1. Introduction
WRKY transcription factors (TFs) are named based on the conserved residue WRKY, which constitutes an integral part of their DNA-binding domain (DBD) and is approximately 60 residues in length. The key structural features of this domain are the conserved DNA-binding motif WRKYGQK and a zinc-finger motif at the C-terminus [
1]. Previous research has indicated that WRKY TFs should be classified into three groups based on the number of WRKY domains and the type of zinc-finger motif they contain [
2]. The group 1 WRKY TFs contain two WRKY domains, and each of these has a C2H2 motif. The group 2 TFs contain one WRKY domain with a C2H2 zinc-finger motif, whereas the group 3 TFs contain one WRKY domain with a C2HC zinc-finger motif. This grouping of WRKY TFs has been widely used, but molecular evidence has now shown that it is inconsistent with phylogeny [
2,
3,
4]. For example, the
WRKY genes from
Arabidopsis thaliana can be grouped into group Ia, comprising
AtWRKY1, -
2,
-3,
-4,
-10,
-25,
-26,
-32,
-33,
-34,
-44, and -
58, and group Ib, comprising
AtWRKY8,
-12,
-13,
-23,
-24,
-28,
-43,
-45,
-48,
-56,
-57,
-68,
-71, and
-75; these two groups are sister groups. Group IIa, comprising
AtWRKY6,
-9, -18,
-31,
-36,
-40,
-42,
-47,
-60,
-61, and
-72, is followed [
4]. However, group Ia and IIa members were treated as a monophyletic group named group IIa by Mohanta et al. [
3]. In addition, some members of the groups Ia and Ib that were defined by Wang et al. [
4] (
AtWRKY1,
-2,
-3,
-4,
-10,
-20,
-25,
-26,
-32,
-33,
-34,
-44,
-45, and
-58) were designated group I by Eulgem et al. [
2]. The remaining members of Wang et al.’s groups Ia and Ib (
AtWRKY8,
-12,
-13,
-23,
-24,
-28,
-43,
-48,
-56,
-68,
-71, and
-75) were absent from the group I defined by Eulgem et al. [
2]. This inconsistency among studies leads not only to improper citations of WRKY TFs, but also confuses postulations about their evolutionary scenario. The confusion may result from the fact that WRKY genes have been classified based on whether they have one or two WRKY domains. [
2,
3,
4]. Furthermore, which type of WRKY gene is closest to the ancestral form remains unclear. Recently, it has been assumed that both types of gene (having one or two WRKY domains) evolved from one ancestral WRKY TF, following diversification and amino acid substitutions in the derived WRKY genes [
5]. However, a lack of phylogenetic support makes this proposal debatable.
WRKY TFs are reported to be involved in plant growth, development, metabolism, responses to environmental stresses, and senescence [
5]. For example, the WRKY TFs WRKY53, WRKY54, and WRKY70 have been characterized as leaf-senescence regulators [
6,
7,
8]. A line of
Arabidopsis thaliana overexpressing
WRKY53 showed whole-plant senescence two weeks earlier than the wild type, and a knock-out line showed delayed senescence compared to the wild type [
7]. In addition, the expression level of
WRKY53 increased in rosette leaves prior to the expression of several senescence-associated genes (
SAG), such as
SAG12 [
9]. These studies suggest that
WRKY53 is a positive senescence regulator in the senescence process. In contrast, a loss-of-function
wrky70 line showed earlier senescence than wild-type plants, suggesting that
WRKY70 functions as a negative regulator of senescence [
8].
WRKY54, a homologue of
WRKY70, displayed similar expression patterns to
WRKY70, suggesting a redundancy between these two WRKY TFs [
6].
WRKY70 and
WRKY54 are also crucial in the response against pathogenic infection, in that they are involved in regulating the salicylic acid and jasmonic acid signaling that is part of plant defense mechanisms [
5,
6,
10]. Restricted lesion diameter and bacterial growth observed in
wrky54wrky70 double mutants suggest that WRKY54 and WRKY70 serve as negative regulators of the leaf senescence process [
6,
10,
11]. Therefore, WRKY53, WRKY54, and WRKY70 are regarded as three key components in the control of plant senescence and defense processes, which act by receiving external factors and conducting subsequent signaling transduction [
5].
To perform their regulatory function in the modulation of gene expression, WRKY TFs bind to a consensus sequence TTGAC-C/T, named W-box, to regulate the expression of target genes [
1,
12,
13]. An extensive survey of binding specificity showed that WRKY TFs exhibit a W-box preference [
10]. According to the report, WRKY11 (a member of group IId) binds to the second and eleventh W-box of
Arabidopsis thaliana senescence-induced receptor-like kinase (
AtSIRK) promoter, while WRKY26 (group I) binds to the eighth W-box of promoter
AtSIRK [
10]. In addition, they found that WRKY11 binds to W-box with a G residue directly at the 5′ end. In contrast, WRKY26 prefers to interact with W-box with a T, C, or A residue at the same site [
10]. As a determinant of the sequence-recognition profile of WRKY TFs, the flanking region of the W-box sequence exerts a profound influence on the DNA-binding preferences of structurally similar members of this highly conserved TF family.
At present, the recognition of specific W-box nucleotide sequence preferences in
A. thaliana has been elucidated by studying group I (WRKY26, as per Wang et al. [
4]) and group II (WRKY11, as per Wang et al. [
4]), while the preferences of group III WRKY TFs remains unknown. In the present study, the phylogenetic tree of WRKY across the plant kingdom was reconstructed and the DNA-binding characteristics of WRKY54, a representative of the group III WRKY TFs, were determined. We selected 528 WRKY genes to investigate their origin across the plant kingdom and identify whether the ancestral form of WRKY has single or double WRKY domains. To investigate the DNA-binding preferences of WRKY TFs containing a single WRKY domain, we probed the binding profile of the WRKY54 DBD against a series of W-box DNA sequences by means of a fluorescence-based electrophoretic mobility shift assay (fEMSA) and quartz crystal microbalance (QCM) analysis. In summary, a novel perspective on the evolutionary origin of WRKY TFs is provided through our reconstructed phylogenetic tree. In addition, the binding profile we describe for the WRKY54 DBD sheds light on the long-standing question of the protein–DNA recognition mechanism and the sequence-selective binding of the highly conserved WRKY TFs.
2. Results
2.1. Retrieval and Identification of WRKY Transcription Factor Genes
WRKY TF gene family was identified by using 16 species from across the plant kingdom (
Table 1). These included a rhodophyte, a chlorophyte, embryophytes (two), a tracheophyte, monocots (two), and dicots (eight). In total, we obtained 528 WRKY TF genes. In the selected species, we identified no WRKY TFs in
P. umbilicalis (rhodophyte), and only two in
Micromonas pusilla (chlorophyte). Large numbers of WRKY TFs were identified in the monocots (
Oryza sativa: 75;
Zea mays: 62) and dicots (
A. thaliana: 70). We only found WRKY TFs with two WRKY domains in the embryophytes. Based on multiple sequence alignment, there were significant differences in the sequence characteristics of the chlorophyte (
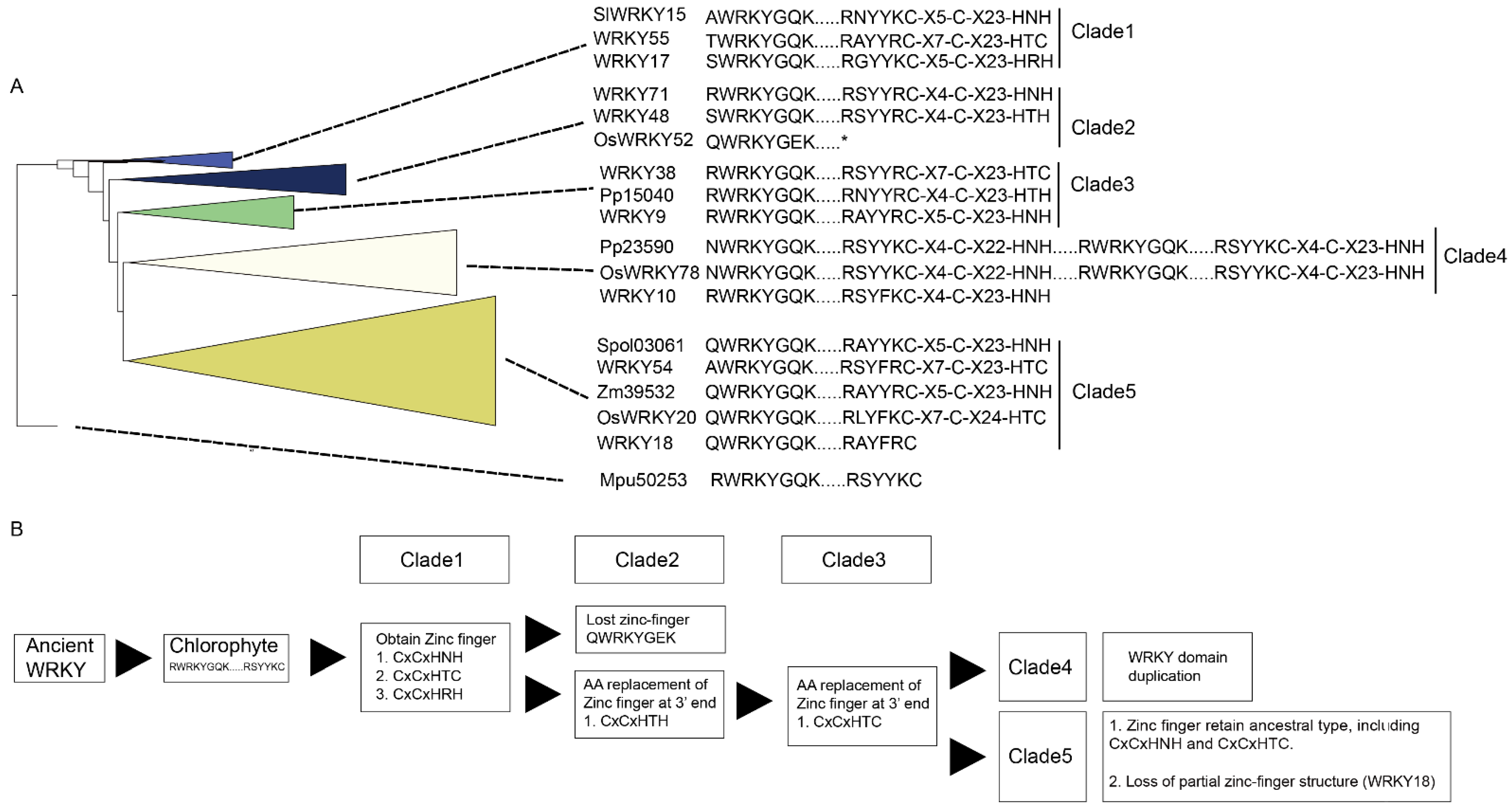
M. pusilla) WRKY TF and those of most of the other species. Almost all of the WRKY TFs were characterized by a conserved WRKY motif (xWRKYGQK or xWRKYGEK) with a zinc-finger motif (CxCxHTC or CxCxHxH), where x denotes any amino acid. However, the WRKY TF from
M. pusilla (Mpu50253) contained a conserved WRKY motif (RWRKYGQK) with only part of the zinc-finger motif (C), suggesting that three or four conserved amino acid substitutions (C → CxCxHxH or C → CxCxHTC) occurred after plants had colonized land.
2.2. Phylogenetic Reconstruction for WRKY Transcription Factors
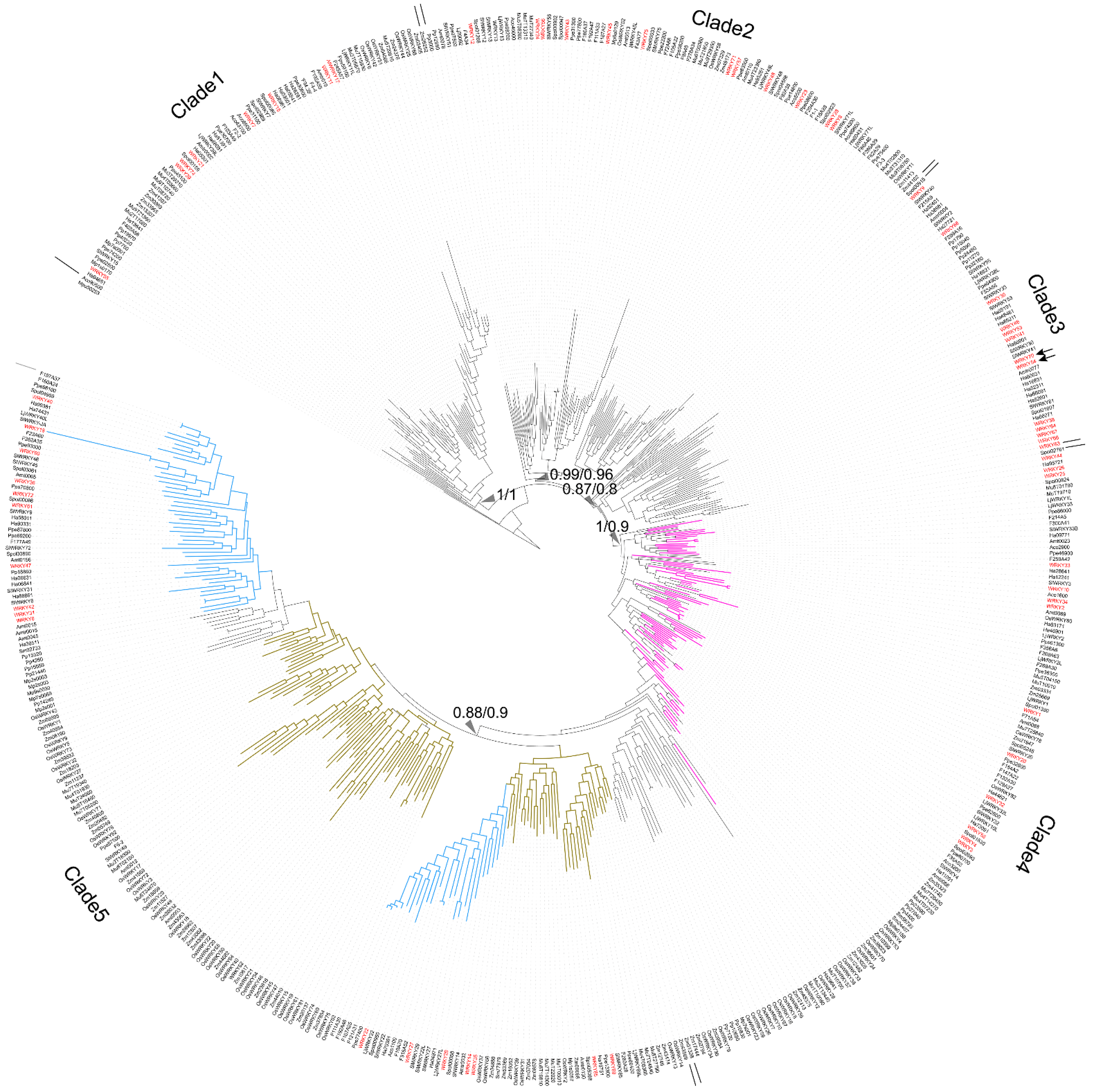
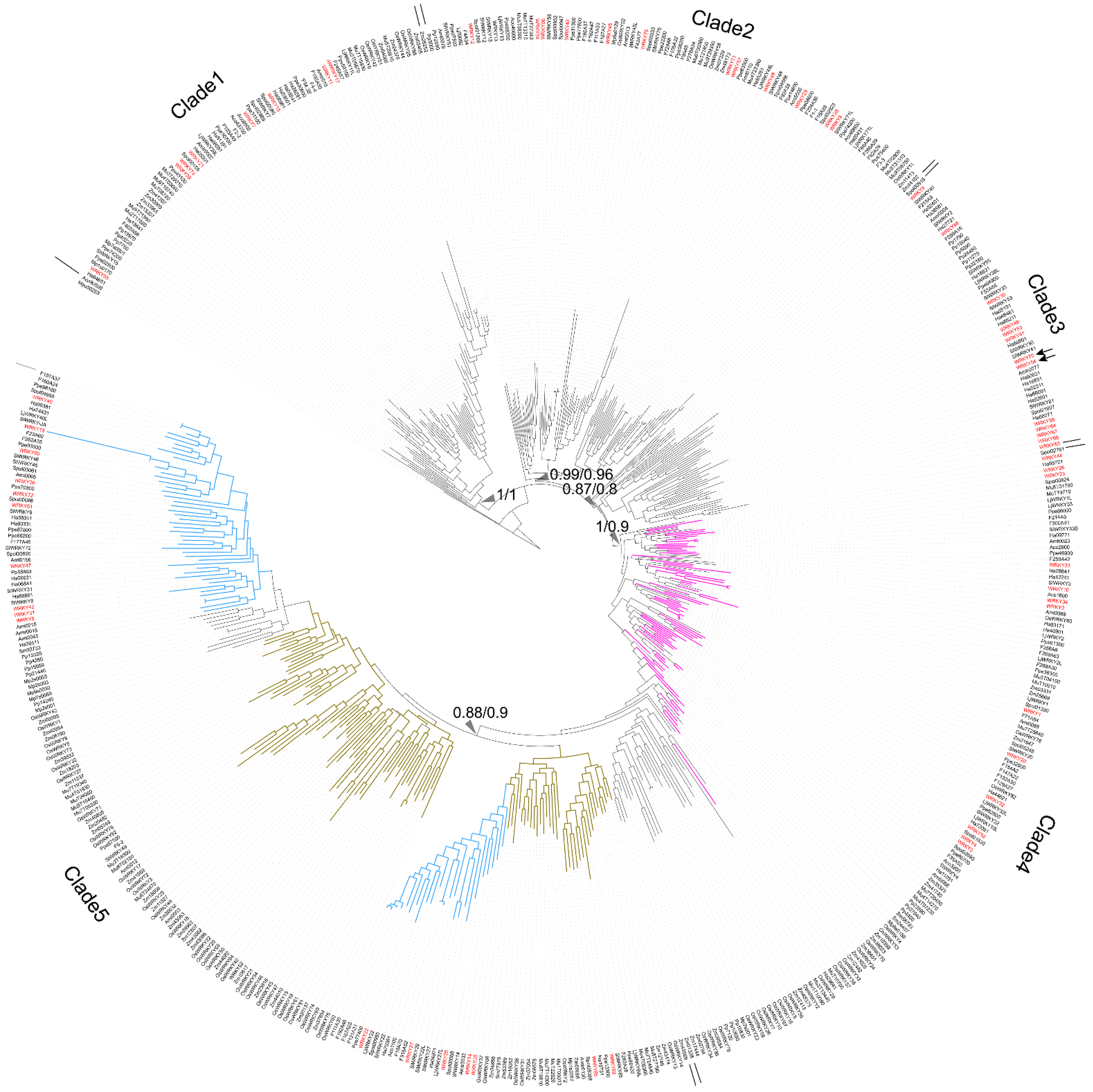
To trace the evolution of WRKY TFs, the WRKY TF obtained from
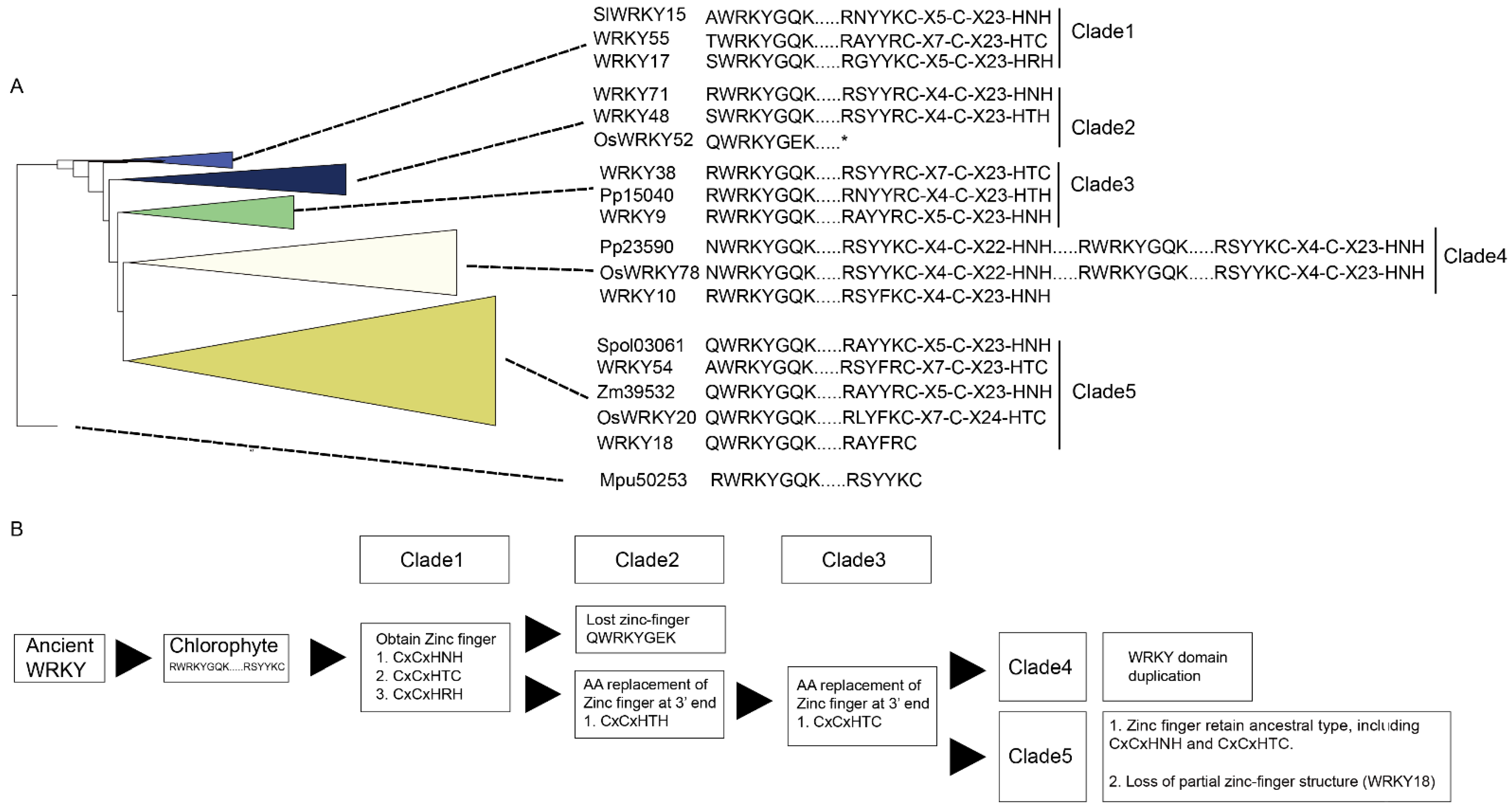
M. pusilla (Mpu50253, no zinc-finger motif) was chosen as the root of the reconstructed WRKY phylogeny (
Figure 1). In the resulting phylogeny, the TFs were grouped into five clades: Clade 1 (BI: 1; ML: 1), Clade 2 (BI: 0.99; ML: 0.96), Clade 3 (BI: 0.87; ML: 0.8), Clade 4 (BI: 1; ML: 0.9), and Clade 5 (BI: 0.8; ML: 0.9) (
Figure 1).
Clade 1 comprised the WRKY TFs obtained from O. sativa, A. thaliana, S. lycopersicum, P. persica, Fragaria x ananassa, M. acuminata, A. trichopoda and H. annuus. Three types of zinc-finger motifs were identified in this clade: CxCxHNH (SlWRKY15, AtWRKY55, and OsWRKY57), CxCxHTC (Ha94651) and CxCxHRH (OsWRKY68 and WRKY17).
Clade 2 contained WRKY TFs from across the plant kingdom, from Physcomitrium patens (e.g., Pp3000) to Prunus persica (e.g., Ppe74300). There were two main zinc-finger structure types identified in Clade 2: CxCxHNH (e.g., WRKY71, Mp6s0129, Spol02923, and Aco99600) and CxCxHTH (e.g., WRKY48, Zm07329, OsWRKY11, and Spol04568). A WRKY TF lacking a zinc-finger motif, OsWRKY52, was also located in Clade 2.
In Clade 3, the major zinc-finger motif types were CxCxHNH (e.g., Ha38981, WRKY9, and F215A8), CxCxHTH (e.g., Pp15040 and Pp32160) and CxCxHTC (e.g., WRKY38, SlWRKY55 and WRKY46). Notably, no WRKY TFs from the monocots was included in Clade 3.
In Clade 4, the WRKY TFs containing two WRKY domains were dominant; these were found in embryophytes, monocots, and dicots (purple branches in
Figure 1). The predominant zinc-finger motif was CxCxHNH-CxCxHNH (e.g., Pp23590, OsWRKY78, and WRKY1). The zinc-finger motif in the WRKY TFs with only one WRKY domain was CxCxHNH (e.g., WRKY10).
Lastly, Clade 5 comprised only TFs with a single WRKY domain and a single zinc-finger motif, and these were assigned to monocot (brown-yellow branches in
Figure 1) and dicot subclades (pale blue branches in
Figure 1). Within the Clade, the main zinc-finger motif types were CxCxHNH (e.g., Zm39532, Spol03061, SlWRKY27, and Ha36511), CxCxHSH (e.g., OsWRKY5) and CxCxHTC (e.g., OsWRKY20 and Zm23616). There were also WRKY TFs without a complete zinc-finger motif (e.g., WRKY18) in the Clade 5.
Finally, the long branches observed in the reconstructed WRKY phylogeny suggest a high degree of nucleotide variation across the plant kingdom.
2.3. The Single WRKY Domain of WRKY54 Exists as Both a Monomer and in an Aggregated Form In Vitro
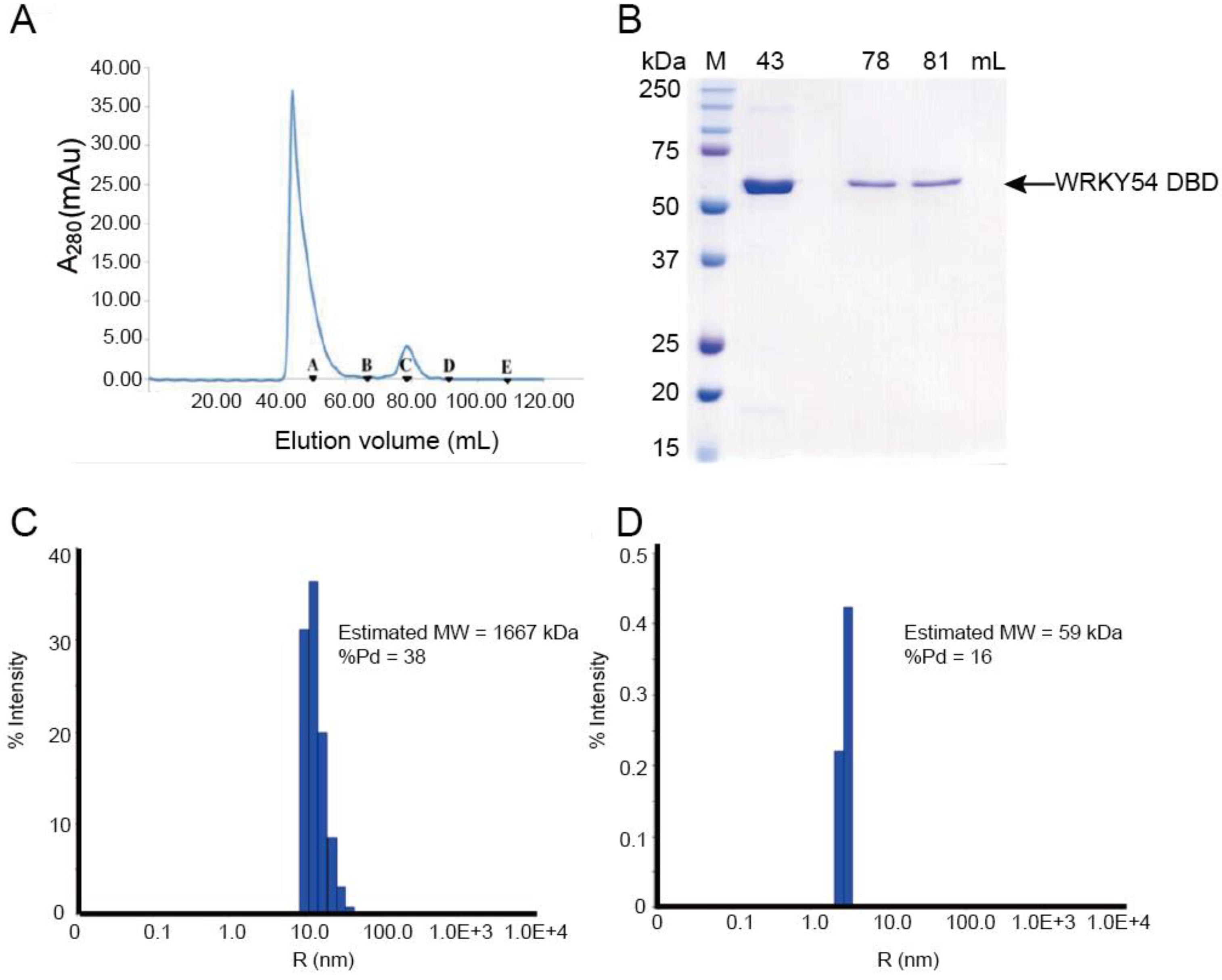
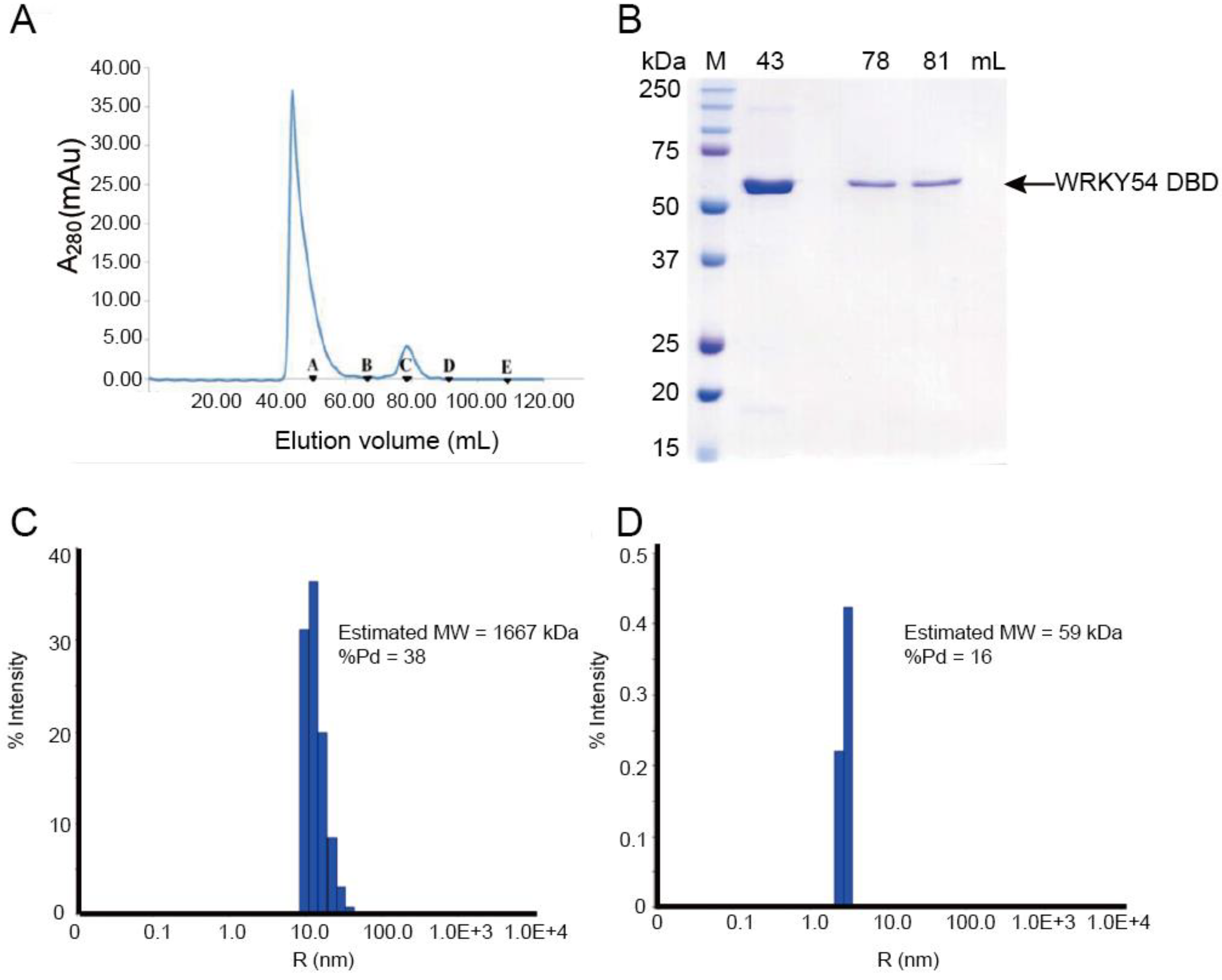
To study the binding ability of the single WRKY domain in Clade 3, the recombinant WRKY54 protein was expressed and purified. In the first trial, the full length of the WRKY54 recombinant protein exhibited an aggregated form with no DNA binding ability in solution after cell lysis. The DNA binding domain of WRKY54 from residues 133–224 was constructed. After protein expression and purification, the recombinant WRKY54 DBD protein exhibited two forms—an aggregated form and a monomeric form—that corresponded to two major peaks in the SEC elution profile (
Figure 2A). SDS-PAGE showed that main protein product size of the eluted solution collected at 43 mL, 78 mL, and 81 mL was 50–75 kDa (
Figure 2B). To verify the oligomerization state of the WRKY54 DBD, protein solutions collected at 43, 78 and 81 mL were subjected to DLS. The protein solutions collected at 78 and 81 mL may have contained the same protein product, so these solutions were pooled for the DLS analysis. The DLS results showed that the molecular mass of the WRKY54 DBD fraction collected at 43 mL was 1667 kDa, suggesting that the WRKY54 DBD in this fraction was in the aggregate form (
Figure 2C). In contrast, the protein size measured in the pooled solutions collected at 78 and 81 mL was 59 kDa, suggesting that the WRKY54 DBD in these fractions was in the monomer form (
Figure 2D).
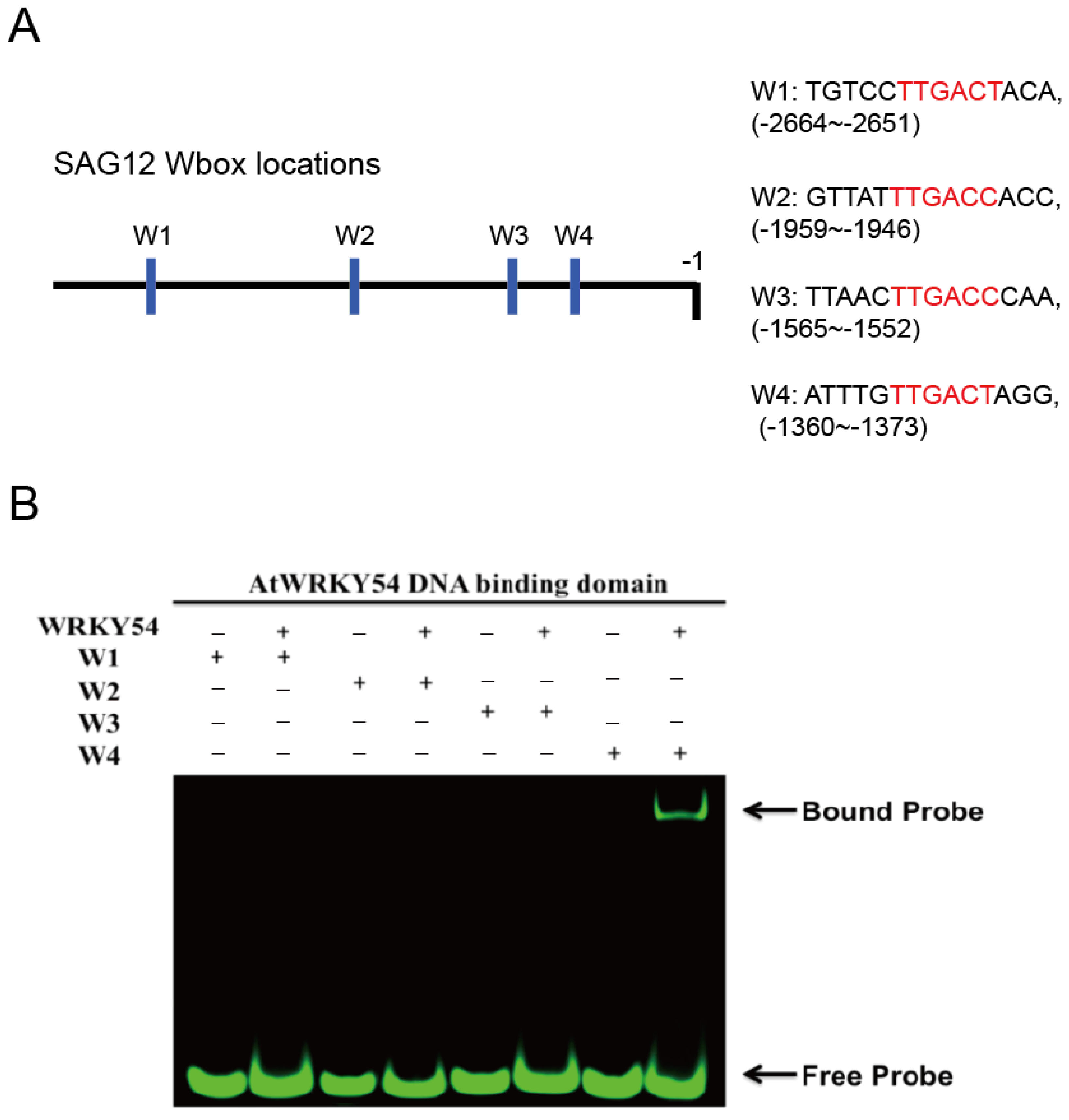
2.4. WRKY54 DNA-Binding Domain Can Bind to W4 Box from SAG12 Upstream Sequence
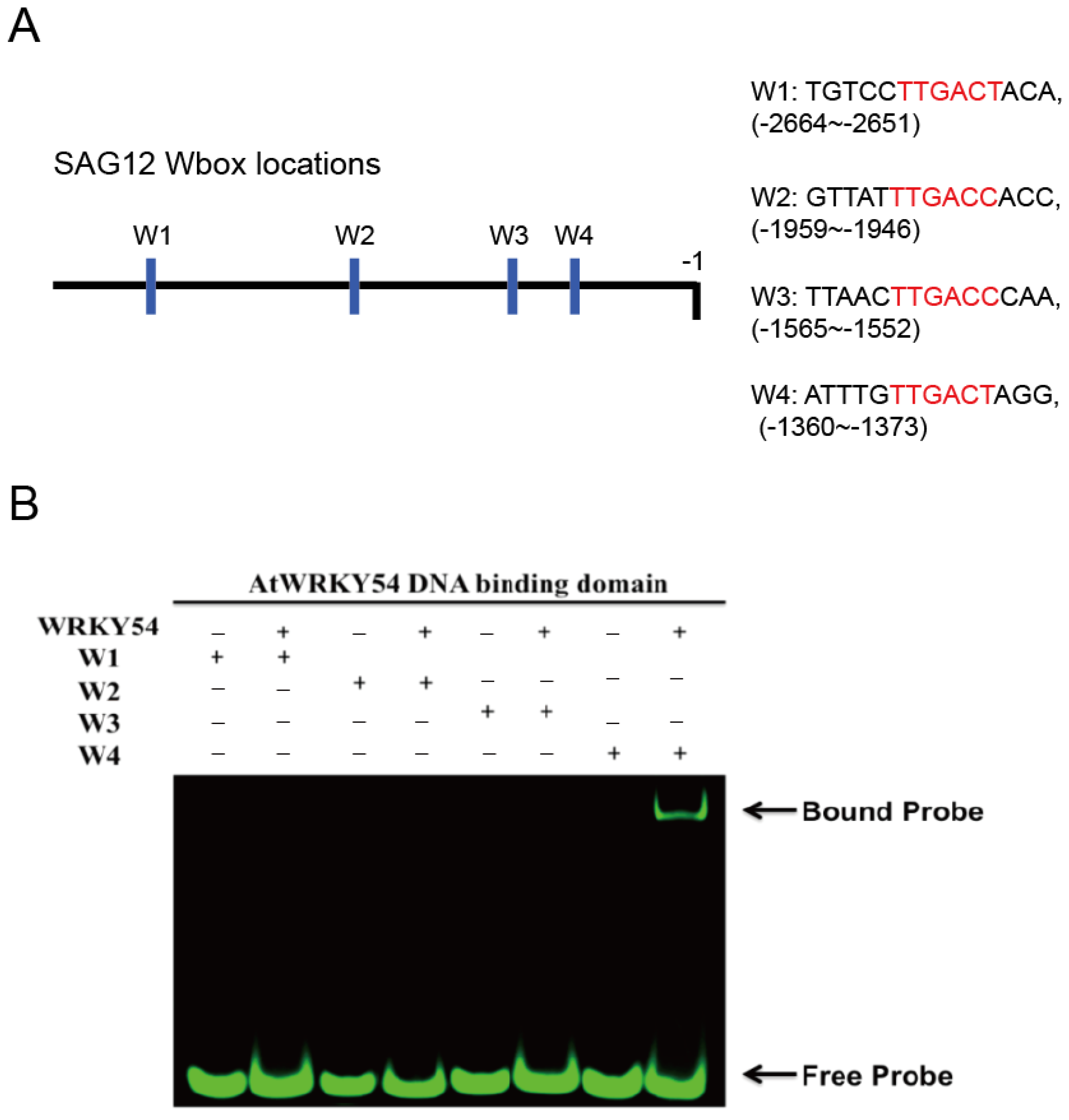
It is known that WRKY TFs bind to the DNA sequence named W-box (5′-TTGAC-C/T-3′). However, several sequences upstream of the promoters triggered by WRKY TFs contain W-box–centered sequences with TTGACC or TTGACT. Therefore, we first used a fEMSA to examine whether the WRKY54 DBD can bind to W-box sequences with TTGACC or TTGACT from the
SAG12 upstream region. Four types of W-box (W1–W4) obtained from
SAG12 upstream regions were used as probes (
Figure 3A). Those four types of W-box were labeled with fluorescein and incubated with purified WRKY54 DBD protein. The results confirmed binding of the WRKY54 DBD protein to W4, but no binding to W1, W2, or W3 (
Figure 3B). These four W-box sequences differ in the nucleotides at either the 5′ or the 3′ flanking region (
Figure 3A). To examine whether the nucleotides located adjacent to W4 box affect the binding ability of WRKY54, we designed a series of W4 probes.
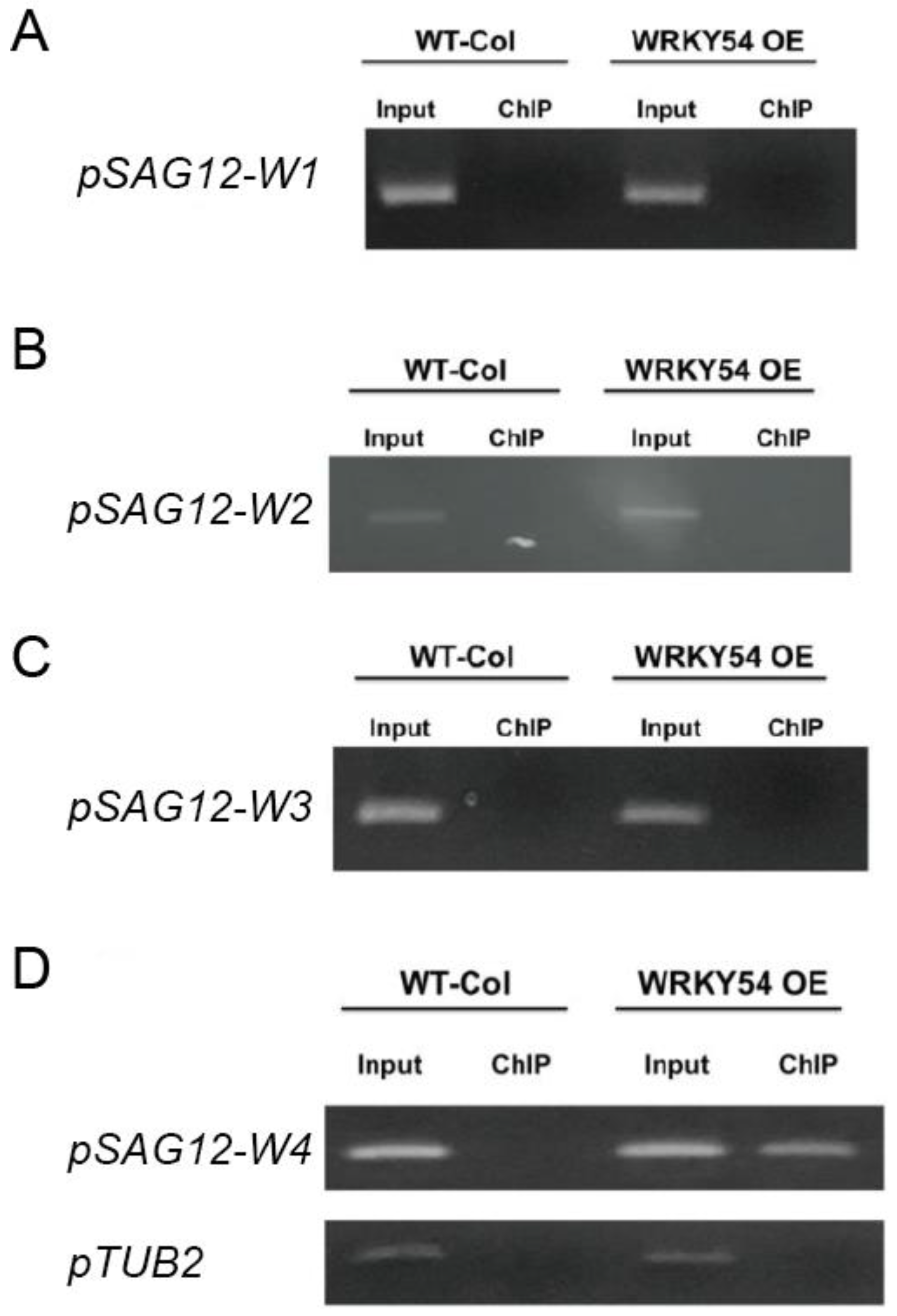
2.5. WRKY54 Can Bind to the W4 of SAG12 In Vivo
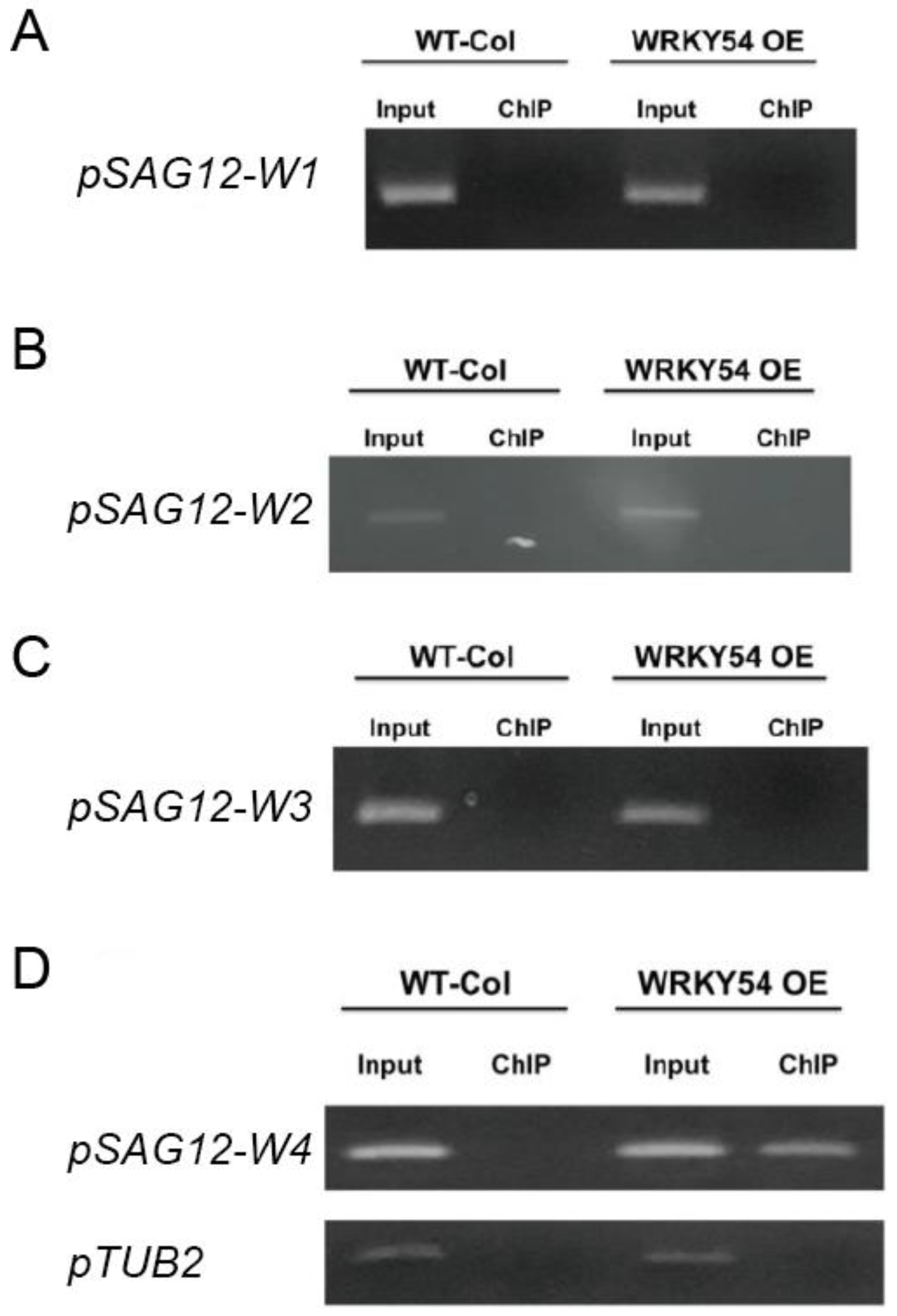
The ChIP-PCR technique was used to confirm whether WRKY54 binds specifically to W4 in vivo. Consistent with our fEMSA results, WRKY54 failed to bind to W1–W3 in either wild type
Arabidopsis or a WRKY54-overexpression line (
Figure 4A–C). However, for W4, a clear banding pattern could be seen for the WRKY54-overexpression line, although it was weaker for the wild type (
Figure 4D).
2.6. Length of the Flanking Region Adjacent to W-Box Affects the Binding Ability of WRKY54 DBD
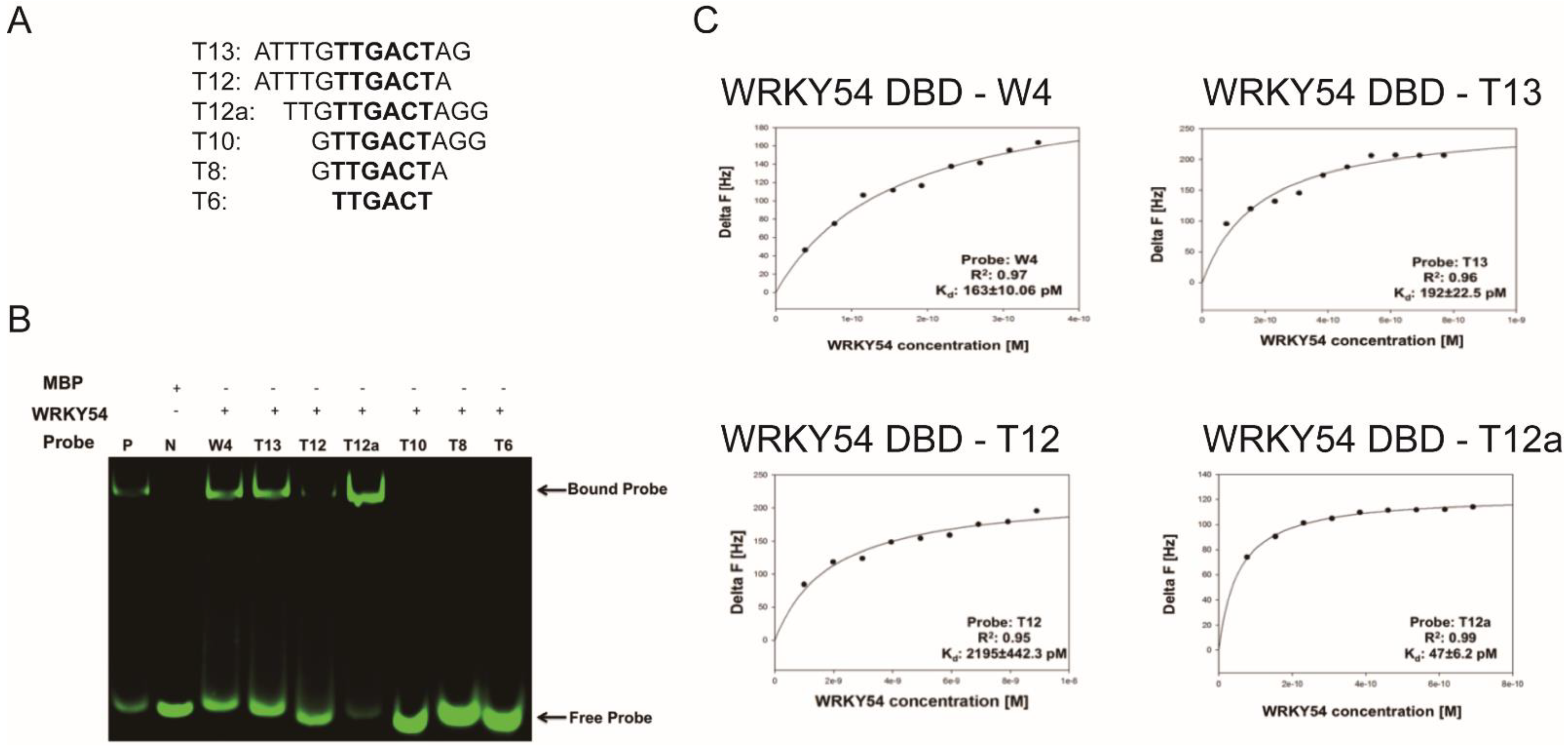
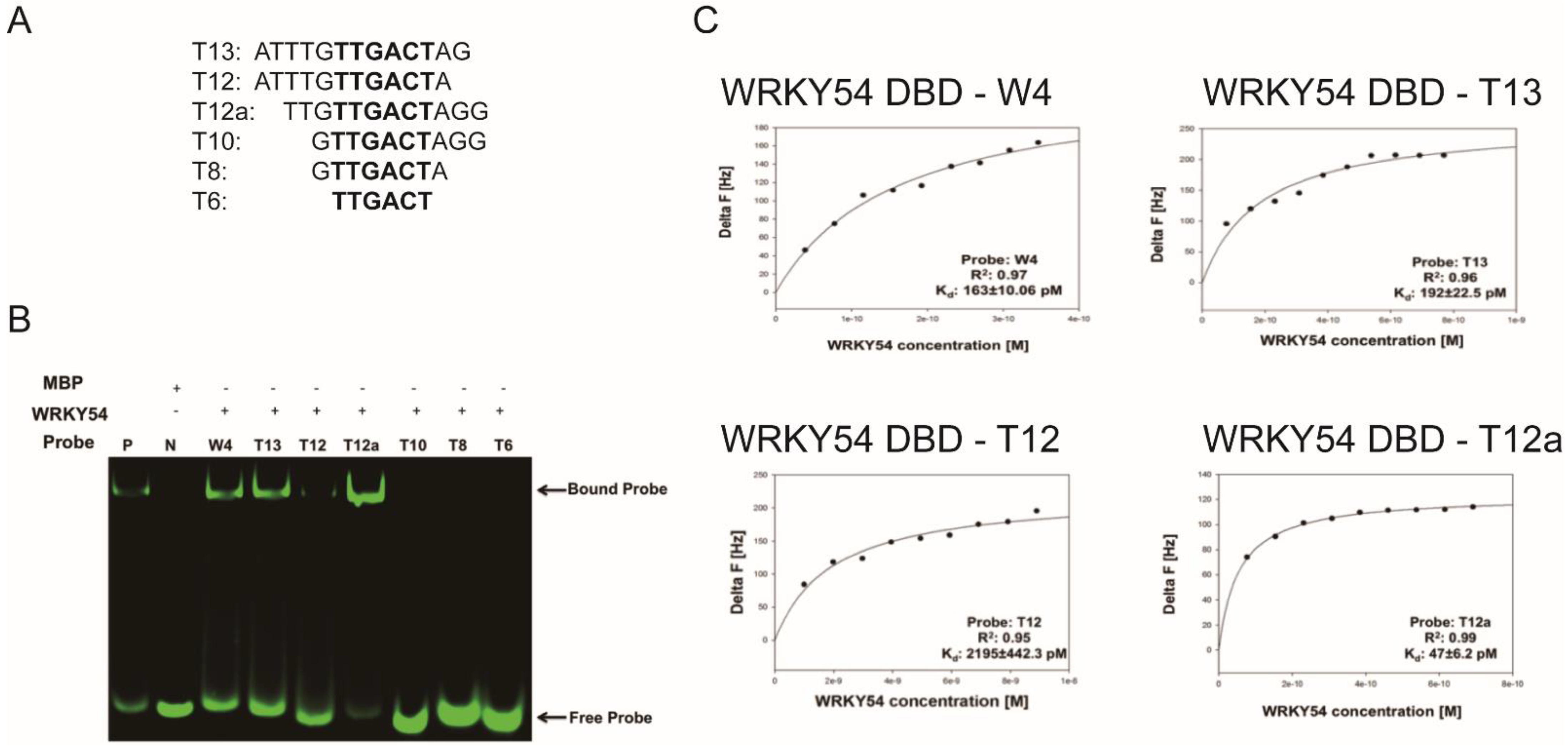
We then examined whether the composition of the flanking region adjacent to W4 box affected the specific binding ability of the WRKY54 DBD. Six artificial W4 variants were synthesized for this purpose (
Figure 5A). Clear banding shift patterns were observed for the WRKY54 DBD–W4, WRKY54 DBD–T13, and WRKY54 DBD–T12a pairs (
Figure 5B). In contrast, the WRKY54 DBD–T12 pair displayed a relatively weak banding shift. For the pairs involving T6, T8, and T10, no banding shift was observed, suggesting that the WRKY54 DBD was unable to bind to those artificial W4-like nucleotides (
Figure 5B).
The QCM was then used to examine the binding constants of the WRKY54 DBD to W4 and the W4-like nucleotides (
Table 2). The
Kd value obtained from the W4–WRKY54 DBD pair was 163 ± 10.06 pM. The
Kd values were 192 ± 22.5 pM and 2195 ± 442.3 pM for the T13–WRKY54 DBD and T12–WRKY54 DBD pairs, respectively. When the 5′-end nucleotides were removed, the
Kd values were 47 ± 6.2 pM (T12a–WRKY54 DBD pair) and 68 ± 11.73 pM (T13–WRKY54 DBD pair;
Table 1,
Figure 5C). There were no signals detected in the T6, T8, or T10 pairs. These results showed that a flanking region with at least three nucleotides at the 5′ end of TTGACT is required for the binding of the WRKY54 DBD.
2.7. Structural Insights into the DNA Binding of WRKY54 to W-Box
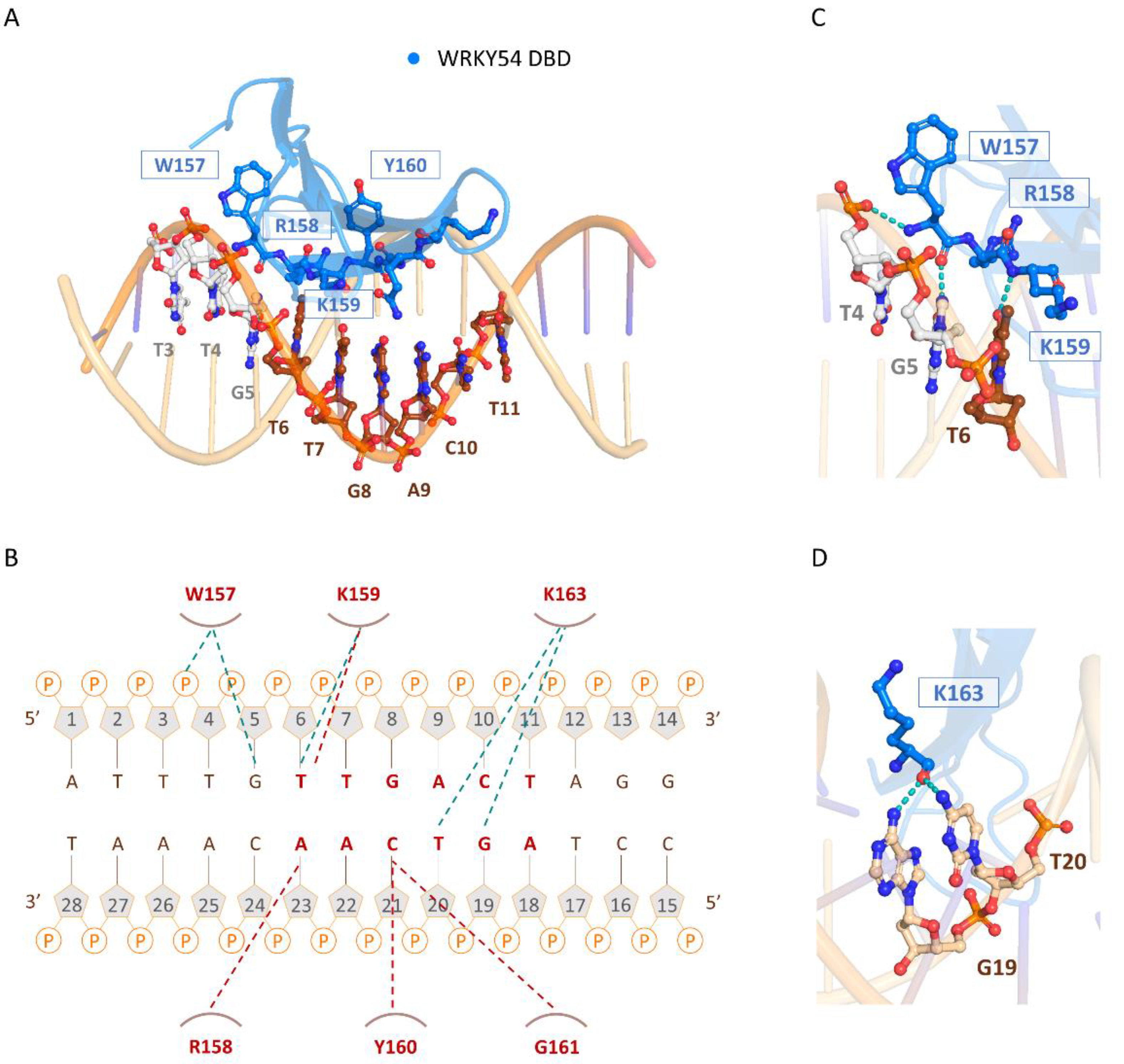
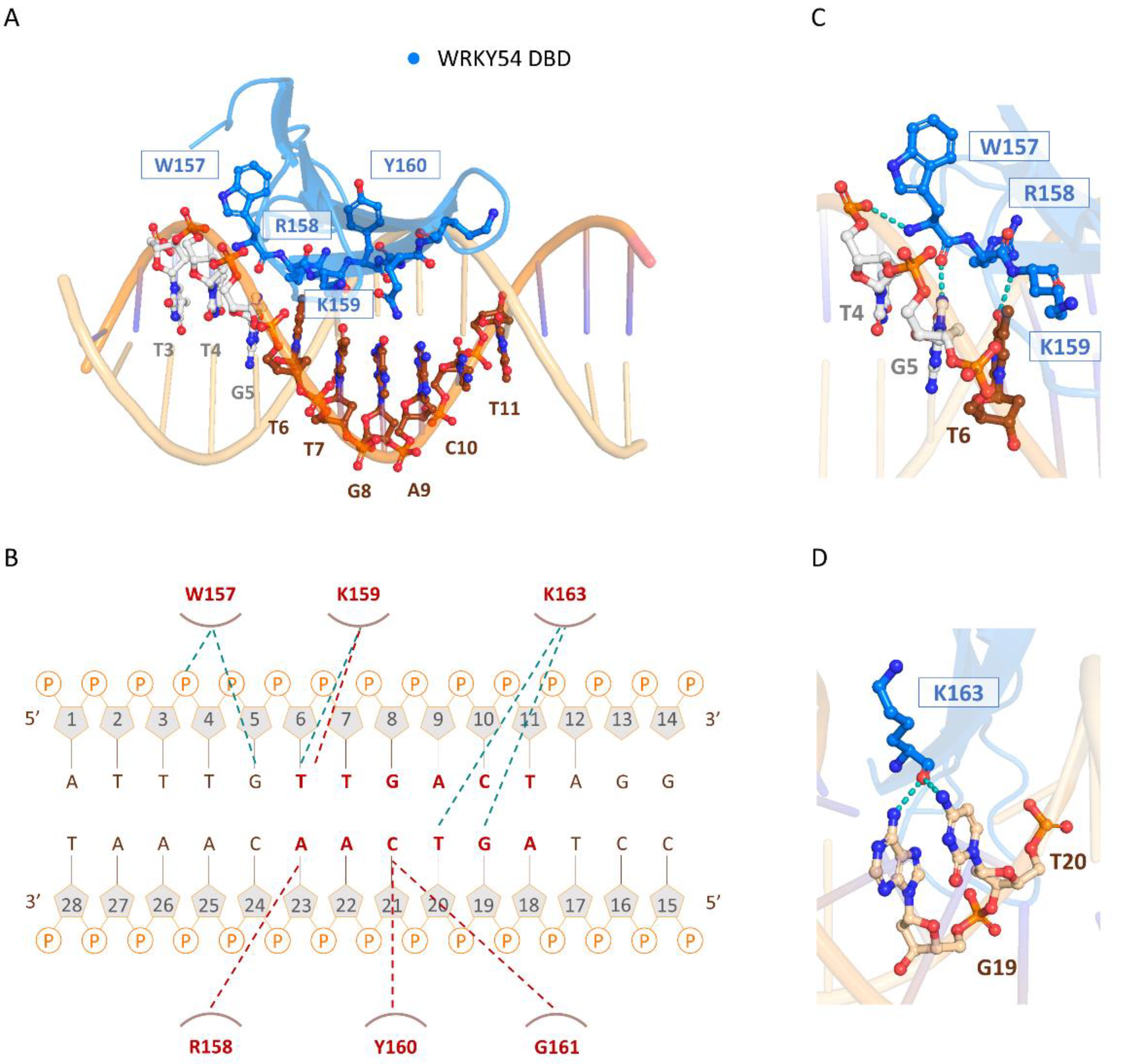
The three-dimensional model of the docked structure of the WRKY54 DBD revealed a four-stranded antiparallel β-sheet with the conserved WRKYGQK (W157, R158, K159, Y160, G161, Q162, and K163) motif located in the β1 strand. Insertion of the four-stranded antiparallel β-sheet into the major groove of W-box DNA permits extensive interactions between the residues of the WRKY54 DBD and the DNA nucleotides (5′-ATTTG
TTGACTAGG-3′, W-box sequence underlined), as illustrated in
Figure 6A. At the protein–DNA interface, intermolecular contacts between the WRKY54 DBD and the W4 nucleotides are primarily between the conserved residues W157, R158, K159, Y160, G161, and K163 on the β1 strand and dT4–dT7 and dG19–dA23 on the DNA. In this context, molecular interactions are mediated mainly by the formation of apolar contacts and H-bonds between the conserved residues of the WRKYRQK binding motif (W157, R158, K159, and Y160) and the DNA nucleotides dG5, dT6, dG19, dT20, dC21 and dA22 (
Figure 6B). In this postulated scenario, intermolecular hydrogen-bonding interactions formed between W157 and dT4 and/or dG5; K159 and dT6; and K163 and dG19 and/or dT20 are involved in the specific binding of the WRKY54 DBD to the W-box DNA sequence (
Figure 6C,D). Based on the binding mode of the WRKY54 DBD to W4, a 5′ flanking region of W-box with at least three additional nucleotides is thus required for WRKY54 binding.
4. Materials and Methods
4.1. WRKY Phylogeny Reconstruction
First, the core residues of WRKY domains (53–55 residues in length, on average) of green plants were labeled using Bioedit to identify conserved
WRKY genes [
23]. Then, to investigate
WRKY gene evolution across the plant kingdom,
WRKY genes were retrieved from the online Phytozome database (
https://phytozome.jgi.doe.gov/pz/portal.html# accessed on 23 January 2022) [
24] and the database of the National Center for Biotechnology Information (NCBI). The 73
WRKY genes of
Arabidopsis thaliana (
AtWRKY) were used as templates for identifying the
WRKY genes of the following selected species:
Micromonas pusilla,
Marchantia polymorpha,
Selaginella moellendorffii,
Amborella trichopoda, basal angiosperms (such as
Amborella trichopoda), monocots (including
Zea mays and
Oryza sativa), and eudicots (including
Aquilegia coerulea, Solanum lycopersicum and
Arabidopsis thaliana) (for details see
Table 1 and
Table S1). In addition, two polyploidy species are also included in this study, such as
Musa acuminate (triploid) and
Fragaria x
ananassa (octoploid). These species are chosen to represent major plant lineages. Although we searched the
Porphyra umbilicalis (Rhodophyta) database, implemented in Phytozome, we found no WRKY-containing genes in that database. Also, a
WRKY gene found in
M. pusilla, Mpu62993, caused long-branch attraction in preliminary phylogenetic reconstruction. Thus, Mpu62993 was excluded from the reconstruction. To conduct a comprehensive search for the
WRKY genes of the selected species, the default threshold value and tblastx were used for each search. Only sequences containing a WRKY domain were used for the phylogenetic reconstruction.
First, all
WRKY nucleotide sequences were aligned using the MUSCLE algorithm [
25] in MEGA v.6 [
26]. Next, the resulting alignment matrix was visually refined based on amino acid translations using Bioedit [
23]. The best-fitting model of the aligned WRKY matrix was GTR + G + I, as evaluated using the Bayesian information criterion (BIC) [
27]. Both Bayesian inference (BI) and maximum likelihood (ML) algorithms were applied to infer the relationships among the identified WRKY genes using the PhyML 3.0 online interface [
28]. Branch support for nodes was assessed using an approximate likelihood ratio test (aLRT) [
29] and a Bayesian-like transformation of the aLRT (aBayes) [
30] for the ML and BI findings, respectively (see
Supplementary Figure S1 for details). A conventional bootstrap algorithm was conducted with 1000 replicates. The reconstructed phylogeny was visualized using the online interface Interactive Tree of Life (iTOL) v5 [
31].
4.2. Vector Construction for the WRKY54 DNA-Binding Domain
For protein expression and purification, the wild type WRKY54 gene (AT2G40750) was amplified via polymerase chain reaction (PCR) from the plasmid pET21a-WRKY54, which was provided by Dr. Shih-Tong Jeng (Institute of Plant Biology, National Taiwan University, Taipei, Taiwan). The DBD of WRKY54 (WRKY54 DBD) was amplified with the forward BamH1 primer (5′-CCCGGATCCGGATGCTACACTAGAA-3′) and the reverse Pst1 primer (5′-CCCCTGCAGGAAAAGGCTCGGTCTT-3′). It was then subcloned into the expression vector pMALH12-c5v, such that the recombinant protein WRKY54 contained a maltose binding protein tag, a thrombin cleavage site at the N-terminus, and a twelve-histidine tag at the C-terminus.
4.3. Protein Expression and Purification
The WRKY54 DBD recombinant protein was expressed using
Escherichia coli Rosetta-gamiB (DE3). Cultures were incubated at 37 °C to an OD
600 of 0.4–0.6 and induced by adding 0.1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG), then grown overnight at 16 °C. Bacterial cells were pelleted and lysed by sonication in 25 mL of lysis buffer (30 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid [HEPES], pH 7.5, 0.5 M NaCl, 20 mM imidazole) to obtain the WRKY54 DBD recombinant protein. After cell lysis, the cell debris were centrifuged for 25 min at 12,500×
g (J2-MC, Beckman, IN, USA). The supernatants were filtered through a 0.45 μm filter (Millipore, Burlington, MA, USA) and loaded into a 5 mL Ni
2+-Sepharose resin column (HisTrap FF, GE Healthcare, Chicago, IL, USA). The column was washed with 10 times the column volume of binding buffer prior to eluting the WRKY54 DBD recombinant protein. After the column washing, 50 times the column volume of elution buffer, complemented with 40 mM imidazole, was injected to elute the recombinant protein. The molecular mass of the purified 12 × His-WRKY54 DBD was estimated as 59 kDa based on sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS-PAGE) (
Figure 1).
For advanced purification, all proteins were passed through a Superdex S-75 column (GE Healthcare) with elution buffer (30 mM HEPES, pH 7.5, 0.5 M NaCl, 500 mM imidazole). The molecular mass of the WRKY54 recombinant protein was estimated using a Superdex S-200 column and calibrated with gel filtration standard protein markers (BioRad, Hercules, CA, USA: thyroglobulin, 670 kDa; γ-globulin [bovine], 158 kDa; ovalbumin [chicken], 44 kDa; myoglobin [horse], 17 kDa; vitamin B12, 1.35 kDa). After size-exclusion chromatography (SEC), all proteins were concentrated using Amicon Ultra-15 centrifugal filters (Merck, Darmstadt, Germany) and quantified using a DS-11 spectrophotometer (DeNovix, Wilmington, DE, USA).
4.4. Determination of Molecular Size via Dynamic Light Scattering
The protein samples were subjected to dynamic light scattering (DLS) after advanced purification by SEC. Data were collected using a Zetasizer Nano ZS DLS instrument (Malvern Instruments, Malvern, UK) equipped with 50 mW laser fiber. An appropriate refractive index, viscosity (10% glycerol), and temperature (25 °C) was set for each sample.
4.5. DNA Binding Assays via fEMSA
To visualize whether the WRKY54 DBD interacted with specific W-box regions of the SAG12 gene, a series of fEMSA assays were conducted. All W-box nucleotides (including W1 to W4, which were identified from the SAG12 promoter region, and artificial synthesized W-box–like nucleotides) were labeled with fluorescein at the 5′ end and used as DNA probes in the fEMSA. Prior to native polyacrylamide gel electrophoresis, the DNA probe–protein mix was incubated in the dark for 30 min. The DNA–protein binding reaction was performed by incubating purified WRKY54 DBD recombinant protein with the aforementioned double-stranded W-box and W-box–like nucleotides at 4 °C for 90 min at 120 V. After electrophoresis, the DNA probe–protein binding pattern was observed using a fluorescent luminescence image analyzer (FluorChem M, San Jose, CA, USA) at National Taiwan University.
4.6. Determination of Binding Constant between WRKY54 DBD Protein and W4 or W4-like DNA Using Quartz Crystal Microbalance Technique
The QCM technique was used to determine the dissociation constant (
Kd) values and examine the binding ability of the WRKY54 DBD protein with W4 DNA or W4-like DNA. The protein–nucleotide interactions in WRKY54 DBD–W4 DNA and –W4-like DNA pairs were analyzed by using an AffinixQN QCM biosensor (Initium, Tokyo, Japan). To measure
Kd, one has to describe the relationship between pressure and the number of active sites on the surface undergoing adsorption by applying the Langmuir equation ([
32]):
Here, [Host] represents the adsorbate surface and [Guest] represents the adsorbate molecule, while [Host][Guest] indicates that the [Host] and the [Guest] have formed an adsorbed complex. Kon and Koff represent the association and dissociation rate constants, respectively. The ratio of Koff to Kon is Kd, the dissociation constant.
Prior to usage, the QCM biosensor was washed twice with 3 μL of piranha solution (H2SO4 and H2O2 in a 3:1 ratio) and incubated with 1% SDS for 5 min. Then, 400 μL of reaction buffer (30 mM HEPES, pH 7.5, 0.5 M NaCl, and 2 mM tris(2-carboxyethyl) phosphine [TCEP]) was applied to the dried sensor to balance and set up the magnetic stir at 1000 rpm at 25 °C. Avidin (1 mg/mL) was injected into the reaction buffer and coated on the Au electrode plate until saturation. Excess proteins were washed out with the reaction buffer until the oscillation frequency became a static horizontal line. Next, 5 μL of a double-stranded DNA probe with a biotin-labeled 5′-end (50 μM) was added. When the frequency had stabilized at ca. 0.3 Hz/s, 1 mg/mL of WRKY54 DBD recombinant protein was injected every 10 min, 10 times. The continuous titration method was used to determine the binding constant of the protein–DNA pairs, and the values were recorded as multiple binding curves using the AffinixQN v2 software (Initium, Tokyo, Japan). Data from three independent repeats were processed using AQUA v2 software (Initium, Tokyo, Japan).
4.7. Examination of WRKY54 Protein and W-Box through Chromatin Immunoprecipitation In Vivo
The chromatin immunoprecipitation (ChIP) technique was used to examine whether WRKY54 interacts with the W-box identified from
SAG12 gene in vivo. Chromatin was extracted from 5.5-week-old
Arabidopsis plants. After fixation with 0.37% formaldehyde, the chromatin was sheared to an average length of 500–1000 bp by sonication and then immunoprecipitated with rabbit-anti-myc antibodies (Abcam, Cambridge, UK, ab9106). The cross-linking was then reversed, and the quantity of each precipitated DNA fragment was determined via PCR using specific primers (
Supplementary Table S2). Three biological replicates were performed.
4.8. Homology Modeling and Protein–DNA Complex Docking
To gain insight into the probable binding mode of WRKY54, homology modeling of the WRKY54 DBD was performed in PyMod 3 [
33] with the crystal structure of the
WRKY1 domain (PDB ID: 6J4E) as a template model. Next, protein–DNA docking was carried out using the ZDOCK server [
34]. From the 10 top-scoring docking poses generated by the rigid-body docking algorithm, a final model was selected based on information provided in previous protein–DNA binding studies [
14].





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}