
Systematic Analysis and Expression Profiles of the 4-Coumarate: CoA Ligase (4CL) Gene Family in Pomegranate (Punica granatum L.)



Abstract
:1. Introduction
2. Results
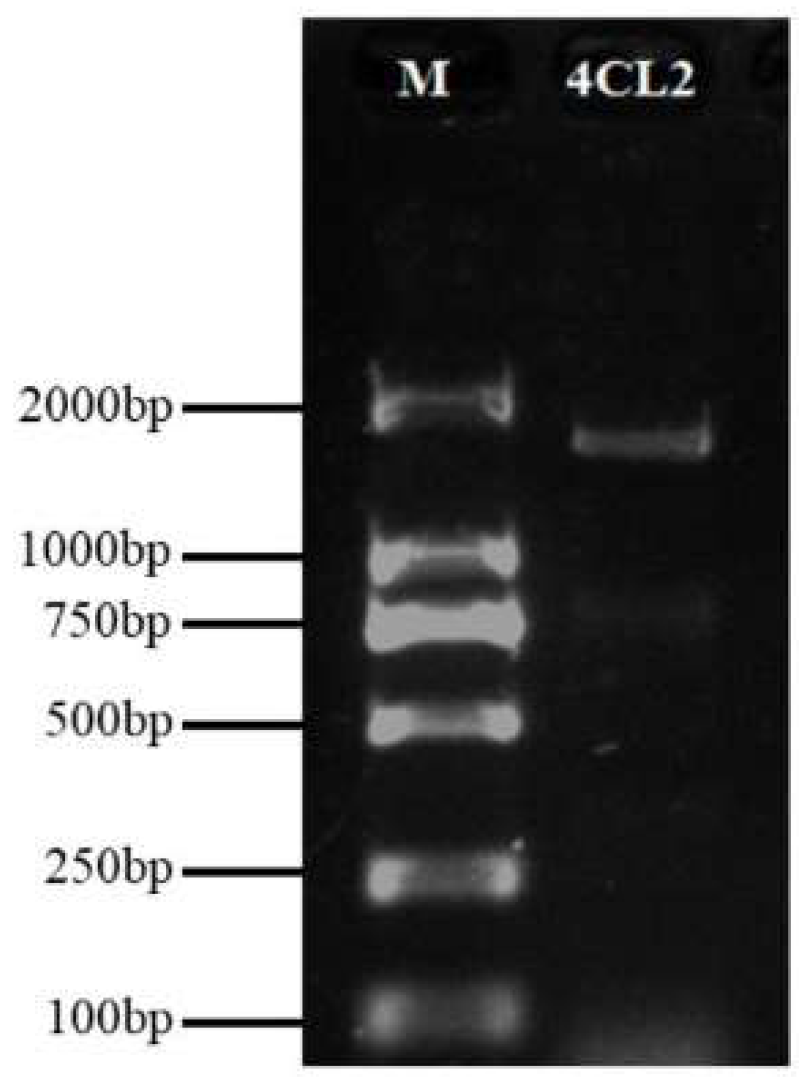
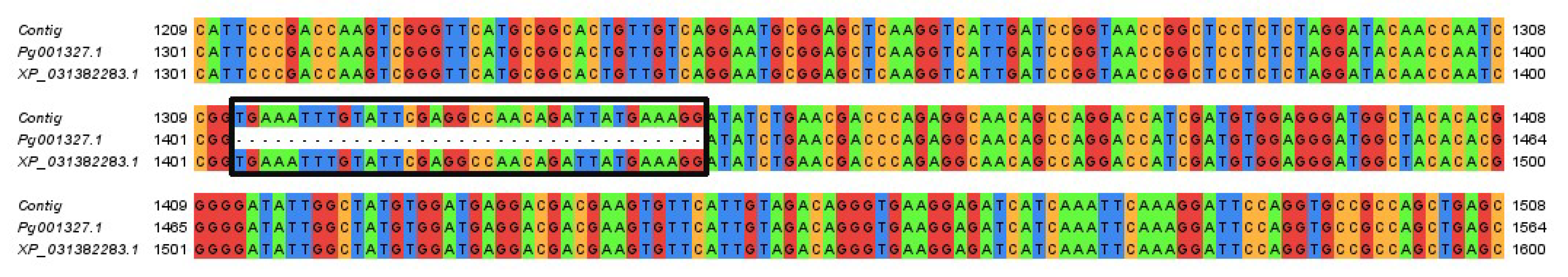
2.1. Identification of Pg4CL Family Genes and Gene Cloning of Pg001327.1
2.2. Analysis of Physical and Chemical Properties
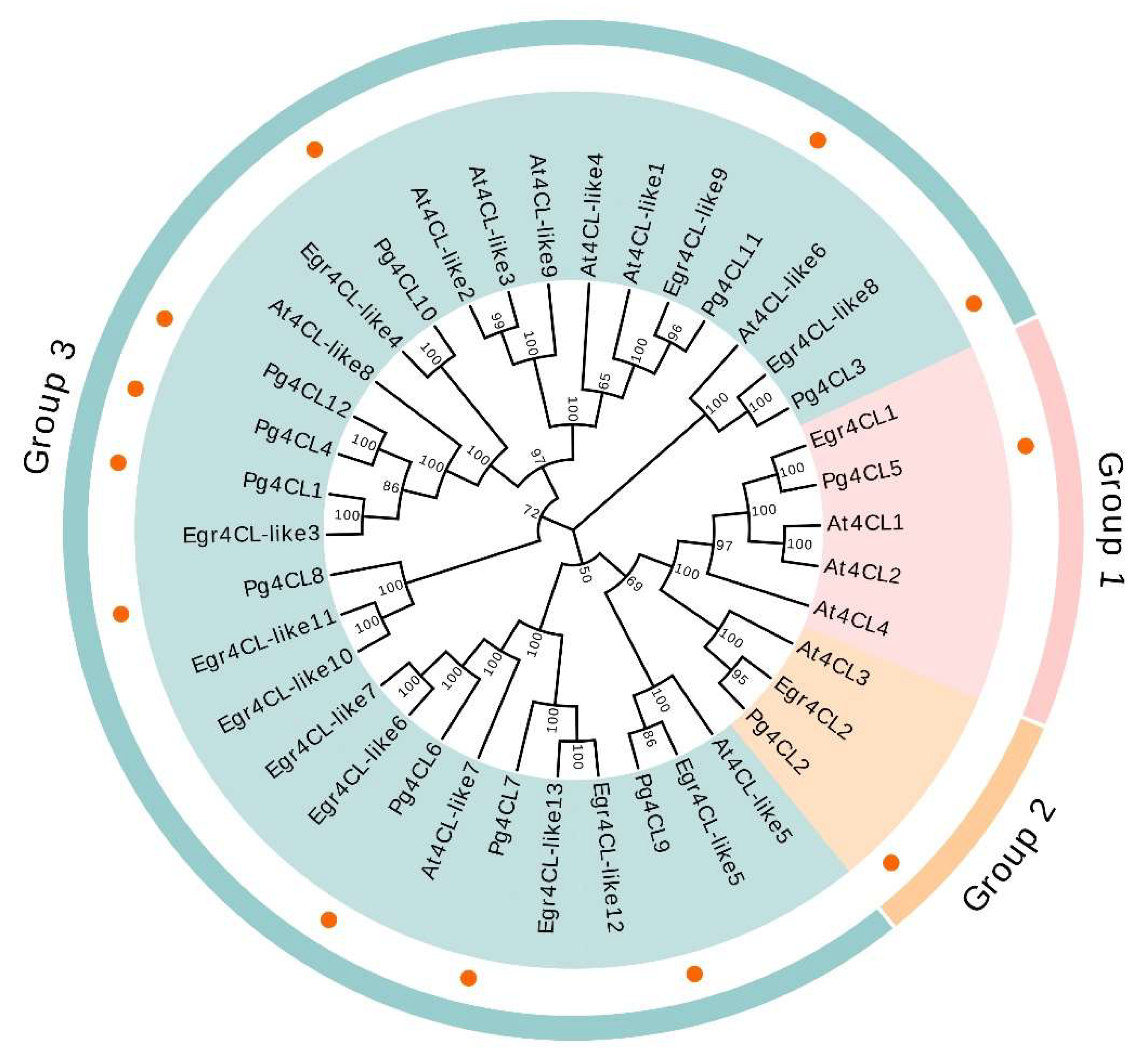
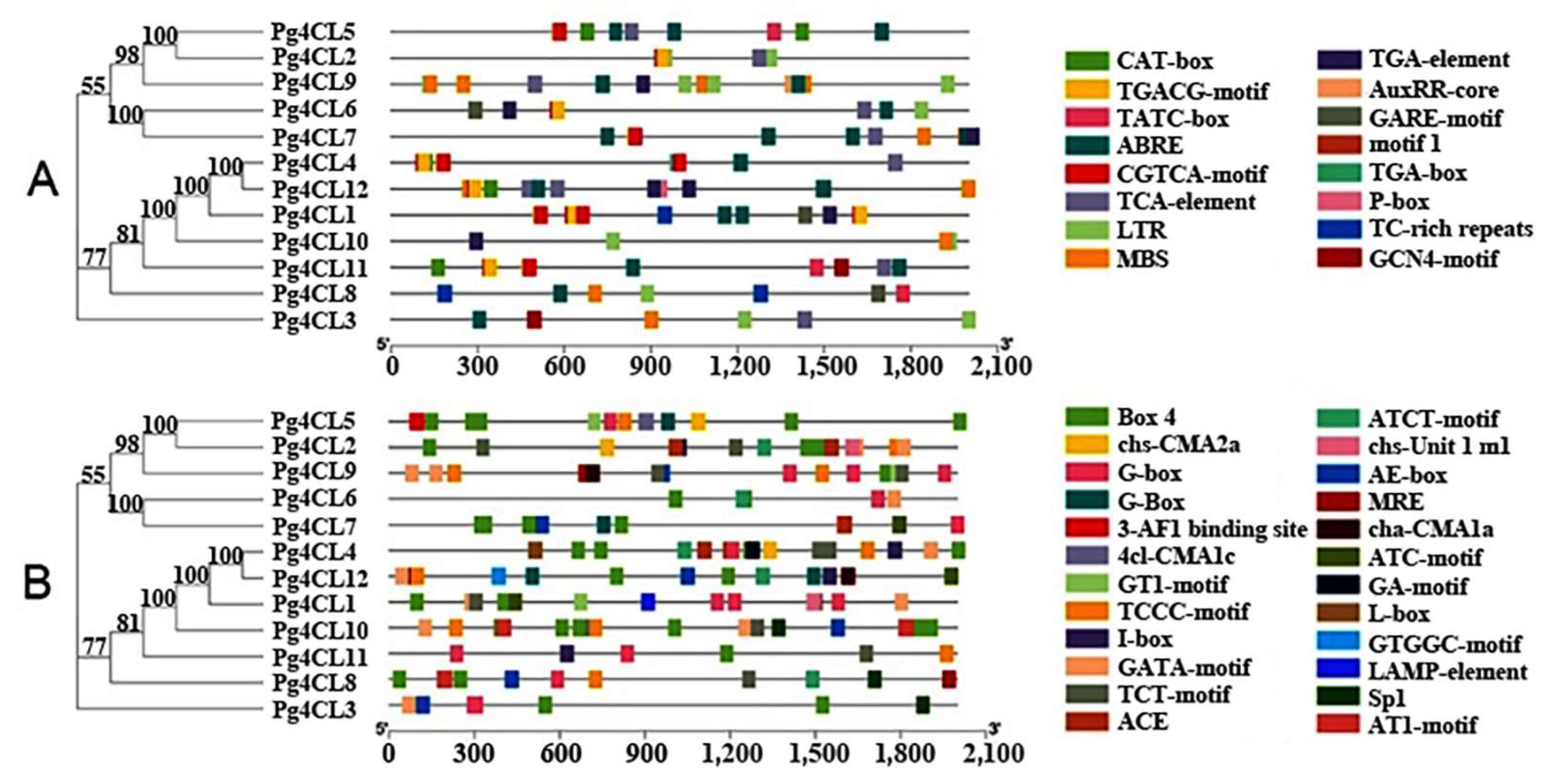
2.3. Phylogenetic Analysis and Classification of the Pg4CL Gene Family
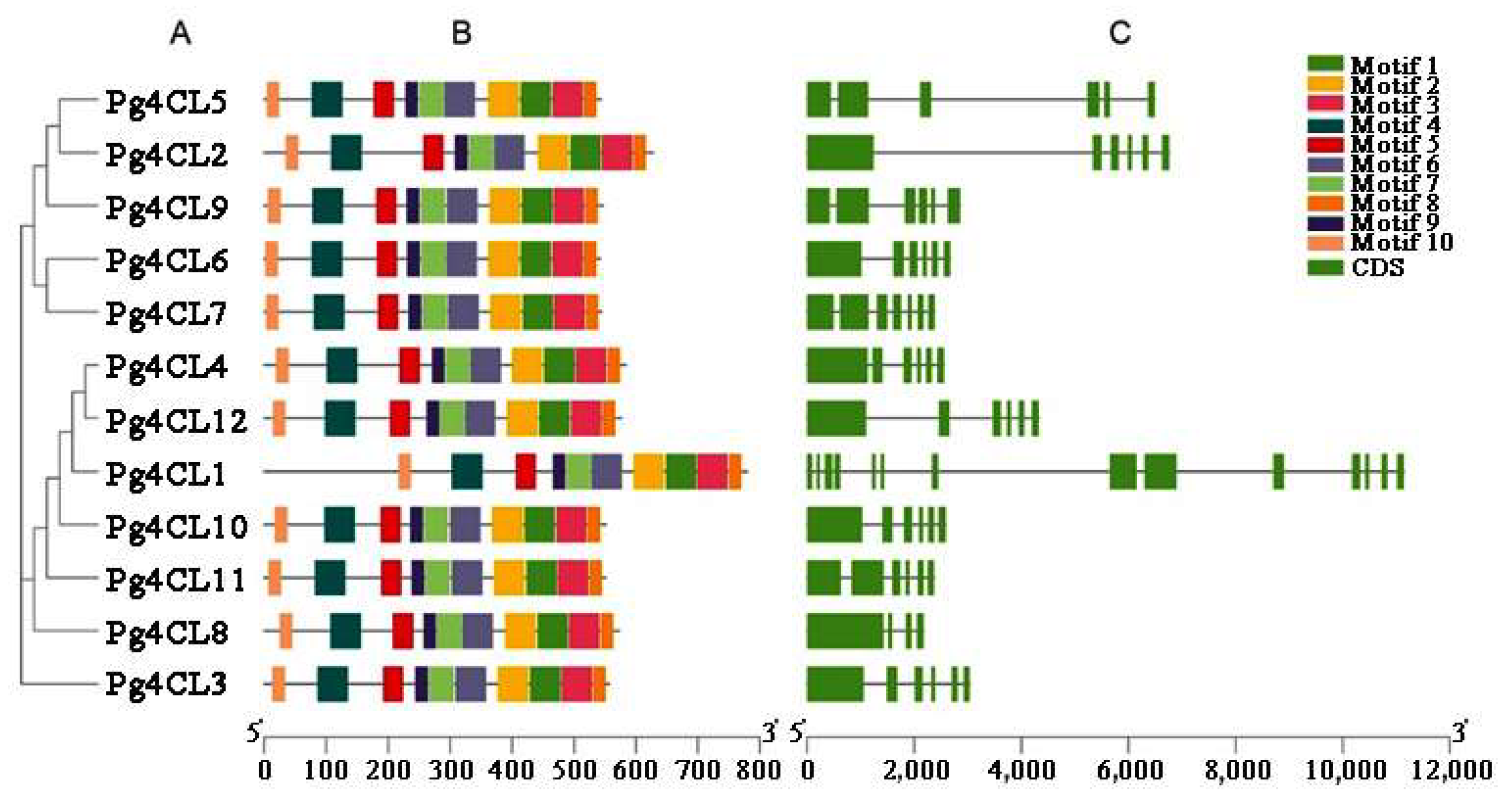
2.4. Analysis of Conserved Motifs and Gene Structures of Pg4CL Gene Family
2.5. Cis-Acting Elements in the Promoter Region of Pg4CLs
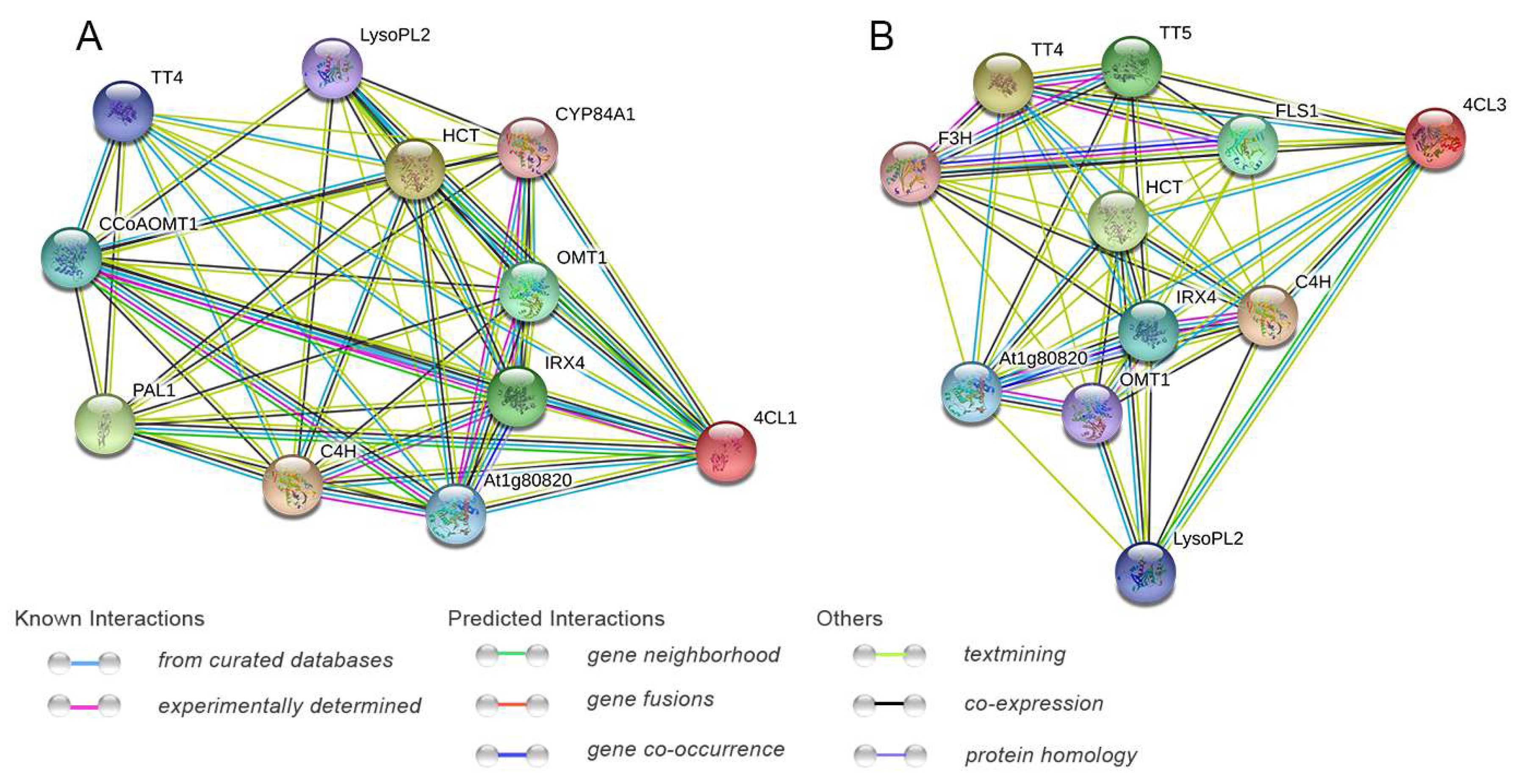
2.6. Protein Interaction Networks of Pg4CL Gene Family
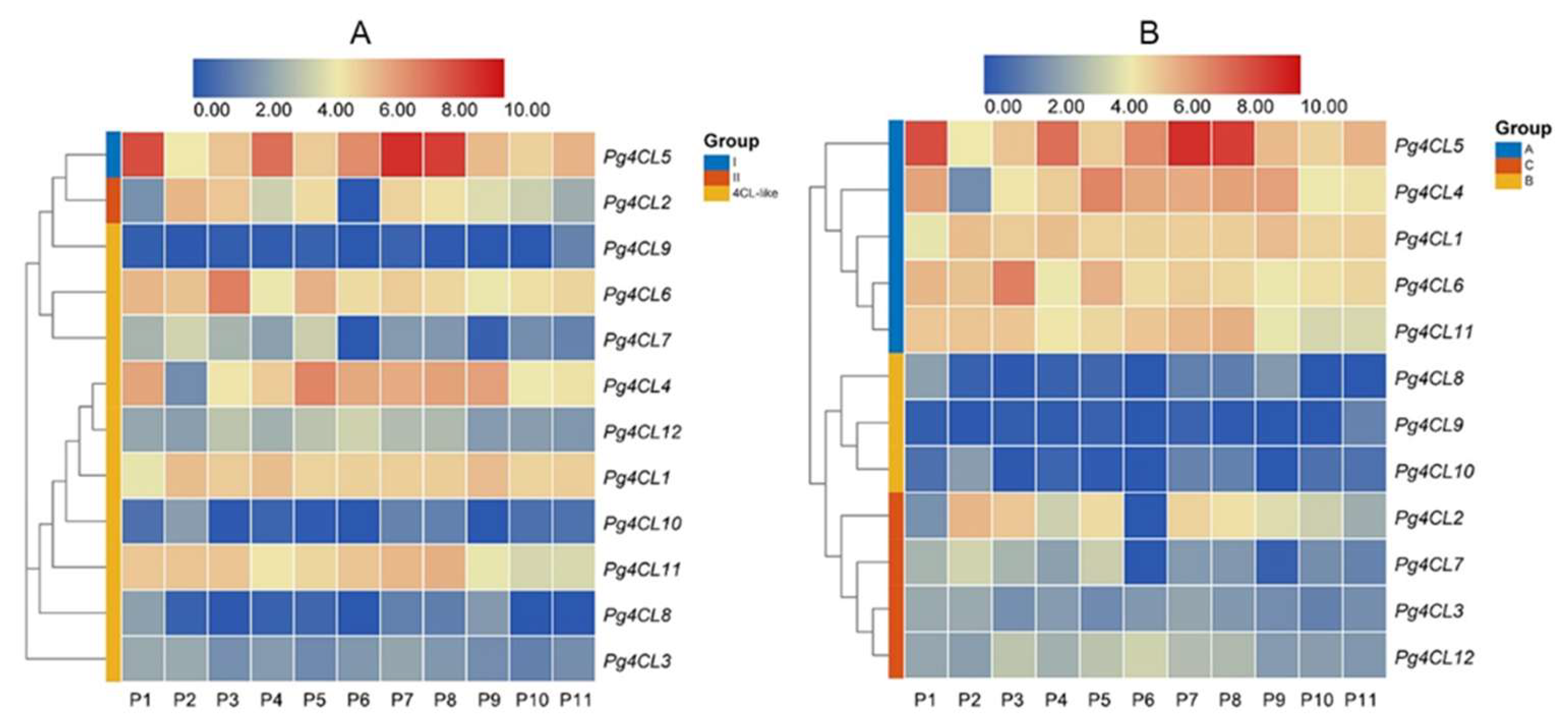
2.7. Expression Analysis of Pg4CL Genes with RNA-Seq
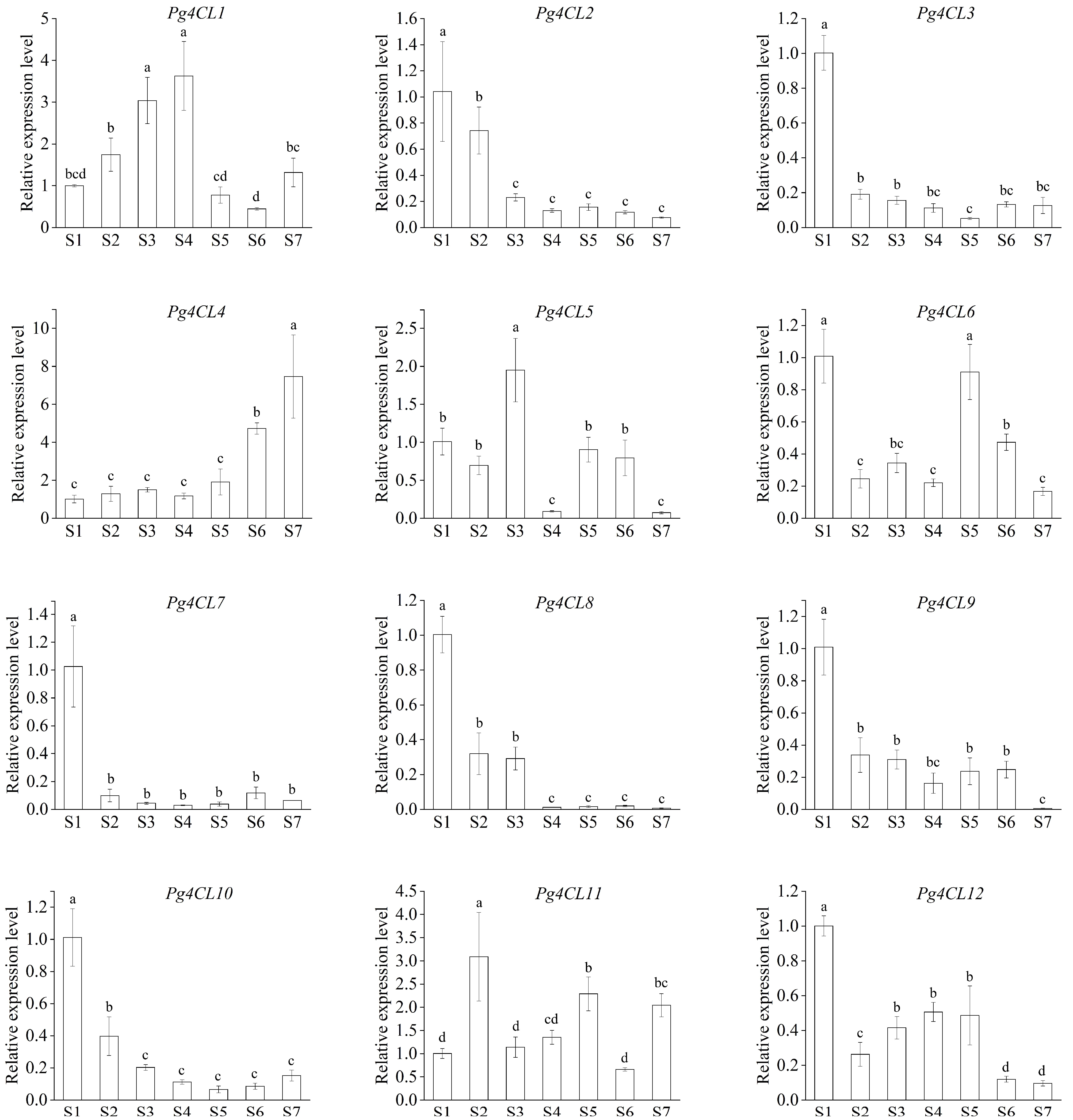
2.8. qRT-PCR Analysis of Pg4CLs during Fruit Development in Pomegranate
3. Discussion
4. Materials and Methods
4.1. Genome and Transcriptome Data Sources
4.2. Identification of Pg4CL Family Genes
4.3. Gene Cloning of Pg001327.1
4.4. Virtual Prediction of Physical and Chemical Properties
4.5. Multiple Sequences Alignment and Phylogenetic Analysis
4.6. Analysis of Gene Structure and Protein Conserved Motifs
4.7. Analysis of Cis-Acting Elements and Protein–Protein Interaction Networks
4.8. RNA-Seq Analysis of 4CL Gene Family in Pomegranate
4.9. Expression Patterns by Quantitative Real-Time PCR (qRT-PCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, Y.; Qiao, L.; Shen, Y.; Jiang, P.; Chen, J.; Ye, X. Phytochemical Profile and Antioxidant Activity of Physiological Drop of Citrus Fruits. J. Food Sci. 2013, 78, C37–C42. [Google Scholar] [CrossRef] [PubMed]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Li, Y.; Kim, J.I.; Pysh, L.; Chapple, C. Four isoforms of Arabidopsis thaliana 4-coumarate: CoA ligase (4CL) have overlapping yet distinct roles in phenylpropanoid metabolism. Plant Physiol. 2015, 169, 2409–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamberger, B.; Hahlbrock, K. The 4-coumarate: CoA ligase gene family in Arabidopsis thaliana comprises one rare, sinapate-activating and three commonly occurring isoenzymes. Proc. Natl. Acad. Sci. USA 2004, 101, 2209–2214. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Li, M.; Li, D.; Zhang, J.; Jin, Q.; Sheng, L.; Cai, Y.; Lin, Y. Correction: Characterization and analysis of CCR and CAD gene families at the whole-genome level for lignin synthesis of stone cells in pear (Pyrus bretschneideri) fruit. Biol. Open 2020, 9, 9. [Google Scholar] [CrossRef]
- Dixon, R.A.; Steele, C.L. Flavonoids and isoflavonoids—A gold mine for metabolic engineering. Trends Plant Sci. 1999, 4, 394–400. [Google Scholar] [CrossRef]
- Raes, J.; Rohde, A.; Christensen, J.H.; Van de Peer, Y.; Boerjan, W. Genome-Wide Characterization of the Lignification Toolbox in Arabidopsis. Plant Physiol. 2003, 133, 1051–1071. [Google Scholar] [CrossRef] [Green Version]
- Gui, J.; Shen, J.; Li, L. Functional Characterization of Evolutionarily Divergent 4-Coumarate:Coenzyme A Ligases in Rice. Plant Physiol. 2011, 157, 574–586. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.-C.; Xiong, X.-P.; Zhang, X.-L.; Feng, H.-J.; Zhu, Q.-H.; Sun, J.; Li, Y.-J. Characterization of the Gh4CL gene family reveals a role of Gh4CL7 in drought tolerance. BMC Plant Biol. 2020, 20, 125. [Google Scholar] [CrossRef]
- Carocha, V.; Soler, M.; Hefer, C.; Cassan-Wang, H.; Fevereiro, P.; Myburg, A.A.; Paiva, J.A.; Grima-Pettenati, J. Genome-wide analysis of the lignin toolbox of Eucalyptus grandis. New Phytol. 2015, 206, 1297–1313. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Li, X.; Jiang, L. Integrative Analysis of the Core Fruit Lignification Toolbox in Pear Reveals Targets for Fruit Quality Bioengineering. Biomology 2019, 9, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, K.; Hövel, K.; Witzel, K.; Hamberger, B.; Schomburg, D.; Kombrink, E.; Stuible, H.-P. The substrate specificity-determining amino acid code of 4-coumarate: CoA ligase. Proc. Natl. Acad. Sci. USA 2003, 100, 8601–8606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavhale, S.G.; Kalunke, R.M.; Giri, A.P. Structural, functional and evolutionary diversity of 4-coumarate-CoA ligase in plants. Planta 2018, 248, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Hamberger, B.; Ellis, M.; Friedmann, M.; de Azevedo Souza, C.; Barbazuk, B.; Douglas, C.J. Genome-wide analyses of phe-nylpropanoid-related genes in Populus trichocarpa, Arabidopsis thaliana, and Oryza sativa: The Populus lignin toolbox and con-servation and diversification of angiosperm gene families. Botany 2007, 85, 1182–1201. [Google Scholar]
- Cukovica, D.; Ehlting, J.; Van Ziffle, J.A.; Douglas, C.J. Structure and Evolution of 4-Coumarate:Coenzyme A Ligase (4CL) Gene Families. Biol. Chem. 2001, 382, 76. [Google Scholar] [CrossRef]
- Wang, B.; Sun, W.; Li, Q.; Li, Y.; Luo, H.; Song, J.; Sun, C.; Qian, J.; Zhu, Y.; Hayward, A.; et al. Genome-wide identification of phenolic acid biosynthetic genes in Salvia miltiorrhiza. Planta 2014, 241, 711–725. [Google Scholar] [CrossRef]
- Chen, X.; Wang, H.; Li, X.; Ma, K.; Zhan, Y.; Zeng, F. Molecular cloning and functional analysis of 4-Coumarate: CoA ligase 4 (4CL-like 1) from Fraxinus mandshurica and its role in abiotic stress tolerance and cell wall synthesis. BMC Plant Biol. 2019, 19, 231. [Google Scholar] [CrossRef] [Green Version]
- Qin, G.; Liu, C.; Li, J.; Qi, Y.; Gao, Z.; Zhang, X.; Yi, X.; Pan, H.; Ming, R.; Xu, Y. Diversity of metabolite accumulation patterns in inner and outer seed coats of pomegranate: Exploring their relationship with genetic mechanisms of seed coat development. Hortic. Res. 2020, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Saminathan, T.; Bodunrin, A.; Singh, N.V.; Devarajan, R.; Nimmakayala, P.; Jeff, M.; Aradhya, M.; Reddy, U.K. Genome-wide identification of microRNAs in pomegranate (Punica granatum L.) by high-throughput sequencing. BMC Plant Biol. 2016, 16, 122. [Google Scholar] [CrossRef] [Green Version]
- Pirzadeh, M.; Caporaso, N.; Rauf, A.; Shariati, M.A.; Yessimbekov, Z.; Khan, M.U.; Imran, M.; Mubarak, M.S. Pomegranate as a source of bioactive constituents: A review on their characterization, properties and applications. Crit. Rev. Food Sci. Nutr. 2021, 61, 982–999. [Google Scholar] [CrossRef]
- Patel, C.; Dadhaniya, P.; Hingorani, L.; Soni, M. Safety assessment of pomegranate fruit extract: Acute and subchronic toxicity studies. Food Chem. Toxicol. 2008, 46, 2728–2735. [Google Scholar] [CrossRef] [PubMed]
- Kandylis, P.; Kokkinomagoulos, E. Food Applications and Potential Health Benefits of Pomegranate and its Derivatives. Foods 2020, 9, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhtar, S.; Ismail, T.; Fraternale, D.; Sestili, P. Pomegranate peel and peel extracts: Chemistry and food features. Food Chem. 2015, 174, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Fang, Y.; Zhang, T.; Fei, Z.; Han, F.; Liu, C.; Liu, M.; Xiao, W.; Zhang, W.; Wu, S.; et al. The pomegranate (Punica granatum L.) genome provides insights into fruit quality and ovule developmental biology. Plant Biotechnol. J. 2017, 16, 1363–1374. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Li, H.; Wu, Z.; Yao, W.; Zhao, P.; Cao, D.; Yu, H.; Li, K.; Poudel, K.; Zhao, D. The pomegranate (Punica granatum L.) draft genome dissects genetic divergence between soft-and hard-seeded cultivars. Plant Biotechnol. J. 2020, 18, 955–968. [Google Scholar] [CrossRef] [Green Version]
- Qin, G.; Xu, C.; Ming, R.; Tang, H.; Guyot, R.; Kramer, E.M.; Hu, Y.; Yi, X.; Qi, Y.; Xu, X.; et al. The pomegranate (Punica granatum L.) genome and the genomics of punicalagin biosynthesis. Plant J. 2017, 91, 1108–1128. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.-H.; Ma, T.; Luo, W.-C.; Xu, J.-M.; Liu, J.-Q.; Wan, D.-S. Identification of 4CL Genes in Desert Poplars and Their Changes in Expression in Response to Salt Stress. Genes 2015, 6, 901–917. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Geng, Z.; Zhang, C.; Wang, K.; Jiang, X. Whole-genome characterization of Rosa chinensis AP2/ERF transcription factors and analysis of negative regulator RcDREB2B in Arabidopsis. BMC Genom. 2021, 22, 90. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis Transcription Factors: Genome-Wide Comparative Analysis Among Eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef]
- Cao, Y.; Han, Y.; Li, D.; Lin, Y.; Cai, Y. Systematic Analysis of the 4-Coumarate:Coenzyme A Ligase (4CL) Related Genes and Expression Profiling during Fruit Development in the Chinese Pear. Genes 2016, 7, 89. [Google Scholar] [CrossRef] [Green Version]
- Hong, P.; Xu, X.; Wang, J.; Chen, X.; Shen, X.; Lin, Y.; Lai, Z. Identification and functional analysis of 4CL gene family during early somatic embryogenesis in Dimocarpus longan Lour. Chin. J. Trop. Crops 2021, 42, 909–919. (In Chinese) [Google Scholar]
- Dong, L.; Abdurazak, I.; Hu, Q.; Li, C.; Zhang, S. Cloning and Expression Analysis of Pomegranate 4-coumarate-CoA Ligase Pg4CL. Acta Bot. Sin. 2016, 36, 2146–2151. (In Chinese) [Google Scholar]
- Luo, X.; Cao, D.; Li, H.; Zhao, D.; Xue, H.; Niu, J.; Chen, L.; Zhang, F.; Cao, S. Complementary iTRAQ-based proteomic and RNA sequencing-based transcriptomic analyses reveal a complex network regulating pomegranate (Punica granatum L.) fruit peel colour. Sci. Rep.-UK 2018, 8, 12362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldi, P.; Moser, M.; Brilli, M.; Pindo, M.; Si-Ammour, A.; Vrhovsek, U. Fine-tuning of the flavonoid and monolignol pathways during apple early fruit development. Planta 2017, 245, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Yu, J.; Cai, Y.-X.; Zhu, P.-P.; Liu, C.-Y.; Zhao, A.-C.; Lü, R.-H.; Li, M.-J.; Xu, F.-X.; Yu, M.-D. Characterization and functional analysis of 4-coumarate: CoA ligase genes in mulberry. PLoS ONE 2016, 11, e0155814. [Google Scholar]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. A simple method to control over-alignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.-C.; Shen, H.-B. Plant-mPLoc: A Top-Down Strategy to Augment the Power for Predicting Plant Protein Subcellular Localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.-H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, w202–w208. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Ono, N.N.; Britton, M.T.; Fass, J.N.; Nicolet, C.M.; Lin, D.; Tian, L. Exploring the Transcriptome Landscape of Pomegranate Fruit Peel for Natural Product Biosynthetic Gene and SSR Marker Discovery F. J. Integr. Plant Biol. 2011, 53, 800–813. [Google Scholar] [CrossRef] [PubMed]
- Ophir, R.; Sherman, A.; Rubinstein, M.; Eshed, R.; Schwager, M.S.; Harel-Beja, R.; Bar-Ya’Akov, I.; Holland, D. Single-Nucleotide Polymorphism Markers from De-Novo Assembly of the Pomegranate Transcriptome Reveal Germplasm Genetic Diversity. PLoS ONE 2014, 9, e88998. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Location | CDS (bp) | Exon No. | AA | pI | Instability Index | GRAVY | Signal Peptide | Protein Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| Pg4CL1 | Pg000024.1 | scaffold1:6623488:6634629 | 2343 | 14 | 780 | 8.81 | 47.22 | 0.059 | No | Plasma membrane |
| Pg4CL2 | Pg001327.1 | scaffold10:76276:83034 | 1887 | 6 | 628 | 5.83 | 46.64 | 0.066 | No | Cytosol |
| Pg4CL3 | Pg004702.1 | scaffold124:253916:256949 | 1674 | 6 | 557 | 6.55 | 39.93 | 0.159 | No | Plasma membrane |
| Pg4CL4 | Pg005072.1 | scaffold13:4695163:4697720 | 1755 | 6 | 584 | 8.09 | 49.09 | −0.057 | No | Extracellular |
| Pg4CL5 | Pg007092.1 | scaffold15:2592574:2599061 | 1635 | 6 | 544 | 5.65 | 36.60 | −0.034 | No | Extracellular |
| Pg4CL6 | Pg007674.1 | scaffold152:355399:358070 | 1629 | 6 | 542 | 8.64 | 26.44 | 0.044 | No | Plasma membrane |
| Pg4CL7 | Pg013255.1 | scaffold22:4058452:4060831 | 1638 | 7 | 545 | 7.65 | 38.71 | −0.023 | No | Plasma membrane |
| Pg4CL8 | Pg015184.1 | scaffold27:1309358:1311530 | 1722 | 4 | 573 | 5.49 | 40.47 | 0.024 | No | Plasma membrane |
| Pg4CL9 | Pg016048.1 | scaffold3:3588368:3591221 | 1644 | 6 | 547 | 5.64 | 35.48 | 0.120 | No | Cytosol |
| Pg4CL10 | Pg016524.1 | scaffold3:3410537:3413121 | 1659 | 6 | 552 | 8.45 | 42.89 | 0.153 | No | Peroxisome |
| Pg4CL11 | Pg018200.1 | scaffold35:541618:543985 | 1656 | 6 | 551 | 8.7 | 41.33 | 0.037 | No | Plasma membrane |
| Pg4CL12 | Pg029185.1 | scaffold85:64794:69120 | 1731 | 6 | 576 | 8.91 | 44.18 | −0.010 | No | Plasma membrane |
| Motif Code | Sequences | E_Value | Characteristic Domain |
|---|---|---|---|
| Motif 1 | DGWLHTGDLCYFDDEGFLFIVDRJKELIKYKGYQVAPAELEALLLSHPEI | 6.2 × 10−341 | AMP-binding enzyme |
| Motif 2 | GRLVPNMEAKIVDPETGAALPPNQSGELWLRGPTIMKGYLNBPEATAETI | 5.20 × 10−242 | AMP-binding enzyme |
| Motif 3 | DAAVIPYPDEEAGEIPMAFVVRKPGSSITEEDVIDFVAKQVAPYKKIRRV | 7.40 × 10−236 | No |
| Motif 4 | KGDVVLILLPNSJHFPIIYLSILSLGAVITTANPLSTPSEIAKQVRDSKP | 9.60 × 10−162 | AMP-binding enzyme |
| Motif 5 | VRVDQDDTAAJLYSSGTTGTSKGVVLTHRNLIA | 1.90 × 10−160 | AMP-binding enzyme |
| Motif 6 | KSSLTDKYDLSSLRQVGSGAAPLGKEVAEQFRKKFPHVELLQGYGMTEST | 2.90 × 10−160 | AMP-binding enzyme |
| Motif 7 | LRAGETVVVMQKFEFEAMLRAIEKYRVTYLPVVPPIILALV | 4.60 × 10−102 | No |
| Motif 8 | FIESIPKSPAGKILRRELIKK | 2.30 × 10−81 | No |
| Motif 9 | NVFLCVLPLFHIYGLAVIILG | 2.70 × 10−53 | No |
| Motif 10 | SGFCSKTGIFHSLRPPVPLPP | 3.90 × 10−32 | No |
| NO. | Accession NO. | Cultivars | Sample Type | Library | Platform | Reference |
|---|---|---|---|---|---|---|
| P1 | SRR5279396 | ‘Dabenzi’ | root | Paired end | Illumina HiSeq 4000 | [26] |
| P2 | SRR5279397 | ‘Dabenzi’ | leaf | Paired end | Illumina HiSeq 4000 | [26] |
| P3 | SRR5279395 | ‘Dabenzi’ | flower | Paired end | Illumina HiSeq 4000 | [26] |
| P4 | SRR5279391 | ‘Dabenzi’ | Pericarp | Paired end | Illumina HiSeq 4000 | [26] |
| P5 | SRR5279388 | ‘Dabenzi’ | Exocarp | Paired end | Illumina HiSeq 4000 | [26] |
| P6 | SRR5279394 | ‘Dabenzi’ | Pericarp | Paired end | Illumina HiSeq 4000 | [26] |
| P7 | SRR5678820 | ‘Tunisia’ | Pericarp | Paired end | Illumina HiSeq 4000 | [26] |
| P8 | SRR5678819 | ‘Baiyushizi’ | Pericarp | Paired end | Illumina HiSeq 4000 | [26] |
| P9 | SRR080723 | ‘Wonderful’ | Pericarp | Paired end | Illumina HiSeq 2000 | [52] |
| P10 | SRR1055290 | ‘Nana’ | Mixed samples of the root, | Single end | 454 GS FLX Titanium | [53] |
| leaves, flowers, and fruit | ||||||
| P11 | SRR1054190 | ‘Black127’ | Mixed samples of the root, | Single end | 454 GS FLX Titanium | [53] |
| leaves, flowers, and fruit |
| Gene | Primer Sequence (5′-3′) |
|---|---|
| Pg4CL1 | F: GGATGGCTGGTTGAGGACAG; R: GCAGCATCTGCAATTTCGGG |
| Pg4CL2 | F: CAGTGCTCTCGATGTGCCTA; R: TCATAATCTGTTGGCCTCGAA |
| Pg4CL3 | F: AGAATGTGGGGAGGTCCCAG; R: CCTGCTGGAGACTTCGGTATC |
| Pg4CL4 | F: AGAAGCAGGACAGATCCCGA; R: CTTCCCTGCCGGAGATTTCG |
| Pg4CL5 | F: GCATTCGTCATCCGATCCAA; R: CGGATGCCAATTTCGCTCT |
| Pg4CL6 | F: GCTTTCAGATTGCTCCTGCG; R: ATTCTGCACGTCCTCTTCGG |
| Pg4CL7 | F: CCGGAGACCTCGGGTATTTC; R: TCGGGAAACGGAATGACCAC |
| Pg4CL8 | F: AGTTGTACCGGTCGAGGATG; R: CTTGTAAGGCGCAACCTGAG |
| Pg4CL9 | F: CGAATTGTGTGTCCGAAGCC; R: ACGACAAAGACGTCCCCATC |
| Pg4CL10 | F: TACTCCCATCCCGACATTGC; R: GTCGTTTCACCACGAAAGCC |
| Pg4CL11 | F: TCCGAACGGGAGATCTTTGC; R: TCGGGAAACGGGATAACAGC |
| Pg4CL12 | F: GCAGATGCCGCTGTTATTCC; R: TTCCCTGCCGGAGATTTCG |
| PgActin | F: AGTCCTCTTCCAGCCATCTC; R: CACTGAGCACAATGTTTCCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Guo, L.; Zhao, Y.; Zhao, X.; Yuan, Z. Systematic Analysis and Expression Profiles of the 4-Coumarate: CoA Ligase (4CL) Gene Family in Pomegranate (Punica granatum L.). Int. J. Mol. Sci. 2022, 23, 3509. https://doi.org/10.3390/ijms23073509
Wang Y, Guo L, Zhao Y, Zhao X, Yuan Z. Systematic Analysis and Expression Profiles of the 4-Coumarate: CoA Ligase (4CL) Gene Family in Pomegranate (Punica granatum L.). International Journal of Molecular Sciences. 2022; 23(7):3509. https://doi.org/10.3390/ijms23073509
Chicago/Turabian StyleWang, Yuying, Linhui Guo, Yujie Zhao, Xueqing Zhao, and Zhaohe Yuan. 2022. "Systematic Analysis and Expression Profiles of the 4-Coumarate: CoA Ligase (4CL) Gene Family in Pomegranate (Punica granatum L.)" International Journal of Molecular Sciences 23, no. 7: 3509. https://doi.org/10.3390/ijms23073509
APA StyleWang, Y., Guo, L., Zhao, Y., Zhao, X., & Yuan, Z. (2022). Systematic Analysis and Expression Profiles of the 4-Coumarate: CoA Ligase (4CL) Gene Family in Pomegranate (Punica granatum L.). International Journal of Molecular Sciences, 23(7), 3509. https://doi.org/10.3390/ijms23073509