
Antioxidative and Anti-Inflammatory Activities of Chrysin and Naringenin in a Drug-Induced Bone Loss Model in Rats

 ,
,  , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Retinoic Acid Is an Effective Drug for Induction of Osteoporosis in Rats
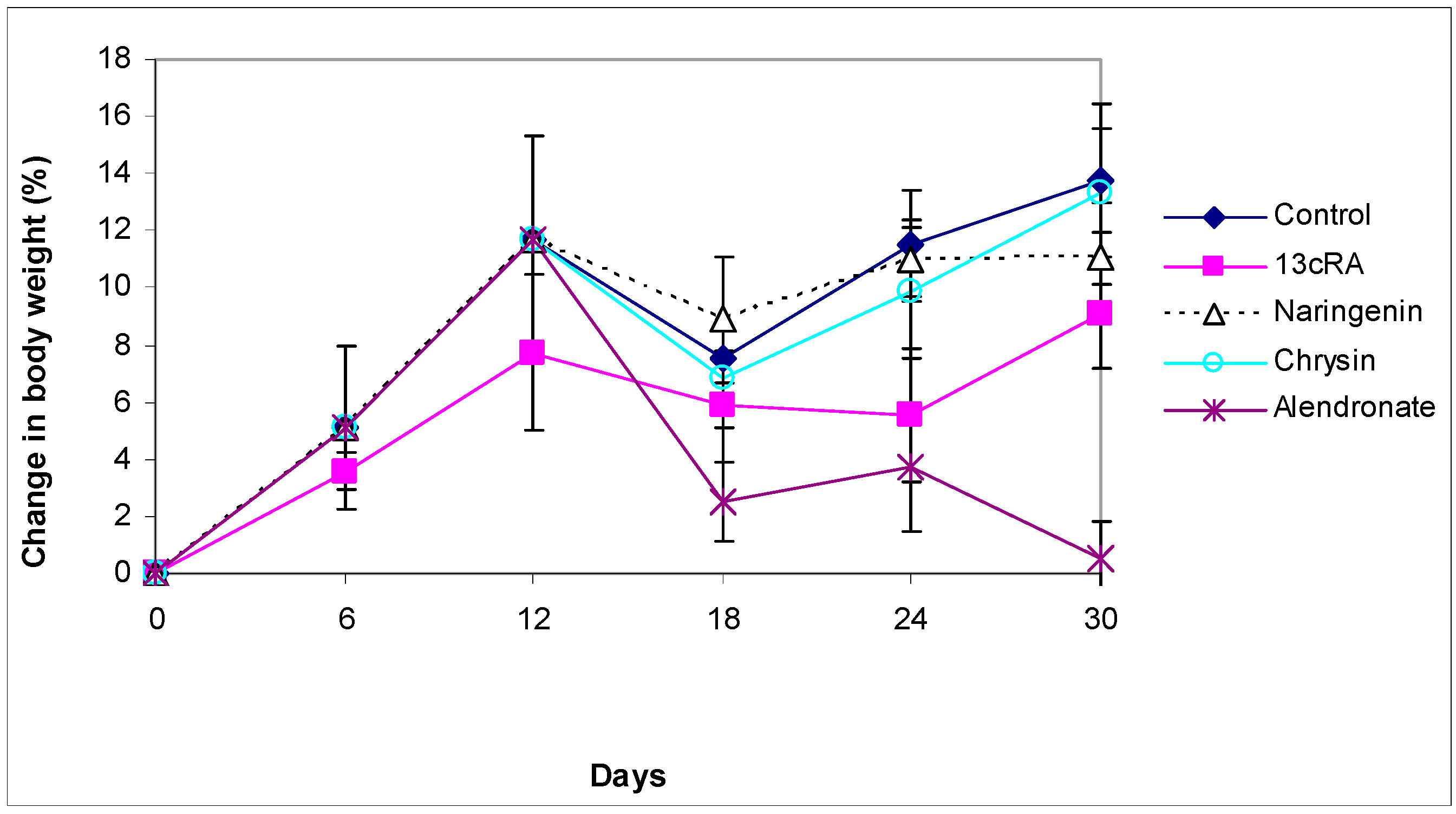
2.2. Body Weight Change
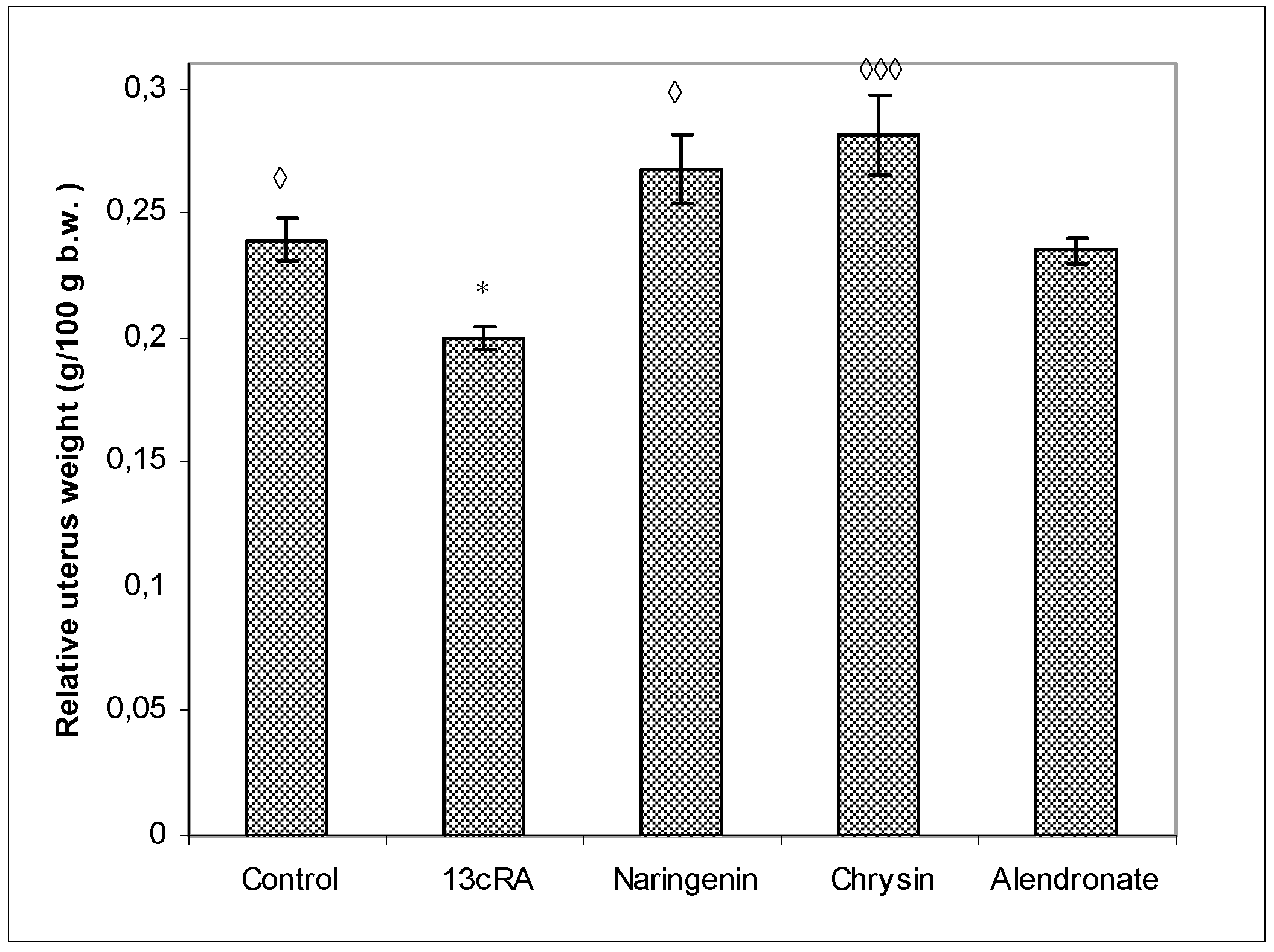
2.3. Relative Weight Changes of Uterus, Liver, Kidney, and Spleen
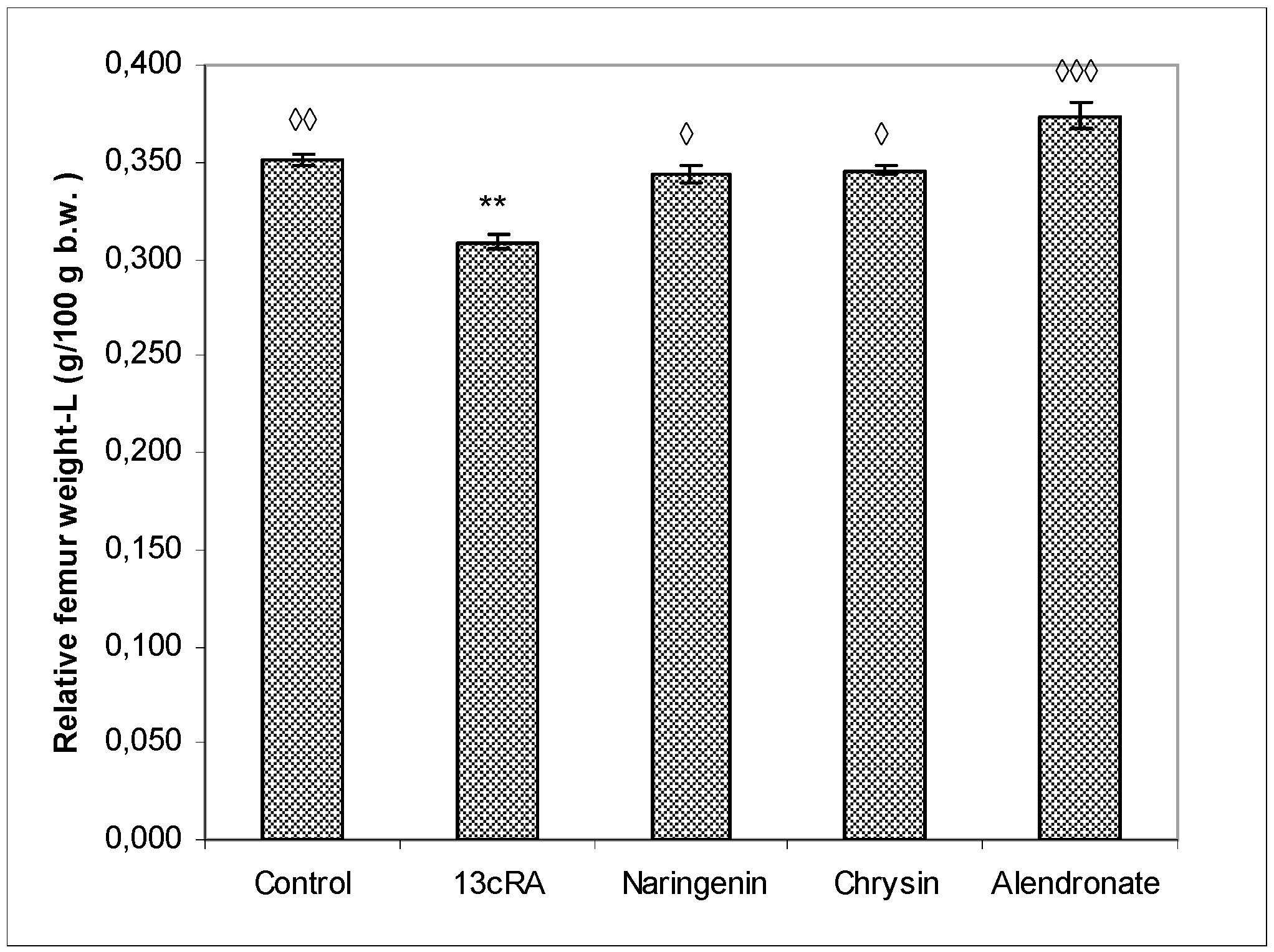
2.4. Relative Bone Weight, Length, and Diameter
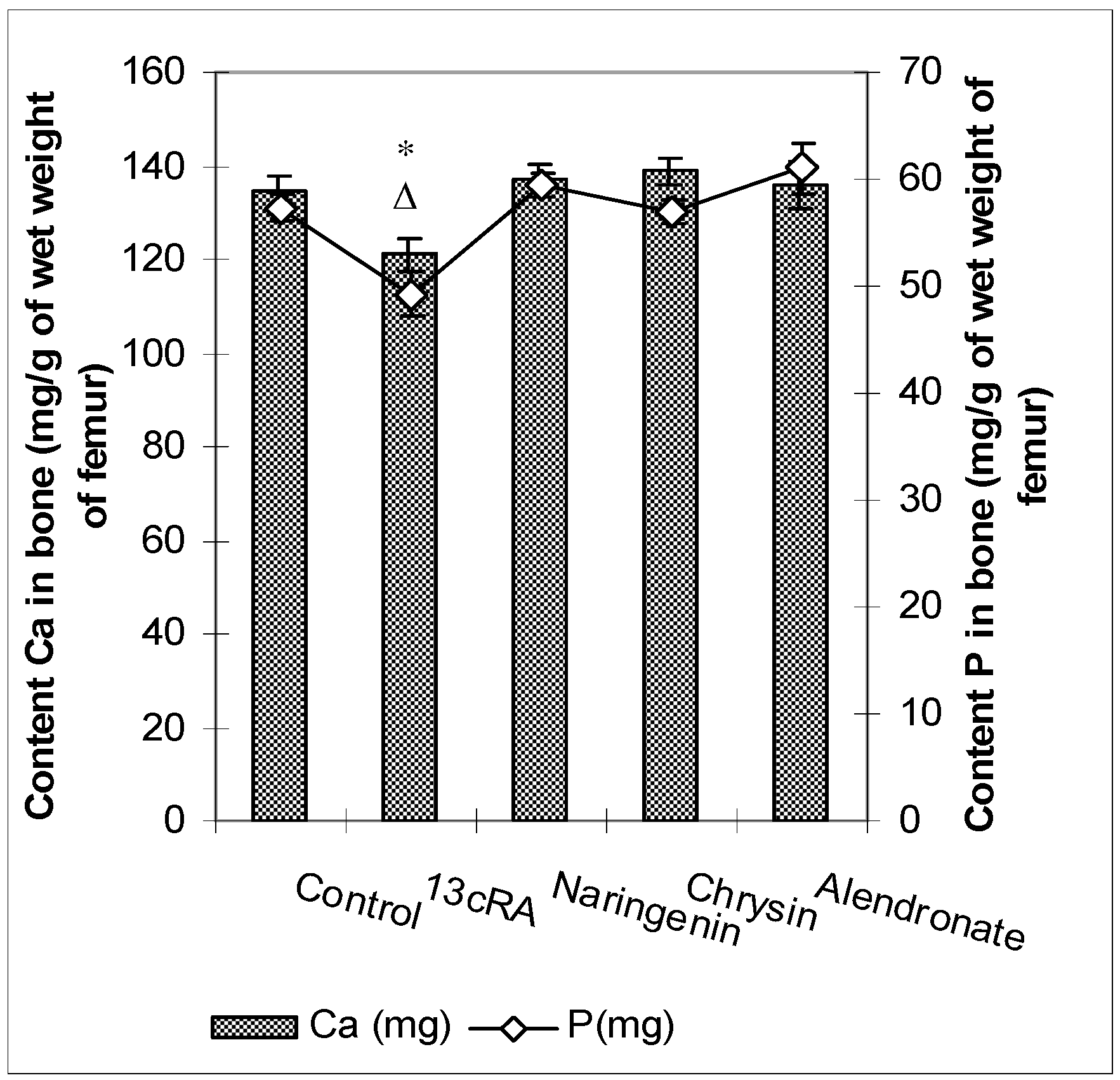
2.5. Ca and P Levels in the Femur and Serum
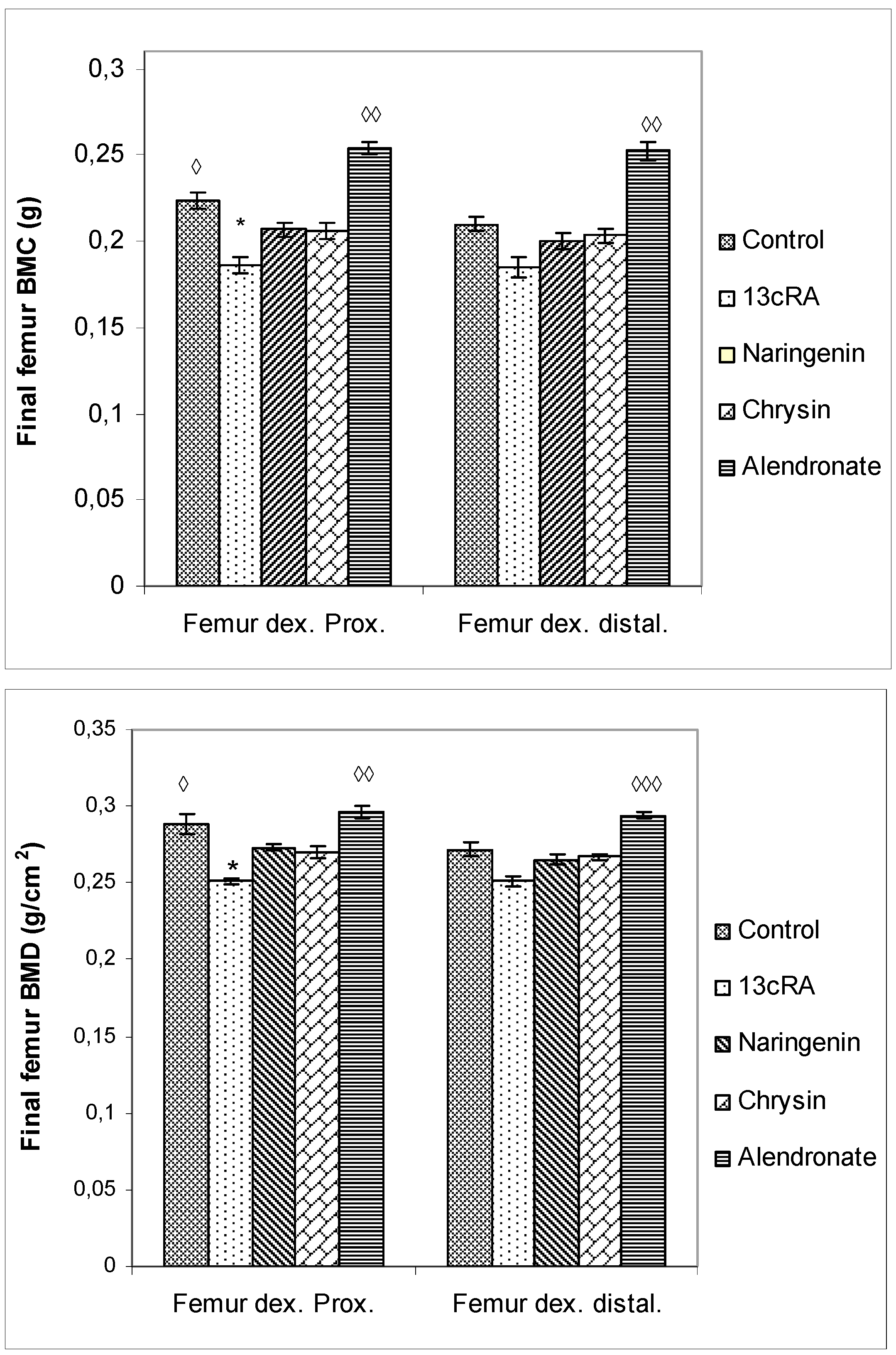
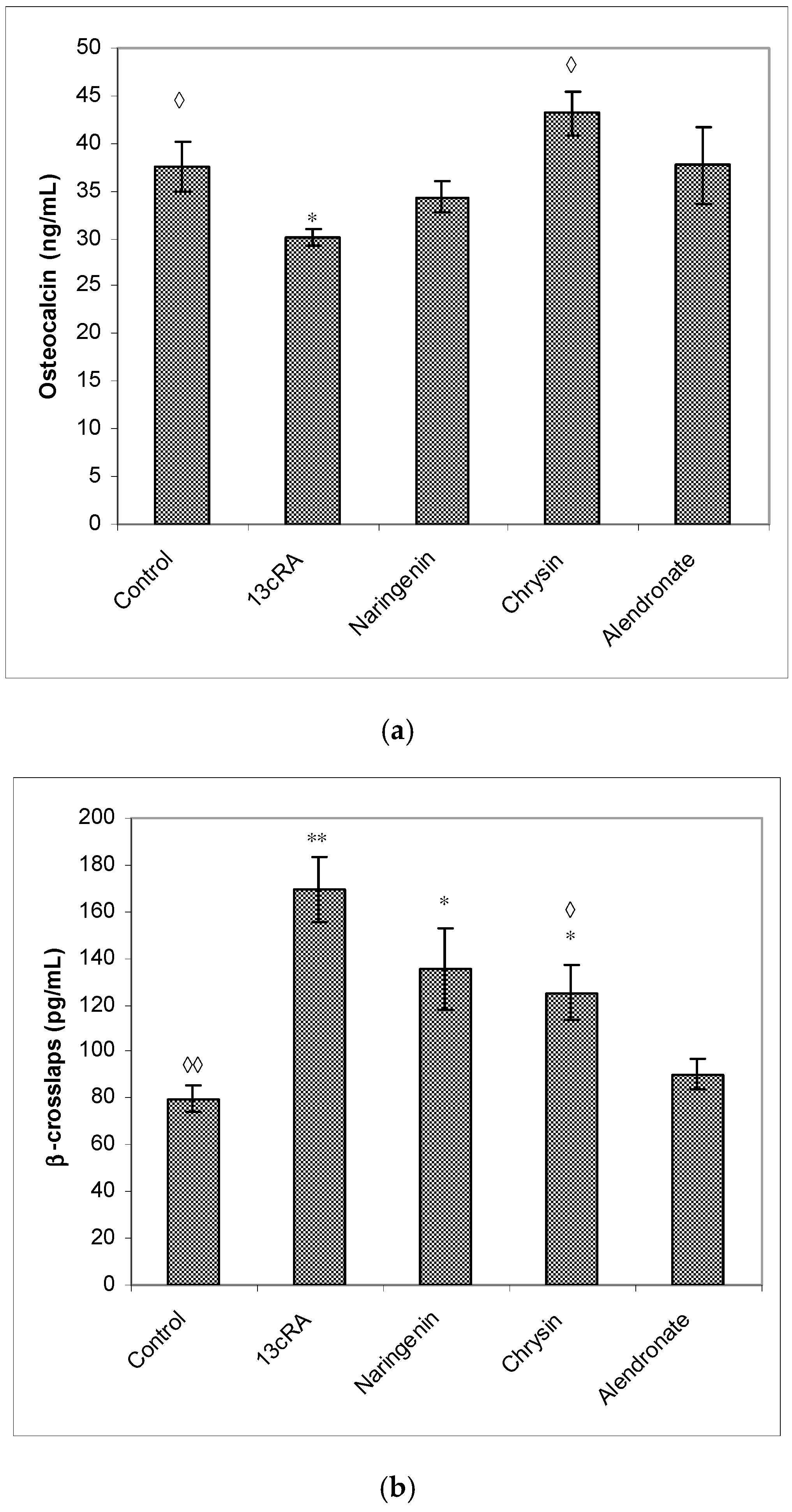
2.6. Bone Mineral Content, Bone Mineral Density, and Serum Markers of Bone Turnover
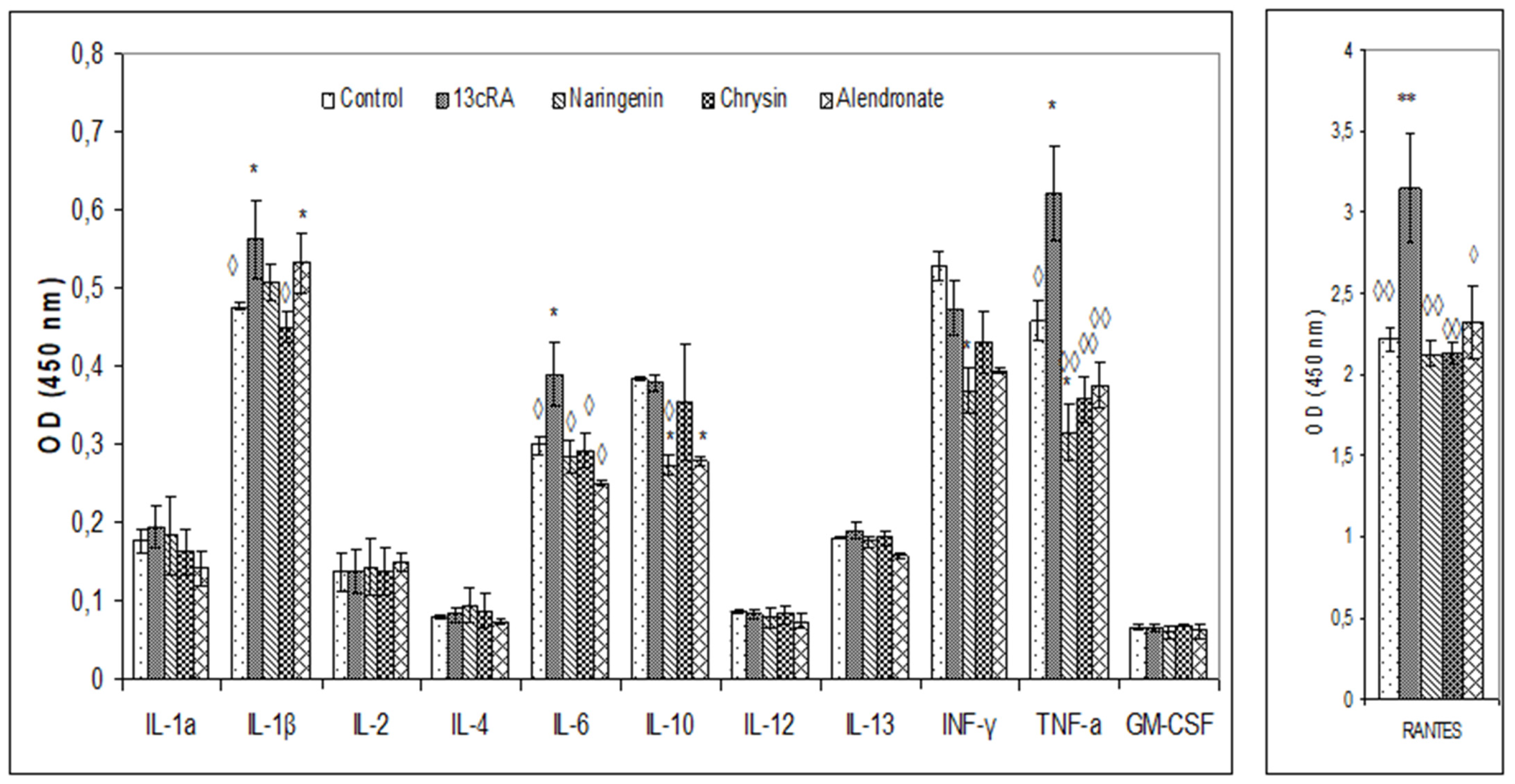
2.7. Changes in Biochemical and Haematological Parameters and Pro-Inflammatory Cytokines
2.8. Effect of Naringenin, Chrysin, and Alendronate on the Oxido-Reduction Status in Liver, Kidney, Spleen, and Ovary Tissues
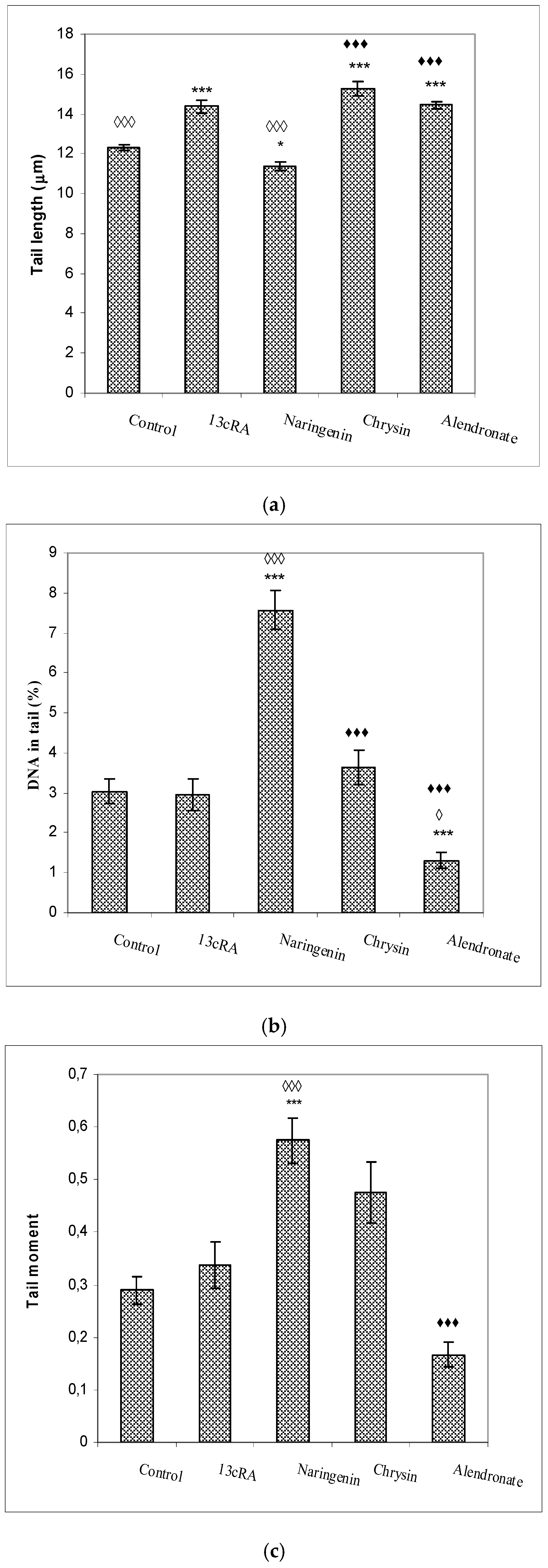
2.9. DNA Damage of Peripheral Blood Cells
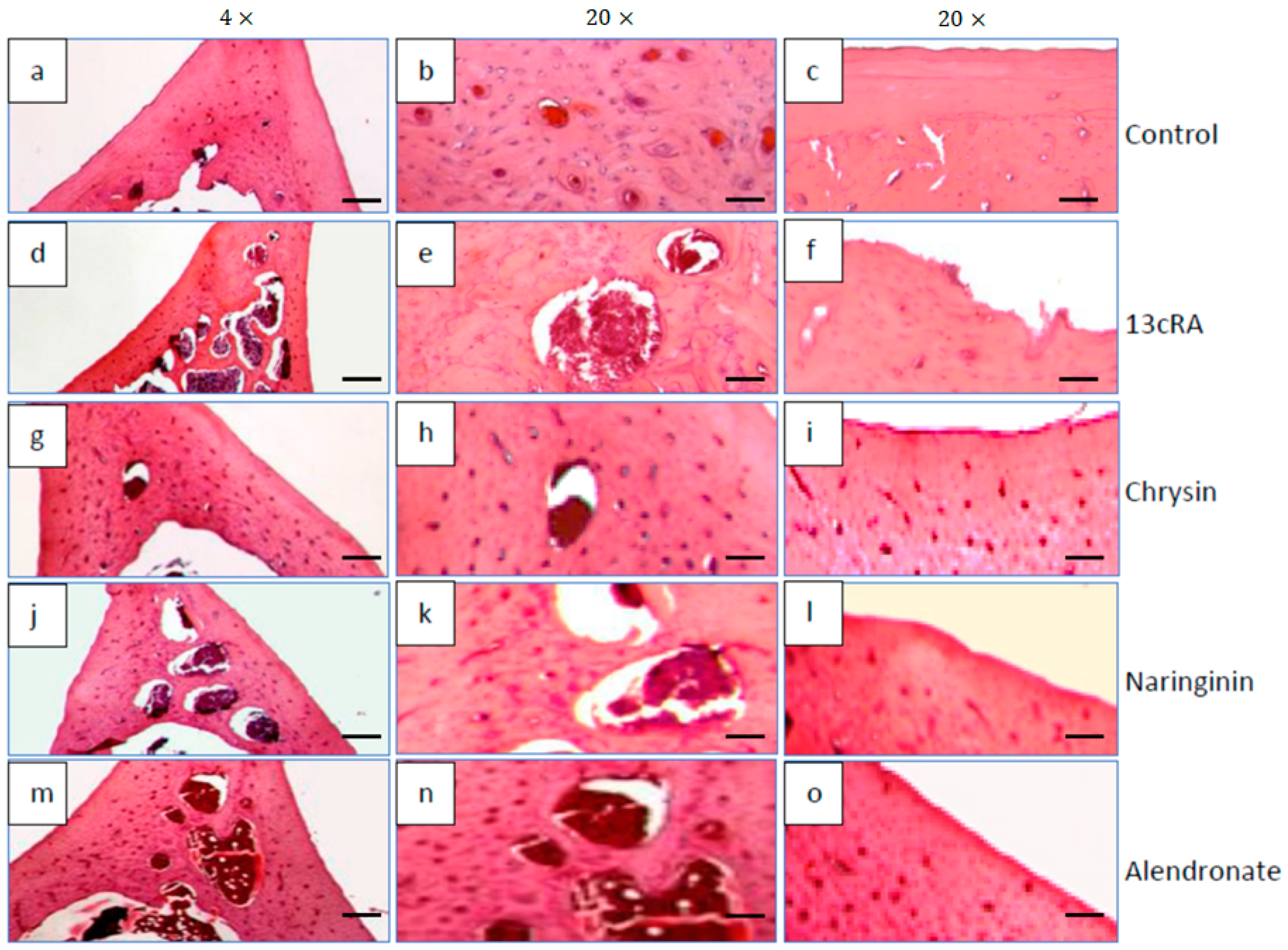
2.10. Histological Changes in Bone Tissue
3. Discussion
4. Material and Methods
4.1. Reagents
4.2. Chrysin and Naringenin
4.3. Experimental Animals, Study Design, and Organ Processing
4.4. Monitoring Weight Changes of Animals and Selected Organs
4.5. Bone Harvesting and Analysis of Bone Physical Parameters
4.6. Serum Markers of Bone Turnover
4.7. Analysis of Haematological and Biochemical Parameters
4.8. Histological Analysis
4.9. Tissue Preparations for Quantitative Analysis of Oxidative Stress Biomarkers
4.10. Analysis of Inflammatory Cytokines
4.11. The Alkaline Comet Assay
4.12. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Swaim, R.A.; Barner, J.C.; Brown, C.M. The relationship of calcium intake and exercise to osteoporosis health beliefs in postmenopausal women. Res. Soc. Adm. Pharm. 2008, 4, 153–163. [Google Scholar] [CrossRef]
- Byreddy, D.V.; Bouchonville, M.F., 2nd; Lewiecki, E.M. Drug-induced osteoporosis: From Fuller Albright to aromatase inhibitors. Climacteric 2015, 18, 39–46. [Google Scholar] [CrossRef]
- Panday, K.; Gona, A.; Humphrey, M.B. Medication-induced osteoporosis: Screening and treatment strategies. Ther. Adv. Musculoskelet. Dis. 2014, 6, 185–202. [Google Scholar] [CrossRef]
- Mitra, R. Adverse effects of corticosteroids on bone metabolism: A review. PM R 2011, 3, 466–471. [Google Scholar] [CrossRef]
- Fahmy, S.R.; Soliman, A.M. Oxidative Stress as a Risk Factor of Osteoporotic Model Induced by Vitamin A in Rats. Aust. J. Basic Appl. Sci. 2009, 3, 1559–1568. [Google Scholar]
- Kneissel, M.; Studer, A.; Cortesi, R.; Susa, M. Retinoid-induced bone thinning is caused by subperiosteal osteoclast activity in adult rodents. Bone 2005, 36, 202–214. [Google Scholar] [CrossRef]
- Scheven, B.A.; Hamilton, N.J. Retinoic acid and 1, 25-dihydroxy vitamin D stimulate osteoclast formation by different mechanisms. Bone 1990, 11, 53–59. [Google Scholar] [CrossRef]
- Oršolić, N.; Goluža, E.; Đikić, D.; Lisičić, D.; Sašilo, K.; Rođak, E.; Jeleč, Ž.; Lazarus, M.V.; Orct, T. Role of flavonoids on oxidative stress and mineral contents in the retinoic acid-induced bone loss model of rat. Eur. J. Nutr. 2014, 53, 1217–1227. [Google Scholar] [CrossRef]
- Yang, J.; Wu, N.; Peng, J.; Yang, X.; Guo, J.; Yin, S.; Wang, J. Prevention of retinoic acid-induced osteoporosis in mice by isoflavone-enriched soy protein. J. Sci. Food Agric. 2016, 96, 331–338. [Google Scholar] [CrossRef]
- Zhao, S.; Niu, F.; Xu, C.Y.; Liu, Y.; Ye, L.; Bi, G.B.; Chen, L.; Tian, G.; Nie, T.H. Diosgenin prevents bone loss on retinoic acid-induced osteoporosis in rats. Ir. J. Med. Sci. 2016, 185, 581–587. [Google Scholar] [CrossRef]
- Bonaccorsi, G.; Piva, I.; Greco, P.; Cervellati, C. Oxidative stress as a possible pathogenic cofactor of post-menopausal osteoporosis: Existing evidence in support of the axis oestrogen deficiency-redox imbalance-bone loss. Indian J. Med. Res. 2018, 147, 341–351. [Google Scholar]
- Weitzmann, M.N.; Pacifici, R. Estrogen regulation of immune cell bone interactions. Ann. N. Y. Acad. Sci. 2006, 1068, 256–274. [Google Scholar] [CrossRef]
- Oršolić, N.; Car, N. Quercetin and hyperthermia modulate cisplatin-induced DNA damage in tumor and normal tissues in vivo. Tumour Biol. 2014, 35, 6445–6454. [Google Scholar] [CrossRef]
- Oršolić, N.; Bašić, I. Honey bee products and their polyphenolic compounds in treatment of diabetes. Phytopharmacol. Ther. Values IV 2008, 22, 455–553. [Google Scholar]
- Oršolić, N.; Jeleč, Ž.; Nemrava, J.; Balta, V.; Gregorović, G.; Jeleč, D. The effect of quercetin on bone mineral status and markers of bone turnover in retinoic acid-induced osteoporosis. Pol. J. Food Nutr. Sci. 2018, 68, 149–162. [Google Scholar] [CrossRef]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef]
- Oršolić, N.; Skurić, J.; Đikić, Đ.; Stanić, G. Inhibitory effect of a propolis on Di-n-Propyl Disulfide or n-Hexyl salicilate-induced skin irritation, oxidative stress and inflammatory responses in mice. Fitoterapia 2014, 93, 18–30. [Google Scholar] [CrossRef]
- Oršolić, N.; Kunštić, M.; Kukolj, M.; Gračan, R.; Nemrava, J. Oxidative stress, polarization of macrophages and tumour angiogenesis: Efficacy of caffeic acid. Chem. Biol. Interact. 2016, 256, 111–124. [Google Scholar] [CrossRef]
- Sirovina, D.; Oršolić, N.; Zovko Končić, M.; Kovačević, G.; Benković, V.; Gregorović, G. Quercetin vs. chrysin: Effect on liver histopathology in diabetic mice. Hum. Exp. Toxicol. 2013, 2, 1058–1066. [Google Scholar] [CrossRef]
- Jembrek, M.J.; Vlainić, J.; Radovanović, V.; Erhardt, J.; Oršolić, N. Effects of copper overload in P19 neurons: Impairment of glutathione redox homeostasis and crosstalk between caspase and calpain protease systems in ROS-induced apoptosis. Biometals 2014, 27, 1303–1322. [Google Scholar] [CrossRef]
- Zorić, N.; Kopjar, N.; Kraljić, K.; Oršolić, N.; Tomić, S.; Kosalec, I. Olive leaf extract activity against Candida albicans and C. dubliniensis—the in vitro viability study. Acta Pharm. 2016, 66, 411–421. [Google Scholar] [CrossRef]
- Madeddu, C.; Gramignano, G.; Floris, C.; Murenu, G.; Sollai, G.; Macciò, A. Role of inflammation and oxidative stress in post-menopausal oestrogen-dependent breast cancer. J. Cell. Mol. Med. 2014, 18, 2519–2529. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.K.; Gupta, S.C.; Karelia, D.; Gilhooley, P.J.; Shakibaei, M.; Aggarwal, B.B. Dietary nutraceuticals as backbone for bone health. Biotechnol. Adv. 2018, 36, 1633–1648. [Google Scholar] [CrossRef] [PubMed]
- Oršolić, N.; Balta, V.; Odeh, D.; Mataković, M.; Skurić, J. Oxidative stress and inflammation caused by n-Hexyl salicylate in mouse skin: The effectiveness of flavonoids. Int. J. Phytomed. 2017, 9, 279–288. [Google Scholar] [CrossRef]
- Zeng, W.; Yan, Y.; Zhang, F.; Zhang, C.; Liang, W. Chrysin promotes osteogenic differentiation via ERK/MAPK activation. Protein Cell 2013, 4, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Mohd Effendy, N.; Mohamed, N.; Muhammad, N.; Mohamad, I.N.; Shuid, A.N. The effects of tualang honey on bone metabolism of postmenopausal women. Evid.-Based Complement. Altern. Med. 2012, 2012, 938574. [Google Scholar] [CrossRef]
- Guo, D.; Wang, J.; Wang, X.; Luo, H.; Zhang, H.; Cao, D.; Chen, L.; Huang, N. Double directional adjusting estrogenic effect of naringin from Rhizoma drynariae (Gusuibu). J. Ethnopharmacol. 2011, 138, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhen, L.; Zhang, G.; Wong, M.S.; Qin, L.; Yao, X. Osteogenic effects of flavonoid aglycones from an osteoprotective fraction of Drynaria fortunei--an in vitro efficacy study. Phytomedicine 2011, 18, 868–872. [Google Scholar] [CrossRef]
- Liao, E.Y.; Luo, X.H.; Wang, W.B.; Wu, X.P.; Zhou, H.D.; Dai, R.C.; Liao, H.-J.; Yang, C. Effects of Different Nylestriol/Levonorgestrel Dosages on Bone Metabolism in Female Sprague–Dawley Rats with Retinoic Acid-Induced Osteoporosis. Endocr. Res. 2003, 29, 23–42. [Google Scholar] [CrossRef]
- Toker, H.; Ozdemir, H.; Balci Yuce, H.; Goze, F. The effect of boron on alveolar bone loss in osteoporotic rats. J. Dent. Sci. 2016, 11, 331–337. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Dar, H.Y.; Mishra, P.K. Immunoporosis: Immunology of Osteoporosis-Role of T Cells. Front. Immunol. 2018, 9, 657. [Google Scholar] [CrossRef] [PubMed]
- Ajuebor, M.N.; Hogaboam, C.M.; Kunkel, S.L.; Proudfoot, A.E.; Wallace, J.L. The chemokine RANTES is a crucial mediator of the progression from acute to chronic colitis in the rat. J. Immunol. 2001, 166, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Lechner, J.; Aschoff, J.; Rudi, T. The vitamin D receptor and the etiology of RANTES/CCL-expressive fatty-degenerative osteolysis of the jawbone: An interface between osteoimmunology and bone metabolism. Int. J. Gen. Med. 2018, 11, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Whisner, C.M.; Castillo, L.F. Prebiotics, bone and mineral metabolism. Calcif. Tissue Int. 2017, 102, 443–479. [Google Scholar] [CrossRef]
- McCabe, L.; Britton, R.A.; Parameswaran, N. Prebiotic and probiotic regulation of bone health: Role of the intestine and its microbiome. Curr. Osteoporos. Rep. 2015, 13, 363–371. [Google Scholar] [CrossRef]
- Franceschi, R.T.; Ge, C.; Xiao, G.; Roca, H.; Jiang, D. Transcriptional regulation of osteoblasts. Ann. N. Y. Acad. Sci. 2007, 1116, 196–207. [Google Scholar] [CrossRef]
- Jana, K.; Yin, X.; Schiffer, R.B.; Chen, J.J.; Pandey, A.K.; Stocco, D.M.; Grammas, P.; Wang, X.J. Chrysin, a natural flavonoid enhances steroidogenesis and steroidogenic acute regulatory protein gene expression in mouse Leydig cells. J. Endocrinol. 2008, 197, 315–323. [Google Scholar] [CrossRef]
- Mohamad, N.V.; Soelaiman, I.N.; Chin, K.Y. A concise review of testosterone and bone health. Clin. Interv. Aging 2016, 11, 1317–1324. [Google Scholar] [CrossRef]
- Breinholt, V.M.; Svendsen, G.W.; Dragsted, L.O.; Hossaini, A. The citrus-derived flavonoid naringenin exerts uterotrophic effects in female mice at human relevant doses. Basic Clin. Pharmacol. Toxicol. 2004, 94, 30–36. [Google Scholar] [CrossRef]
- Assini, J.M.; Mulvihill, E.E.; Huff, M.W. Citrus flavonoids and lipid metabolism. Curr. Opin. Lipidol. 2013, 24, 34–40. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Rahman, M.M.; Uddin, S.J.; Reza, H.M.; Sarker, S.D. Effect of citrus flavonoids, naringin and naringenin, on metabolic syndrome and their mechanisms of action. Adv. Nutr. 2014, 5, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.E.; Breinholt, V.M. Non-nutritive bioactive food constituents of plants: Bioavailability of flavonoids. Int. J. Vitam. Nutr. Res. 2003, 73, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Erlund, I.; Meririnne, E.; Alfthan, G.; Aro, A. Plasma kinetics and urinary excretion of the flavanones naringenin and hesperetin in humans after ingestion of orange juice and grapefruit juice. J. Nutr. 2001, 131, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Istasse, T.; Jacquet, N.; Berchem, T.; Haubruge, E.; Nguyen, B.K.; Richel, A. Extraction of Honey Polyphenols: Method Development and Evidence of Cis Isomerization. Anal. Chem. Insights 2016, 11, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Premratanachai, P.; Chanchao, C. Review of the anticancer activities of bee products. Asian Pac. J. Trop. Biomed. 2014, 4, 337–344. [Google Scholar] [CrossRef]
- Samarghandian, S.; Farkhondeh, T.; Azimi-Nezhad, M. Protective Effects of Chrysin Against Drugs and Toxic Agents. Dose-Response 2017, 15, 1559325817711782. [Google Scholar] [CrossRef]
- Luo, K.; Ma, S.; Guo, J.; Huang, Y.; Yan, F.; Xiao, Y. Association between postmenopausal osteoporosis and experimental periodontitis. Biomed. Res. Int. 2014, 2014, 316134. [Google Scholar] [CrossRef]
- Kacamak, P.; Ozogul, C.; Saribas, G.S.; Akarca, D.; Saadet, O.; Emniyet, S.; Asiye, A. The Effect of Isotretinoin on Oocyte Maturation in Adolescent Female Rats. Gynecol. Obstet. Investig. 2020, 85, 27–335. [Google Scholar] [CrossRef]
- Abali, R.; Yuksel, M.A.; Aktas, C.; Celik, C.; Guzel, S.; Erfan, G.; Sahin, O. Decreased ovarian reserve in female Sprague–Dawley rats induced by isotretinoin (retinoic acid) exposure. Reprod. BioMed. Online 2013, 27, 184–191. [Google Scholar] [CrossRef][Green Version]
- Perazzella, M.A.; Markowitz, G.S. Bisphosphonate Nephrotoxicity. Kidney Int. 2008, 74, 1385–1393. [Google Scholar] [CrossRef]
- Saita, Y.; Ishijima, M.; Kaneko, K. Atypical femoral fractures and bisphosphonate use: Current evidence and clinical implications. Ther. Adv. Chronic Dis. 2015, 6, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.H.; Huang, C.C.; Chan, W.L.; Huang, P.H.; Chen, T.J.; Chung, C.M.; Lin, S.-J.; Chen, J.-W.; Leu, H.-B. Oral alendronate use and risk of cancer in postmenopausal women with osteoporosis: A nationwide study. J. Bone Miner. Res. 2012, 27, 1951–1958. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Yu, Z.; Funayama, H.; Yamaguchi, K.; Sasano, T.; Sugawara, S.; Endo, Y. Histidine decarboxylase-stimulating and inflammatory effects of alendronate in mice: Involvement of mevalonate pathway, TNFalpha, macrophages, and T-cells. Int. Immunopharmacol. 2007, 7, 152–1561. [Google Scholar] [CrossRef] [PubMed]
- Benghuzzi, H.; Mohamed, A.; Aneja, A.; Cameron, J.A.; Tucci, M. The effects of sustained delivery of alendronate on the kidney in ovariectomized female rats. Biomed. Sci. Instrum. 2012, 48, 35–42. [Google Scholar]
- Garimella, P.S.; Rennke, H.G.; Strom, J.A. Alendronate associated focal segmental glomerulosclerosis: A case report and review of the literature. CEN Case Rep. 2015, 4, 20–23. [Google Scholar] [CrossRef]
- Russell, R.G.; Watts, N.B.; Ebetino, F.H.; Rogers, M.J. Mechanisms of action of bisphosphonates: Similarities and differences and their potential influence on clinical efficacy. Osteoporos. Int. 2008, 19, 733–759. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups a | Femur Physical Characteristics (cm/100 g) (X ± SE) | ||||||
|---|---|---|---|---|---|---|---|
| AP ϕ Proximal Epiphysis | ML ϕ Proximal Epiphysis | AP ϕ Mid-Diaphysis | ML ϕ Mid-Diaphysis | AP ϕ Distal Epiphysis | ML Distal Epiphysis | Femur Length Trochanter-Condyle | |
| Control | 0.153 ± 0.004 | 0.223 ± 0.010 | 0.129 ± 0.004 | 0.172 ± 0.021 ◊ | 0.154 ± 0.009 | 0.207 ± 0.015 | 1.405 ± 0.050 |
| 13cRA | 0.137 ± 0.009 | 0.214 ± 0.011 | 0.118 ± 0.005 | 0.141 ± 0.010 * | 0.150 ± 0.008 | 0.213 ± 0.009 | 1.325 ± 0.044 |
| Naringenin | 0.152 ± 0.004 | 0.209 ± 0.009 | 0.136 ± 0.009 | 0.148 ± 0.010 | 0.153 ± 0.010 | 0.207 ± 0.016 | 1.489 ± 0.065 ◊ |
| Chrysin | 0.140 ± 0.010 | 0.198 ± 0.012 | 0.120 ± 0.009 | 0.145 ± 0.019 | 0.144 ± 0.017 | 0.193 ± 0.011 | 1.389 ± 0.102 |
| Alendronate | 0.146 ± 0.004 | 0.222 ± 0.005 | 0.128 ± 0.005 | 0.149 ± 0.004 | 0.169 ± 0.015 | 0.228 ± 0.001 | 1.390 ± 0.036 |
| Groups a | Biochemical Parameters-Enzymes (X ± SE) | |||||
|---|---|---|---|---|---|---|
| AST (U/L) | ALT (U/L) | ALP (U/L) | GGT (U/L) | LDH (U/L) | Amylase (U/L) | |
| Control | 78.33 ± 7.73 | 46.40 ± 9.52 | 170.13 ± 24.54 ◊ | 0.00 ± 0.00 | 464.50 ± 130.97 | 1751.00 ± 609.96 |
| 13cRA | 96.16 ± 13.34 | 43.16 ± 4.66 | 224.00 ± 15.01 * | 0.66 ± 0.51 | 725.66 ± 157.59 * | 2046.16 ± 213.69 |
| Naringenin | 74.00 ± 5.25 ◊ | 41.00 ± 3.89 | 147.50 ± 30.09 ◊ | 0.33 ± 0.51 | 201.50 ± 97.65 ◊◊◊ | 1897.00 ± 311.35 |
| Chrysin | 83.00 ± 4.00 | 41.00 ± 4.19 | 186.33 ± 45.60 | 0.16 ± 0.40 | 325.16 ± 121.79 ◊ | 1912.16 ± 510.88 |
| Alendronate | 87.66 ± 7.50 | 42.00 ± 2.64 | 115.00 ± 12.00 ◊◊ | 0.33 ± 0.57 | 402.00 ± 122.74 | 2184.00 ± 419.06 |
| Groups a | Leukocytes (×109 L−1) | Differential Blood Count (%) (X ± SE) | ||||
|---|---|---|---|---|---|---|
| Lymphocytes | Monocytes | Neutrophils | Basophils | Eosinophils | ||
| Control | 2.70 ± 1.17 | 79.06 ± 7.99 | 0.42 ± 0.21 | 13.46 ± 2.97 | 0.38 ± 0.21 | 0.61 ± 0.27 |
| 13cRA | 3.88 ± 1.12 | 83.96 ± 2.74 | 0.45 ± 0.16 | 15.18 ± 2.28 | 0.55 ± 0.17 | 0.73 ± 0.36 |
| Naringenin | 3.85 ± 0.47 | 86.95 ± 2.02 | 0.38 ± 0.21 | 11.45 ± 2.37 | 0.36 ± 0.23 | 0.85 ± 0.38 |
| Chrysin | 4.51 ± 1.33 | 79.55 ± 6.86 | 0.36 ± 0.22 | 15.33 ± 3.29 | 0.46 ± 0.15 | 1.05 ± 0.25 |
| Alendronate | 5.35 ± 1.36 * | 70.16 ± 3.29 | 0.86 ± 0.20 * | 28.00 ± 4.33 * | 1.23 ± 0.50 | 0.73 ± 0.41 |
| Groups a | MDA (nmol/mg Proteins) | |||
|---|---|---|---|---|
| Liver | Spleen | Kidney | Ovary | |
| Control | 10.22 ± 0.56 ◊ | 16.24 ± 1.05 | 4.09 ± 0.60 ◊◊ | 12.12 ± 1.63 ◊◊ |
| 13cRA | 15.58 ± 0.63 * | 17.61 ± 1.56 | 12.33 ± 0.72 ** | 35.78 ± 3.45 ** |
| Naringenin | 13.36 ± 0.33 | 13.66 ± 0.92 ♦ | 3.90 ± 0.46 ◊◊ | 20.06 ± 2.01 ◊ |
| Chrysin | 14.04 ± 0.42 | 19.11 ± 1.70 | 4.81 ± 0.53 ◊◊ | 28.99 ± 1.33 *◊ |
| Alendronate | 18.05 ± 0.83 ** | 23.62 ± 2.15 | 9.84 ± 0.87 * | 37.41 ± 2.14 ** |
| GSH (µg/mg proteins) | ||||
| Control | 8.90 ± 0.71 | 14.78 ± 2.25 | 8.46 ± 0.11 ◊ | 0.95 ± 0.15 ◊ |
| 13cRA | 5.65 ± 0.54 | 8.39 ± 0.38 | 3.61 ± 0.32 * | 0.20 ± 0.02 * |
| Naringenin | 7.08 ± 1.05 | 15.11 ± 3.18 | 10.07 ± 0.54 ◊ | 0.56 ± 0.10 |
| Chrysin | 7.64 ± 1.63 | 13.38 ± 1.77 | 6.75 ± 0.69 | 1.01 ± 0.09 ◊ |
| Alendronate | 5.09 ± 1.20 | 12.69 ± 1.27 | 10.63 ± 0.61 ◊◊ | 0.37 ± 0.20 |
| SOD (U/mg proteins) | ||||
| Control | 3.67 ± 0.69 ◊ | 5.38 ± 0.52 | 4.07 ± 0.48 | 15.46 ± 1.81 ♦♦ |
| 13cRA | 1.66 ± 0.13 * | 4.22 ± 1.14 | 3.94 ± 0.52 | 7.44 ± 0.69 |
| Naringenin | 4.61 ± 0.35 ◊◊♦ | 9.03 ± 1.80 ◊ | 4.56 ± 0.86 | 9.28 ± 1.01 ♦ |
| Chrysin | 2.50 ± 0.22 | 4.93 ± 1.06 | 4.06 ± 1.40 | 11.54 ± 1.81 ♦ |
| Alendronate | 1.77 ± 0.38 | 5.29 ± 1.37 | 7.74 ± 1.75 | 2.46 ± 0.43 ** |
| CAT (U/mg proteins) | ||||
| Control | 4.98 ± 0.42 | 4.22 ± 0.28 | 21.99 ± 1.58 | 14.03 ± 0.06 |
| 13cRA | 4.67 ± 0.61 | 2.76 ± 0.40 | 8.94 ± 0.73 | 11.64 ± 1.73 |
| Naringenin | 4.45 ± 0.62 | 4.09 ± 0.87 | 43.77 ± 5.71 ◊◊ | 15.92 ± 1.85 |
| Chrysin | 4.70 ± 0.34 | 2.82 ± 0.33 ♦ | 41.24 ± 4.87 ◊◊ | 24.53 ± 0.24 **◊ |
| Alendronate | 4.08 ± 0.86 | 6.99 ± 0.56 ◊ | 66.45 ± 4.21 ◊◊◊ | 20.92 ± 1.10 *◊ |
| Parameter | Method | |
|---|---|---|
| Serum biochemical parameters | Aspartate aminotransferase (AST), Alanine aminotransferase (ALT), Alkaline phosphatase (ALP), Glutamyl transferase (GGT), Amylase Urea, creatinine, Blood glucose levels (glucose), Lactate dehydrogenase (LDH), Total protein and serum Ca and P levels | Colorimetric method |
| Hematological parameters | Erythrocytes (E), The average cellular volume of erythrocytes (MCV), Haemoglobin (Hgb), Haematocrit (Hct), Mean cell haemoglobin (MCH), Mean cell Haemoglobin concentration (MCHC), Total leukocyte count (L), and The total number of platelets (Plt) | Colorimetric method |
| Serum Inflammatory cytokines | IL1α, IL1β, IL2, IL4, IL6, IL10, IL12, IL17A, IFNγ, TNFα, GM-CSF, and RANTES | ELISA method |
| Biochemical bone parameters | Ca and P analyses | Atomic absorption spectrophotometry |
| Bone mineral density (BMD) | Dual-energy X-ray absorptiometry (DXA). | |
| Serum markers of bone turnover Osteocalcin (OC) β-CrossLaps | Electrochemiluminescence “ECLIA” method | |
| Bone physical parameters | Relative bone weight index, Length of the bone (large trochanter-condyle), The distal and proximal epiphyseal diameters of the femur were measured in mediolateral (ML) and anteroposterior directions (AP) | Digital scale Small bone caliper |
| Histological parameters | Histological analyses | Haematoxylin and eosin (HE) methods |
| Oxidative stress parameters | Lipid peroxidation level (MDA), Glutathione level (GSH) Superoxide dismutase (SOD) activity Catalase (CAT) activity | Colorimetric method |
| DNA damage | Alkaline Comet Assay |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oršolić, N.; Nemrava, J.; Jeleč, Ž.; Kukolj, M.; Odeh, D.; Jakopović, B.; Jazvinšćak Jembrek, M.; Bagatin, T.; Fureš, R.; Bagatin, D. Antioxidative and Anti-Inflammatory Activities of Chrysin and Naringenin in a Drug-Induced Bone Loss Model in Rats. Int. J. Mol. Sci. 2022, 23, 2872. https://doi.org/10.3390/ijms23052872
Oršolić N, Nemrava J, Jeleč Ž, Kukolj M, Odeh D, Jakopović B, Jazvinšćak Jembrek M, Bagatin T, Fureš R, Bagatin D. Antioxidative and Anti-Inflammatory Activities of Chrysin and Naringenin in a Drug-Induced Bone Loss Model in Rats. International Journal of Molecular Sciences. 2022; 23(5):2872. https://doi.org/10.3390/ijms23052872
Chicago/Turabian StyleOršolić, Nada, Johann Nemrava, Željko Jeleč, Marina Kukolj, Dyana Odeh, Boris Jakopović, Maja Jazvinšćak Jembrek, Tomica Bagatin, Rajko Fureš, and Dinko Bagatin. 2022. "Antioxidative and Anti-Inflammatory Activities of Chrysin and Naringenin in a Drug-Induced Bone Loss Model in Rats" International Journal of Molecular Sciences 23, no. 5: 2872. https://doi.org/10.3390/ijms23052872
APA StyleOršolić, N., Nemrava, J., Jeleč, Ž., Kukolj, M., Odeh, D., Jakopović, B., Jazvinšćak Jembrek, M., Bagatin, T., Fureš, R., & Bagatin, D. (2022). Antioxidative and Anti-Inflammatory Activities of Chrysin and Naringenin in a Drug-Induced Bone Loss Model in Rats. International Journal of Molecular Sciences, 23(5), 2872. https://doi.org/10.3390/ijms23052872