
Estrogen Receptors Mediated Negative Effects of Estrogens and Xenoestrogens in Teleost Fishes—Review

 ,
,  ,
,  ,
,
Abstract
1. Introduction

2. Carcinogenic Potential
3. Impact on Circulatory System
4. Impact on Metabolism
5. Impact on Immune System
6. Impact on Reproductive System
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jensen, E.V.; Jordan, V.C. The estrogen receptor: A model for molecular medicine. Clin. Cancer Res. 2003, 9, 1980–1989. [Google Scholar]
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef]
- Olde, B.; Leeb-Lundberg, L.F. GPR30/GPER1: Searching for a role in estrogen physiology. Trends Endocrinol. Metab. 2009, 20, 409–416. [Google Scholar] [CrossRef]
- Ignatov, T.; Modl, S.; Thulig, M.; Weißenborn, C.; Treeck, O.; Ortmann, O.; Ignatov, A. GPER-1 acts as a tumor suppressor in ovarian cancer. J. Ovarian Res. 2013, 6, 51. [Google Scholar] [CrossRef]
- Toran-Allerand, C.D.; Guan, X.; Maclusky, N.J.; Horvath, T.L.; Diano, S.; Singh, M.; Tinnikov, A.A. ER-X: A novel, plasma membrane-associated, putative Estrogen Receptor that is regulated during development and after ischemic brain injury. J. Neurosci. 2002, 22, 8391–8401. [Google Scholar] [CrossRef]
- Micevych, P.E.; Kelly, J.M. Membrane estrogen receptor regulation of hypothalamic function. Neuroendocrinology 2021, 96, 103–110. [Google Scholar] [CrossRef]
- Prossnitz, E.R.; Barton, M. The G-Protein-Coupled Estrogen Receptor GPER in health and disease. Nat. Rev. Endocrinol. 2011, 7, 715–726. [Google Scholar] [CrossRef]
- Qiu, J.; Bosch, M.A.; Tobias, S.C.; Grandy, D.K.; Scanlan, T.S.; Rønnekleiv, O.K.; Kelly, M.J. Rapid signaling of estrogen in hypothalamic neurons involves a novel G-Protein-Coupled estrogen receptor that activates protein kinase C. J. Neurosci. 2003, 23, 9529–9540. [Google Scholar] [CrossRef]
- O’Lone, R.; Frith, M.C.; Karlsson, E.K.; Hansen, U. Genomic targets of nuclear estrogen receptors. Mol. Endocrinol. 2004, 18, 1859–1875. [Google Scholar] [CrossRef]
- Jacob, J.; Sebastian, K.S.; Devassy, S.; Priyadarsini, L.; Farook, M.F.; Shameem, A.; Thampan, R.V. Membrane Estrogen Receptors: Genomic actions and post transcriptional regulation. Mol. Cell. Endocrinol. 2006, 246, 34–41. [Google Scholar] [CrossRef]
- Lecomte, S.; Demay, F.; Ferrière, F.; Pakdel, F. Phytochemicals targeting estrogen receptors: Beneficial rather than adverse effects? Int. J. Mol. Sci. 2017, 18, 1381. [Google Scholar] [CrossRef]
- Acconcia, F.; Kumar, R. Signaling regulation of genomic and nongenomic functions of estrogen receptors. Cancer Lett. 2006, 238, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Paris, M.; Pettersson, K.; Schubert, M.; Bertrand, S.; Pongratz, I.; Escriva, H.; Laudet, V. An amphioxus orthologue of the estrogen receptor that does not bind estradiol: Insights into estrogen receptor evolution. BMC Evol. Biol. 2008, 8, 219. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, M.B.; Godwin, J.; Crews, D.; Thomas, P. The distributions of the duplicate oestrogen receptors ER-βa and ER-βb in the forebrain of the Atlantic croaker (Micropogonias undulatus): Evidence for subfunctionalization after gene duplication. Proc. R. Soc. B: Biol. Sci. 2005, 272, 633–641. [Google Scholar] [CrossRef]
- Pinto, P.I.; Andrade, A.R.; Estêvão, M.D.; Alvarado, M.V.; Felip, A.; Power, D.M. Duplicated membrane estrogen receptors in the European sea bass (Dicentrarchus labrax): Phylogeny, expression and regulation throughout the reproductive cycle. J. Steroid Biochem. Mol. Biol. 2018, 178, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Booth, A.N.; Bickoff, E.M.; Kohler, G.O. Estrogen-like activity in vegetable oils and mill by-products. Science 1960, 131, 1807–1808. [Google Scholar] [CrossRef]
- Ostrovsky, D.; Kitts, W.D. Estrogen-Like substances in legumes and grasses: The influence of fractionation and route of administration on the estrogenic activity of plant materials. Can. J. Bioch. Physi. 1962, 40, 159–164. [Google Scholar] [CrossRef]
- Bennetts, H.W.; Uuderwood, E.J.; Shier, F.L. A specific breeding problem of sheep on subterranean clover pastures in western australia. Aust. Vet. J. 1946, 22, 348–352. [Google Scholar] [CrossRef]
- Paterni, I.; Granchi, C.; Minutolo, F. Risks and benefits related to alimentary exposure to xenoestrogens. Crit. Rev. Food Sci. Nutr. 2017, 57, 3384–3404. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; Ropero, A.B.; Soriano, S.; García-Arévalo, M.; Ripoll, C.; Fuentes, E.; Nadal, Á. Bisphenol-A acts as a potent estrogen via non-classical estrogen triggered pathways. Molecul. Cell. Endocrin. 2012, 355, 201–207. [Google Scholar] [CrossRef]
- Cao, L.Y.; Ren, X.M.; Guo, L.H. Estrogen-related receptor Γ is a novel target for lower-chlorinated polychlorinated biphenyls and their hydroxylated and sulfated metabolites. Environ. Pollut. 2019, 254, 113088. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Ko, Y.; Pang, C.; Ko, Y.J.; Choi, Y.K.; Kim, K.H.; Kang, K.S. Estrogenic activity of mycoestrogen (3β, 5α, 22E)-Ergost-22-En-3-Ol via estrogen receptor α-dependent signaling pathways in MCF-7 cells. Molecules 2022, 27, 36. [Google Scholar] [CrossRef] [PubMed]
- Kinkade, C.W.; Rivera-Núñez, Z.; Gorcyzca, L.; Aleksunes, L.M.; Barrett, E.S. Impact of fusarium-derived mycoestrogens on female reproduction: A systematic review. Toxins 2021, 13, 373. [Google Scholar] [CrossRef] [PubMed]
- Černá, T.; Ezechiáš, M.; Semerád, J.; Grasserová, A.; Cajthaml, T. Evaluation of estrogenic and antiestrogenic activity in sludge and explanation of individual compound contributions. J. Hazard. Mater. 2022, 423, 127108. [Google Scholar] [CrossRef] [PubMed]
- Shirdel, I.; Kalbassi, M.R.; Esmaeilbeigi, M.; Tinoush, B. Disruptive effects of nonylphenol on reproductive hormones, antioxidant enzymes, and histology of liver, kidney and gonads in caspian trout smolts. Compar. Biochem. Physio. Part C Toxicol. Pharmacol. 2020, 232, 108756. [Google Scholar] [CrossRef] [PubMed]
- Dobbs, R.W.; Malhotra, N.R.; Greenwald, D.T.; Wang, A.Y.; Prins, G.S.; Abern, M.R. Estrogens and prostate cancer. Prostate Cancer Prostatic Dis. 2019, 22, 185–194. [Google Scholar] [CrossRef]
- Gore, A.C. (Ed.) Endocrine-Disrupting Chemicals: From Basic Research to Clinical Practice; Humana Press: Totowa, NJ, USA, 2007; Volume 8. [Google Scholar]
- Ronchetti, S.A.; Miler, E.A.; Duvilanski, B.H.; Cabilla, J.P. Cadmium mimics estrogen-driven cell proliferation and prolactin secretion from anterior pituitary cells. PLoS ONE 2013, 8, e81101. [Google Scholar] [CrossRef]
- Georgescu, B.; Georgescu, C.; Dărăban, S.; Bouaru, A.; Paşcalău, S. Heavy metals acting as endocrine disrupters. Sci. Pap. Anim. Sci. Biotechnol. 2011, 44, 89–93. [Google Scholar]
- Chmielewski, J.; Łuszczki, J.; Czarny-Działak, M.; Dutkiewicz, E.; Król, H.; Gworek, B.; Nowak-Starz, G. Environmental exposition to xenoestrogens and related health effects. J. Elem. 2021, 26, 717–730. [Google Scholar] [CrossRef]
- Silva, I.P.; Brito, D.C.C.; Silva, T.E.S.; Silva, R.F.; Guedes, M.I.F.; Silva, J.Y.G.; Figueiredo, J.R. In vitro exposure of sheep ovarian tissue to the xenoestrogens zearalenone and enterolactone: Effects on preantral follicles. Theriogenology 2021, 174, 124–130. [Google Scholar] [CrossRef]
- Ratajczak-Wrona, W.; Rusak, M.; Nowak, K.; Dabrowska, M.; Radziwon, P.; Jablonska, E. Effect of bisphenol a on human neutrophils immunophenotype. Sci. Rep. 2020, 10, 3083. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Li, Y.W.; Chen, Q.L.; Liu, Z.H. Long-term exposure of xenoestrogens with environmental relevant concentrations disrupted spermatogenesis of zebrafish through altering sex hormone balance, stimulating germ cell proliferation, meiosis and enhancing apoptosis. Environ. Pollut. 2019, 244, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Mas, M.; Roca, P. Phytoestrogens for cancer prevention and treatment. Biology 2020, 9, 427. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.J.; Hsu, Y.L.; Huang, Y.F.; Tsai, E.M. Molecular mechanisms of anticancer effects of phytoestrogens in breast cancer. Curr. Protein Pept. Sci. 2018, 19, 323–332. [Google Scholar] [CrossRef]
- Wang, X.; Ha, D.; Yoshitake, R.; Chan, Y.S.; Sadava, D.; Chen, S. Exploring the biological activity and mechanism of xenoestrogens and phytoestrogens in cancers: Emerging methods and concepts. Int. J. Mol. Sci. 2021, 22, 8798. [Google Scholar] [CrossRef]
- Petrie, B.; Lopardo, L.; Proctor, K.; Youdan, J.; Barden, R.; Kasprzyk-Hordern, B. Assessment of bisphenol-A in the urban water cycle. Sci. Total Environ. 2019, 650, 900–907. [Google Scholar] [CrossRef]
- Samia, K.; Dhouha, A.; Anis, C.; Ammar, M.; Rim, A.; Abdelkrim, C. Assessment of organic pollutants (PAH and PCB) in surface water: Sediments and shallow groundwater of grombalia watershed in northeast of tunisia. Arab. J. Geosci. 2018, 11, 34. [Google Scholar] [CrossRef]
- Lu, S.; Lin, C.; Lei, K.; Xin, M.; Wang, B.; Ouyang, W.; He, M. Endocrine-disrupting chemicals in a typical urbanized bay of yellow sea, china: Distribution, risk assessment, and identification of priority pollutants. Environ. Pollut. 2021, 287, 117588. [Google Scholar] [CrossRef]
- Luo, Z.; Tu, Y.; Li, H.; Qiu, B.; Liu, Y.; Yang, Z. Endocrine-disrupting compounds in the Xiangjiang River of China: Spatio-temporal distribution, source apportionment, and risk assessment. Ecotoxicol. Environ. Saf. 2019, 167, 476–484. [Google Scholar] [CrossRef]
- Radwan, E.K.; Ibrahim, M.B.M.; Adel, A.; Farouk, M. The occurrence and risk assessment of phenolic endocrine-disrupting chemicals in egypt’s drinking and source water. Environ. Sci. Pollut. Res. 2020, 27, 1776–1788. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, Z.; Luo, Z.; Li, H.; Chen, G. Endocrine disrupting chemicals in wild freshwater fishes: Species, tissues, sizes and human health risks. Environ. Pollut. 2019, 244, 462–468. [Google Scholar] [CrossRef]
- Ribeiro, C.; Tiritan, M.E.; Rocha, E.; Rocha, M.J. Seasonal and spatial distribution of several endocrine-disrupting compounds in the Douro River Estuary, Portugal. Arch. Environ. Contam. Toxicol. 2009, 56, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yu, W.; Yang, L.; Du, B.; Chen, S.; Sun, W.; Tang, J. Occurrence and fate of steroid estrogens in a Chinese typical concentrated dairy farm and slurry irrigated soil. J. Agric. Food Chem. 2020, 69, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wojnarowski, K.; Podobiński, P.; Cholewińska, P.; Smoliński, J.; Dorobisz, K. Impact of estrogens present in environment on health and welfare of animals. Animals 2021, 11, 2152. [Google Scholar] [CrossRef] [PubMed]
- Renaud, L.; Agarwal, N.; Richards, D.J.; Falcinelli, S.; Hazard, E.S.; Carnevali, O.; Hardiman, G. Transcriptomic analysis of short-term 17α-ethynylestradiol exposure in two californian sentinel fish species sardine (Sardinops Sagax) and mackerel (Scomber Japonicus). Environ. Pollut. 2019, 244, 926–937. [Google Scholar] [CrossRef]
- Meng, Q.; Yeung, K.; Kwok, M.L.; Chung, C.T.; Hu, X.L.; Chan, K.M. Toxic effects and transcriptome analyses of zebrafish (Danio Rerio) larvae exposed to benzophenones. Environ. Pollut. 2020, 265, 114857. [Google Scholar] [CrossRef]
- Anderson, J.C.; Beyger, L.; Guchardi, J.; Holdway, D.A. The effects of 17α-Ethinylestradiol on the heart rate of embryonic Japanese Medaka (Oryzias Latipes). Toxicol. Chem. 2020, 39, 904–912. [Google Scholar] [CrossRef]
- Elvin, T.; Malini, N.; George, K. Deleterious effect of short term exposure to xenoestrogen-bisphenol a on certain haematological and physiological profile of Freshwater Murrel. Channa Striata. Bloch 1793, 39, 126–133. [Google Scholar]
- Kirk, C.J.; Bottomley, L.; Minican, N.; Carpenter, H.; Shaw, S.; Kohli, N.; Harris, R.M. Environmental endocrine disrupters dysregulate estrogen metabolism and Ca2+ homeostasis in fish and mammals via receptor-independent mechanisms. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 135, 1–8. [Google Scholar] [CrossRef]
- Nelson, E.R.; Habibi, H.R. Estrogen receptor function and regulation in fish and other vertebrates. Gen. Comp. Endocrinol. 2013, 192, 15–24. [Google Scholar] [CrossRef]
- Sun, S.-X.; Wu, J.-L.; Lv, H.-B.; Zhang, H.-Y.; Zhang, J.; Limbu, S.M.; Du, Z.-Y. Environmental estrogen exposure converts lipid metabolism in male fish to a female pattern mediated by AMPK And mtor signaling pathways. J. Hazard. Mater. 2020, 394, 122537. [Google Scholar] [CrossRef] [PubMed]
- Warner, K.E.; Jenkins, J.J. Effects of 17α-Ethinylestradiol and bisphenol A on vertebral development in the fathead Minnow (Pimephales Promelas). Toxicol. Chem. Int. J. 2007, 26, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhong, H.; Han, Z.; Tang, Z.; Xiao, J.; Guo, Z.; Zhou, Y. Effects of waterborne exposure to 17β-estradiol on hepatic lipid metabolism genes in Tilapia (Oreochromis Niloticus). Aquac. Rep. 2020, 17, 100382. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, Z.; Liu, Q.; Zhang, M.; Yang, H.; Wei, W. Bisphenol A disrupts Apolipoprotein e expression through estrogen-related receptor gamma and DNA methlylation in the liver of male rare Minnow (Gobiocypris Rarus). Ecotoxicol. Environ. Saf. 2021, 228, 113041. [Google Scholar] [CrossRef]
- Nilsson, B.O.; Olde, B.; Leeb-Lundberg, L.F. G protein-coupled oestrogen receptor 1 (GPER1)/GPR30: A new player in cardiovascular and metabolic oestrogenic signalling. Br. J. Pharmacol. 2011, 163, 1131–1139. [Google Scholar] [CrossRef]
- Thomas, P.; Alyea, R.; Pang, Y.; Peyton, C.; Dong, J.; Berg, A.H. Conserved estrogen binding and signaling functions of the G Protein-coupled estrogen receptor 1 (GPER) in mammals and fish. Steroids 2010, 75, 595–602. [Google Scholar] [CrossRef]
- Teng, J.; Zhao, Y.; Chen, H.J.; Xue, L.Y.; Ji, X.S. Global expression response of genes in sex-undifferentiated Nile tilapia gonads after exposure to trace letrozole. Ecotoxicol. Environ. Saf. 2021, 217, 112255. [Google Scholar] [CrossRef]
- Orozco-Hernández, L.; Gutiérrez-Gómez, A.A.; SanJuan-Reyes, N.; Islas-Flores, H.; García-Medina, S.; Galar-Martínez, M.; Gómez-Oliván, L.M. 17β-Estradiol induces cyto-genotoxicity on blood cells of common carp (Cyprinus carpio). Chemosphere 2018, 191, 118–127. [Google Scholar] [CrossRef]
- Sator, P.G.; Schmidt, J.B.; Rabe, T.; Zouboulis, C.C. Skin aging and sex hormones in women–clinical perspectives for intervention by hormone replacement therapy. Exp. Dermatol. 2004, 13, 36–40. [Google Scholar] [CrossRef]
- Gill, A.; Patranabis, S. Phytohormones as potential anticancer agents. Int. J. Res. Appl. Sci. Biotechnol. 2021, 8, 37–43. [Google Scholar] [CrossRef]
- Sehmisch, S.; Hammer, F.; Christoffel, J.; Seidlova-Wuttke, D.; Tezval, M.; Wuttke, W.; Stuermer, E.K. Comparison of the phytohormones genistein, resveratrol and 8-Prenylnaringenin as agents for preventing osteoporosis. Planta Med. 2008, 74, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Page, Y.L.; Percevault, F.; Ferriere, F.; Flouriot, G.; Pakdel, F. Apigenin, a partial antagonist of the estrogen receptor (ER), inhibits ER-Positive breast cancer cell proliferation through Akt/FOXM1 signaling. Int. J. Mol. Sci. 2021, 22, 470. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, T.F.R.; Colleta, S.J.; De Campos Zuccari, D.A.P.; Vilamaior, P.S.L.; Leonel, E.C.R.; Taboga, S.R. Hormone receptor expression in aging mammary tissue and carcinoma from a rodent model after xenoestrogen disruption. Life Sci. 2021, 285, 120010. [Google Scholar] [CrossRef] [PubMed]
- Calaf, G.M.; Ponce Cusi, R.; Aguayo, F.; Muñoz, J.P.; Bleak, T.C. Endocrine disruptors from the environment affecting breast cancer. Oncol. Lett. 2020, 20, 19–32. [Google Scholar] [CrossRef]
- Henderson, B.E.; Feigelson, H.S. Hormonal carcinogenesis. Carcinogenesis 2000, 21, 427–433. [Google Scholar] [CrossRef]
- Bohra, A.; Bhateja, S. Carcinogenesis and sex hormones: A review. Endocrinol. Metab. Syndrome. 2015, 4, 1–14. [Google Scholar]
- Bhardwaj, P.; Au, C.C.; Benito-Martin, A.; Ladumor, H.; Oshchepkova, S.; Moges, R.; Brown, K.A. Estrogens and breast cancer: Mechanisms involved in obesity-related development, growth and progression. J. Steroid Biochem. Mol. Biol. 2019, 189, 161–170. [Google Scholar] [CrossRef]
- Jackson, L.; Klerks, P. Effects of the synthetic estrogen 17α-Ethinylestradiol on heterandria formosa populations: Does matrotrophy circumvent population collapse? Aquat. Toxicol. 2020, 229, 105659. [Google Scholar] [CrossRef]
- Karki, N.P.; Colombo, R.E.; Gaines, K.F.; Maia, A. Exposure to 17β estradiol causes erosion of sexual dimorphism in bluegill (Lepomis Macrochirus). Environ. Sci. Pollut. Res. 2021, 28, 6450–6458. [Google Scholar] [CrossRef]
- Lam, S.H.; Lee, S.G.; Lin, C.Y.; Thomsen, J.S.; Fu, P.Y.; Murthy, K.R.; Mathavan, S. Molecular conservation of estrogen-response associated with cell cycle regulation, hormonal carcinogenesis and cancer in zebrafish and human. Cancer Cell Lines. BMC Med. Genom. 2011, 4, 41. [Google Scholar] [CrossRef]
- Chaturantabut, S.; Shwartz, A.; Evason, K.J.; Cox, A.G.; Labella, K.; Schepers, A.G.; Goessling, W. Estrogen Activation of G-Protein–coupled estrogen receptor 1 regulates phosphoinositide 3-Kinase and Mtor signaling to promote liver growth in zebrafish and proliferation of human hepatocytes. Gastroenterology 2019, 156, 1788–1804. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Mitra, A.; Homechaudhuri, S. Differential expression of genes responsible for reproduction of male and female Danio rerio following interaction between estrogen receptor and cadmium. Proc. Zool. Soc. Springer India 2021, 74, 456–465. [Google Scholar] [CrossRef]
- Brama, M.; Gnessi, L.; Basciani, S.; Cerulli, N.; Politi, L.; Spera, G.; Migliaccio, S. Cadmium induces mitogenic signaling in breast cancer cell by an Erα-dependent mechanism. Mol. Cell. Endocrinol. 2007, 264, 102–108. [Google Scholar] [CrossRef]
- Stoica, A.; Katzenellenbogen, B.S.; Martin, M.B. Activation of estrogen receptor—A by the heavy metal Cadmium. Mol. Endocrinol. 2000, 14, 545–553. [Google Scholar] [PubMed]
- Kim, Y.J.; Lee, N.; Woo, S.; Ryu, J.C.; Yum, S. Transcriptomic change as evidence for Cadmium-induced endocrine disruption in marine fish model of Medaka, Oryzias Javanicus. Mol. Cell. Toxicol. 2016, 12, 409–420. [Google Scholar] [CrossRef]
- Christiansen, T.; Korsgaard, B.; Jespersen, A. Effects of nonylphenol and 17 beta-oestradiol on vitellogenin synthesis, testicular structure and cytology in male eelpout Zoarces viviparus. J. Exp. Biol. 1998, 201, 179–192. [Google Scholar] [CrossRef]
- Azevedo, D.D.A.; Lacorte, S.; Viana, P.; Barceló, D. Occurrence of nonylphenol and bisphenol-A in surface waters from Portugal. J. Braz. Chem. Soc. 2001, 12, 532–537. [Google Scholar] [CrossRef]
- Ganesan, S.; Keating, A.F. Bisphenol A-induced ovotoxicity involves DNA damage induction to which the ovary mounts a protective response indicated by increased expression of proteins involved in DNA repair and xenobiotic biotransformation. Toxicol. Sci. 2016, 152, 169–180. [Google Scholar] [CrossRef][Green Version]
- Khan, N.G.; Correia, J.; Adiga, D.; Rai, P.S.; Dsouza, H.S.; Chakrabarty, S.; Kabekkodu, S.P. A comprehensive review on the carcinogenic potential of bisphenol A: Clues and evidence. Environ. Scicience Pollut. Res. 2021, 28, 19643–19663. [Google Scholar] [CrossRef]
- Dumitrascu, M.C.; Mares, C.; Petca, R.C.; Sandru, F.; Popescu, R.I.; Mehedintu, C.; Petca, A. Carcinogenic effects of bisphenol a in breast and ovarian cancers. Oncol. Lett. 2020, 20, 282. [Google Scholar] [CrossRef]
- Cervantes-Camacho, I.; Guerrero-Estévez, S.M.; López, M.F.; Alarcón-Hernández, E.; López-López, E. Effects of Bisphenol a on Foxl2 gene expression and DNA damage in adult viviparous fish Goodea Atripinnis. J. Toxicol. Environ. Health Part A 2020, 83, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Major, K.M.; Decourten, B.M.; Li, J.; Britton, M.; Settles, M.L.; Mehinto, A.C.; Brander, S.M. Early life exposure to environmentally relevant levels of endocrine disruptors drive multigenerational and transgenerational epigenetic changes in a fish model. Front. Mar. Sci. 2020, 7, 471. [Google Scholar] [CrossRef]
- Callard, G.V.; Tchoudakova, A.V.; Kishida, M.; Wood, E. Differential tissue distribution, developmental programming, estrogen regulation and promoter characteristics of Cyp19 genes in teleost fish. J. Steroid Biochem. Mol. Biol. 2001, 79, 305–314. [Google Scholar] [CrossRef]
- Filice, M.; Leo, S.; Mazza, R.; Amelio, D.; Garofalo, F.; Imbrogno, S.; Gattuso, A. The heart of the adult goldfish (Carassius auratus) as a target of bisphenol A: A multifaceted analysis. Environ. Pollut. 2021, 269, 116177. [Google Scholar] [CrossRef] [PubMed]
- Moreman, J.; Takesono, A.; Trznadel, M.; Winter, M.J.; Perry, A.; Wood, M.E.; Tyler, C.R. Estrogenic mechanisms and cardiac responses following early life exposure to bisphenol A (BPA) and its metabolite 4-Methyl-2, 4-Bis (P-Hydroxyphenyl) Pent-1-Ene (MBP) in zebrafish. Environ. Sci. Technol. 2018, 52, 6656–6665. [Google Scholar] [CrossRef]
- Bakos, K.; Kovács, R.; Staszny, Á.; Sipos, D.K.; Urbányi, B.; Müller, F.; Kovács, B. Developmental toxicity and estrogenic potency of zearalenone in zebrafish (Danio Rerio). Aquat. Toxicol. 2013, 136, 13–21. [Google Scholar] [CrossRef]
- Amenyogbe, E.; Chen, G.; Wang, Z.; Lu, X.; Lin, M.; Lin, A.Y. A review on sex steroid hormone estrogen receptors in mammals and fish. Int. J. Endocrinol. 2020, 2020, 5386193. [Google Scholar] [CrossRef]
- Budczies, J.; Brockmöller, S.F.; Müller, B.M.; Barupal, D.K.; Richter-Ehrenstein, C.; Kleine-Tebbe, A.; Fiehn, O. Comparative metabolomics of estrogen receptor positive and estrogen receptor negative breast cancer: Alterations in glutamine and beta-alanine metabolism. J. Proteom. 2013, 94, 279–288. [Google Scholar] [CrossRef]
- Loomis, A.K.; Thomas, P. Effects of estrogens and xenoestrogens on androgen production by Atlantic croaker testes in vitro: Evidence for a nongenomic action mediated by an estrogen membrane receptor. Biol. Reprod. 2000, 62, 995–1004. [Google Scholar] [CrossRef]
- Mohapatra, S.; Chakraborty, T.; Shimizu, S.; Ohta, K.; Nagahama, Y.; Ohta, K. Estrogen and estrogen receptors chauffeur the sex-biased autophagic action in liver. Cell Death Differ. 2020, 27, 3117–3130. [Google Scholar] [CrossRef]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and autophagy. Methods Molecural Biol. 2008, 445, 77–88. [Google Scholar] [CrossRef]
- Feswick, A.; Munkittrick, K.R.; Martyniuk, C.J. Estrogen-responsive gene networks in the teleost liver: What are the key molecular indicators? Environ. Toxicol. Pharmacol. 2017, 56, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Knopp, R.H.; Paramsothy, P.; Retzlaff, B.M.; Fish, B.; Walden, C.; Dowdy, A.; Cheung, M.C. Sex differences in lipoprotein metabolism and dietary response: Basis in hormonal differences and implications for cardiovascular disease. Curr. Cardiol. Rep. 2006, 8, 452–459. [Google Scholar] [CrossRef]
- Cleveland, B.M.; Weber, G.M. Effects of steroid treatment on growth, nutrient partitioning, and expression of genes related to growth and nutrient metabolism in adult triploid rainbow trout (Oncorhynchus Mykiss). Domest. Anim. Endocrinol. 2016, 56, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sovadinová, I.; Liedtke, A.; Schirmer, K. Effects of clofibric acid alone and in combination with 17β-Estradiol on mRNA abundance in primary hepatocytes isolated from rainbow trout. Toxicol. Vitr. 2014, 28, 1106–1116. [Google Scholar] [CrossRef]
- Celino-Brady, F.T.; Petro-Sakuma, C.K.; Breves, J.P.; Lerner, D.T.; Seale, A.P. Early-Life exposure to 17β-Estradiol and 4-Nonylphenol impacts the growth hormone/insulin-like growth-factor system and estrogen receptors in Mozambique Tilapia, Oreochromis Mossambicus. Aquat. Toxicol. 2019, 217, 105336. [Google Scholar] [CrossRef]
- Müller, A.K.; Markert, N.; Leser, K.; Kämpfer, D.; Schiwy, S.; Riegraf, C.; Hollert, H. Bioavailability and impacts of estrogenic compounds from suspended sediment on rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2021, 231, 105719. [Google Scholar] [CrossRef]
- Breves, J.P.; Duffy, T.A.; Einarsdottir, I.E.; Björnsson, B.T.; McCormick, S.D. In vivo effects of 17α-ethinylestradiol, 17β-estradiol and 4-nonylphenol on insulin-like growth-factor binding proteins (igfbps) in Atlantic salmon. Aquat. Toxicol. 2018, 203, 28–39. [Google Scholar] [CrossRef]
- Lazaro-Velasco, A.; Isidro-Cristobal, H.M.; Alcántar-Vázquez, J.P.; Antonio-Estrada, C.; Calzada-Ruiz, D.; Torre, R.M.D.L. Effect of the combination of a cold-water temperature and exogenous estrogens on feminization, growth, gonadosomatic index and fat muscle content of Nile tilapia Oreochromis niloticus (Linnaeus, 1758). Lat. Am. J. Aquat. Res. 2019, 47, 52–64. [Google Scholar] [CrossRef]
- Kovats, S. Estrogen receptors regulate innate immune cells and signaling pathways. Cell. Immunol. 2015, 294, 63–69. [Google Scholar] [CrossRef]
- Cunningham, M.; Gilkeson, G. Estrogen receptors in immunity and autoimmunity. Clin. Rev. Allergy Immunol. 2011, 40, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Iwanowicz, L.R.; Ottinger, C.A. Estrogens, estrogen receptors and their role as immunoregulators in fish. Fish Def. 2009, 1, 277–322. [Google Scholar]
- Chaves-Pozo, E.; García-Ayala, A.; Cabas, I. Effects of sex steroids on fish leukocytes. Biology 2018, 7, 9. [Google Scholar] [CrossRef]
- Seemann, F.; Knigge, T.; Duflot, A.; Marie, S.; Olivier, S.; Minier, C.; Monsinjon, T. Sensitive periods for 17β-estradiol exposure during immune system development in Sea Bass Head kidney. J. Appl. Toxicol. 2016, 36, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Moreira, C.; Paiola, M.; Duflot, A.; Varó, I.; Sitjà-Bobadilla, A.; Knigge, T.; Monsinjon, T. the influence of 17β-oestradiol on lymphopoiesis and immune system ontogenesis in juvenile Sea Bass, Dicentrarchus Labrax. Dev. Comp. Immunol. 2021, 118, 104011. [Google Scholar] [CrossRef] [PubMed]
- Woźny, M.; Obremski, K.; Hliwa, P.; Gomułka, P.; Różyński, R.; Wojtacha, P.; Brzuzan, P. Feed contamination with zearalenone promotes growth but affects the immune system of rainbow trout. Fish Shellfish Immunol. 2019, 84, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Tawwab, M.; Khalil, R.H.; Diab, A.M.; Khallaf, M.A.; Abdel-Razek, N.; Abdel-Latif, H.M.; Khalifa, E. Dietary garlic and chitosan enhanced the antioxidant capacity, immunity, and modulated the transcription of HSP70 and cytokine genes in zearalenone-intoxicated european Seabass. Fish Shellfish Immunol. 2021, 113, 35–41. [Google Scholar] [CrossRef]
- Park, K.; Kwak, I.S. Expression of Stress Response HSP70 Gene in Asian Paddle Crabs, Charybdis Japonica, Exposure to endocrine disrupting chemicals, bisphenol A (BPA) and 4-Nonylphenol (NP). Ocean Sci. J. 2013, 48, 207–214. [Google Scholar] [CrossRef]
- Li, X.; Wang, S.; Qi, J.; Echtenkamp, S.F.; Chatterjee, R.; Wang, M.; Gupta, D. zebrafish peptidoglycan recognition proteins are bactericidal amidases essential for defense against bacterial infections. Immunity 2007, 27, 518–529. [Google Scholar] [CrossRef]
- Stamenkovic, I.; Yu, Q.; Merlin, A. “Magic” linker between the extracellular cues and intracellular signaling pathways that regulate cell motility, proliferation, and survival. Curr. Protein Pept. Sci. 2010, 11, 471–484. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Khalifa, E.; Diab, A.M.; Khallaf, M.A.; Abdel-Razek, N.; Khalil, R.H. Dietary garlic and chitosan alleviated zearalenone toxic effects on performance, immunity, and challenge of european sea bass, Dicentrarchus Labrax, to Vibrio Alginolyticus infection. Aquac. Int. 2020, 28, 493–510. [Google Scholar] [CrossRef]
- Rehberger, K.; Von Siebenthal, E.W.; Bailey, C.; Bregy, P.; Fasel, M.; Herzog, E.L.; Segner, H. Long-Term exposure to low 17α-ethinylestradiol (EE2) concentrations disrupts both the reproductive and the immune system of juvenile rainbow trout, Oncorhynchus Mykiss. Environ. Int. 2020, 142, 105836. [Google Scholar] [CrossRef] [PubMed]
- Kernen, L.; Phan, A.; Bo, J.; Herzog, E.L.; Huynh, J.; Segner, H.; Baumann, L. Estrogens as immunotoxicants: 17α-ethinylestradiol exposure retards thymus development in zebrafish (Danio Rerio). Aquat. Toxicol. 2022, 242, 106025. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, F.; Zhang, Y.; Cai, L.; Qiu, W.; Yang, M.G. protein-coupled estrogen receptor 1 mediates estrogen effect in red common carp (Cyprinus carpio). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 239, 108868. [Google Scholar] [CrossRef] [PubMed]
- Maciuszek, M.; Pijanowski, L.; Pekala-Safinska, A.; Kemenade, B.M.; Chadzinska, M. 17β-Estradiol affects the innate immune response in common carp. Fish Physiol. Biochem. 2020, 46, 1775–1794. [Google Scholar] [CrossRef]
- Maciuszek, M.; Pijanowski, L.; Pekala-Safinska, A.; Palichleb, P.; Błachut, M.; Verburg-van Kemenade, B.L.; Chadzińska, M. 17α-ethinylestradiol and 4-tert-octylphenol concurrently disrupt the immune response of common carp. Fish Shellfish Immunol. 2020, 107, 238–250. [Google Scholar] [CrossRef]
- Wang, Y.L.; Zhou, X.Q.; Jiang, W.D.; Wu, P.; Liu, Y.; Jiang, J.; Feng, L. Effects of dietary zearalenone on oxidative stress, cell apoptosis, and tight junction in the intestine of juvenile grass carp (Ctenopharyngodon idella). Toxins 2019, 11, 333. [Google Scholar] [CrossRef]
- Pacheco Passos Neto, O.; Bezerra dos Santos, A.; Feitosa Silva, J.R.; Mota, S. Alterations in the development and gonadal structure of Nile Tilapia (Oreochromis niloticus) exposed to natural and synthetic estrogens. Water Air Soil Pollut. 2021, 232, 1–13. [Google Scholar] [CrossRef]
- Fan, X.; Hou, T.; Jia, J.; Tang, K.; Wei, X.; Wang, Z. Discrepant dose responses of bisphenol A on oxidative stress and DNA methylation in grass carp ovary cells. Chemosphere 2020, 248, 126110. [Google Scholar] [CrossRef]
- Forner-Piquer, I.; Fakriadis, I.; Mylonas, C.C.; Piscitelli, F.; Di Marzo, V.; Maradonna, F.; Carnevali, O. Effects of dietary bisphenol A on the reproductive function of gilthead sea bream (Sparus aurata) testes. Int. J. Mol. Sci. 2019, 20, 5003. [Google Scholar] [CrossRef]
- Xie, Q.P.; Li, B.B.; Wei, F.L.; Yu, M.; Zhan, W.; Liu, F.; Lou, B. Growth and gonadal development retardations after long-term exposure to estradiol in little yellow croaker, Larimichthys polyactis. Ecotoxicol. Environ. Saf. 2021, 222, 112462. [Google Scholar] [CrossRef] [PubMed]
- Voisin, A.S.; Kültz, D.; Silvestre, F. Early-life exposure to the endocrine disruptor 17-α-ethinylestradiol induces delayed effects in adult brain, liver and ovotestis proteomes of a self-fertilizing fish. J. Proteom. 2019, 194, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Łuszczek-Trojnar, E.; Drąg-Kozak, E.; Szczerbik, P.; Socha, M.; Popek, W. Effect of long-term dietary lead exposure on some maturation and reproductive parameters of a female Prussian carp (Carassius gibelio B.). Environ. Sci. Pollut. Res. 2014, 21, 2465–2478. [Google Scholar] [CrossRef] [PubMed]
- Paschoalini, A.L.; Savassi, L.A.; Arantes, F.P.; Rizzo, E.; Bazzoli, N. Heavy metals accumulation and endocrine disruption in Prochilodus argenteus from a polluted neotropical river. Ecotoxicol. Environ. Saf. 2019, 169, 539–550. [Google Scholar] [CrossRef]
- Pojana, G.; Gomiero, A.; Jonkers, N.; Marcomini, A. Natural and synthetic endocrine disrupting compounds (EDCs) in water, sediment and biota of a coastal lagoon. Environ. Int. 2007, 33, 929–936. [Google Scholar] [CrossRef]
- Tang, Z.; Liu, Z.H.; Wang, H.; Dang, Z.; Liu, Y. A review of 17α-ethynylestradiol (EE2) in surface water across 32 countries: Sources, concentrations, and potential estrogenic effects. J. Environ. Manag. 2021, 292, 112804. [Google Scholar] [CrossRef]
- Rotimi, O.A.; Olawole, T.D.; De Campos, O.C.; Adelani, I.B.; Rotimi, S.O. Bisphenol A in Africa: A review of environmental and biological levels. Sci. Total Environ. 2021, 764, 142854. [Google Scholar] [CrossRef]
- Gromadzka, K.; Waśkiewicz, A.; Goliński, P.; Świetlik, J. Occurrence of estrogenic mycotoxin–zearalenone in aqueous environmental samples with various NOM content. Water Res. 2009, 43, 1051–1059. [Google Scholar] [CrossRef]
- Jin, X.; Jiang, G.; Huang, G.; Liu, J.; Zhou, Q. Determination of 4-tert-octylphenol, 4-nonylphenol and bisphenol A in surface waters from the Haihe River in Tianjin by gas chromatography–mass spectrometry with selected ion monitoring. Chemosphere 2004, 56, 1113–1119. [Google Scholar] [CrossRef]
- Domagalski, J.L.; Weston, D.P.; Zhang, M.; Hladik, M. Pyrethroid insecticide concentrations and toxicity in streambed sediments and loads in surface waters of the San Joaquin Valley, California, USA. Environ. Toxicol. Chem. Int. J. 2010, 29, 813–823. [Google Scholar] [CrossRef]
- King, O.C.; van de Merwe, J.P.; McDonald, J.A.; Leusch, F.D. Concentrations of levonorgestrel and ethinylestradiol in wastewater effluents: Is the progestin also cause for concern? Environ. Toxicol. Chem. 2016, 35, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.M.; Yao, L.A.; Ma, Q.L.; Zhou, G.J.; Wang, L.; Fang, Q.L.; Xu, Z.C. Distribution and ecological risk assessment of cadmium in water and sediment in Longjiang River, China: Implication on water quality management after pollution accident. Chemosphere 2018, 194, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.J.; Poulakos, A.G.; Tornero-Velez, R.; Xue, J. Systematic review and meta-analyses of lead (Pb) concentrations in environmental media (soil, dust, water, food, and air) reported in the United States from 1996 to 2016. Sci. Total Environ. 2019, 694, 133489. [Google Scholar] [CrossRef] [PubMed]

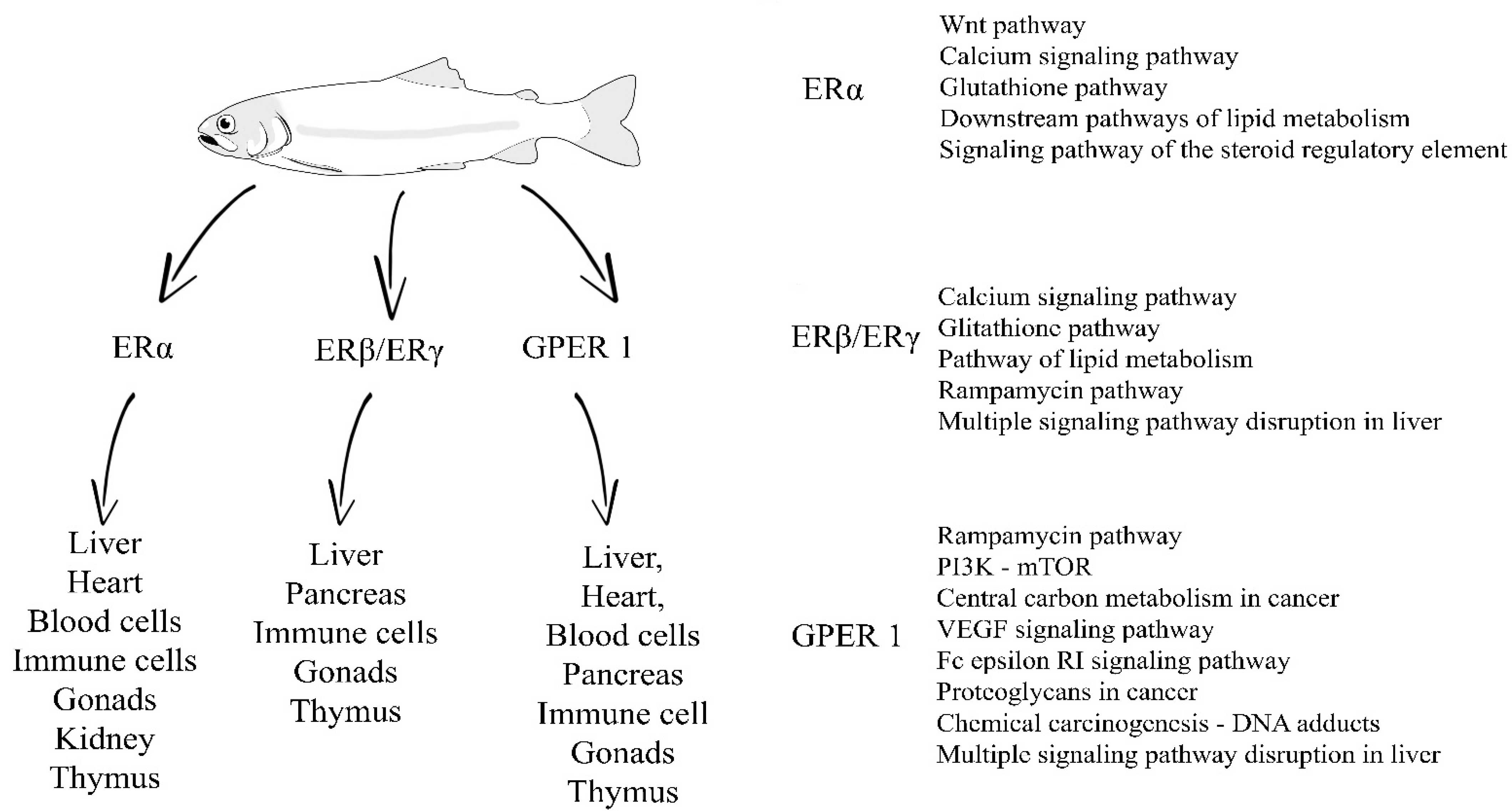{kind=link}
| Compound | Selected Negative Effects | Environmentally Detected Concentrations | References |
|---|---|---|---|
| 17-β-estradiol E2 | DNA damage Carcinogenesis Cardioviscular diseases Brachycardic effect Cyto—genetoxic effect on fish blood Lipid transformation Modification on interleukin 1β Ovotestis and gonadal development retardations; | 175 ng/L—Venice lagoon/February 2002 | [48,59,72,84,85,122,126] |
| 17-α-Ethinylestradiol EE2 | Carcinogenesis Neoplastic processes Hepatocarcinogenetic processes Vtg mRNA level increased Reduced Thymus growth; | 21.5 ng/L—mean concentration, China, 2021 | [46,72,113,127] |
| Bisphenols BPs | Disrupted ER expression Disrupted gene expression DNA damage Impaired myocardial hemodynamics Negative impact on blood cells (RBC, WBC, Hb%) Calcium signaling pathway Glutathione metabolism; Disrupted lipid metabolism Reduced sperm motility Morphopatological alteration in gonads Up regulated: leptin receptors, ERα and ERβ, hepatic genes and oogenesis. | 63.64 ng/L New Calabar River, Nigeria, 2019 | [47,49,54,55,79,80,81,82,85,121,128] |
| Zearaleone ZAE | Anatomical changes of heart muscle Disturbed gene expression Suppresed fish immune answer Negative impact on blood cells Morphological anomalies Intersex specimens; | 43.7 ng/L—Poland, 2009 | [87,107,112,119,129] |
| Nonylophenol NP | DNA damage Carcinogenesis Increased expression of insulin like growth factor Vtg production | 106–296 ng/L—Haihe river, China, 2004 | [97,98,130] |
| Bifenthrin | Disturbed gene expression Deregulation of glutathione conjugtion | 51.0 mg/L—Stanislaus River, USA, 2009 | [47,83,131] |
| Levonorgestrel | Disturbed gene expression Deregulation of glutathione conjugtion | 0.1 ng/L—Australia, 2015 | [47,83,132] |
| Cadmium Cd | Activation of the ERα gene Disruption of expresion of genes responsible for ribosomal structure, biogenesis, RNA processing and modification, cell cycle control, cell division and chromosome division | 45.01 mg/L—Longjiang River, 2012 | [76,125,133] |
| Lead Pb | Increased incidence of histopathologies, Changes in number and morphology of germline cells, Up-regulation of expression of vitellogenin | 4.6 ppb—mean concentration in surface waters, USA, 1996–2016 | [124,134] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojnarowski, K.; Cholewińska, P.; Palić, D.; Bednarska, M.; Jarosz, M.; Wiśniewska, I. Estrogen Receptors Mediated Negative Effects of Estrogens and Xenoestrogens in Teleost Fishes—Review. Int. J. Mol. Sci. 2022, 23, 2605. https://doi.org/10.3390/ijms23052605
Wojnarowski K, Cholewińska P, Palić D, Bednarska M, Jarosz M, Wiśniewska I. Estrogen Receptors Mediated Negative Effects of Estrogens and Xenoestrogens in Teleost Fishes—Review. International Journal of Molecular Sciences. 2022; 23(5):2605. https://doi.org/10.3390/ijms23052605
Chicago/Turabian StyleWojnarowski, Konrad, Paulina Cholewińska, Dušan Palić, Małgorzata Bednarska, Magdalena Jarosz, and Iga Wiśniewska. 2022. "Estrogen Receptors Mediated Negative Effects of Estrogens and Xenoestrogens in Teleost Fishes—Review" International Journal of Molecular Sciences 23, no. 5: 2605. https://doi.org/10.3390/ijms23052605
APA StyleWojnarowski, K., Cholewińska, P., Palić, D., Bednarska, M., Jarosz, M., & Wiśniewska, I. (2022). Estrogen Receptors Mediated Negative Effects of Estrogens and Xenoestrogens in Teleost Fishes—Review. International Journal of Molecular Sciences, 23(5), 2605. https://doi.org/10.3390/ijms23052605