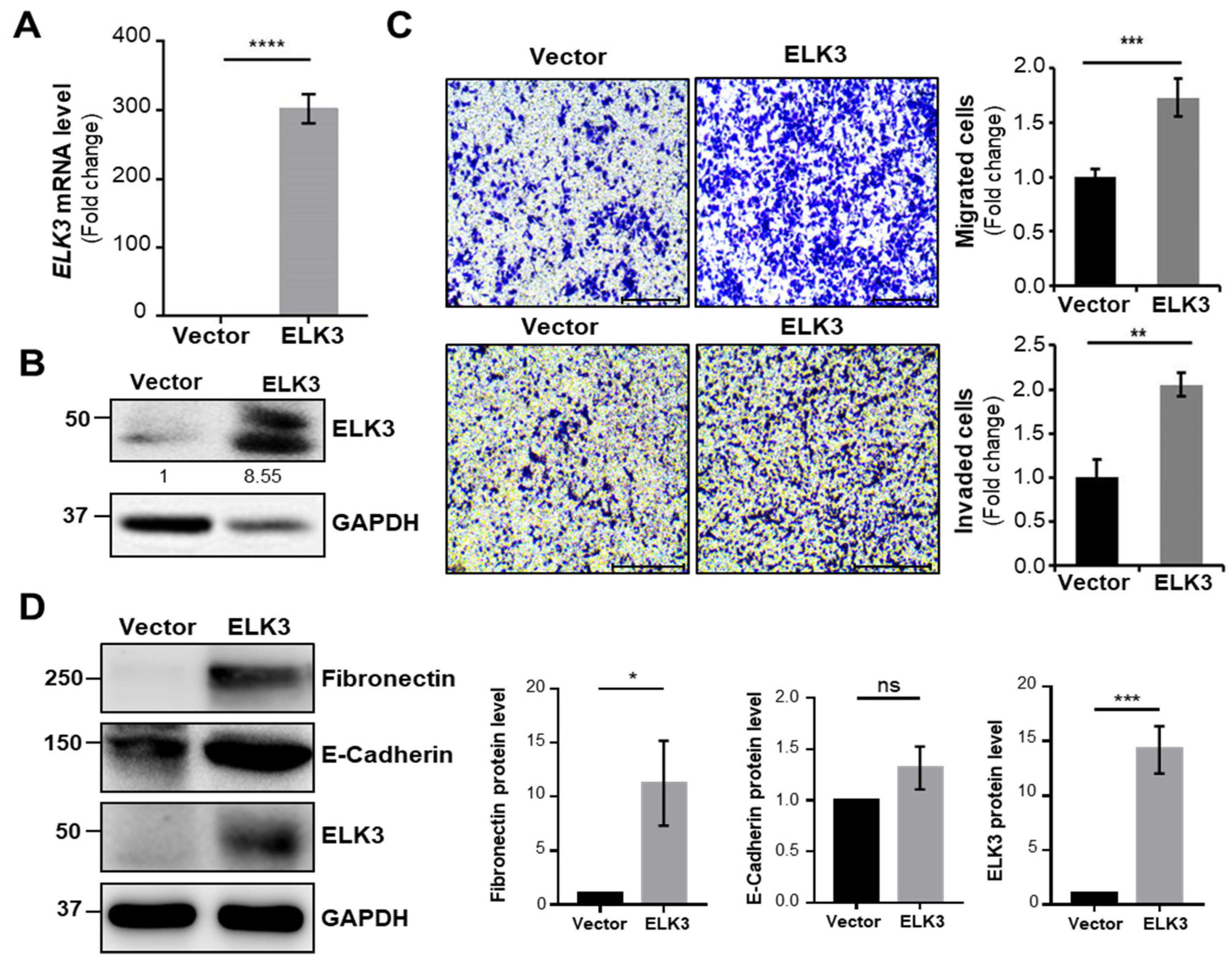
2.1. ELK3 Controls Gastric Cancer Cell Migration and Invasion in E-Cadherin Independent Manner
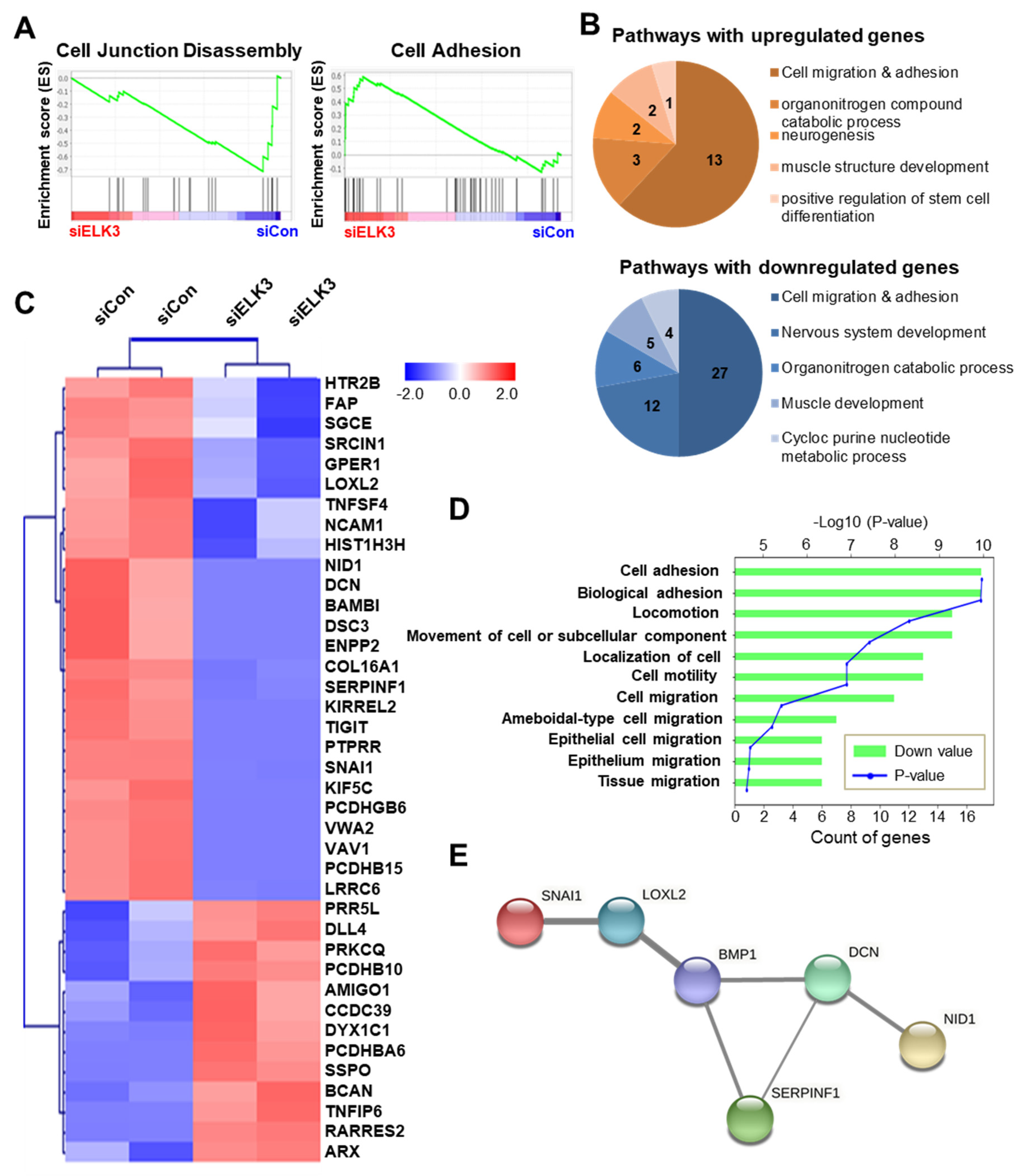
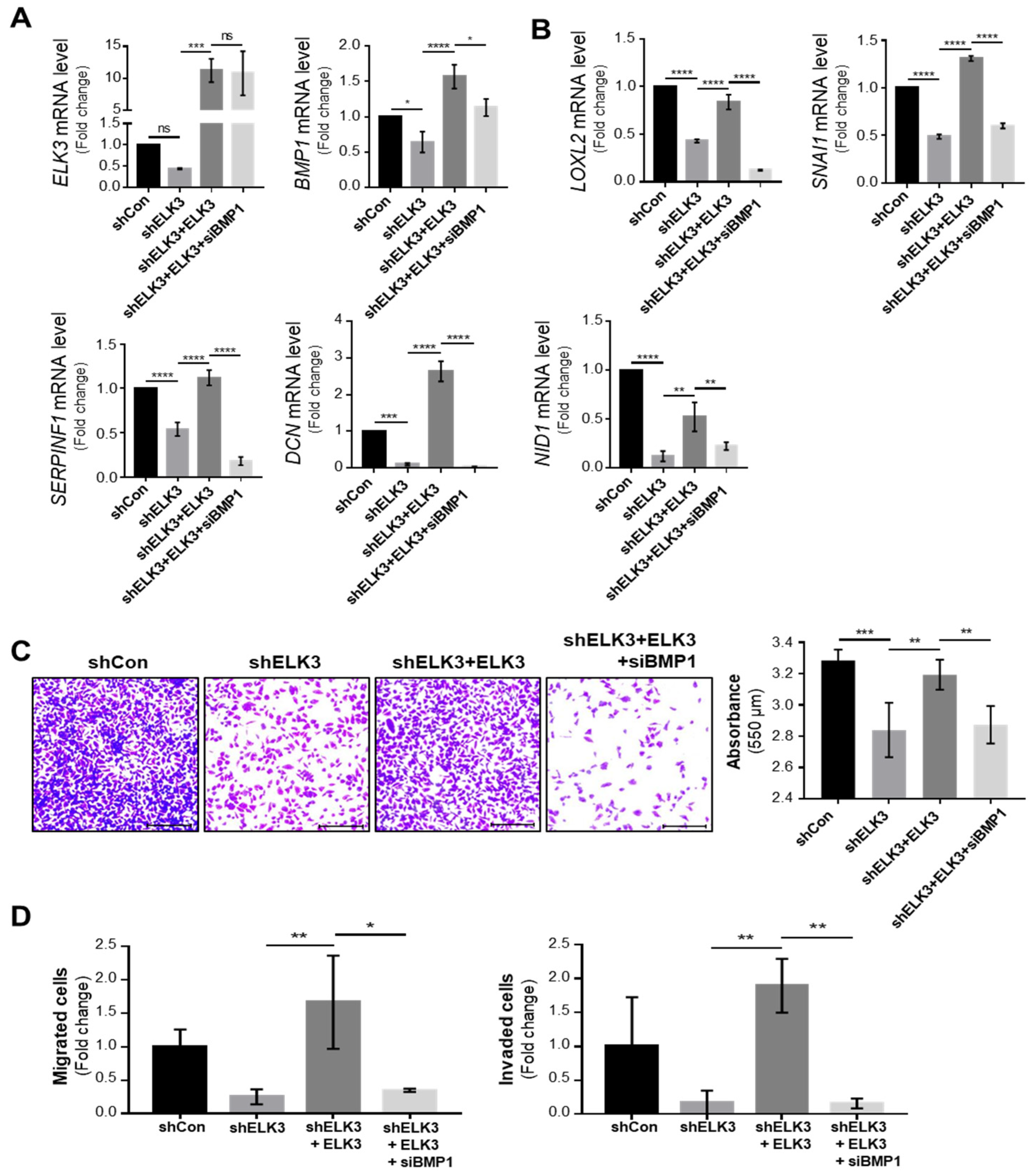
2.2. ELK3 Regulates Extracellular Matrix (ECM) Remodeling-Related Genes to Control Cell Migration and Invasion
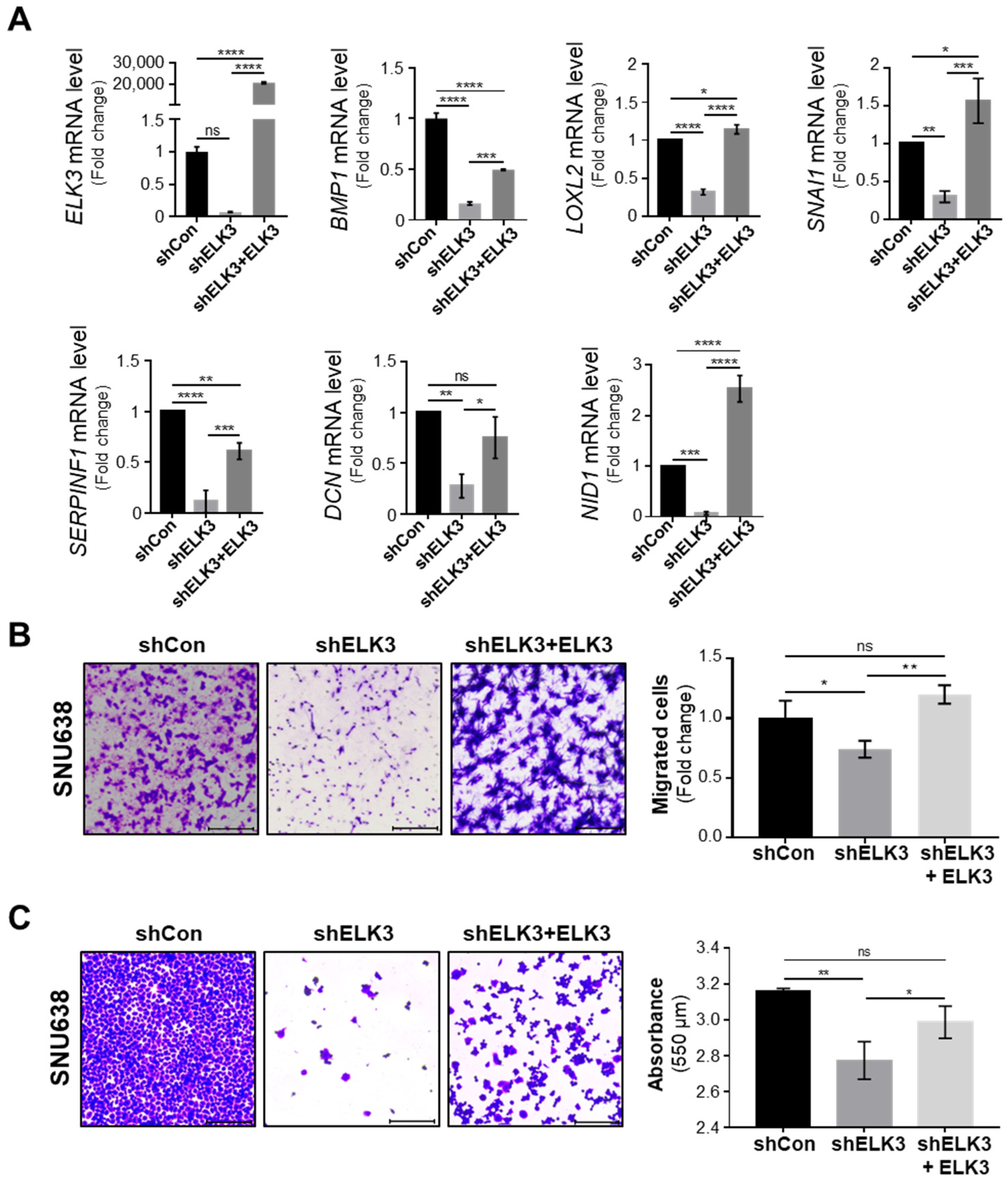
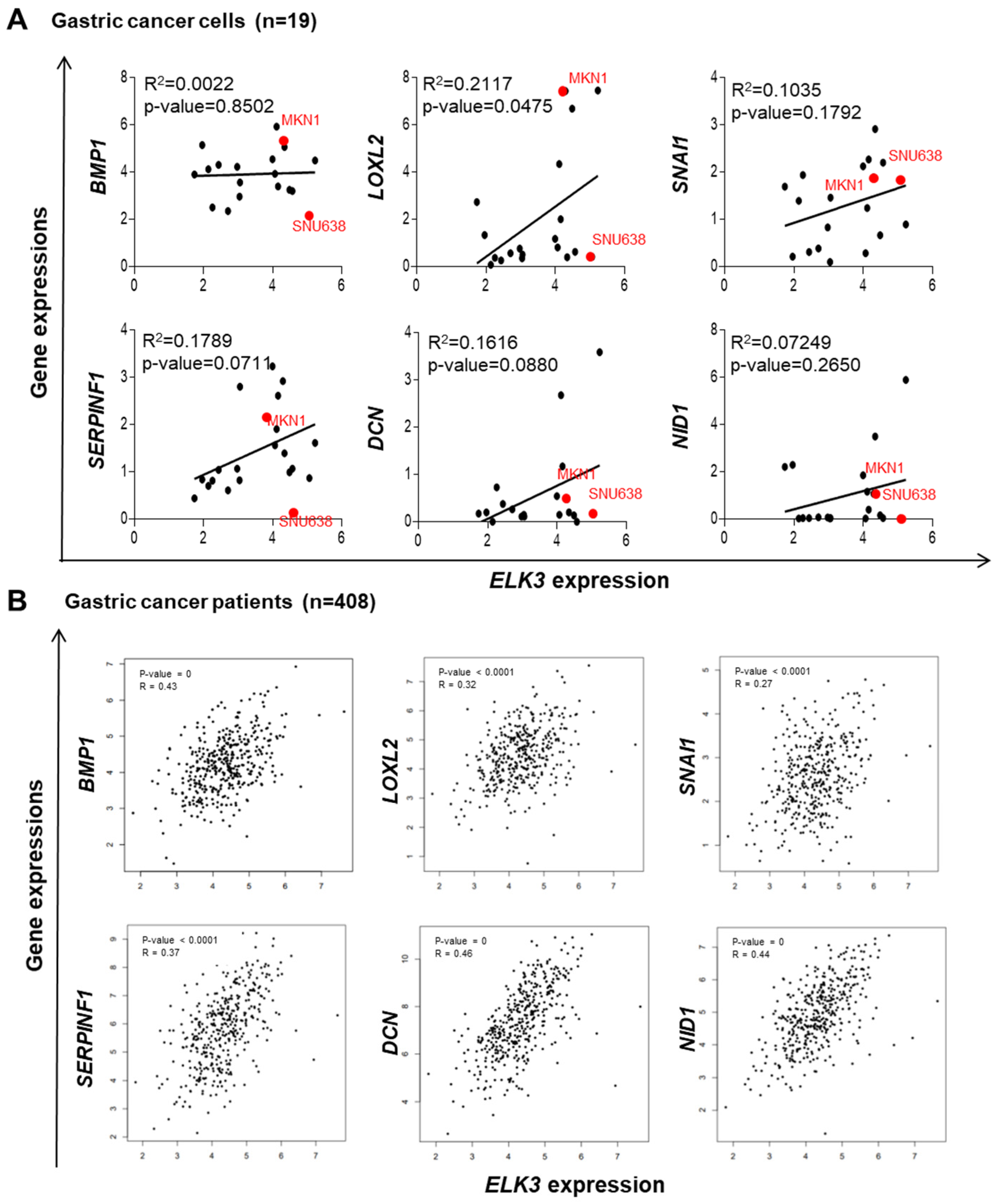
2.3. ELK3 Positively Correlates with ECM Remodeling-Related Genes in Gastric Cancer Cells and Patient Samples
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Plasmid DNA, Small Interfering (si)RNA Transfection, and Short Haripin (sh)RNA Transduction
4.3. Reverse Transcription-Quantitative PCR (RT-qPCR)
4.4. Cell Migration and Invasion Assays
4.5. Cell Adhesion Assay
4.6. Western Blot Analysis
4.7. RNA Sequencing (RNA-Seq) Analysis
4.8. Genomic Analyses of Gastric Cancer Cell Lines and Samples from Patients with Gastric Cancer
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; van Grieken, N.C.; Lordick, F. Gastric cancer. Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef]
- Karimi, P.; Islami, F.; Anandasabapathy, S.; Freedman, N.D.; Kamangar, F. Gastric cancer: Descriptive epidemiology, risk factors, screening, and prevention. Cancer Epidemiol. Biomark. Prev. 2014, 23, 700–713. [Google Scholar] [CrossRef] [PubMed]
- Cenitagoya, G.F.; Bergh, C.K.; Klinger-Roitman, J. A prospective study of gastric cancer. ‘Real’ 5-year survival rates and mortality rates in a country with high incidence. Dig. Surg. 1998, 15, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Chen, X.; Alattar, M.; Wei, J.; Liu, H. MicroRNAs in tumorigenesis, metastasis, diagnosis and prognosis of gastric cancer. Cancer Gene Ther. 2015, 22, 291–301. [Google Scholar] [CrossRef]
- Liu, H.; Xiang, Y.; Zong, Q.B.; Zhang, X.Y.; Wang, Z.W.; Fang, S.Q.; Zhang, T.C.; Liao, X.H. miR-6745-TIMP1 axis inhibits cell growth and metastasis in gastric cancer. Aging 2021, 13, 24402–24416. [Google Scholar] [CrossRef]
- Guan, H.; Li, W.; Li, Y.; Wang, J.; Li, Y.; Tang, Y.; Lu, S. MicroRNA-93 promotes proliferation and metastasis of gastric cancer via targeting TIMP2. PLoS ONE 2017, 12, e0189490. [Google Scholar] [CrossRef]
- Tian, S.; Peng, P.; Li, J.; Deng, H.; Zhan, N.; Zeng, Z.; Dong, W. SERPINH1 regulates EMT and gastric cancer metastasis via the Wnt/beta-catenin signaling pathway. Aging 2020, 12, 3574–3593. [Google Scholar] [CrossRef]
- Yang, X.Z.; Cheng, T.T.; He, Q.J.; Lei, Z.Y.; Chi, J.; Tang, Z.; Liao, Q.X.; Zhang, H.; Zeng, L.S.; Cui, S.Z. LINC01133 as ceRNA inhibits gastric cancer progression by sponging miR-106a-3p to regulate APC expression and the Wnt/beta-catenin pathway. Mol. Cancer 2018, 17, 126. [Google Scholar] [CrossRef]
- Uchihara, T.; Miyake, K.; Yonemura, A.; Komohara, Y.; Itoyama, R.; Koiwa, M.; Yasuda, T.; Arima, K.; Harada, K.; Eto, K.; et al. Extracellular Vesicles from Cancer-Associated Fibroblasts Containing Annexin A6 Induces FAK-YAP Activation by Stabilizing beta1 Integrin, Enhancing Drug Resistance. Cancer Res. 2020, 80, 3222–3235. [Google Scholar] [CrossRef]
- Abbaszadegan, M.R.; Mojarrad, M.; Moghbeli, M. Role of extra cellular proteins in gastric cancer progression and metastasis: An update. Genes Environ. 2020, 42, 18. [Google Scholar] [CrossRef]
- Miki, Y.; Yashiro, M.; Moyano-Galceran, L.; Sugimoto, A.; Ohira, M.; Lehti, K. Crosstalk Between Cancer Associated Fibroblasts and Cancer Cells in Scirrhous Type Gastric Cancer. Front. Oncol. 2020, 10, 568557. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Xing, Z.; Guo, Z.; Liu, Z. LINC01106 post-transcriptionally regulates ELK3 and HOXD8 to promote bladder cancer progression. Cell Death Dis. 2020, 11, 1063. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Park, J.D.; Choi, S.H.; Shin, D.J.; Hwang, S.; Jung, H.Y.; Park, K.S. Functional Link between miR-200a and ELK3 Regulates the Metastatic Nature of Breast Cancer. Cancers 2020, 12, 1225. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Oh, N.; Park, J.H.; Kim, K.S.; Kim, H.K.; Lee, E.; Hwang, S.; Kim, S.J.; Park, K.S. ZEB1 Collaborates with ELK3 to Repress E-Cadherin Expression in Triple-Negative Breast Cancer Cells. Mol. Cancer Res. 2019, 17, 2257–2266. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hur, W.; Hong, S.W.; Kim, J.H.; Kim, S.M.; Lee, E.B.; Yoon, S.K. ELK3 promotes the migration and invasion of liver cancer stem cells by targeting HIF-1alpha. Oncol. Rep. 2017, 37, 813–822. [Google Scholar] [CrossRef]
- Semenchenko, K.; Wasylyk, C.; Cheung, H.; Tourrette, Y.; Maas, P.; Schalken, J.A.; van der Pluijm, G.; Wasylyk, B. XRP44X, an Inhibitor of Ras/Erk Activation of the Transcription Factor Elk3, Inhibits Tumour Growth and Metastasis in Mice. PLoS ONE 2016, 11, e0159531. [Google Scholar] [CrossRef]
- Mao, Y.; Li, W.; Hua, B.; Gu, X.; Pan, W.; Chen, Q.; Xu, B.; Wang, Z.; Lu, C. Silencing of ELK3 Induces S-M Phase Arrest and Apoptosis and Upregulates SERPINE1 Expression Reducing Migration in Prostate Cancer Cells. BioMed Res. Int. 2020, 2020, 2406159. [Google Scholar] [CrossRef]
- Heo, S.H.; Cho, J.Y. ELK3 suppresses angiogenesis by inhibiting the transcriptional activity of ETS-1 on MT1-MMP. Int. J. Biol. Sci. 2014, 10, 438–447. [Google Scholar] [CrossRef][Green Version]
- Buchwalter, G.; Gross, C.; Wasylyk, B. The ternary complex factor Net regulates cell migration through inhibition of PAI-1 expression. Mol. Cell. Biol. 2005, 25, 10853–10862. [Google Scholar] [CrossRef][Green Version]
- Bridgewater, H.E.; Date, K.L.; O’Neil, J.D.; Hu, C.; Arrand, J.R.; Dawson, C.W.; Young, L.S. The Epstein-Barr Virus-Encoded EBNA1 Protein Activates the Bone Morphogenic Protein (BMP) Signalling Pathway to Promote Carcinoma Cell Migration. Pathogens 2020, 9, 594. [Google Scholar] [CrossRef]
- Rafi, J.H.; Jafar, T.; Pathan, M.T.; Reza, R.; Islam, S.; Sourna, I.J.; Alam, R.; Samad, A.; Ahammad, F. High expression of bone morphogenetic protein 1 (BMP1) is associated with a poor survival rate in human gastric cancer, a dataset approaches. Genomics 2021, 113, 1141–1154. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.Y.; Tung, S.Y.; Pan, H.Y.; Yen, C.W.; Xu, H.W.; Deng, Y.F.; Lin, Y.J.; Hsu, W.T.; Wu, C.S.; Li, C. Upregulation of bone morphogenetic protein 1 is associated with poor prognosis of late-stage gastric Cancer patients. BMC Cancer 2018, 18, 508. [Google Scholar] [CrossRef]
- Dazhi, W.; Zheng, J.; Chunling, R. High ELK3 expression is associated with the VEGF-C/VEGFR-3 axis and gastric tumorigenesis and enhances infiltration of M2 macrophages. Future Med. Chem. 2020, 12, 2209–2224. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chen, Z.; Wu, Y.; Lu, H.; Xin, L. Prognostic signature composed of transcription factors accurately predicts the prognosis of gastric cancer patients. Cancer Cell Int. 2021, 21, 357. [Google Scholar] [CrossRef]
- Heo, S.H.; Lee, J.Y.; Yang, K.M.; Park, K.S. ELK3 Expression Correlates With Cell Migration, Invasion, and Membrane Type 1-Matrix Metalloproteinase Expression in MDA-MB-231 Breast Cancer Cells. Gene Expr. 2015, 16, 197–203. [Google Scholar] [CrossRef]
- Kim, K.S.; Kim, J.; Oh, N.; Kim, M.Y.; Park, K.S. ELK3-GATA3 axis modulates MDA-MB-231 metastasis by regulating cell-cell adhesion-related genes. Biochem. Biophys. Res. Commun. 2018, 498, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The extracellular matrix: Tools and insights for the “omics” era. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef]
- Hastings, J.F.; Skhinas, J.N.; Fey, D.; Croucher, D.R.; Cox, T.R. The extracellular matrix as a key regulator of intracellular signalling networks. Br. J. Pharmacol. 2019, 176, 82–92. [Google Scholar] [CrossRef]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nat. Rev. Mol. Cell Biol. 2014, 15, 802–812. [Google Scholar] [CrossRef]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef]
- Xiao, Q.; Ge, G. Lysyl oxidase, extracellular matrix remodeling and cancer metastasis. Cancer Microenviron. 2012, 5, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Salvador, F.; Martin, A.; Lopez-Menendez, C.; Moreno-Bueno, G.; Santos, V.; Vazquez-Naharro, A.; Santamaria, P.G.; Morales, S.; Dubus, P.R.; Muinelo-Romay, L.; et al. Lysyl Oxidase-like Protein LOXL2 Promotes Lung Metastasis of Breast Cancer. Cancer Res. 2017, 77, 5846–5859. [Google Scholar] [CrossRef] [PubMed]
- Peinado, H.; Del Carmen Iglesias-de la Cruz, M.; Olmeda, D.; Csiszar, K.; Fong, K.S.; Vega, S.; Nieto, M.A.; Cano, A.; Portillo, F. A molecular role for lysyl oxidase-like 2 enzyme in snail regulation and tumor progression. EMBO J. 2005, 24, 3446–3458. [Google Scholar] [CrossRef] [PubMed]
- Aleckovic, M.; Wei, Y.; LeRoy, G.; Sidoli, S.; Liu, D.D.; Garcia, B.A.; Kang, Y. Identification of Nidogen 1 as a lung metastasis protein through secretome analysis. Genes Dev. 2017, 31, 1439–1455. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequence |
|---|---|
| ELK3 | F: 5′-ACCCAAAGGCTTGGAAATCT-3′ |
| R: 5′-TGTATGCTGGAGAGCAGTGG-3′ | |
| BMP1 | F: 5′-TGGCCGACTACACCTATGAC-3′ |
| R: 5′-GGAGGACTTACGAGCTGTGT-3′ | |
| LOXL2 | F: 5′-GGACATGTACCGCCATGACA-3′ |
| R: 5′-ATAGCGGCTCCTGCATTTCA-3′ | |
| SNAI1 | F: 5′-GCAAATACTGCAACAAGG-3′ |
| R: 5′-GCACTGGTACTTCTTGACA-3′ | |
| SERPINF1 | F: 5′-GCTATGACCTGTACCGGGTG-3′ |
| R: 5′-GTCTGGGCTGCTGATCAAGT-3′ | |
| DCN | F: 5′-AATTGAAAATGGGGCTTTCC-3′ |
| R: 5′-GCCATTGTCAACAGCAGAGA-3′ | |
| NID1 | F: 5′-CCTGCTACATCGGCACTCAT-3′ |
| R: 5′-TGAAAACTGGTAGCCCTCCAC-3′ | |
| GAPDH | F: 5′-GGGTGTGAACCATGAGAA-3′ |
| R: 5′-GTCTTCTGGGTGGCAGTGAT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.; Cho, H.-J.; Park, K.-S.; Jung, H.-Y. ELK3 Controls Gastric Cancer Cell Migration and Invasion by Regulating ECM Remodeling-Related Genes. Int. J. Mol. Sci. 2022, 23, 3709. https://doi.org/10.3390/ijms23073709
Lee M, Cho H-J, Park K-S, Jung H-Y. ELK3 Controls Gastric Cancer Cell Migration and Invasion by Regulating ECM Remodeling-Related Genes. International Journal of Molecular Sciences. 2022; 23(7):3709. https://doi.org/10.3390/ijms23073709
Chicago/Turabian StyleLee, Minwook, Hyeon-Ju Cho, Kyung-Soon Park, and Hae-Yun Jung. 2022. "ELK3 Controls Gastric Cancer Cell Migration and Invasion by Regulating ECM Remodeling-Related Genes" International Journal of Molecular Sciences 23, no. 7: 3709. https://doi.org/10.3390/ijms23073709
APA StyleLee, M., Cho, H.-J., Park, K.-S., & Jung, H.-Y. (2022). ELK3 Controls Gastric Cancer Cell Migration and Invasion by Regulating ECM Remodeling-Related Genes. International Journal of Molecular Sciences, 23(7), 3709. https://doi.org/10.3390/ijms23073709