
Inflammatory Response and Secondary White Matter Damage to the Corpus Callosum after Focal Striatal Stroke in Rats

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
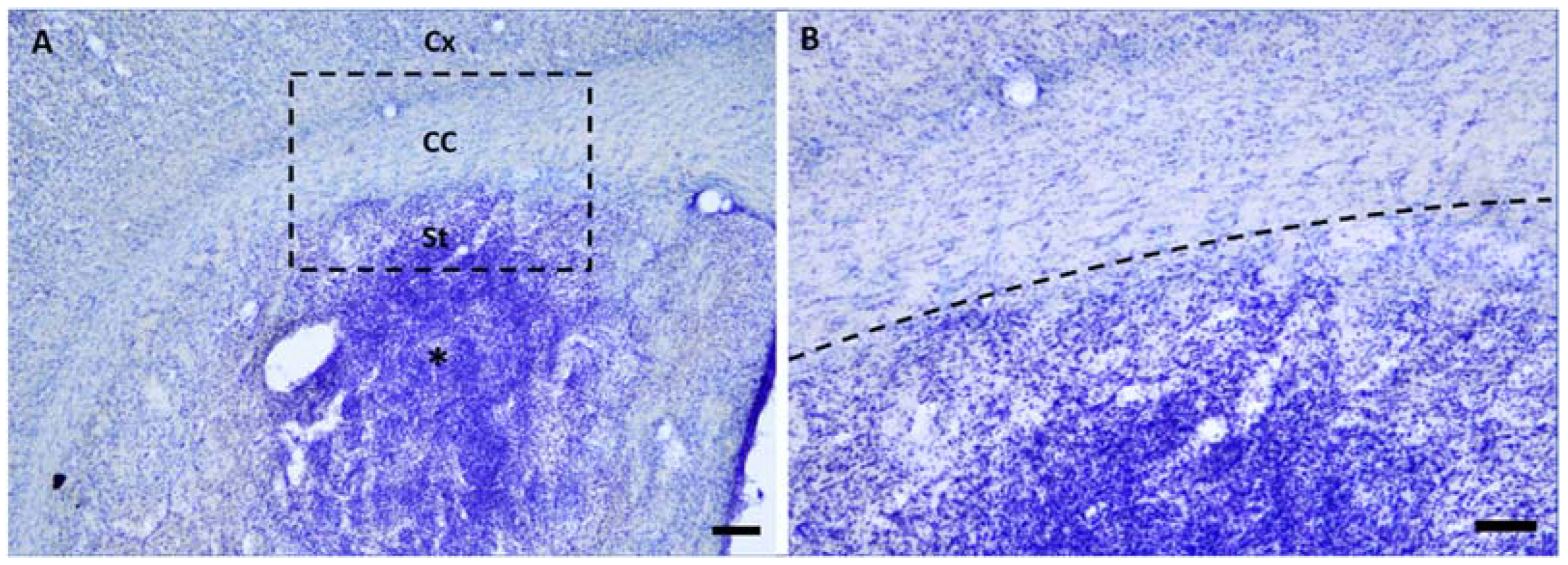
2.1. Microinjections of ET-1 Induced Focal Ischemia Restricted to the Striatum
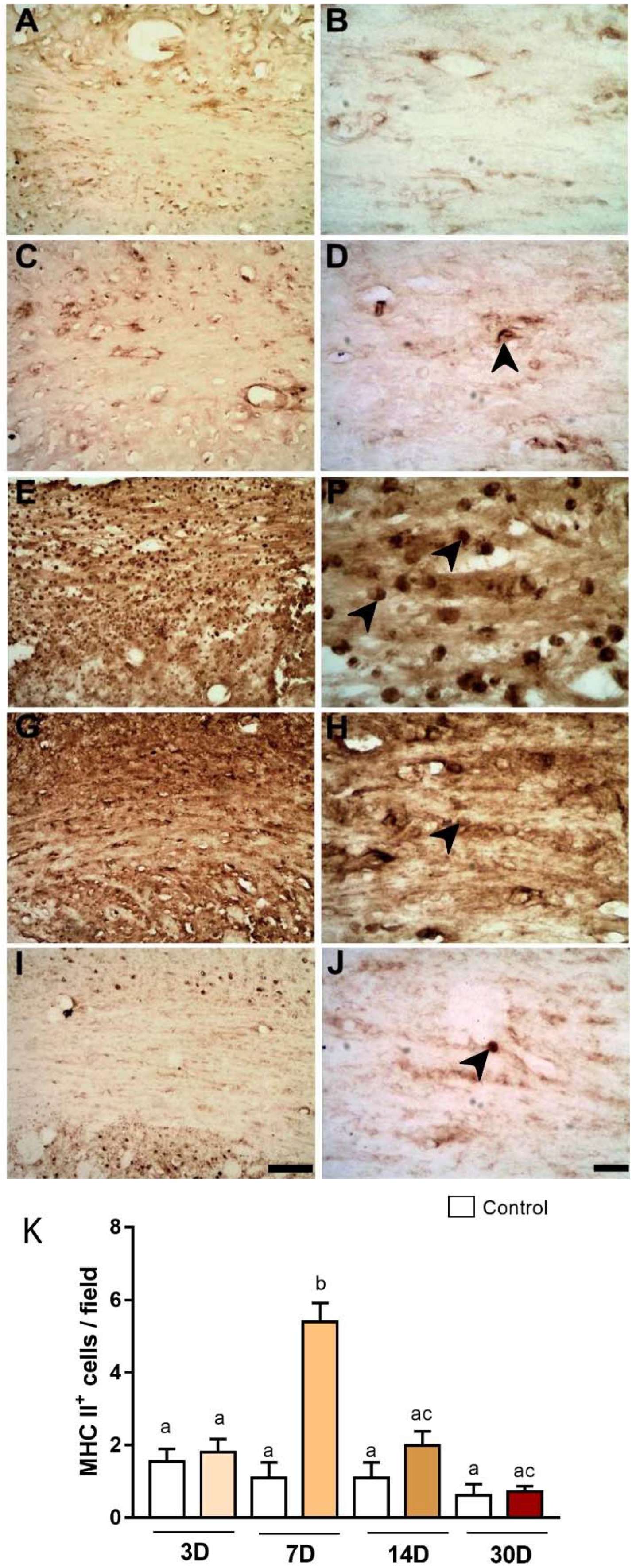
2.2. Microinjections of ET-1 Induced Microglial/Macrophage Activation in the Corpus Callosum
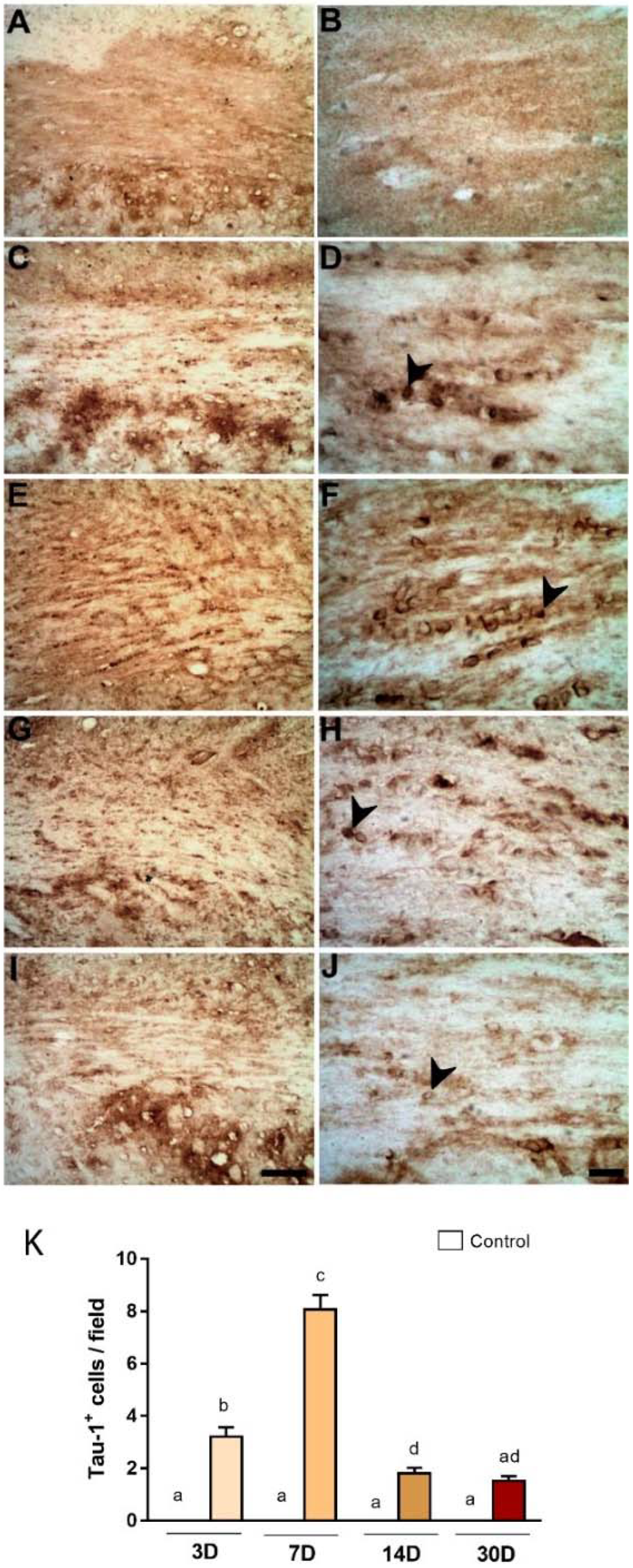
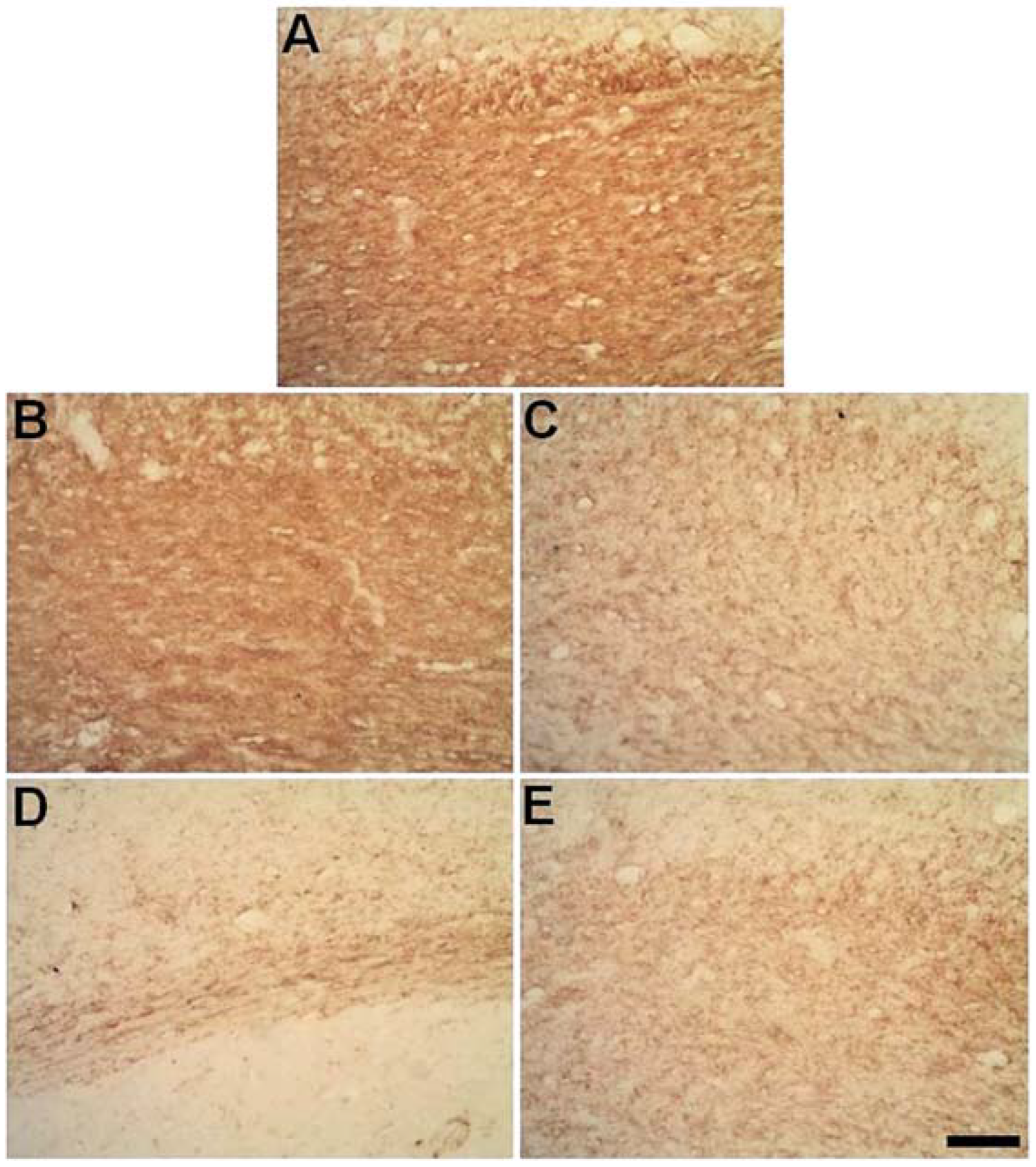
2.3. Degeneration of Oligodendrocytes in the Corpus Callosum after Primary Striatal Ischemia
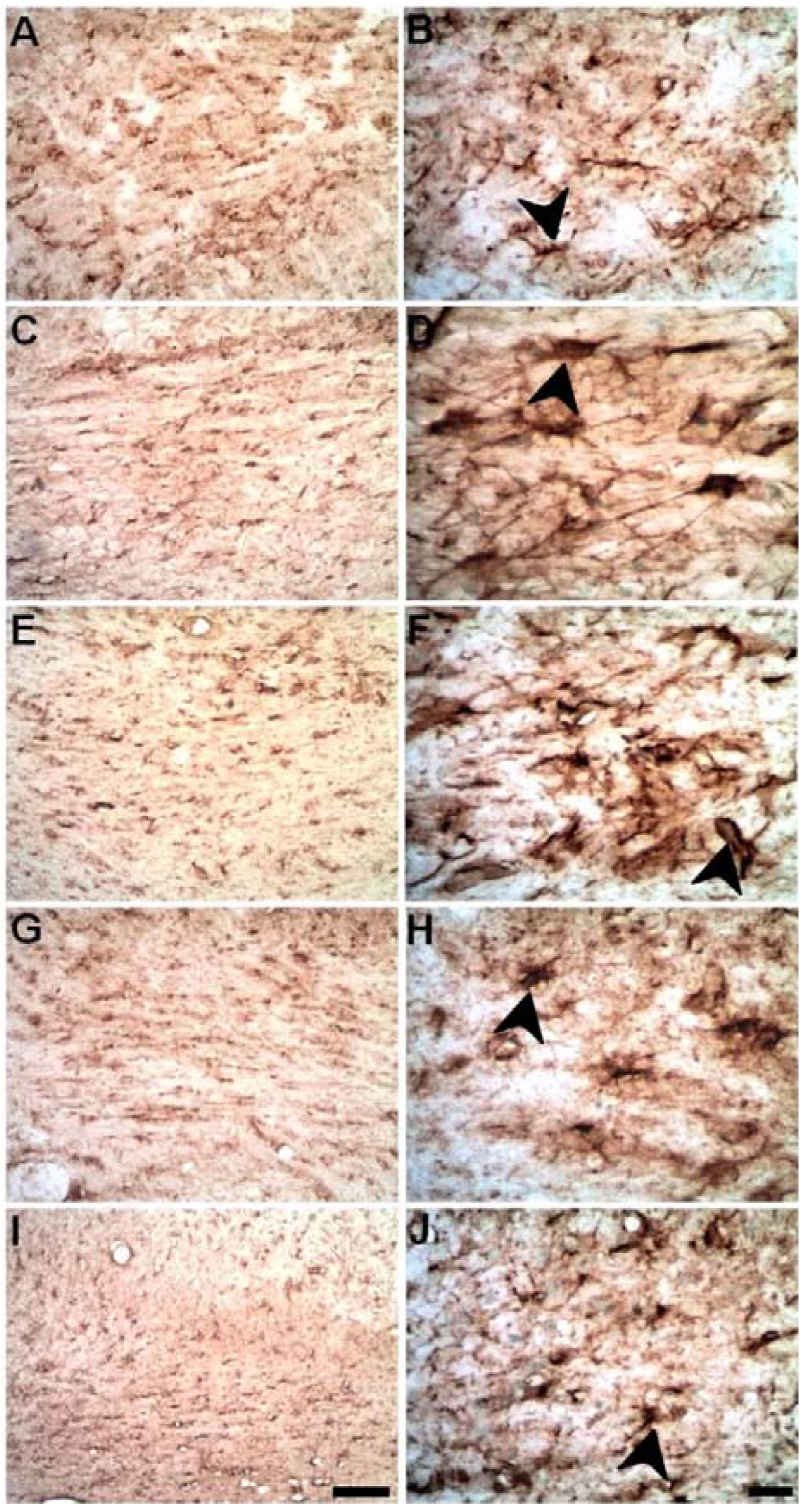
2.4. Microinjections of ET-1 into the Striatum Increase Astrocytosis in the Corpus Callosum
2.5. Progressive Myelin Impairment in Rat’s the Corpus Callosum Secondary to Striatal Ischemia
3. Discussion
4. Materials and Methods
4.1. Ethical Statement and Experimental Animals
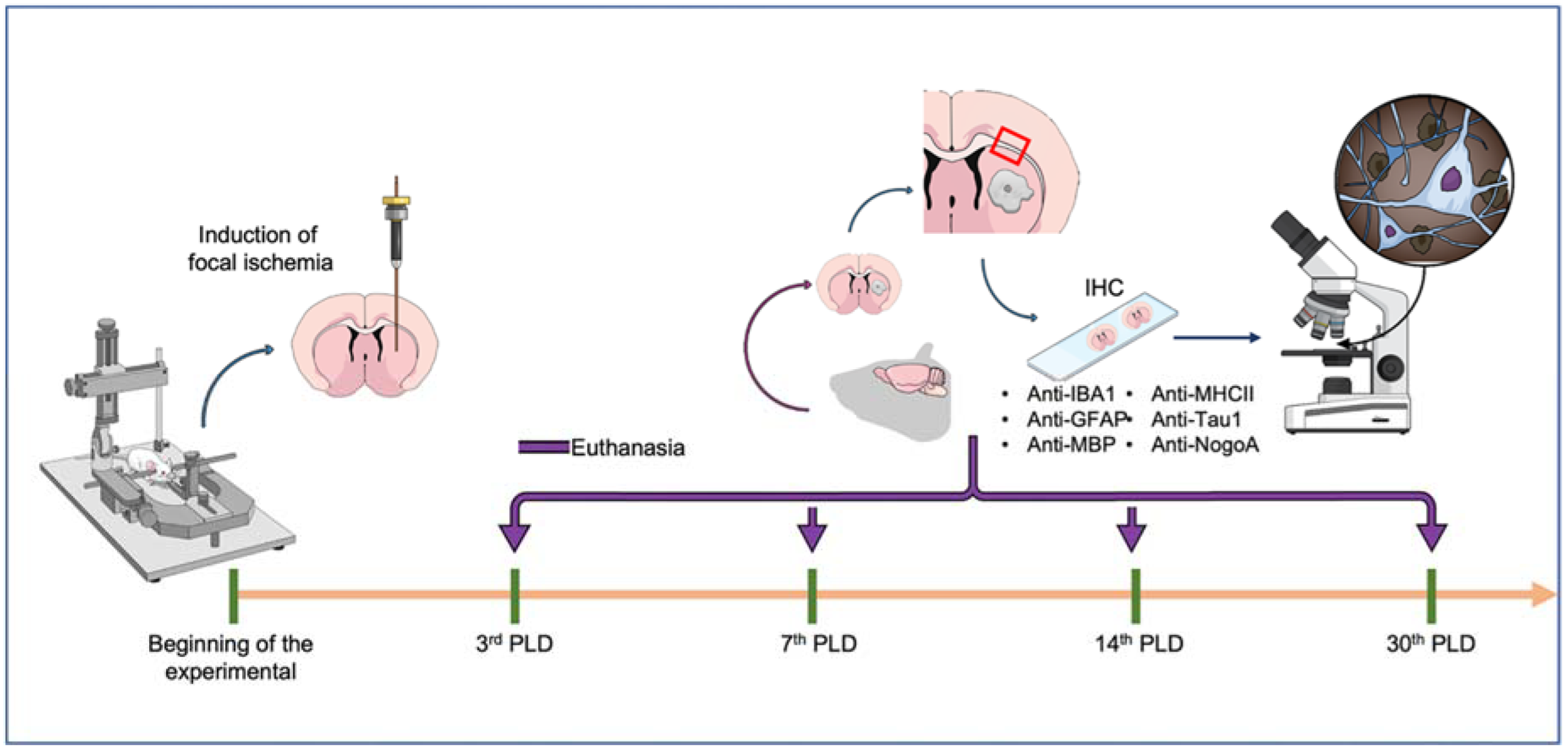
4.2. Induction of Focal Striatal Ischemia
4.3. Perfusion and Histological Procedures
4.3.1. Immunolabeling Protocol
4.3.2. Qualitative and Quantitative Analyses
4.4. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NMDA | N-Methyl-D-Aspartate |
| ET-1 | Endothelin-1 |
| CC | Corpus callosum |
| Cx | Cortex |
| St | Striatum |
| PDL | Post-Lesion Day |
| PBS | Phosphate buffer saline |
| SCI | Spinal cord injury |
References
- Powers, W.J. Acute Ischemic Stroke. N. Engl. J. Med. 2020, 16, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. American Heart Association Council on Epidemiology and Prevention Statistics Committee and Stroke Statistics Subcommittee. Heart Disease and Stroke Statistics-2020 Update: A Report from the American Heart Association. Circulation 2020, 3, e139–e596. [Google Scholar]
- Guzik, A.; Bushnell, C. Stroke Epidemiology and Risk Factor Management. CONTINUUM Lifelong Learn. Neurol. 2017, 23, 15–39. [Google Scholar] [CrossRef]
- Caprio, F.Z.; Sorond, F.A. Cerebrovascular Disease: Primary and Secondary Stroke Prevention. Med. Clin. N. Am. 2019, 103, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Gallacher, K.I.; Jani, B.D.; Hanlon, P.; Nicholl, B.I.; Mair, F.S. Multimorbidity in stroke. Stroke 2019, 50, 919–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnan, G.A.; Fisher, M.; Macleod, M.; Davis, S.M. Stroke. Lancet 2008, 371, 1612–1623. [Google Scholar] [CrossRef]
- Moskowitz, M.A.; Lo, E.H.; Iadecola, C. The science of stroke: Mechanisms in search of treatments. Neuron 2010, 67, 181–198. [Google Scholar] [CrossRef] [Green Version]
- Khoshnam, S.E.; Winlow, W.; Farzaneh, M.; Farbood, Y.; Moghaddam, H.F. Pathogenic mechanisms following ischemic stroke. Neurol. Sci. 2017, 38, 1167–1186. [Google Scholar] [CrossRef]
- Radak, D.; Katsiki, N.; Resanovic, I.; Jovanovic, A.; Sudar-Milovanovic, E.; Zafirovic, S.; Mousad, S.A.; Isenovic, E.R. Apoptosis and Acute Brain Ischemia in Ischemic Stroke. Curr. Vasc. Pharmacol. 2017, 15, 115–122. [Google Scholar] [CrossRef]
- Curcio, M.; Salazar, I.L.; Mele, M.; Canzoniero, L.M.; Duarte, C.B. Calpains and neuronal damage in the ischemic brain: The swiss knife in synaptic injury. Prog. Neurobiol. 2016, 143, 1–35. [Google Scholar] [CrossRef]
- Orellana-Urzúa, S.; Rojas, I.; Líbano, L.; Rodrigo, R. Pathophysiology of Ischemic Stroke: Role of Oxidative Stress. Curr. Pharm. Des. 2020, 26, 4246–4260. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Leal, W. Why microglia kill neurons after neural disorders? The friendly fire hypothesis. Neural Regen. Res. 2019, 14, 1499–1502. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, R.L.; Azimullah, S.; Beiram, R.; Jalal, F.Y.; Rosenberg, G.A. Neuroinflammation: Friend and foe for ischemic stroke. J. Neuroinflamm. 2019, 16, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, K.; Tian, D.C.; Li, Z.G.; Ducruet, A.F.; Lawton, M.T.; Shi, F.D. Global brain inflammation in stroke. Lancet Neurol. 2019, 18, 1058–1066. [Google Scholar] [CrossRef]
- Lo, E.; Dalkara, T.; Moskowitz, M.A. Mechanisms, challenges and opportunities in stroke. Nat. Rev. Neurosci. 2003, 4, 399–415. [Google Scholar] [CrossRef]
- Guimaraes, J.S.; Freire, M.A.M.; Lima, R.R.; Picanco-Diniz, C.W.; Pereira, A.; Gomes-Leal, W. Minocycline treatment reduces white matter damage after excitotoxic striatal injury. Brain Res. 2010, 6, 182–193. [Google Scholar] [CrossRef]
- Thiel, A.; Heiss, W.D. Imaging of microglia activation in stroke. Stroke 2011, 42, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Mattos, D.J.S.; Rutlin, J.; Hong, X.; Zinn, K.; Shimony, J.S.; Carter, A.R. White matter integrity of contralesional and transcallosal tracts may predict response to upper limb task-specific training in chronic stroke. Neuroimage Clin. 2021, 31, 102710. [Google Scholar] [CrossRef]
- Garcia-Martin, G.; Alcover-Sanchez, B.; Wandosell, F.; Cubelos, B. Pathways involved in remyelination after cerebral ischemia. Curr. Neuropharmacol. 2021. In press. [Google Scholar] [CrossRef]
- Gomes-Leal, W.; Corkill, D.J.; Freire, M.A.M.; Picanco-Diniz, C.W.; Perry, V.H. Astrocytosis, microglia activation, oligodendrocyte degeneration, and pyknosis following acute spinal cord injury. Exp. Neurol. 2004, 190, 456–467. [Google Scholar] [CrossRef]
- Gomes-Leal, W.; Corkill, D.J.; Picanco-Diniz, C.W. Systematic analysis of axonal damage and inflammatory response in different white matter tracts of acutely injured rat spinal cord. Brain Res. 2005, 1066, 57–70. [Google Scholar] [CrossRef]
- Shahsavani, N.; Kataria, H.; Karimi-Abdolrezaee, S. Mechanisms and repair strategies for white matter degeneration in CNS injury and diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1, 166117. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, W.A.; Bahia, C.P.; Teixeira, J.C.; Gomes-Leal, W.; Pereira, A. Interlimb Dynamic after Unilateral Focal Lesion of the Cervical Dorsal Corticospinal Tract with Endothelin-1. Front. Neuroanat. 2017, 11, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, R.R.; Guimaraes-Silva, J.; Oliveira, J.L.; Costa, A.M.R.; Souzarodrigues, R.; Dos Santos, C.D.; Picanço-Diniz, C.W.; Gomes-Leal, W. Diffuse Axonal Damage, myelin impairment astrocytosis and Inflammatory Response Following Microinjections of NMDA into The Rat Striatum. Inflammation 2008, 3, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Freire, M.A.M.; Guimaraes, J.S.; Santos, J.R.; Simplício, H.; Gomes-Leal, W. Morphometric analysis of NADPH diaphorase reactive neurons in a rat model of focal excitotoxic striatal injury. Neuropathology 2016, 36, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Paul, L.K.; Brown, W.S.; Adolphs, R.; Tyszka, J.M.; Richards, L.J.; Mukherjee, P.; Sherr, E.H. Agenesis of the corpus callosum: Genetic, developmental and functional aspects of connectivity. Nat. Rev. Neurosci. 2007, 8, 287–299. [Google Scholar] [CrossRef]
- Irving, E.A.; Yatsushiro, K.; McCulloch, J.; Dewar, D. Rapid alteration of tau in oligodendrocytes after focal ischemic injury in the rat: Involvement of free radicals. J. Cereb. Blood Flow Metab. 1997, 17, 612–622. [Google Scholar] [CrossRef]
- Chen, M.S.; Huber, A.B.; van der Haar, M.E.; Frank, M.; Schnell, L.; Spillmann, A.A.; Christ, F.; Schwab, M.E. Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature 2000, 403, 434–439. [Google Scholar] [CrossRef]
- Lima, R.R.; Santana, L.N.; Fernandes, R.M.; Nascimento, E.M.; Oliveira, A.C.A.; Fernandes, L.M.; Santos, E.M.N.D.; Tavares, P.A.N.; Santos, I.R.D.; Gimarães-Santos, A.; et al. Neurodegeneration and Glial Response after Acute Striatal Stroke: Histological Basis for Neuroprotective Studies. Oxid. Med. Cell Longev. 2016, 2016, 3173564. [Google Scholar] [CrossRef]
- Szczupak, D.; Kossmann Ferraz, M.; Gemal, L.; Oliveira-Szejnfeld, P.S.; Monteiro, M.; Bramati, I.; Vargas, F.R.; IRC5 Consortium; Lent, R.; Silva, A.C.; et al. Corpus callosum dysgenesis causes novel patterns of structural and functional brain connectivity. Brain Commun. 2021, 14, fcab057. [Google Scholar] [CrossRef]
- Gomes-Leal, W. Microglial physiopathology: How to explain the dual role of microglia after acute neural disorders? Brain Behav. 2012, 2, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Butovsky, O.; Talpalar, A.E.; Ben-Yaakov, K.; Schwartz, M. Activation of microglia by aggregated beta-amyloid or lipopolysaccharide impairs MHC-II expression and renders them cytotoxic whereas IFN-gamma and IL-4 render them protective. Mol. Cell. Neurosci. 2005, 29, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.B.; Buss, A.; Breuer, S.; Brook, G.A.; Pech, K.; Martin, D.; Schoenen, J.; Noth, J.; Love, S.; Schröder, J.M.; et al. Major histocompatibility complex class II expression by activated microglia caudal to lesions of descending tracts in the human spinal cord is not associated with a T cell response. Acta Neuropathol. 2000, 100, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Yrjanheikki, J.; Tikka, T.; Keinanen, R.; Goldsteins, G.; Chan, P.H.; Koistinaho, J. A tetracycline derivative, minocycline, reduces inflammation and protects against focal cerebral ischemia with a wide therapeutic window. Proc. Natl. Acad. Sci. USA 1999, 96, 13496–13500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, E.C.; Cardoso, M.M.; Gouveia, A.; Pereira, A.; Gomes-Leal, W. Modulation of microglial activation enhances neuroprotection and functional recovery derived from bone marrow mononuclear cell transplantation after cortical ischemia. Neurosci. Res. 2012, 73, 122–132. [Google Scholar] [CrossRef]
- Cardoso, M.M.; Franco, E.C.; de Souza, C.C.; da Silva, M.C.; Gouveia, A.; Gomes-Leal, W. Minocycline treatment and bone marrow mononuclear cell transplantation after endothelin-1 induced striatal ischemia. Inflammation 2013, 36, 197–205. [Google Scholar] [CrossRef]
- Han, R.T.; Kim, R.D.; Molofsky, A.V.; Liddelow, S.A. Astrocyte-immune cell interactions in physiology and pathology. Immunity 2021, 54, 211–224. [Google Scholar] [CrossRef]
- Hasel, P.; Rose, I.V.L.; Sadick, J.S.; Kim, R.D.; Liddelow, S.A. Neuroinflammatory astrocyte subtypes in the mouse brain. Nat. Neurosci. 2021, 24, 1475–1487. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Marsh, S.E.; Stevens, B. Microglia and Astrocytes in Disease: Dynamic Duo or Partners in Crime? Trends Immunol. 2020, 41, 820–835. [Google Scholar] [CrossRef]
- Irving, E.A.; Nicoll, J.; Graham, D.I.; Dewar, D. Increased tau immunoreactivity in oligodendrocytes following human stroke and head injury. Neurosci. Lett. 1996, 213, 189–192. [Google Scholar] [CrossRef]
- Irving, E.A.; Bentley, D.L.; Parsons, A.A. Assessment of white matter injury following prolonged focal cerebral ischaemia in the rat. Acta Neuropathol. 2001, 102, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Valeriani, V.; Dewar, D.; McCulloch, J. Quantitative assessment of ischemic pathology in axons, oligodendrocytes, and neurons: Attenuation of damage after transient ischemia. J. Cereb. Blood Flow Metab. 2000, 20, 765–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weingarten, M.D.; Lockwood, A.H.; Hwo, S.Y.; Kirschner, M.W. A protein factor essential for microtubule assembly. Proc. Natl. Acad. Sci. USA 1975, 72, 1858–1862. [Google Scholar] [CrossRef] [Green Version]
- Stirling, D.P.; Khodarahmi, K.; Liu, J.; McPhail, L.T.; McBride, C.B.; Steeves, J.D.; Ramer, M.S.; Tetzlaff, W. Minocycline treatment reduces delayed oligodendrocyte death, attenuates axonal dieback, and improves functional outcome after spinal cord injury. J. Neurosci. 2004, 24, 2182–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedraza, S.; Puig, J.; Blasco, G.; Prados, F.; Thiel, A. Diffusion tensor imaging, permanent pyramidal tract damage, and outcome in subcortical stroke. Neurology 2011, 3, 1606. [Google Scholar] [CrossRef] [PubMed]
- Wiessner, C.; Bareyre, F.M.; Allegrini, P.R.; Mir, A.K.; Frentzel, S.; Zurini, M.; Schnell, L.; Oertle, T.; Schwab, M.E. Anti-Nogo-Aantibody infusion 24 hours after experimental stroke improved behavioral outcome and corticospinal plasticity in normotensive and spontaneously hypertensive rats. J. Cereb. Blood Flow Metab. 2003, 23, 154–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheatwood, J.L.; Emerick, A.J.; Schwab, M.E.; Kartje, G.L. Nogo-a expression after focal ischemic stroke in the adult rat. Stroke 2008, 39, 2091–2098. [Google Scholar] [CrossRef] [Green Version]
- Schnell, L.; Schwab, M.E. Axonal regeneration in the rat spinal cord produced by an antibody against myelin associated neurite growth inhibitors. Nature 1990, 343, 269–272. [Google Scholar] [CrossRef]
- Seymour, A.B.; Andrews, E.M.; Tsai, S.Y.; Markus, T.M.; Bollnow, M.R.; Brenneman, M.M.; Obrien, T.E.; Castro, A.J.; Schwab, M.E.; Kartje, G.L. Delayed treatment with monoclonal antibody IN-1 1 week after stroke results in recovery of function and corticorubral plasticity in adult rats. J. Cereb. Blood Flow Metab. 2005, 5, 1366–1375. [Google Scholar] [CrossRef] [Green Version]
- Freund, P.; Schmidlin, E.; Wannier, T.; Bloch, J.; Mir, A.; Schwab, M.E.; Rouiller, E.M. Anti-Nogo-A antibody treatment promotes recovery of manual dexterity after unilateral cervical lesion in adult primates--re-examination and extension of behavioral data. Eur. J. Neurosci. 2009, 29, 983–996. [Google Scholar] [CrossRef] [Green Version]
- Kilic, E.; ElAli, A.; Kilic, Ü.; Guo, Z.; Ugur, M.; Uslu, U.; Bassetti, C.L.; Schwab, M.E.; Hermann, D.M. Role of Nogo-A in neuronal survival in the reperfused ischemic brain. J. Cereb. Blood Flow Metab. 2010, 30, 969–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-de-Puig, I.; Miró-Mur, F.; Ferrer-Ferrer, M.; Gelpi, E.; Pedragosa, J.; Justicia, C.; Urra, X.; Chamorro, A.; Planas, A.M. Neutrophil recruitment to the brain in mouse and human ischemic stroke. Acta Neuropathol. 2015, 129, 239–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhaskar, S.; Bivard, A.; Stanwell, P.; Parsons, M.; Attia, J.R.; Nilsson, M.; Levi, C. Baseline collateral status and infarct topography in post-ischaemic perilesional hyperperfusion: An arterial spin labelling study. J. Cereb. Blood Flow Metab. 2017, 37, 1148–1162. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, A.; Weissert, R.; Bhaskar, S.M.M. Leukoaraiosis severity and post-reperfusion outcomes in acute ischaemic stroke: A meta-analysis. Acta Neurol. Scand. 2022, 145, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.E.; Tittgemeyer, M.; Imperati, D.; Diekhoff, S.; Ameli, M.; Fink, G.R.; Grefkes, C. Degeneration of corpus callosum and recovery of motor function after stroke: A multimodal magnetic resonance imaging study. Hum. Brain Mapp. 2012, 33, 2941–2956. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier Science: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Freire, M.A.M.; Faber, J.; Picanco-Diniz, C.W.; Franca, J.G.; Pereira, A. Morphometric variability of nicotinamide adenine dinucleotide phosphate diaphorase neurons in the primary sensory areas of the rat. Neuroscience 2012, 205, 140–153. [Google Scholar] [CrossRef]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-specific localisation of a novel calcium binding protein, Iba1. Mol. Brain Res. 1998, 57, 1–9. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Meth. 2007, 39, 175–191. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Secondary Antibodies | Normal Serum (10%) | Labeling Purpose |
|---|---|---|---|
| Anti-MHC-II (1:100, Serotec, Oxford, UK) | Horse anti-mouse (1:100, Vector Labs, Burlingame, CA, USA) | Horse | Microglia |
| Anti-Iba-1 (1:1000, WAKO, Richmond, VA, USA) | Goat anti-rabbit (1:200, Vector Labs, Burlingame, CA, USA) | Goat | Microglia/macrophages |
| Anti-GFAP (GFAP, 1:1000, DAKO, Glostup, Denmark) | Goat anti-rabbit (1:200, Vector Labs, Burlingame, CA, USA) | Goat | Astrocytes |
| Anti-MBP (1:100, Serotec, Oxford, UK) | Horse anti-mouse (1:100, Vector Labs, Burlingame, CA, USA) | Horse | Myelin |
| Anti-Tau-1 (1:500, Chemicon, Temecula, CA, USA) | Horse anti-mouse (1:100, Vector Labs, Burlingame, CA, USA) | Horse | Pathological oligodendrocytes |
| Anti-Nogo-A (1:100, BD Transduction Lab, La Jolla, CA, USA) | Horse anti-mouse (1:100, Vector Labs, Burlingame, CA, USA) | Horse | Inhibition of axonal regeneration in oligodendrocytes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, R.R.; Oliveira, A.C.A.; Fernandes, R.M.; Nascimento, P.C.; Freire, M.A.M.; Gomes-Leal, W. Inflammatory Response and Secondary White Matter Damage to the Corpus Callosum after Focal Striatal Stroke in Rats. Int. J. Mol. Sci. 2022, 23, 3179. https://doi.org/10.3390/ijms23063179
Lima RR, Oliveira ACA, Fernandes RM, Nascimento PC, Freire MAM, Gomes-Leal W. Inflammatory Response and Secondary White Matter Damage to the Corpus Callosum after Focal Striatal Stroke in Rats. International Journal of Molecular Sciences. 2022; 23(6):3179. https://doi.org/10.3390/ijms23063179
Chicago/Turabian StyleLima, Rafael Rodrigues, Ana Carolina Alves Oliveira, Rafael Monteiro Fernandes, Priscila Cunha Nascimento, Marco Aurelio M. Freire, and Walace Gomes-Leal. 2022. "Inflammatory Response and Secondary White Matter Damage to the Corpus Callosum after Focal Striatal Stroke in Rats" International Journal of Molecular Sciences 23, no. 6: 3179. https://doi.org/10.3390/ijms23063179
APA StyleLima, R. R., Oliveira, A. C. A., Fernandes, R. M., Nascimento, P. C., Freire, M. A. M., & Gomes-Leal, W. (2022). Inflammatory Response and Secondary White Matter Damage to the Corpus Callosum after Focal Striatal Stroke in Rats. International Journal of Molecular Sciences, 23(6), 3179. https://doi.org/10.3390/ijms23063179