
Immune Dysregulation in Autism Spectrum Disorder: What Do We Know about It?


 ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
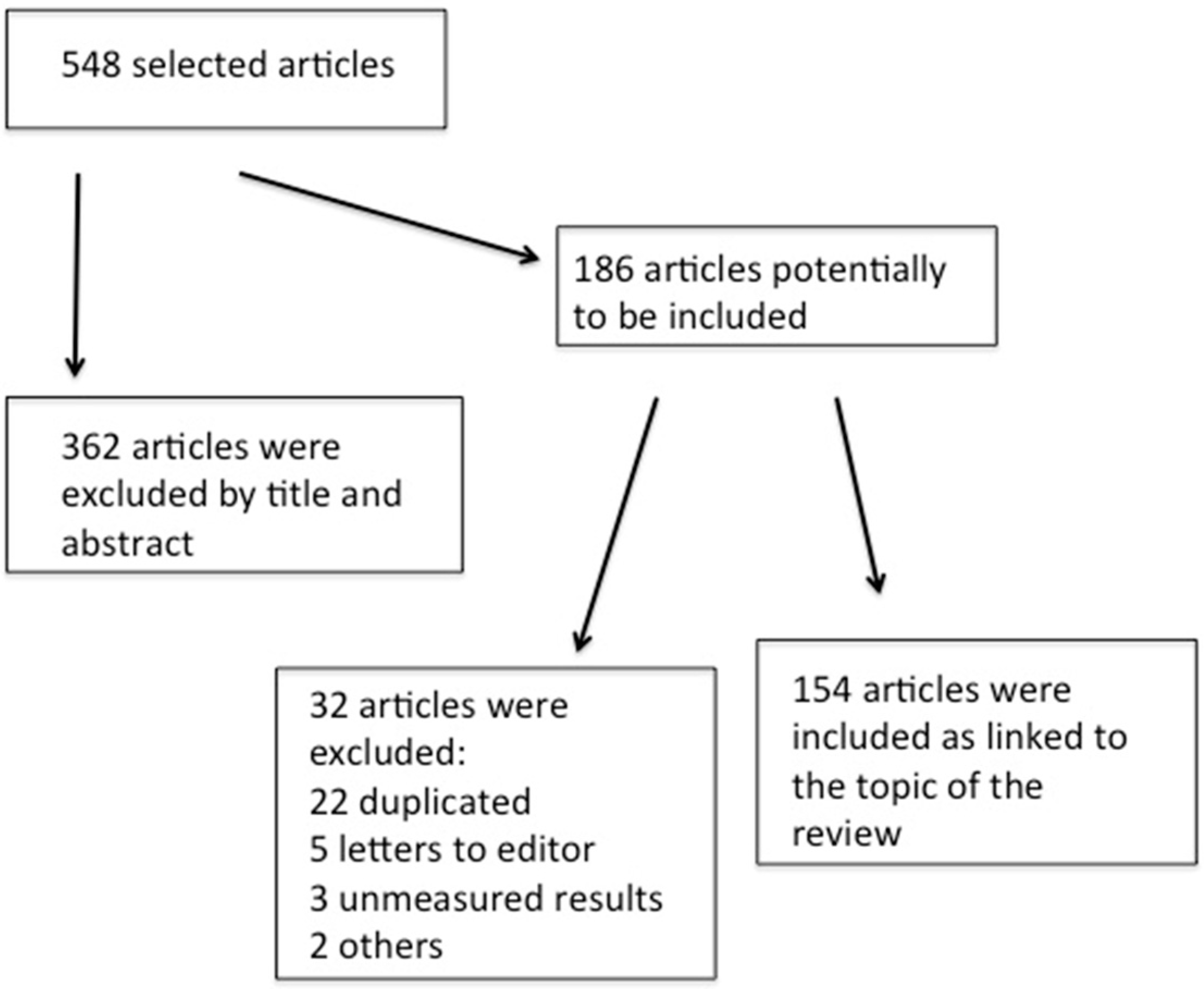
2. Methods
3. Genetic Factors and Immunological Disturbance in Autism Spectrum Disorders
3.1. Major Histocompatibility Complex (MHC) and Autism Spectrum Disorder
3.2. Adaptive Immune Reaction in Autism Spectrum Disorders
3.2.1. Cellular Immune Reaction in Autism
3.2.2. Adaptive Immune Response in Autism
Antibody Reaction in ASD Brain Tissue
Peripheral Immunoglobulins Response in Autism
Maternal Autoantibodies Influencing Gestational Environment in Risk for ASD
3.3. Innate Immunity in ASD
3.3.1. Natural Killer Cells
3.3.2. Monocytes
3.3.3. Microglia
4. Cytokines and Chemokines in Brain and Peripheral Compartment in Autism
4.1. Cytokines Affecting the Gestational Environment Increasing Risk for Autism
4.2. Cytokines and Clinical Phenotype in ASD
4.3. Immune Dysregulation and Comorbidities with High Incidence in ASD
5. Obesity, Fatty Acids, and Risk for Autism Pathology
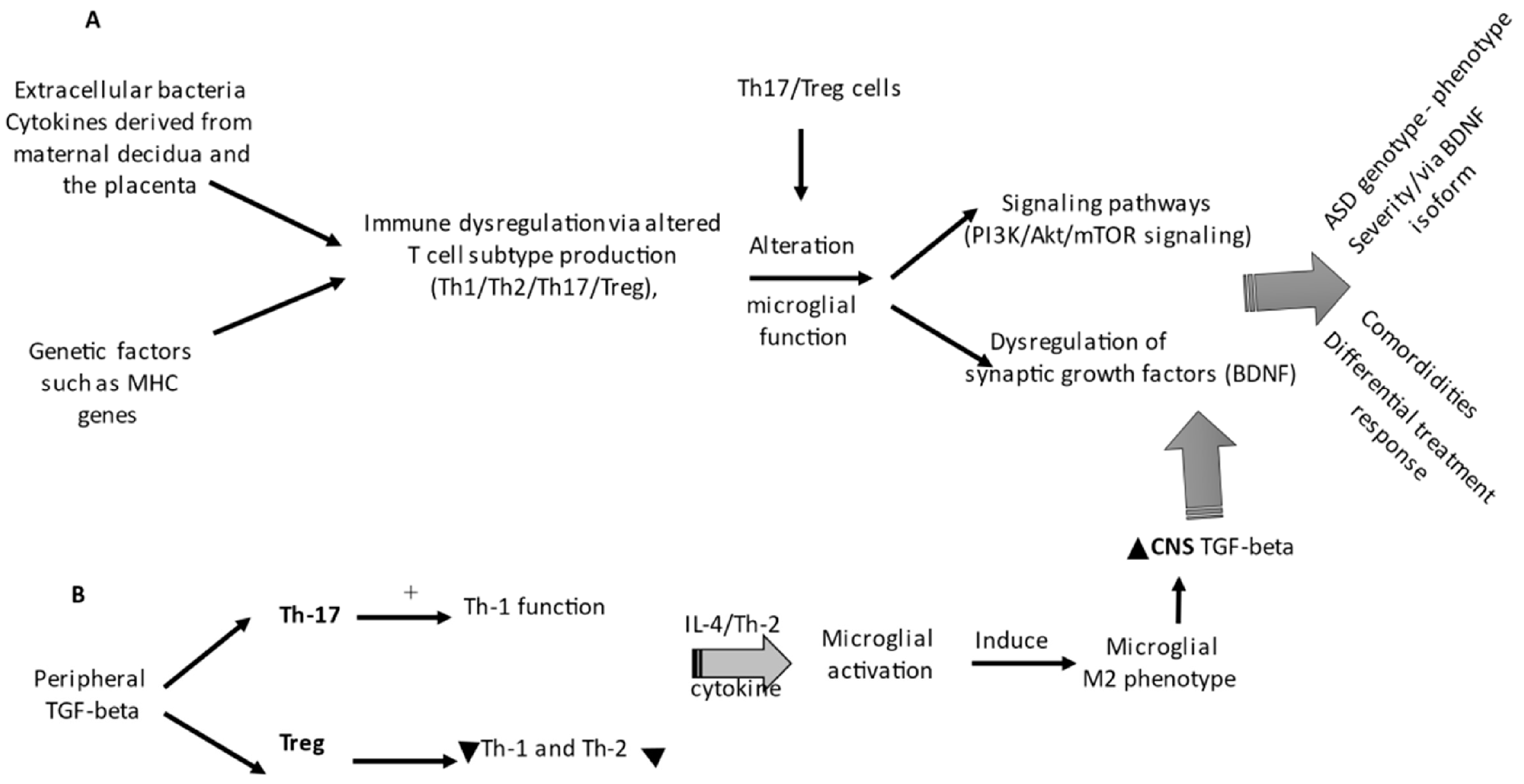
6. Immune Signaling Pathway Overlapping Autism and Neurodegenerative Disorders
Immunopathology in Both ASD and PD
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chovatiya, R.; Medzhitov, R. Stress, inflammation, and defense of homeostasis. Mol. Cell 2014, 54, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raphael, I.; Joern, R.R.; Forsthuber, T.G. Memory CD4+ T cells in immunity and autoimmune diseases. Cells 2020, 9, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meltzer, A.; Van de Water, J. The role of the immune system in autism spectrum disorder. Neuropsychopharmacology 2017, 42, 284–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawabe, T.; Yi, J.; Kawajiri, A.; Hilligan, K.; Fang, D.; Ishii, N.; Yamane, H.; Zhu, J.; Jankovic, D.; Kim, K.S.; et al. Requirements for the differentiation of innate t-bethigh memory-phenotype cd4+ t lymphocytes under steady state. Nat. Commun. 2020, 11, 3366. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Maternal immune activation: Implications for neuropsychiatric disorders. Science 2016, 353, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.L.; Van de Water, J. Maternal autoantibody related autism: Mechanisms and pathways. Mol. Psychiatry 2019, 24, 252–265. [Google Scholar] [CrossRef]
- Careaga, M.; Rogers, S.; Hansen, R.L.; Amaral, D.G.; de Water, J.V.; Ashwood, P. Immune endophenotypes in children with autism spectrum disorder. Biol. Psychiatry 2017, 81, 434–441. [Google Scholar] [CrossRef] [Green Version]
- Krakowiak, P.; Goines, P.E.; Tancredi, D.J.; Ashwood, P.; Hansen, R.L.; Hertz-Picciotto, I.; Van de Water, J. Neonatal cytokine profiles associated with autism spectrum disorder. Biol. Psychiatry 2017, 81, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Lord, C.; Elsabbagh, M.; Baird, G.; Veenstra-Vanderweele, J. Autism spectrum disorder. Lancet 2018, 392, 508–520. [Google Scholar] [CrossRef]
- Weizman, A.; Weizman, R.; Szekely, G.A.; Wijsenbeek, H.; Livni, E. Abnormal immune response to brain tissue antigen in the syndrome of autism. Am. J. Psychiatry 1982, 139, 1462–1465. [Google Scholar]
- Cohly, H.H.; Panja, A. Immunological findings in autism. Int. Rev. Neurobiol. 2005, 71, 317–341. [Google Scholar] [PubMed]
- Hosomichi, K.; Shiina, T.; Tajima, A.; Inoue, I. The impact of next-generation sequencing technologies on hla research. J. Hum. Genet. 2015, 60, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Palmen, S.J.; van Engeland, H. Review on structural neuroimaging findings in autism. J. Neural Transm. 2004, 111, 903–929. [Google Scholar] [CrossRef] [PubMed]
- Sinkus, M.L.; Adams, C.E.; Logel, J.; Freedman, R.; Leonard, S. Expression of immune genes on chromosome 6p21.3-22.1 in schizophrenia. Brain Behav. Immun. 2013, 32, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hakbany, M.; Awadallah, S.; Al-Ayadhi, L. The relationship of hla class i and ii alleles and haplotypes with autism: A case control study. Autism Res. Treat. 2014, 2014, 242048. [Google Scholar] [CrossRef] [PubMed]
- Bennabi, M.; Gaman, A.; Delorme, R.; Boukouaci, W.; Manier, C.; Scheid, I.; Si Mohammed, N.; Bengoufa, D.; Charron, D.; Krishnamoorthy, R.; et al. Hla-class II haplotypes and autism spectrum disorders. Sci. Rep. 2018, 8, 7639. [Google Scholar] [CrossRef] [Green Version]
- Harville, T.; Rhodes-Clark, B.; Bennuri, S.C.; Delhey, L.; Slattery, J.; Tippett, M.; Wynne, R.; Rose, S.; Kahler, S.; Frye, R.E. Inheritance of hla-cw7 associated with autism spectrum disorder (asd). Front. Psychiatry 2019, 10, 612. [Google Scholar] [CrossRef]
- Needleman, L.A.; McAllister, A.K. The major histocompatibility complex and autism spectrum disorder. Dev. Neurobiol. 2012, 72, 1288–1301. [Google Scholar] [CrossRef]
- Torres, A.R.; Westover, J.B.; Rosenspire, A.J. Hla immune function genes in autism. Autism Res. Treat. 2012, 2012, 959073. [Google Scholar] [CrossRef] [Green Version]
- Sweeten, T.L.; Odell, D.W.; Odell, J.D.; Torres, A.R. C4b null alleles are not associated with genetic polymorphisms in the adjacent gene cyp21a2 in autism. BMC Med. Genet. 2008, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Warren, R.P.; Singh, V.K.; Cole, P.; Odell, J.D.; Pingree, C.B.; Warren, W.L.; White, E. Increased frequency of the null allele at the complement c4b locus in autism. Clin. Exp. Immunol. 1991, 83, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Hashem, S.; Nisar, S.; Bhat, A.A.; Yadav, S.K.; Azeem, M.W.; Bagga, P.; Fakhro, K.; Reddy, R.; Frenneaux, M.P.; Haris, M. Genetics of structural and functional brain changes in autism spectrum disorder. Transl. Psychiatry 2020, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Hu, V.W. From genes to environment: Using integrative genomics to build a “systems level” understanding of autism spectrum disorders. Child Dev. 2013, 84, 89–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippi, A.; Tores, F.; Carayol, J.; Rousseau, F.; Letexier, M.; Roschmann, E.; Lindenbaum, P.; Benajjou, A.; Fontaine, K.; Vazart, C.; et al. Association of autism with polymorphisms in the paired-like homeodomain transcription factor 1 (pitx1) on chromosome 5q31: A candidate gene analysis. BMC Med. Genet. 2007, 8, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siniscalco, D.; Schultz, S.; Brigida, A.L.; Antonucci, N. Inflammation and neuro-immune dysregulations in autism spectrum disorders. Pharmaceuticals 2018, 11, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enstrom, A.M.; Lit, L.; Onore, C.E.; Gregg, J.P.; Hansen, R.L.; Pessah, I.N.; Hertz-Picciotto, I.; Van de Water, J.A.; Sharp, F.R.; Ashwood, P. Altered gene expression and function of peripheral blood natural killer cells in children with autism. Brain Behav. Immun. 2009, 23, 124–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, D.B.; Sutcliffe, J.S.; Ebert, P.J.; Militerni, R.; Bravaccio, C.; Trillo, S.; Elia, M.; Schneider, C.; Melmed, R.; Sacco, R.; et al. A genetic variant that disrupts met transcription is associated with autism. Proc. Natl. Acad. Sci. USA 2006, 103, 16834–16839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuer, L.; Braunschweig, D.; Ashwood, P.; Van de Water, J.; Campbell, D.B. Association of a met genetic variant with autism-associated maternal autoantibodies to fetal brain proteins and cytokine expression. Transl. Psychiatry 2011, 1, e48. [Google Scholar] [CrossRef] [Green Version]
- Huh, G.S.; Boulanger, L.M.; Du, H.; Riquelme, P.A.; Brotz, T.M.; Shatz, C.J. Functional requirement for class I mhc in cns development and plasticity. Science 2000, 290, 2155–2159. [Google Scholar] [CrossRef] [Green Version]
- Needleman, L.A.; Liu, X.B.; El-Sabeawy, F.; Jones, E.G.; McAllister, A.K. Mhc class I molecules are present both pre- and postsynaptically in the visual cortex during postnatal development and in adulthood. Proc. Natl. Acad. Sci. USA 2010, 107, 16999–17004. [Google Scholar] [CrossRef] [Green Version]
- Glynn, M.W.; Elmer, B.M.; Garay, P.A.; Liu, X.B.; Needleman, L.A.; El-Sabeawy, F.; McAllister, A.K. Mhc class i negatively regulates synapse density during the establishment of cortical connections. Nat. Neurosci. 2011, 14, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goddard, C.A.; Butts, D.A.; Shatz, C.J. Regulation of cns synapses by neuronal mhc class i. Proc. Natl. Acad. Sci. USA 2007, 104, 6828–6833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autry, A.E.; Monteggia, L.M. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol. Rev. 2012, 64, 238–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drescher, H.K.; Bartsch, L.M.; Weiskirchen, S.; Weiskirchen, R. Intrahepatic th17/treg cells in homeostasis and disease—It’s all about the balance. Front. Pharmacol. 2020, 11, 1598. [Google Scholar] [CrossRef]
- McAllister, A.K. Immune contributions to cause and effect in autism spectrum disorder. Biol. Psychiatry 2017, 81, 380–382. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.P.; Li, J.; Garbett, K.; Mirnics, K.; Patterson, P.H. Maternal immune activation alters fetal brain development through interleukin-6. J. Neurosci. 2007, 27, 10695–10702. [Google Scholar] [CrossRef] [Green Version]
- Pentón-Rol, G.; Cervantes-Llanos, M.; Martínez-Sánchez, G.; Cabrera-Gómez, J.A.; Valenzuela-Silva, C.M.; Ramírez-Nuñez, O.; Casanova-Orta, M.; Robinson-Agramonte, M.A.; Lopategui-Cabezas, I.; López-Saura, P.A. Tnf-α and il-10 downregulation and marked oxidative stress in neuromyelitis optica. J. Inflamm. 2009, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves Pereira, M.H.; Figueiredo, M.M.; Queiroz, C.P.; Magalhães, T.V.B.; Mafra, A.; Diniz, L.M.O.; da Costa, Ú.L.; Gollob, K.J.; Antonelli, L.V.; Santiago Hda, C. T-cells producing multiple combinations of infγ, tnf and il10 are associated with mild forms of dengue infection. Immunology 2020, 160, 90–102. [Google Scholar] [CrossRef]
- Xie, J.; Huang, L.; Li, X.; Li, H.; Zhou, Y.; Zhu, H.; Pan, T.; Kendrick, K.M.; Xu, W. Immunological cytokine profiling identifies tnf-α as a key molecule dysregulated in autistic children. Oncotarget 2017, 8, 82390–82398. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Chauhan, A.; Sheikh, A.M.; Patil, S.; Chauhan, V.; Li, X.M.; Ji, L.; Brown, T.; Malik, M. Elevated immune response in the brain of autistic patients. J. Neuroimmunol. 2009, 207, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Manzardo, A.; Henkhaus, R.; Dhillon, S.; Butler, M. Plasma cytokine levels in children with autistic disorder and unrelated siblings. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2012, 30, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Grigorenko, E.L.; Han, S.S.; Yrigollen, C.M.; Leng, L.; McDonald, C.; Mizue, Y.; Anderson, G.M.; Mulder, E.J.; de Bildt, A.; Minderaa, R.B.; et al. Macrophage migration inhibitory factor and stereotypical behavior in autism spectrum disorders. Pediatrics 2008, 122, e438–e445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordeleau, M.; Fernández de Cossío, L.; Chakravarty, M.M.; Tremblay, M. From maternal diet to neurodevelopmental disorders: A story of neuroinflammation. Front. Cell. Neurosci. 2020, 14, 461. [Google Scholar] [CrossRef] [PubMed]
- Jaini, R.; Wolf, M.R.; Yu, Q.; King, A.T.; Frazier, T.W.; Eng, C. Maternal genetics influences fetal neurodevelopment and postnatal autism spectrum disorder-like phenotype by modulating in-utero immunosuppression. Transl. Psychiatry 2021, 11, 348. [Google Scholar] [CrossRef] [PubMed]
- Cuchillo-Ibáñez, I.; Andreo-Lillo, P.; Pastor-Ferrándiz, L.; Carratalá-Marco, F.; Sáez-Valero, J. Elevated plasma reelin levels in children with autism. Front. Psychiatry 2020, 11, 242. [Google Scholar] [CrossRef]
- Hernández-García, I.; Chamorro, A.J.; Ternavasio-de la Vega, H.G.; Carbonell, C.; Marcos, M.; Mirón-Canelo, J.A. Association of allelic variants of the reelin gene with autistic spectrum disorder: A systematic review and meta-analysis of candidate gene association studies. Int. J. Environ. Res. Public Health 2020, 17, 8010. [Google Scholar] [CrossRef]
- Ishii, K.; Kubo, K.; Nakajima, K. Reelin and neuropsychiatric disorders. Front. Cell. Neurosci. 2016, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Hughes, H.K.; Mills Ko, E.; Rose, D.; Ashwood, P. Immune dysfunction and autoimmunity as pathological mechanisms in autism spectrum disorders. Front. Cell. Neurosci. 2018, 12, 405. [Google Scholar] [CrossRef] [Green Version]
- Estes, M.L.; McAllister, A.K. Immune mediators in the brain and peripheral tissues in autism spectrum disorder. Nat. Rev. Neurosci. 2015, 16, 469–486. [Google Scholar] [CrossRef] [Green Version]
- Pardo, C.A.; Farmer, C.A.; Thurm, A.; Shebl, F.M.; Ilieva, J.; Kalra, S.; Swedo, S. Serum and cerebrospinal fluid immune mediators in children with autistic disorder: A longitudinal study. Mol. Autism 2017, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Chua, R.X.Y.; Tay, M.J.Y.; Ooi, D.S.Q.; Siah, K.T.H.; Tham, E.H.; Shek, L.P.C.; Meaney, M.J.; Broekman, B.F.P.; Loo, E.X.L. Understanding the link between allergy and neurodevelopmental disorders: A current review of factors and mechanisms. Front. Neurol. 2020, 11, 1965. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Snetselaar, L.G.; Jing, J.; Liu, B.; Strathearn, L.; Bao, W. Association of food allergy and other allergic conditions with autism spectrum disorder in children. JAMA Netw. Open 2018, 1, e180279. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Karlsson, H.; Dalman, C.; Widman, L.; Rai, D.; Gardner, R.M.; Magnusson, C.; Schendel, D.E.; Newschaffer, C.J.; Lee, B.K. Family history of mental and neurological disorders and risk of autism. JAMA Netw. Open 2019, 2, e190154. [Google Scholar] [CrossRef] [PubMed]
- Sotgiu, S.; Manca, S.; Gagliano, A.; Minutolo, A.; Melis, M.C.; Pisuttu, G.; Scoppola, C.; Bolognesi, E.; Clerici, M.; Guerini, F.R.; et al. Immune regulation of neurodevelopment at the mother-foetus interface: The case of autism. Clin. Transl. Immunol. 2020, 9, e1211. [Google Scholar] [CrossRef]
- Pronovost, G.N.; Hsiao, E.Y. Perinatal interactions between the microbiome, immunity and neurodevelopment. Immunity 2019, 50, 18–36. [Google Scholar] [CrossRef]
- Marks, K.; Coutinho, E.; Vincent, A. Maternal-autoantibody-related (mar) autism: Identifying neuronal antigens and approaching prospects for intervention. J. Clin. Med. 2020, 9, 2564. [Google Scholar] [CrossRef]
- O’Rourke, J.A.; Ravichandran, C.; Howe, Y.J.; Mullett, J.E.; Keary, C.J.; Golas, S.B.; Hureau, A.R.; McCormick, M.; Chung, J.; Rose, N.R.; et al. Accuracy of self-reported history of autoimmune disease: A pilot study. PLoS ONE 2019, 14, e0216526. [Google Scholar] [CrossRef]
- Choi, G.B.; Yim, Y.S.; Wong, H.; Kim, S.; Kim, H.; Kim, S.V.; Hoeffer, C.A.; Littman, D.R.; Huh, J.R. The maternal interleukin-17a pathway in mice promotes autismlike phenotypes in offspring. Science 2016, 351, 933–939. [Google Scholar] [CrossRef] [Green Version]
- Graham, A.M.; Rasmussen, J.M.; Rudolph, M.D.; Heim, C.M.; Gilmore, J.H.; Styner, M.; Potkin, S.G.; Entringer, S.; Wadhwa, P.D.; Fair, D.A.; et al. Maternal systemic interleukin-6 during pregnancy is associated with newborn amygdala phenotypes and subsequent behavior at 2-years-of-age. Biol. Psychiatry 2018, 83, 109–119. [Google Scholar] [CrossRef]
- Rudolph, M.; Graham, A.; Feczko, E.; Miranda-Dominguez, O.; Rasmussen, J.; Nardos, R.; Entringer, S.; Wadhwa, P.; Buss, C.; Fair, D. Maternal il-6 during pregnancy can be estimated from newborn brain connectivity and predicts future working memory in offspring. Nat. Neurosci. 2018, 21, 765–772. [Google Scholar] [CrossRef]
- Zerbo, O.; Leong, A.; Barcellos, L.; Bernal, P.; Fireman, B.; Croen, L.A. Immune mediated conditions in autism spectrum disorders. Brain Behav. Immun. 2015, 46, 232–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, A.L.-A.; Mostafa, G.A. Serum antinucleosome-specific antibody as a marker of autoimmunity in children with autism. J. Neuroinflamm. 2014, 11, 69. [Google Scholar]
- Zerbo, O.; Yoshida, C.; Gunderson, E.P.; Dorward, K.; Croen, L.A. Interpregnancy interval and risk of autism spectrum disorders. Pediatrics 2015, 136, 651–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, A.L.-A.; Mostafa, G.A. Elevated serum levels of interleukin-17a in children with autism. J. Neuroinflamm. 2012, 9, 158. [Google Scholar]
- Casanova, M.F.; Frye, R.E.; Gillberg, C.; Casanova, E.L. Editorial: Comorbidity and autism spectrum disorder. Front. Psychiatry 2020, 1273. [Google Scholar] [CrossRef]
- Rose, D.R.; Yang, H.; Careaga, M.; Angkustsiri, K.; Van de Water, J.; Ashwood, P. T cell populations in children with autism spectrum disorder and co-morbid gastrointestinal symptoms. Brain Behav. Immun.—Health 2020, 2, 100042. [Google Scholar] [CrossRef]
- Ly, A.L.-A.; Mostafa, G.A. Elevated serum levels of macrophage-derived chemokine and thymus and activation-regulated chemokine in autistic children. J. Neuroinflamm. 2013, 10, 72. [Google Scholar]
- Masi, A.; Glozier, N.; Dale, R.; Guastella, A.J. The immune system, cytokines, and biomarkers in autism spectrum disorder. Neurosci. Bull. 2017, 33, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Jyonouchi, H.; Geng, L.; Davidow, A.L. Cytokine profiles by peripheral blood monocytes are associated with changes in behavioral symptoms following immune insults in a subset of asd subjects: An inflammatory subtype? J. Neuroinflamm. 2014, 11, 187. [Google Scholar] [CrossRef] [Green Version]
- Enstrom, A.M.; Onore, C.E.; Van de Water, J.A.; Ashwood, P. Differential monocyte responses to tlr ligands in children with autism spectrum disorders. Brain Behav. Immun. 2009, 24, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Napolioni, V.; Ober-Reynolds, B.; Szelinger, S.; Corneveaux, J.J.; Pawlowski, T.; Ober-Reynolds, S.; Kirwan, J.; Persico, A.M.; Melmed, R.D.; Craig, D.W.; et al. Plasma cytokine profiling in sibling pairs discordant for autism spectrum disorder. J. Neuroinflamm. 2013, 10, 38. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Shen, C.; Li, J.; Yuan, J.; Wei, J.; Huang, F.; Wang, F.; Li, G.; Li, Y.; Xing, L.; et al. Plasma IP-10 and MCP-3 levels are highly associated with disease severity and predict the progression of covid-19. J. Allergy Clin. Immunol. 2020, 146, 119–127.e114. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Matsuzaki, H.; Iwata, K.; Kameno, Y.; Shimmura, C.; Kawai, S.; Yoshihara, Y.; Wakuda, T.; Takebayashi, K.; Takagai, S.; et al. Plasma cytokine profiles in subjects with high-functioning autism spectrum disorders. PLoS ONE 2011, 6, e20470. [Google Scholar] [CrossRef] [PubMed]
- Bryn, V.; Aass, H.C.; Skjeldal, O.H.; Isaksen, J.; Saugstad, O.D.; Ormstad, H. Cytokine profile in autism spectrum disorders in children. J. Mol. Neurosci. 2017, 61, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Pessah, I.; Van de Water, J. Elevated plasma cytokines in autism spectrum disorders provide evidence of immune dysfunction and are associated with impaired behavioral outcome. Brain Behav. Immun. 2010, 25, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Guloksuz, S.A.; Abali, O.; Aktas, C.E.; Bilgic, G.S.; Deniz, G.; Yildirim, A.; Kawikova, I.; Guloksuz, S.; Leckman, J.F. Elevated plasma concentrations of s100 calcium-binding protein b and tumor necrosis factor alpha in children with autism spectrum disorders. Rev. Bras. Psiquiatr. 2017, 39, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.J.; Liu, C.L.; Sang, B.; Zhu, X.M.; Du, Y.J. The combined role of serotonin and interleukin-6 as biomarker for autism. Neuroscience 2015, 284, 290–296. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Marler, S.; Altstein, L.L.; Lee, E.B.; Mazurek, M.O.; McLaughlin, A.; Macklin, E.A.; McDonnell, E.; Davis, D.J.; Belenchia, A.M.; et al. Associations between cytokines, endocrine stress response, and gastrointestinal symptoms in autism spectrum disorder. Brain Behav. Immun. 2016, 58, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Ashwood, P.; Enstrom, A.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.L.; Croen, L.A.; Ozonoff, S.; Pessah, I.N.; Van de Water, J. Decreased transforming growth factor beta1 in autism: A potential link between immune dysregulation and impairment in clinical behavioral outcomes. J. Neuroimmunol. 2008, 204, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Makinodan, M.; Iwata, K.; Ikawa, D.; Yamashita, Y.; Yamamuro, K.; Toritsuka, M.; Kimoto, S.; Okumura, K.; Yamauchi, T.; Yoshino, H.; et al. Tumor necrosis factor-alpha expression in peripheral blood mononuclear cells correlates with early childhood social interaction in autism spectrum disorder. Neurochem. Int. 2017, 104, 1–5. [Google Scholar] [CrossRef]
- Han, Y.M.; Cheung, W.K.; Wong, C.K.; Sze, S.L.; Cheng, T.W.; Yeung, M.K.; Chan, A.S. Distinct cytokine and chemokine profiles in autism spectrum disorders. Front. Immunol. 2017, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libbey, J.E.; Coon, H.H.; Kirkman, N.J.; Sweeten, T.L.; Miller, J.N.; Stevenson, E.K.; Lainhart, J.E.; McMahon, W.M.; Fujinami, R.S. Are there enhanced mbp autoantibodies in autism? J. Autism Dev. Disord. 2008, 38, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Inga Jácome, M.C.; Morales Chacòn, L.M.; Vera Cuesta, H.; Maragoto Rizo, C.; Whilby Santiesteban, M.; Ramos Hernandez, L.; Noris García, E.; González Fraguela, M.E.; Fernandez Verdecia, C.I.; Vegas Hurtado, Y.; et al. Peripheral inflammatory markers contributing to comorbidities in autism. Behav. Sci. 2016, 6, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, N.K.; Sweeten, T.L.; McMahon, W.M.; Fujinami, R.S. Hyperserotoninemia and altered immunity in autism. J. Autism Dev. Disord. 2006, 36, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Sink, V.K. Phenotypic expression of autoimmune disorder (aad): A major subset of autism. Ann. Clin. Psychiatry 2009, 21, 148–161. [Google Scholar]
- Sink, V.K.; Lin, S.X.; Newell, E.; Nelson, C. Abnormal measles-munps-rubella antiboodies and CNS autoimmunity in children with autism. J. Biomed. Sci. 2000, 9, 359–364. [Google Scholar]
- Singh, V.K.; Lin, S.X.; Yang, V.C. Serological association of measles virus and humanherpesvirus-6 with brain autoantibodies in autism. Clin. Immunol. Immunopathol. 1998, 89, 105–108. [Google Scholar] [CrossRef]
- Wasilewska, J.; Kaczmarski, M.; Stasiak-Barmuta, A.; Tobolczyk, J.; Kowalewska, E. Low serum iga and increased expression of cd23 on b lymphocytes in peripheral blood in children with regressive autism aged 3–6 years old. Arch. Med. Sci. AMS 2012, 8, 324–331. [Google Scholar] [CrossRef]
- Heuer, L.S.; Rose, M.; Ashwood, P.; Van de Water, J. Decreased levels of total immunoglobulin in children with autism is not a result of b cell dysfunction. J. Neuroimmunol. 2012, 251, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, D.A.; Frye, R.E. A systematic review and meta-analysis of immunoglobulin G abnormalities and the therapeutic use of intravenous immunoglobulins (ivig) in autism spectrum disorder. J. Pers. Med. 2021, 11, 488. [Google Scholar] [CrossRef]
- Heuer, L.; Ashwood, P.; Schauer, J.; Goines, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Croen, L.A.; Pessah, I.N.; Van de Water, J. Reduced levels of immunoglobulin in children with autism correlates with behavioral symptoms. Autism Res. 2008, 1, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Spiroski, M.; Trajkovski, V.; Trajkov, D.; Petlichkovski, A.; Efinska-Mladenovska, O.; Hristomanova, S.; Djulejic, E.; Paneva, M.; Bozhikov, J. Family analysis of immunoglobulin classes and subclasses in children with autistic disorder. Bosn. J. Basic Med. Sci. 2009, 9, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piras, I.; Haapanen, L.; Napolioni, V.; Sacco, R.; Van de Water, J.; Persico, A. Anti-brain antibodies are associated with more severe cognitive and behavioral profiles in italian children with autism spectrum disorder. Brain Behav. Immun. 2014, 38, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, G.A.; Al-Ayadhi, L.Y. Increased serum levels of anti-ganglioside m1 autoantibodies in autistic children: Relation to the disease severity. J. Neuroinflamm. 2011, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Bobrowski-khoury, N.; Ramaekers, V.; Sequeira, J.M.; Quadros, E.V. Folate receptor alpha autoantibodies in autism spectrum disorders diagnosis, treatment and prevention. J. Pers. Med. 2021, 11, 710. [Google Scholar] [CrossRef]
- Racicot, K.; Kwon, J.Y.; Aldo, P.; Silasi, M.; Mor, G. Understanding the complexity of the immune system during pregnancy. Am. J. Reprod. Immunol. 2014, 72, 107–116. [Google Scholar] [CrossRef]
- Mostafa, G.A.; Al-Ayadhi, L.Y. The relationship between the increased frequency of serum antineuronal antibodies and the severity of autism in children. Eur. J. Paediatr. Neurol. 2012, 16, 464–468. [Google Scholar] [CrossRef]
- Kusunoki, S.; Kaida, K. Antibodies against ganglioside complexes in Guillain-Barre syndrome and related disorders. J. Neurochem. 2011, 116, 828–832. [Google Scholar] [CrossRef]
- Careaga, M.; Hansen, R.L.; Hertz-Piccotto, I.; Van De Water, J.; Ashwood, P. Increased anti-phospholipid antibodies in autism spectrum disorders. Mediat. Inflamm. 2013, 2013, 935608. [Google Scholar] [CrossRef]
- Mostafa, G.A.; Al-Ayadhi, L.Y. The possible relationship between allergic manifestations and elevated serum levels of brain specific autoantibodies in autistic children. J. Neuroimmunol. 2013, 261, 77–81. [Google Scholar] [CrossRef]
- Mostafa, G.A.; El-Sherif, D.F.; Al-Ayadhi, L.Y. Systemic autoantibodies in children with autism. J. Neuroimmunol. 2014, 272, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Angelidou, A.; Alysandratos, K.D.; Vasiadi, M.; Francis, K.; Asadi, S.; Theoharides, A.; Sideri, K.; Lykouras, L.; Kalogeromitros, D.; et al. Mitochondrial DNA and anti-mitochondrial antibodies in serum of autistic children. J. Neuroinflamm. 2010, 7, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frye, R.E.; Sequeira, J.M.; Quadros, E.V.; James, S.J.; Rossignol, D.A. Cerebral folate receptor autoantibodies in autism spectrum disorder. Mol. Psychiatry 2013, 18, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Ramaekers, V.T.; Quadros, E.V.; Sequeira, J.M. Role of folate receptor autoantibodies in infantile autism. Mol. Psychiatry 2013, 18, 270–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldo, P.B.; Racicot, K.; Craviero, V.; Guller, S.; Romero, R.; Mor, G. Trophoblast induces monocyte differentiation into CD14+/CD16+ macrophages. Am. J. Reprod. Immunol. 2014, 72, 270–284. [Google Scholar] [CrossRef] [Green Version]
- Braunschweig, D.; Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Croen, L.A.; Pessah, I.N.; Van de Water, J. Autism: Maternally derived antibodies specific for fetal brain proteins. Neurotoxicology 2008, 29, 226–231. [Google Scholar] [CrossRef] [Green Version]
- Dekel, N.; Gnainsky, Y.; Granot, I.; Racicot, K.; Mor, G. The role of inflammation for a successful implantation. Am. J. Reprod. Immunol. 2014, 72, 141–147. [Google Scholar] [CrossRef]
- Albrecht, M.; Arck, P.C. Vertically transferred immunity in neonates: Mothers, mechanisms and mediators. Front. Immunol. 2020, 11, 555. [Google Scholar] [CrossRef]
- Ghosh, M.K.; Muller, H.K.; Walker, A.M. Lactation-based maternal educational immunity crosses mhc class i barriers and can impart th1 immunity to th2-biased recipients. J. Immunol. 2017, 199, 1729–1736. [Google Scholar] [CrossRef] [Green Version]
- Mor, G.; Cardenas, I. The immune system in pregnancy: A unique complexity. Am. J. Reprod. Immunol. 2010, 63, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Edmiston, E.; Ashwood, P.; Van de Water, J. Autoimmunity, autoantibodies, and autism spectrum disorders (asd). Biol. Psychiatry 2017, 81, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clements, T.; Rice, T.F.; Vamvakas, G.; Barnett, S.; Barnes, M.; Donaldson, B.; Jones, C.E.; Kampmann, B.; Holder, B. Update on transplacental transfer of IgG subclasses: Impact of maternal and fetal factors. Front. Immunol. 2020, 11, 1920. [Google Scholar] [CrossRef] [PubMed]
- Gata-Garcia, A.; Diamond, B. Maternal antibody and asd: Clinical data and animal models. Front. Immunol. 2019, 10, 1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onore, C.; Enstrom, A.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Van de Water, J.; Ashwood, P. Decreased cellular il-23 but not il-17 production in children with autism spectrum disorders. J. Neuroimmunol. 2009, 216, 126–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jyonouchi, H.; Geng, L.; Cushing-Ruby, A.; Quraishi, H. Impact of innate immunity in a subset of children with autism spectrum disorders: A case control study. J Neuroinflam. 2008, 5, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Ding, Y.; Wu, F.; Li, R.; Xie, G.; Hou, J.; Mao, P. Family history of autoimmune diseases is associated with an increased risk of autism in children: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2015, 55, 322–332. [Google Scholar] [CrossRef]
- Martin, L.A.; Ashwood, P.; Braunschweig, D.; Cabanlit, M.; Van De Water, J.; Amaral, D.G. Stereotypies and hyperactivity in rhesus monkeys; exposed to IgG from mothers of children with autism. Brain Behav. Immun. 2008, 22, 806–816. [Google Scholar] [CrossRef] [Green Version]
- Braunschweig, D.; Golub, M.S.; Koenig, C.M.; Qi, L.; Pessah, I.N.; Van de Water, J.; Berman, R.F. Maternal autism-associated IgG antibodies delay developmentand produce anxiety in a mouse gestational transfer model. J. Neuroimmunol. 2012, 252, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Bauman, M.D.; Iosif, A.-M.; Smith, S.E.P.; Bregere, C.; Amaral, D.G.; Patterson, P.H. Activation of the maternal immune system during pregnancy alters behavioral development of rhesus monkeyoffspring. Biol. Psychiatry 2014, 75, 332–341. [Google Scholar] [CrossRef] [Green Version]
- Atladóttir, H.Ó.; Thorsen, P.; Østergaard, L.; Schendel, D.E.; Lemcke, S.; Abdallah, M.; Parner, E.T. Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders. J. Autism Dev. Disord. 2010, 40, 1423–1430. [Google Scholar] [CrossRef]
- Zerbo, O.; Iosif, A.M.; Walker, C.; Ozonoff, S.; Hansen, R.L.; Hertz-Picciotto, I. Is maternal influenza or fever during pregnancy associated with autism or developmental delays? Results from the CHARGE (Childhood Autism Risks from Genetics and Environment) study. J. Autism Dev. Disord. 2013, 43, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural killer cells: Development, maturation, and clinical utilization. Front. Immunol. 2018, 9, 1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucuksezer, U.C.; Aktas Cetin, E.; Esen, F.; Tahrali, I.; Akdeniz, N.; Gelmez, M.Y.; Deniz, G. The role of natural killer cells in autoimmune diseases. Front. Immunol. 2021, 12, 79. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Tian, Z.; Wei, H. Developmental and functional control of natural killer cells by cytokines. Front. Immunol. 2017, 8, 930. [Google Scholar] [CrossRef]
- Gładysz, D.; Krzywdzińska, A.; Hozyasz, K.K. Immune abnormalities in autism spectrum disorder-could they hold promise for causative treatment? Mol. Neurobiol. 2018, 55, 6387–6435. [Google Scholar] [CrossRef] [Green Version]
- Tye, C.; Runicles, A.K.; Whitehouse, A.J.O.; Alvares, G.A. Characterizing the interplay between autism spectrum disorder and comorbid medical conditions: An integrative review. Front. Psychiatry 2018, 9, 751. [Google Scholar] [CrossRef] [Green Version]
- Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Pessah, I.N.; Van de Water, J. Altered T cell responses in children with autism. Brain Behav. Immun. 2011, 25, 840–849. [Google Scholar] [CrossRef] [Green Version]
- Edilova, M.I.; Akram, A.; Abdul-Sater, A.A. Innate immunity drives pathogenesis of rheumatoid arthritis. Biomed. J. 2021, 44, 172–182. [Google Scholar] [CrossRef]
- Laria, A.; Lurati, A.; Marrazza, M.; Mazzocchi, D.; Re, K.A.; Scarpellini, M. The macrophages in rheumatic diseases. J. Inflamm. Res. 2016, 9, 1. [Google Scholar]
- Nagafuchi, Y.; Shoda, H.; Sumitomo, S.; Nakachi, S.; Kato, R.; Tsuchida, Y.; Tsuchiya, H.; Sakurai, K.; Hanata, N.; Tateishi, S.; et al. Immunophenotyping of rheumatoid arthritis reveals a linkage between hla-drb1 genotype, cxcr4 expression on memory CD4+ T cells, and disease activity. Sci. Rep. 2016, 6, 29338. [Google Scholar] [CrossRef] [Green Version]
- Takai, C.; Matsumoto, I.; Inoue, A.; Umeda, N.; Tanaka, Y.; Kurashima, Y.; Wada, Y.; Narita, I.; Sumida, T. Specific overexpression of tumour necrosis factor-α-induced protein (tnfaip)9 in CD14+CD16− monocytes in patients with rheumatoid arthritis: Comparative analysis with tnfaip3. Clin. Exp. Immunol. 2015, 180, 458–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grozdanov, V.; Bliederhaeuser, C.; Ruf, W.P.; Roth, V.; Fundel-Clemens, K.; Zondler, L.; Brenner, D.; Martin-Villalba, A.; Hengerer, B.; Kassubek, J.; et al. Inflammatory dysregulation of blood monocytes in parkinson’s disease patients. Acta Neuropathol. 2014, 128, 651–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier, L.P.; York, E.M.; Kamyabi, A.; Choi, H.B.; Weilinger, N.L.; MacVicar, B.A. Microglial metabolic flexibility supports immune surveillance of the brain parenchyma. Nat. Commun. 2020, 11, 1559. [Google Scholar] [CrossRef] [PubMed]
- Eyo, U.B.; Wu, L.J. Microglia: Lifelong patrolling immune cells of the brain. Prog. Neurobiol. 2019, 179, 101614. [Google Scholar] [CrossRef]
- Bronzuoli, M.R.; Facchinetti, R.; Ingrassia, D.; Sarvadio, M.; Schiavi, S.; Steardo, L.; Verkhratsky, A.; Trezza, V.; Scuderi, C. Neuroglia in the autistic brain: Evidence from a preclinical model. Mol. Autism 2018, 9, 66. [Google Scholar] [CrossRef]
- Maezawa, I.; Calafiore, M.; Wulff, H.; Jin, L.W. Does microglial dysfunction play a role in autism and rett syndrome? Neuron Glia Biol. 2011, 7, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.I.; Kern, J.K. Evidence of microglial activation in autism and its possible role in brain underconnectivity. Neuron Glia Biol. 2011, 7, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Tsilioni, I.; Pantazopoulos, H.; Conti, P.; Leeman, S.E.; Theoharides, T.C. Il-38 inhibits microglial inflammatory mediators and is decreased in amygdala of children with autism spectrum disorder. Proc. Natl. Acad. Sci. USA 2020, 117, 16475–16480. [Google Scholar] [CrossRef]
- Dean, D.C.; Travers, B.G.; Adluru, N.; Tromp, D.P.; Destiche, D.J.; Samsin, D.; Prigge, M.B.; Zielinski, B.A.; Fletcher, P.T.; Anderson, J.S.; et al. Investigating the microstructural correlation of white matter in autism spectrum disorder. Brain Connect. 2016, 6, 415–433. [Google Scholar] [CrossRef] [Green Version]
- Lenz, K.M.; Nelson, L.H. Microglia and beyond: Innate immune cells as regulators of brain development and behavioral function. Front. Immunol. 2018, 9, 698. [Google Scholar] [CrossRef] [Green Version]
- Michell-Robinson, M.A.; Touil, H.; Healy, L.M.; Owen, D.R.; Durafourt, B.A.; Bar-Or, A.; Antel, J.P.; Moore, C.S. Roles of microglia in brain development, tissue maintenance and repair. Brain 2015, 138, 1138–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Choi, J.; Yoon, B.E. Neuron-glia interactions in neurodevelopmental disorders. Cells 2020, 9, 2176. [Google Scholar] [CrossRef] [PubMed]
- Haruwaka, K.; Ikegami, A.; Tachibana, Y.; Ohno, N.; Konishi, H.; Hashimoto, A.; Matsumoto, M.; Kato, D.; Ono, R.; Kiyama, H.; et al. Dual microglia effects on blood brain barrier permeability induced by systemic inflammation. Nat. Commun. 2019, 10, 5816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davoli-Ferreira, M.; Thomson, C.A.; McCoy, K.D. Microbiota and microglia interactions in asd. Front. Immunol. 2021, 12, 2024. [Google Scholar] [CrossRef]
- Ashwood, P.; Corbett, B.A.; Kantor, A.; Schulman, H.; Van de Water, J.; Amaral, D.G. In search of cellular immunophenotypes in the blood of children with autism. PLoS ONE 2011, 6, e19299. [Google Scholar] [CrossRef] [Green Version]
- Depino, A.M.; Robinson-Agramonte, M.A. Understanding on Neuroimmunology in Autism Spectrum Disorder; Robinson-Agramonte, M.A., Ed.; Translational Approaches to Autism Spectrum Disorder; Springer International Publishing: Cham, Switzerland, 2015; Chapter 9; pp. 155–180. [Google Scholar] [CrossRef]
- Jones, K.L.; Croen, L.A.; Yoshida, C.K.; Heuer, L.; Hansen, R.; Zerbo, O.; DeLorenze, G.N.; Kharrazi, M.; Yolken, R.; Ashwood, P.; et al. Autism with intellectual disability is associated with increased levels of maternal cytokines and chemokines during gestation. Mol. Psychiatry 2017, 22, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Sharon, G.; Cruz, N.J.; Kang, D.W.; Gandal, M.J.; Wang, B.; Kim, Y.M.; Zink, E.M.; Casey, C.P.; Taylor, B.C.; Lane, C.J.; et al. Human gut microbiota from autism spectrum disorder promote behavioral symptoms in mice. Cell 2019, 177, 1600–1618.e17. [Google Scholar] [CrossRef] [Green Version]
- Goines, P.; Van de Water, J. The immune system’s role in the biology of autism. Curr. Opin. Neurol. 2010, 23, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Masi, A.; Breen, E.J.; Alvares, G.A.; Glozier, N.; Hickie, I.B.; Hunt, A.; Hui, J.; Beilby, J.; Ravine, D.; Wray, J.; et al. Cytokine levels and associations with symptom severity in male and female children with autism spectrum disorder. Mol. Autism 2017, 8, 63. [Google Scholar] [CrossRef]
- Schwartzer, J.J.; Careaga, M.; Coburn, M.A.; Rose, D.R.; Hughes, H.K.; Ashwood, P. Behavioral impact of maternal allergic-asthma in two genetically distinct mouse strains. Brain Behav. Immun. 2017, 63, 99–107. [Google Scholar] [CrossRef]
- Volk, H.E.; Park, B.; Hollingue, C.; Jones, K.L.; Ashwood, P.; Windham, G.C.; Lurman, F.; Alexeeff, S.E.; Kharrazi, M.; Pearl, M.; et al. Maternal immune response and air pollution exposure during pregnancy: Insights from the early markers for autism (ema) study. J. Neurodev. Disord. 2020, 12, 42. [Google Scholar] [CrossRef] [PubMed]
- Dunaevsky, A.; Bergdolt, L. Brain changes in a maternal immune activation model of neurodevelopmental brain disorders. Prog. Neurobiol. 2019, 175, 1–19. [Google Scholar]
- Gustafsson, H.C.; Sullivan, E.L.; Nousen, E.K.; Sullivan, C.A.; Huang, E.; Rincon, M.; Nigg, J.T.; Loftis, J.M. Maternal prenatal depression predicts infant negative affect via maternal inflammatory cytokine levels. Brain Behav. Immun. 2018, 73, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Kentner, A.C.; Bilbo, S.D.; Brown, A.S.; Hsiao, E.Y.; McAllister, A.K.; Meyer, U.; Pearce, B.D.; Pletnikov, M.V.; Yolken, R.H.; Bauman, M.D. Maternal immune activation: Reporting guidelines to improve the rigor, reproducibility, and transparency of the model. Neuropsychopharmacology 2019, 44, 245–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, H.; Hoeffer, C. Maternal il-17a in autism. Exp. Neurol. 2018, 299, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Southcombe, J.H.; Redman, C.W.G.; Sargent, I.L.; Granne, I. Interleukin-1 family cytokines and their regulatory proteins in normal pregnancy and pre-eclampsia. Clin. Exp. Immunol. 2015, 181, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Grev, J.; Berg, M.; Soll, R. Maternal probiotic supplementation for prevention of morbidity and mortality in preterm infants. Cochrane Database Syst. Rev. 2018, 2018, 1465–1858. [Google Scholar] [CrossRef]
- Ravaccia, D.; Ghafourian, T. Critical role of the maternal immune system in the pathogenesis of autism spectrum disorder. Biomedicines 2020, 8, 557. [Google Scholar] [CrossRef]
- Beversdorf, D.Q. Phenotyping, etiological factors, and biomarkers: Toward precision medicine in autism spectrum disorders. J. Dev. Behav. Pediatrics 2016, 37, 659–673. [Google Scholar] [CrossRef] [Green Version]
- Bölte, S.; Girdler, S.; Marschik, P.B. The contribution of environmental exposure to the etiology of autism spectrum disorder. Cell. Mol. Life Sci. 2019, 76, 1275–1297. [Google Scholar] [CrossRef] [Green Version]
- Didion, S.P. Cellular and oxidative mechanisms associated with interleukin-6 signaling in the vasculature. Int. J. Mol. Sci. 2017, 18, 2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose-John, S. Interleukin-6 signalling in health and disease. F1000Research 2020, 9, 1013. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Overview of the il-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Pyrillou, K.; Burzynski, L.C.; Clarke, M.C.H. Alternative pathways of il-1 activation, and its role in health and disease. Front. Immunol. 2020, 11, 3288. [Google Scholar] [CrossRef] [PubMed]
- Ting, E.Y.C.; Yang, A.C.; Tsai, S.J. Role of interleukin-6 in depressive disorder. Int. J. Mol. Sci. 2020, 21, 2194. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.L.; Hsiao, E.Y.; Yan, Z.; Mazmanian, S.K.; Patterson, P.H. The placental interleukin-6 signaling controls fetal brain development and behavior. Brain Behav. Immun. 2017, 62, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, S.L.; Porter, K.; Christian, L.M. Adaptation of the inflammatory immune response across pregnancy and postpartum in Black and White women. J. Reprod. Immunol. 2016, 114, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Pinto-Martin, J.A.; Levy, S.E.; Feldman, J.F.; Lorenz, J.M.; Paneth, N.; Whitaker, A.H. Prevalence of autism spectrum disorder in adolescents born weighing <2000 grams. Pediatrics 2011, 128, 2010–2016. [Google Scholar] [CrossRef] [Green Version]
- Patterson, P.H. Maternal infections and immune involvement in autism. Trends Mol. Med. 2011, 17, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Steinman, G. Predicting autism at birth. Med. Hypotheses 2013, 81, 21–25. [Google Scholar] [CrossRef]
- Bianchi, V.E.; Locatelli, V.; Rissi, L. Neurotrophic and neuro-regenerative efforts of GH/IGF1. Int. J. Mol. Sci. 2017, 18, 2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinman, G. The putative etiology and prevention of autism. In Progress in Molecular Biology and Translational Science; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 173, ISSN 1877-1173. [Google Scholar]
- Janušonis, S. Statistical distribution of blood serotonin as a predictor of early autistic brain abnormalities. Theor. Biol. Med. Model. 2005, 2, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janušonis, S. Origin of the blood hyperserotonemia of autism. Theor. Biol. Med. Model. 2008, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frye, R.E.; Vassall, S.; Kaur, G.; Lewis, C.; Karim, M.; Rossignol, D. Emerging biomarkers in autism spectrum disorder: A systematic review. Ann. Transl. Med. 2019, 7, 792. [Google Scholar] [CrossRef] [PubMed]
- Coleman, D.M.; Adams, J.B.; Anderson, A.L.; Frye, R.E. Rating of the effectiveness of 26 psychiatric and seizure medications for autism spectrum disorder: Results of a national survey. J. Child Adolesc. Psychopharmacol. 2019, 29, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Al-Mazidi, S.; Al-Ayadhi, L.Y. Plasma levels of alpha and gamma synucleins in autism spectrum disorder: An indicator of severity. Med. Princ. Pract. 2021, 30, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zang, H.; Liu, S.; Luo, W.; Jiang, Y.; Gao, J. Association of peripheral blood levels of cytokines with autism spectrum disorder: A meta-analysis. Front. Psychiatry 2021, 12, 1006. [Google Scholar] [CrossRef]
- Ashwood, P.; Van de Water, J. A review of autism and the immune response. Clin. Dev. Immunol. 2004, 11, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Enstrom, A.; Onore, C.; Hertz-Picciotto, I.; Hansen, R.; Croen, L.; Van de Water, J.; Ashwood, P. Detection of il-17 and il-23 in plasma samples of children with autism. Am. J. Biochem. Biotechnol. 2008, 4, 114–120. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Marler, S.; Altstein, L.L.; Lee, E.B.; Akers, J.; Sohl, K.; McLaughlin, A.; Hartnett, K.; Kille, B.; Mazurek, M.; et al. Psychophysiological associations with gastrointestinal symptomatology in autism spectrum disorder. Autism Res. Off. J. Int. Soc. Autism Res. 2017, 10, 276–288. [Google Scholar] [CrossRef]
- Burton, P.B.; Quirke, P.; Sorensen, C.M.; Nehlsen-Cannarella, S.L.; Bailey, L.L.; Knight, D.E. Growth factor expression during rat development: A comparison of tgf-beta 3, tgf-alpha, bfgf, pdgf and pdgf-r. Int. J. Exp. Pathol. 1993, 74, 87–96. [Google Scholar] [PubMed]
- Galbiati, M.; Crippa, V.; Rusmini, P.; Cristofani, R.; Messi, E.; Piccolella, M.; Tedesco, B.; Ferrari, V.; Casarotto, E.; Chierichetti, M.; et al. Multiple roles of transforming growth factor beta in amyotrophic lateral sclerosis. Int. J. Mol. Sci. 2020, 21, 4291. [Google Scholar] [CrossRef] [PubMed]
- Pandiyan, P.; Zhu, J. Origin and functions of pro-inflammatory cytokine producing foxp3+ regulatory T cells. Cytokine 2015, 76, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partridge, M.; Green, M.R.; Langdin, J.D.; Feldmann, M. Production of transforming growth factor beta by human t lymphocytes and its potential role in the regulation of T cell growth. J. Exp. Med. 1986, 163, 1037–1050. [Google Scholar]
- Choi, J.E.; Widjaja, F.; Careaga, M.; Bent, S.; Ashwood, P.; Hendren, R.L. Change in plasma cytokine levels during risperidone treatment in children with autism. J. Child Adolesc. Psychopharmacol. 2014, 24, 586–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohja, K.; Gozal, E.; Fahnestock, M.; Cai, L.; Cai, J.; Freedman, J.H.; Switala, A.; El-Baz, A.; Barnes, G.N. Neuroimmunologic and neurotrophic interactions in autism spectrum disorders: Relationship to neuroinflammation. Neuromolecular Med. 2018, 20, 161–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.; Niederkorn, J.Y. Transforming growth factor-beta down-regulates major histocompatibility complex class i antigen expression and increases the susceptibility of uveal melanoma cells to natural killer cell-mediated cytolysis. Immunology 1995, 86, 263–269. [Google Scholar] [PubMed]
- Dobolyi, A.; Vincze, C.; Pál, G.; Lovas, G. The neuroprotective functions of transforming growth factor beta proteins. Int. J. Mol. Sci. 2012, 13, 8219–8258. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, P.; Wakefield, A.J. Immune activation of peripheral blood and mucosal CD3+ lymphocyte cytokine profiles in children with autism and gastrointestinal symptoms. J. Neuroimmunol. 2006, 173, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Barcia, G.; Posar, A.; Santucci, M.; Parmeggiani, A. Autism and celiac disease. J. Autism Dev. Disord. 2008, 38, 407–408. [Google Scholar] [CrossRef] [PubMed]
- Garcia Garcìa, R.J.; Robinson-Agramonte, M.A. Comorbilidad en pacientes con trastornos del neurodesarollo. Rev. Cub. Pediatr. 2020, 92, e1108. [Google Scholar]
- Mostafa, G.A.; Hamza, R.T.; El-Shahawi, H.H. Allergic manifestations in autistic children: Relation to disease severity. J. Pediatr. Neurol. 2008, 6, 115–123. [Google Scholar]
- Shibata, A.; Hitomi, Y.; Kambayashi, Y.; Hibino, Y.; Yamazaki, M.; Mitoma, J.; Asakura, H.; Hayashi, K.; Otaki, N.; Sagara, T.; et al. Epidemiological study on the involvements of environmental factors and allergy in child mental health using the Autism Screening Questionnaire. Res. Autism Spectr. Disord. 2013, 7, 132–140. [Google Scholar] [CrossRef]
- Kotey, S.; Ertel, K.; Whitcomb, B. Co-occurrence of autism and asthma in a nationally-representative sample of children in the United States. J. Autism Dev. Disord. 2014, 44, 3083–3088. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.H.; Su, T.P.; Chen, Y.S.; Hsu, J.W.; Huang, K.L.; Chang, W.H.; Chen, T.J.; Bai, Y.M. Comorbidity of allergic and autoimmune diseases in patients with autism spectrum disorder: A nationwide population-based study. Res. Autism Spectr. Disord. 2013, 7, 205–212. [Google Scholar] [CrossRef]
- Buie, T.; Campbell, D.B.; Fuchs, G.J.; Furuta, G.T.; Levy, J.; Van de Water, J.; Whitaker, A.H.; Atkins, D.; Bauman, M.L.; Beaudet, A.L.; et al. Evaluation, diagnosis, and treatment of gastrointestinal disorders in individuals with ASDs: A consensus report. Pediatrics 2010, 125 (Suppl. 1), S1–S18. [Google Scholar] [CrossRef] [Green Version]
- Jyonouchi, H.; Geng, L.; Ruby, A.; Zimmerman-Bier, B. Dysregulated innate immune responses in young children with autism spectrum disorders: Their relationship to gastrointestinal symptoms and dietary intervention. Neuropsychobiology 2005, 51, 77–85. [Google Scholar] [CrossRef]
- Ludvigsson, J.F.; Reichenberg, A.; Hultman, C.M.; Murray, J.A. A nationwide study of the association between celiac disease and the risk of autistic spectrum disorders. JAMA Psychiatry 2013, 70, 1224–1230. [Google Scholar] [CrossRef] [Green Version]
- Nazeen, S.; Palmer, N.P.; Berger, B.; Kohane, I.S. Integrative analysis of genetic data sets reveals a shared innate immune component in autism spectrum disorder and its co-morbidities. Genome Biol. 2016, 17, 228. [Google Scholar] [CrossRef]
- Mcelhanon, B.O.; Mccracken, C.; Karpen, S.; Sharp, W.G. Gastrointestinal symptoms in autism spectrum disorder: A meta-analysis. Pediatrics 2014, 133, 872–883. [Google Scholar] [CrossRef] [Green Version]
- Ashwood, P.; Anthony, A.; Pellicer, A.A.; Torrente, F.; Walker-Smith, J.A.; Wakefield, A.J. Intestinal lymphocyte populations in children with regressive autism: Evidence for extensive mucosal immunopathology. J. Clin. Immunol. 2003, 23, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, P.; Anthony, A.; Torrente, F.; Wakefield, A.J. Spontaneous mucosal lymphocyte cytokine profiles in children with autism and gastrointestinal symptoms: Mucosal immune activation and reduced counter regulatory interleukin-10. J. Clin. Immunol. 2004, 24, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Chaidez, V.; Hansen, R.L.; Hertz-Picciotto, I. Gastrointestinal problems in children with autism, developmental delays or typical development. J. Autism Dev. Disord. 2014, 44, 1117–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrente, F.; Ashwood, P.; Day, R.; Machado, N.; Furlano, R.I.; Anthony, A.; Davies, S.E.; Wakefield, A.J.; Thomson, M.A.; Walker-Smith, J.A.; et al. Small intestinal enteropathy with epithelial IgG and complement deposition in children with regressive autism. Mol. Psychiatry 2002, 7, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, S.J.; Fortunato, J.; Gonzalez, L.G.; Krigsman, A. Identification of unique gene expression profile in children with regressive Autism Spectrum Disorder (ASD) and ileocolitis. PLoS ONE 2013, 8, e58058. [Google Scholar] [CrossRef]
- Rose, D.R.; Yang, H.; Serena, G.; Sturgeon, C.; Ma, B.; Careaga, M.; Hughes, H.K.; Angkustsiri, K.; Rose, M.; Hertz-Picciotto, I.; et al. Differential immune responses and microbiota profiles in children with autism spectrum disorders and co-morbid gastrointestinal symptoms. Brain Behav. Immun. 2018, 70, 354–368. [Google Scholar] [CrossRef]
- Tomova, A.; Husarova, V.; Lakatosova, S.; Bakos, J.; Vlkova, B.; Babinska, K.; Ostatnikova, D. Gastrointestinal microbiota in children with autismin Slovakia. Physiol. Behav. 2015, 138, 179–187. [Google Scholar] [CrossRef]
- Strati, F.; Cavalieri, D.; Albanese, D.; De Felice, C.; Donati, C.; Hayek, J.; Jousson, O.; Leoncini, S.; Renzi, D.; Calabrò, A.; et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome 2017, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Hughes, H.K.; Rose, D.; Ashwood, P. The gut microbiota and dysbiosis in Autism spectrum disorders. Curr. Neurol. Neurosci. Rep. 2018, 18, 81. [Google Scholar] [CrossRef]
- Coury, D.L.; Ashwood, P.; Fasano, A.; Fuchs, G.; Geraghty, M.; Kaul, A.; Mawe, G.; Patterson, P.; Jones, N.E. Gastrointestinal conditions in children with autism spectrum disorder: Developing a research agenda. Pediatrics 2012, 130 (Suppl. 2), S160–S168. [Google Scholar] [CrossRef] [Green Version]
- Fiorentino, M.; Sapone, A.; Senger, S.; Camhi, S.S.; Kadzielski, S.M.; Buie, T.M.; Kelly, D.L.; Cascella, N.; Fasano, A. Blood-brain barrier and intestinal epithelial barrier alterations in autism spectrum disorders. Mol. Autism 2016, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damier, P.; Hirsch, E.C.; Zhang, P.; Agid, Y.; Javoy-Agid, F. Glutathione peroxidase, glial cells and Parkinson’s disease. Neuroscience 1993, 52, 1–6. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Lombardo, M.V.; Baron-Cohen, S. Autism. Lancet 2014, 383, 896–910. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A.H.V. Parkinson Disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef]
- Han, J.W.; Ahn, Y.D.; Kim, W.S.; Shin, C.M.; Jeong, S.J.; Song, Y.S.; Bae, Y.J.; Kim, J.M. Psychiatric Manifestation in Patients with Parkinson’s Disease. J. Korean Med. Sci. 2018, 33, e300. [Google Scholar] [CrossRef]
- Matsuda, W.; Furuta, T.; Nakamura, K.C.; Hioki, H.; Fujiyama, F.; Arai, R.; Kaneko, T. Single nigrostriatal dopaminergic neurons form widely spread and highly dense axonal arborizations in the neostriatum. J. Neurosci. 2009, 29, 444–453. [Google Scholar] [CrossRef] [Green Version]
- Amaral, D.G.; Schumann, C.M.; Nordahl, C.W. Neuroanatomy of autism. Trends Neurosci. 2008, 31, 137–145. [Google Scholar] [CrossRef]
- Fineberg, N.A.; Potenza, M.N.; Chamberlain, S.R.; Berlin, H.A.; Menzies, L.; Bechara, A.; Sahakian, B.J.; Robbins, T.W.; Bullmore, E.T.; Hollander, E. Probing compulsive and impulsive behaviors, from animal models to endophenotypes: A narrative review. Neuropsychopharmacology 2010, 35, 591–604. [Google Scholar] [CrossRef] [Green Version]
- Wijnhoven, L.A.; Niels-Kessels, H.; Creemers, D.H.; Vermulst, A.A.; Otten, R.; Engels, R.C. Prevalence of comorbid depressive symptoms and suicidal ideation in children with autism spectrum disorder and elevated anxiety symptoms. J. Child Adolesc. Ment. Health 2019, 31, 77–84. [Google Scholar] [CrossRef]
- Chuquilin-Arista, F.; Alvarez-Avellon, T.; Menendez-Gonzalez, M. Prevalence of Depression and Anxietyin Parkinson Disease and Impact on Quality of Life: A Community-Based Study in Spain. J. Geriatr. Psychiatry Neurol. 2019, 34, 207–213. [Google Scholar]
- Antonini, A.; Barone, P.; Bonuccelli, U.; Annoni, K.; Asgharnejad, M.; Stanzione, P. ICARUS study: Prevalence and clinical features of impulse control disorders in Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2017, 88, 317–324. [Google Scholar] [CrossRef]
- Byers, B.; Cord, B.; Nguyen, H.N.; Schule, B.; Fenno, L.; Lee, P.C.; Deisseroth, K.; Langston, J.W.; Pera, R.R.; Palmer, T.D. SNCA triplication Parkinson’s patient’s iPSC-derived DA neurons accumulate alpha-synuclein and are susceptible to oxidative stress. PLoS ONE 2011, 6, e26159. [Google Scholar] [CrossRef] [PubMed]
- Mogi, M.; Harada, M.; Kondo, T.; Narabayashi, H.; Riederer, P.; Nagatsu, T. Transforming growth factor-β1 levels are elevated in the striatum and in ventricular cerebrospinal fluid in Parkinson’s disease. Neurosci. Lett. 1995, 193, 129–132. [Google Scholar] [CrossRef]
- Mogi, M.; Harada, M.; Narabayashi, H.; Inagaki, H.; Minami, M.; Nagatsu, T. Interleukin (IL)-1, IL-2, IL-4, IL-6 and transforming growth factor-levels are elevated in ventricular cerebrospinal fluid in juvenile parkinsonism and Parkinson’s disease. Neurosci. Lett. 1996, 211, 13–16. [Google Scholar] [CrossRef]
- Jha, M.K.; Jo, M.; Kim, J.H.; Suk, K. Microglia-Astrocyte Crosstalk: An Intimate Molecular Conversation. Neuroscientist 2019, 25, 227–240. [Google Scholar] [CrossRef]
- Fung, T.C.; Olson, C.A.; Hsiao, E.Y. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 2017, 20, 145–155. [Google Scholar] [CrossRef]
- Zoghbi, H.Y.; Bear, M.F. Synaptic dysfunction in neurodevelopmental disorders associated with autism and intellectual disabilities. Cold Spring Harb. Perspect. Biol. 2012, 4, a009886. [Google Scholar] [CrossRef] [Green Version]
- Kleijer, K.T.E.; Huguet, G.; Tastet, J.; Bourgeron, T.; Burbach, J.P.H. Anatomy and Cell Biology of Autism Spectrum Disorder: Lessons from Human Genetics. Adv. Anat. Embryol. Cell Biol. 2017, 224, 1–25. [Google Scholar]
- Parikshak, N.N.; Gandal, J.M.; Geschwind, D.H. Systems biology and gene networks in neurodevelopmental and neurodegenerative disorders. Nat. Rev. Genet. 2015, 16, 441–458. [Google Scholar]
- Masi, A.; DeMayo, M.M.; Glozier, N.; Guastella, A.J. An Overview of Autism Spectrum Disorder, Heterogeneity and Treatment Options. Neurosci. Bull. 2017, 33, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermanowicz, N.; Jones, S.A.; Hauser, R.A. Impact of non-motor symptoms in Parkinson’s disease: A PMDAlliance survey. Neuropsychiatr. Dis. Treat. 2019, 15, 2205–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.Y.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. α-synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.L.; Chen, H.I.; Li, L.H.; Chien, Y.L.; Liao, H.M.; Chou, M.C.; Chou, W.J.; Tsai, W.C.; Chiu, Y.N.; Wu, Y.Y.; et al. Genome-wide analysis of copy number variations identifies PARK2 as a candidate gene for autism spectrum disorder. Mol. Autism 2016, 7, 23. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson-Agramonte, M.d.l.A.; Noris García, E.; Fraga Guerra, J.; Vega Hurtado, Y.; Antonucci, N.; Semprún-Hernández, N.; Schultz, S.; Siniscalco, D. Immune Dysregulation in Autism Spectrum Disorder: What Do We Know about It? Int. J. Mol. Sci. 2022, 23, 3033. https://doi.org/10.3390/ijms23063033
Robinson-Agramonte MdlA, Noris García E, Fraga Guerra J, Vega Hurtado Y, Antonucci N, Semprún-Hernández N, Schultz S, Siniscalco D. Immune Dysregulation in Autism Spectrum Disorder: What Do We Know about It? International Journal of Molecular Sciences. 2022; 23(6):3033. https://doi.org/10.3390/ijms23063033
Chicago/Turabian StyleRobinson-Agramonte, Maria de los Angeles, Elena Noris García, Jarasca Fraga Guerra, Yamilé Vega Hurtado, Nicola Antonucci, Neomar Semprún-Hernández, Stephen Schultz, and Dario Siniscalco. 2022. "Immune Dysregulation in Autism Spectrum Disorder: What Do We Know about It?" International Journal of Molecular Sciences 23, no. 6: 3033. https://doi.org/10.3390/ijms23063033
APA StyleRobinson-Agramonte, M. d. l. A., Noris García, E., Fraga Guerra, J., Vega Hurtado, Y., Antonucci, N., Semprún-Hernández, N., Schultz, S., & Siniscalco, D. (2022). Immune Dysregulation in Autism Spectrum Disorder: What Do We Know about It? International Journal of Molecular Sciences, 23(6), 3033. https://doi.org/10.3390/ijms23063033