
Gremlin-1 and BMP-4 Overexpressed in Osteoarthritis Drive an Osteochondral-Remodeling Program in Osteoblasts and Hypertrophic Chondrocytes

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
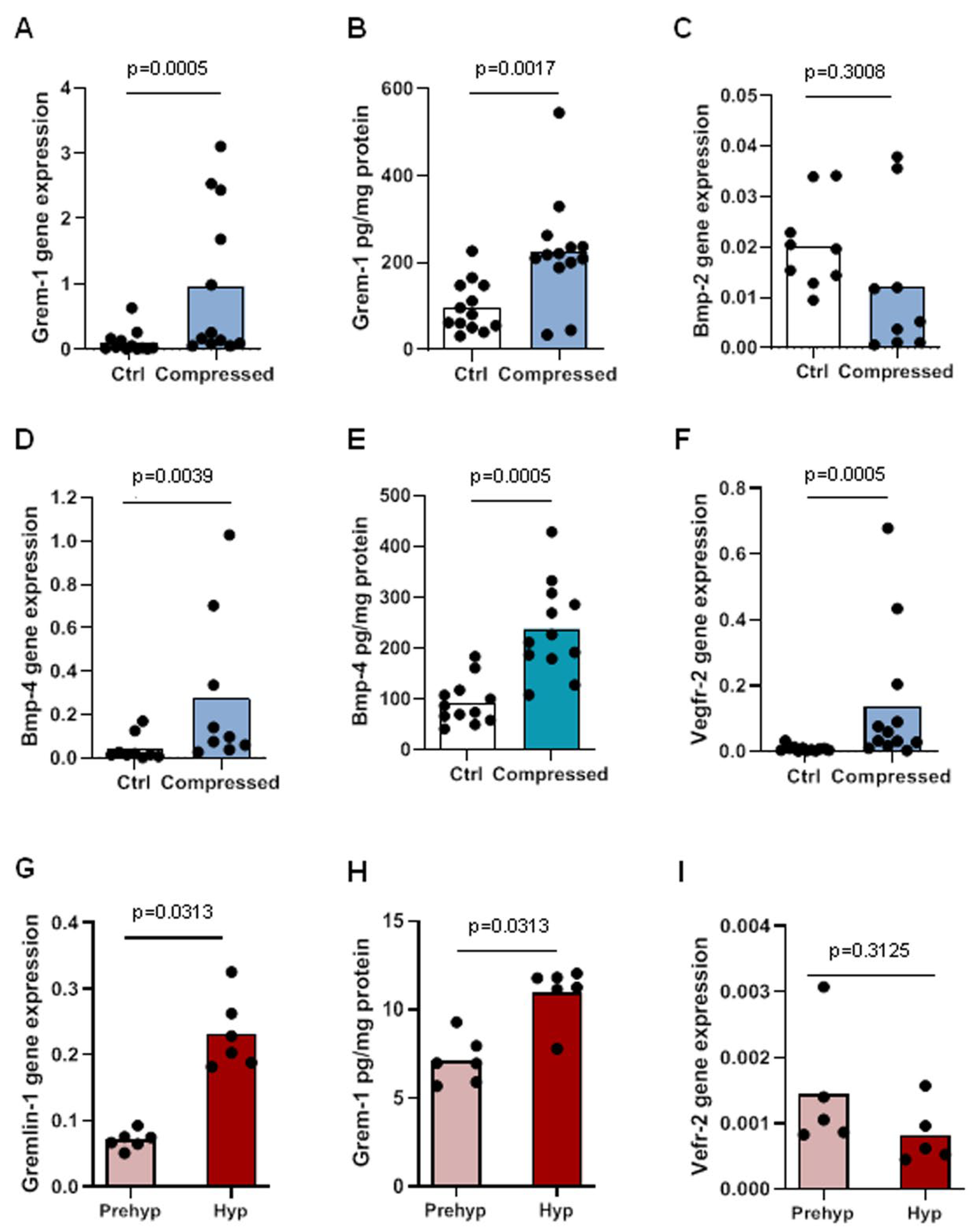
2.1. Overexpression of Gremlin-1 and BMPs at the Osteochondral Junction with OA Progression
2.2. Gremlin-1 Expression and Release Increased under Mechanical Compressive Loading on Osteoblasts
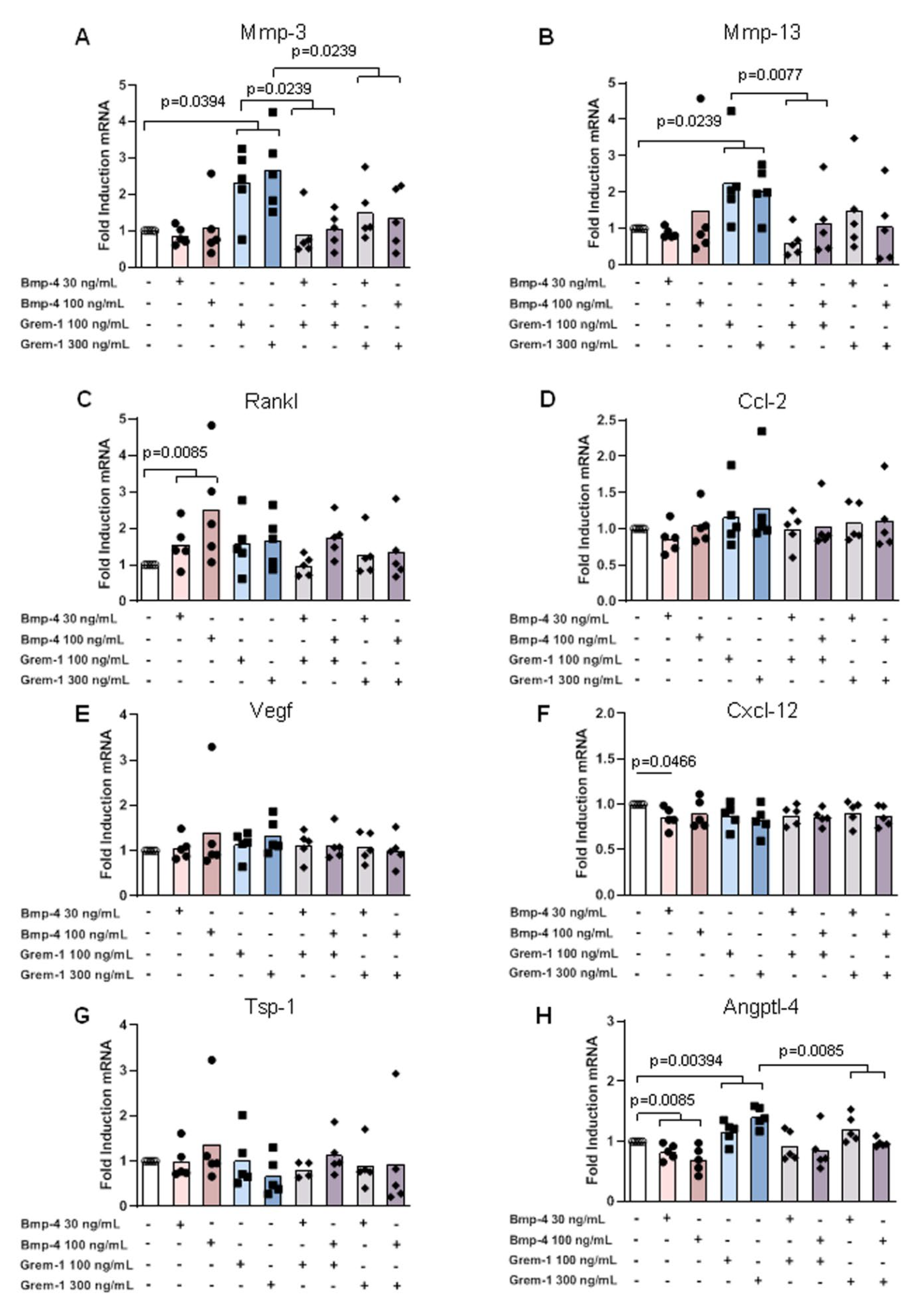
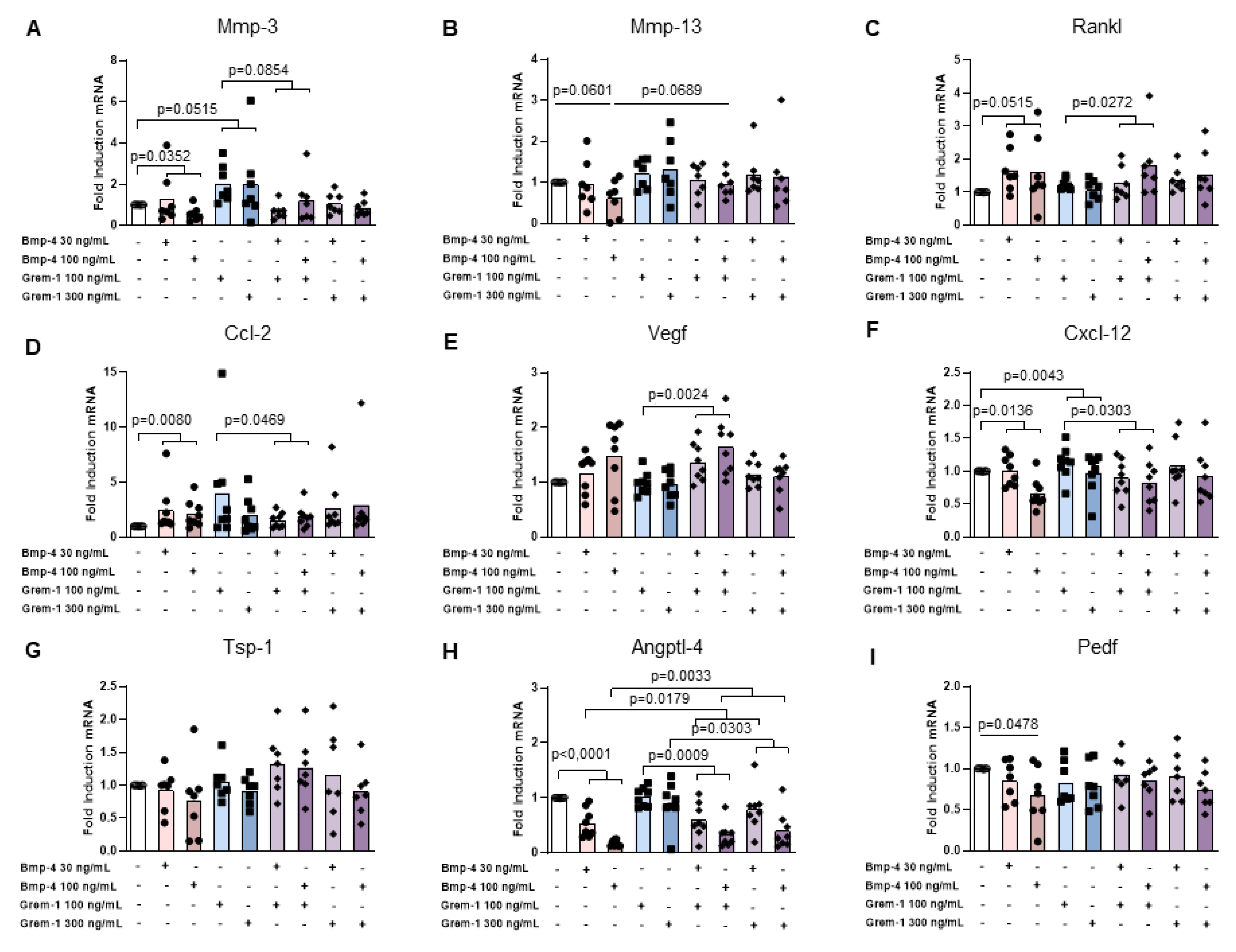
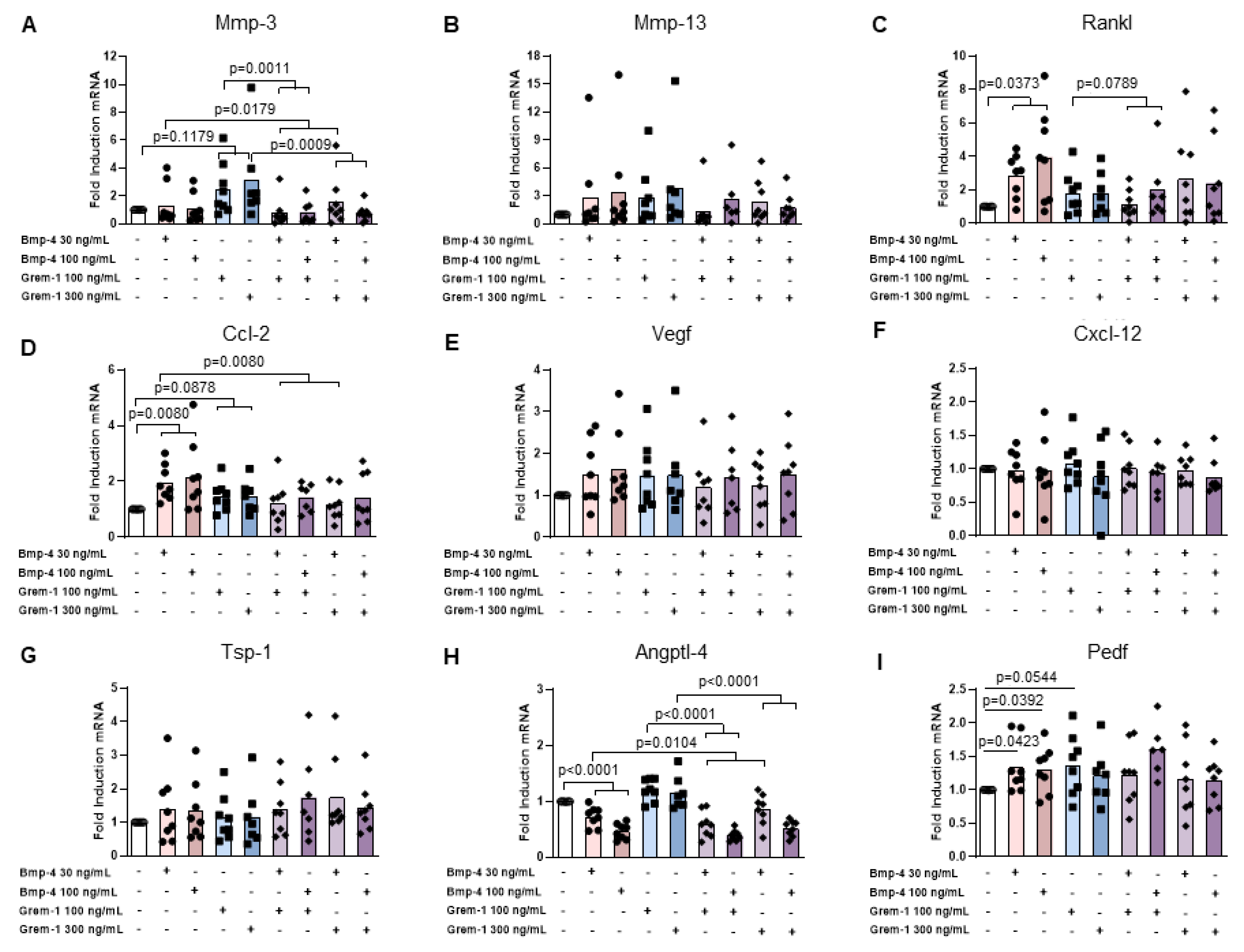
2.3. Grem-1 and BMP-4 Increase the Remodeling Potential of Osteoblasts and Chondrocytes
3. Discussion
4. Materials and Methods
4.1. Collection of Osteoarthritis Human Cartilage and Subchondral Bone
4.2. Isolation, Culture and Differentiation of Primary Calvaria Mouse Osteoblasts
4.3. Isolation, Culture, and Differentiation of Primary Articular Mouse Chondrocytes
4.4. Compression Experiments
4.5. Stimulation of Primary Cultures of Murine Chondrocytes and Osteoblasts
4.6. Real-Time Quantitative PCR Analysis
4.7. Protein Quantification and ELISA Assessment of Grem-1, BMP-2, BMP-4
4.8. Histology and Immunohistochemistry
4.9. Image Analysis and Morphometric Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, D.J.; Schofield, D.; Callander, E. The individual and socioeconomic impact of osteoarthritis. Nat. Rev. Rheumatol. 2014, 10, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Nüesch, E.; Dieppe, P.; Reichenbach, S.; Williams, S.; Iff, S.; Jüni, P. All cause and disease specific mortality in patients with knee or hip osteoarthritis: Population based cohort study. BMJ 2011, 342, d1165. [Google Scholar] [CrossRef] [Green Version]
- Kassebaum, N.J.; Arora, M.; Barber, R.M.; Bhutta, Z.A.; Brown, J.; Carter, A.; Casey, D.C.; Charlson, F.J.; Coates, M.M.; Coggeshall, M.; et al. Global, regional, and national disability-adjusted life-years (DALYs) for 315 diseases and injuries and healthy life expectancy (HALE), 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1603–1658. [Google Scholar] [CrossRef] [Green Version]
- Blagojevic, M.; Jinks, C.; Jeffery, A.; Jordan, K.P. Risk factors for onset of osteoarthritis of the knee in older adults: A systematic review and meta-analysis. Osteoarthr. Cartil. 2010, 18, 24–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldring, S.R.; Goldring, M.B. Changes in the osteochondral unit during osteoarthritis: Structure, function and cartilage-bone crosstalk. Nat. Rev. Rheumatol. 2016, 12, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Burr, D.B.; Gallant, M.A. Bone remodelling in osteoarthritis. Nat. Rev. Rheumatol. 2012, 8, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Lane, L.B.; Villacin, A.; Bullough, P.G. The vascularity and remodelling of subchondrial bone and calcified cartilage in adult human femoral and humeral heads. An age- and stress-related phenomenon. J. Bone Jt. Surg. Br. 1977, 59, 272–278. [Google Scholar] [CrossRef]
- Lories, R.J.; Luyten, F.P. The bone-cartilage unit in osteoarthritis. Nat. Rev. Rheumatol 2011, 7, 43–49. [Google Scholar] [CrossRef]
- Singh, P.; Marcu, K.B.; Goldring, M.B.; Otero, M. Phenotypic instability of chondrocytes in osteoarthritis: On a path to hypertrophy. Ann. N. Y. Acad. Sci. 2019, 1442, 17–34. [Google Scholar] [CrossRef]
- Rubin, C.T.; Lanyon, L.E. Regulation of bone mass by mechanical strain magnitude. Calcif. Tissue Int. 1985, 37, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Biewener, A.A.; Fazzalari, N.L.; Konieczynski, D.D.; Baudinette, R.V. Adaptive changes in trabecular architecture in relation to functional strain patterns and disuse. Bone 1996, 19, 1–8. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Aoyama, T.; Ito, A.; Nagai, M.; Iijima, H.; Zhang, X.; Tajino, J.; Kuroki, H. Effects of exercise level on biomarkers in a rat knee model of osteoarthritis: Effects of exercise on osteoarthritis. J. Orthop. Res. 2013, 31, 1026–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iijima, H.; Aoyama, T.; Ito, A.; Tajino, J.; Yamaguchi, S.; Nagai, M.; Kiyan, W.; Zhang, X.; Kuroki, H. Exercise intervention increases expression of bone morphogenetic proteins and prevents the progression of cartilage-subchondral bone lesions in a post-traumatic rat knee model. Osteoarthr. Cartil. 2016, 24, 1092–1102. [Google Scholar] [CrossRef] [Green Version]
- Iijima, H.; Ito, A.; Nagai, M.; Tajino, J.; Yamaguchi, S.; Kiyan, W.; Nakahata, A.; Zhang, J.; Wang, T.; Aoyama, T.; et al. Physiological exercise loading suppresses post-traumatic osteoarthritis progression via an increase in bone morphogenetic proteins expression in an experimental rat knee model. Osteoarthr. Cartil. 2017, 25, 964–975. [Google Scholar] [CrossRef] [Green Version]
- Leong, D.J.; Gu, X.I.; Li, Y.; Lee, J.Y.; Laudier, D.M.; Majeska, R.J.; Schaffler, M.B.; Cardoso, L.; Sun, H.B. Matrix metalloproteinase-3 in articular cartilage is upregulated by joint immobilization and suppressed by passive joint motion. Matrix Biol. 2010, 29, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Evans, W.J. Exercise and nutritional needs of elderly people: Effects on muscle and bone. Gerodontology 1998, 15, 15–24. [Google Scholar] [CrossRef]
- Cox, L.G.E.; van Rietbergen, B.; van Donkelaar, C.C.; Ito, K. Bone structural changes in osteoarthritis as a result of mechanoregulated bone adaptation: A modeling approach. Osteoarthr. Cartil. 2011, 19, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Cox, L.G.E.; van Donkelaar, C.C.; van Rietbergen, B.; Emans, P.J.; Ito, K. Alterations to the subchondral bone architecture during osteoarthritis: Bone adaptation vs. endochondral bone formation. Osteoarthr. Cartil. 2013, 21, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Gosset, M.; Berenbaum, F.; Levy, A.; Pigenet, A.; Thirion, S.; Saffar, J.-L.; Jacques, C. Prostaglandin E2 synthesis in cartilage explants under compression: mPGES-1 is a mechanosensitive gene. Arthritis Res. Ther. 2006, 8, R135. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, C.; Gabay, O.; Salvat, C.; Henrotin, Y.E.; Berenbaum, F. Mechanical loading highly increases IL-6 production and decreases OPG expression by osteoblasts. Osteoarthr. Cartil. 2009, 17, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, C.; Pesesse, L.; Gabay, O.; Delcour, J.-P.; Msika, P.; Baudouin, C.; Henrotin, Y.E. Regulation of subchondral bone osteoblast metabolism by cyclic compression. Arthritis Rheum. 2012, 64, 1193–1203. [Google Scholar] [CrossRef]
- Priam, S.; Bougault, C.; Houard, X.; Gosset, M.; Salvat, C.; Berenbaum, F.; Jacques, C. Identification of Soluble 14-3-3∊ as a Novel Subchondral Bone Mediator Involved in Cartilage Degradation in Osteoarthritis: Soluble 14-3-3∊ in Bone-Cartilage Communication. Arthritis Rheum. 2013, 65, 1831–1842. [Google Scholar] [CrossRef] [PubMed]
- Laiguillon, M.-C.; Courties, A.; Houard, X.; Auclair, M.; Sautet, A.; Capeau, J.; Fève, B.; Berenbaum, F.; Sellam, J. Characterization of diabetic osteoarthritic cartilage and role of high glucose environment on chondrocyte activation: Toward pathophysiological delineation of diabetes mellitus-related osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1513–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, L.; Huang, X.; Karperien, M.; Post, J. Correlation between Gene Expression and Osteoarthritis Progression in Human. Int. J. Mol. Sci. 2016, 17, 1126. [Google Scholar] [CrossRef] [Green Version]
- Leijten, J.C.; Bos, S.D.; Landman, E.; Georgi, N.; Jahr, H.; Meulenbelt, I.; Post, J.N.; van Blitterswijk, C.A.; Karperien, M. GREM1, FRZB and DKK1 mRNA levels correlate with osteoarthritis and are regulated by osteoarthritis-associated factors. Arthritis Res. Ther. 2013, 15, R126. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.; Jin, Q.; Zhang, B.; Wu, X.; Ge, D. Gremlin-1 Concentrations Are Correlated with the Severity of Knee Osteoarthritis. Med. Sci. Monit. 2016, 22, 4062–4065. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.H.; Mori, D.; Kobayashi, H.; Mori, Y.; Nakamoto, H.; Okada, K.; Taniguchi, Y.; Sugita, S.; Yano, F.; Chung, U.-I.; et al. Excessive mechanical loading promotes osteoarthritis through the gremlin-1–NF-κB pathway. Nat. Commun. 2019, 10, 1442. [Google Scholar] [CrossRef] [Green Version]
- Tardif, G.; Hum, D.; Pelletier, J.-P.; Boileau, C.; Ranger, P.; Martel-Pelletier, J. Differential gene expression and regulation of the bone morphogenetic protein antagonists follistatin and gremlin in normal and osteoarthritic human chondrocytes and synovial fibroblasts. Arthritis Rheum. 2004, 50, 2521–2530. [Google Scholar] [CrossRef]
- Tardif, G.; Pelletier, J.-P.; Boileau, C.; Martel-Pelletier, J. The BMP antagonists follistatin and gremlin in normal and early osteoarthritic cartilage: An immunohistochemical study. Osteoarthr. Cartil. 2009, 17, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Stabile, H.; Mitola, S.; Moroni, E.; Belleri, M.; Nicoli, S.; Coltrini, D.; Peri, F.; Pessi, A.; Orsatti, L.; Talamo, F.; et al. Bone morphogenic protein antagonist Drm/gremlin is a novel proangiogenic factor. Blood 2007, 109, 1834–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitola, S.; Ravelli, C.; Moroni, E.; Salvi, V.; Leali, D.; Ballmer-Hofer, K.; Zammataro, L.; Presta, M. Gremlin is a novel agonist of the major proangiogenic receptor VEGFR2. Blood 2010, 116, 3677–3680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravelli, C.; Mitola, S.; Corsini, M.; Presta, M. Involvement of αvβ3 integrin in gremlin-induced angiogenesis. Angiogenesis 2013, 16, 235–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethi, A.; Wordinger, R.J.; Clark, A.F. Gremlin utilizes canonical and non-canonical TGFβ signaling to induce lysyl oxidase (LOX) genes in human trabecular meshwork cells. Exp. Eye Res. 2013, 113, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, D.; Yamada, A.; Aizawa, R.; Funato, S.; Matsumoto, T.; Suzuki, W.; Takami, M.; Miyamoto, Y.; Suzawa, T.; Yamamoto, M.; et al. BMP2 Differentially Regulates the Expression of Gremlin1 and Gremlin2, the Negative Regulators of BMP Function, During Osteoblast Differentiation. Calcif. Tissue Int. 2012, 91, 88–96. [Google Scholar] [CrossRef]
- Salazar, V.S.; Gamer, L.W.; Rosen, V. BMP signalling in skeletal development, disease and repair. Nat. Rev. Endocrinol. 2016, 12, 203–221. [Google Scholar] [CrossRef]
- Church, R.H.; Krishnakumar, A.; Urbanek, A.; Geschwindner, S.; Meneely, J.; Bianchi, A.; Basta, B.; Monaghan, S.; Elliot, C.; Strömstedt, M.; et al. Gremlin1 preferentially binds to bone morphogenetic protein-2 (BMP-2) and BMP-4 over BMP-7. Biochem. J. 2015, 466, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.-J.; Dutra, E.H.; Mehta, S.; O’Brien, M.H.; Yadav, S. Age-related changes in the cartilage of the temporomandibular joint. GeroScience 2020, 42, 995–1004. [Google Scholar] [CrossRef]
- Nakase, T.; Miyaji, T.; Tomita, T.; Kaneko, M.; Kuriyama, K.; Myoui, A.; Sugamoto, K.; Ochi, T.; Yoshikawa, H. Localization of bone morphogenetic protein-2 in human osteoarthritic cartilage and osteophyte. Osteoarthr. Cartil. 2003, 11, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Van der Kraan, P.M.; van den Berg, W.B. Osteophytes: Relevance and biology. Osteoarthr. Cartil. 2007, 15, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Albilia, J.B.; Tenenbaum, H.C.; Clokie, C.M.L.; Walt, D.R.; Baker, G.I.; Psutka, D.J.; Backstein, D.; Peel, S.A.F. Serum levels of BMP-2, 4, 7 and AHSG in patients with degenerative joint disease requiring total arthroplasty of the hip and temporomandibular joints: Serum BMP and AHSG in DJD. J. Orthop. Res. 2013, 31, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Van Eegher, S.; Perez-Lozano, M.; Toillon, I.; Valour, D.; Pigenet, A.; Citadelle, D.; Bourrier, C.; Courtade-Gaïani, S.; Grégoire, L.; Cléret, D.; et al. The differentiation of prehypertrophic into hypertrophic chondrocytes drives an OA-remodeling program and IL-34 expression. Osteoarthr. Cartil. 2021, 29, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Schmal, H.; Pilz, I.H.; Mehlhorn, A.T.; Dovi-Akue, D.; Kirchhoff, C.; Südkamp, N.P.; Gerlach, U.; Niemeyer, P. Expression of BMP-receptor type 1A correlates with progress of osteoarthritis in human knee joints with focal cartilage lesions. Cytotherapy 2012, 14, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Pufe, T.; Petersen, W.; Tillmann, B.; Mentlein, R. The splice variants VEGF121 and VEGF189 of the angiogenic peptide vascular endothelial growth factor are expressed in osteoarthritic cartilage. Arthritis Rheum. 2001, 44, 1082–1088. [Google Scholar] [CrossRef]
- Enomoto, H.; Inoki, I.; Komiya, K.; Shiomi, T.; Ikeda, E.; Obata, K.-I.; Matsumoto, H.; Toyama, Y.; Okada, Y. Vascular endothelial growth factor isoforms and their receptors are expressed in human osteoarthritic cartilage. Am. J. Pathol. 2003, 162, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Pullig, O.; Weseloh, G.; Ronneberger, D.; Käkönen, S.; Swoboda, B. Chondrocyte differentiation in human osteoarthritis: Expression of osteocalcin in normal and osteoarthritic cartilage and bone. Calcif. Tissue Int. 2000, 67, 230–240. [Google Scholar] [CrossRef]
- Kirsch, T.; Swoboda, B.; Nah, H.-D. Activation of annexin II and V expression, terminal differentiation, mineralization and apoptosis in human osteoarthritic cartilage. Osteoarthr. Cartil. 2000, 8, 294–302. [Google Scholar] [CrossRef] [Green Version]
- Hilal, G.; Martel-Pelletier, J.; Pelletier, J.P.; Ranger, P.; Lajeunesse, D. Osteoblast-like cells from human subchondral osteoarthritic bone demonstrate an altered phenotype in vitro: Possible role in subchondral bone sclerosis. Arthritis Rheum. 1998, 41, 891–899. [Google Scholar] [CrossRef]
- Jaiprakash, A.; Prasadam, I.; Feng, J.Q.; Liu, Y.; Crawford, R.; Xiao, Y. Phenotypic characterization of osteoarthritic osteocytes from the sclerotic zones: A possible pathological role in subchondral bone sclerosis. Int. J. Biol. Sci. 2012, 8, 406–417. [Google Scholar] [CrossRef] [Green Version]
- da Silva Madaleno, C.; Jatzlau, J.; Knaus, P. BMP signalling in a mechanical context—Implications for bone biology. Bone 2020, 137, 115416. [Google Scholar] [CrossRef]
- Hayami, T.; Pickarski, M.; Wesolowski, G.A.; Mclane, J.; Bone, A.; Destefano, J.; Rodan, G.A.; Duong, L.T. The role of subchondral bone remodeling in osteoarthritis: Reduction of cartilage degeneration and prevention of osteophyte formation by alendronate in the rat anterior cruciate ligament transection model. Arthritis Rheum. 2004, 50, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Kadri, A.; Ea, H.K.; Bazille, C.; Hannouche, D.; Liote, F.; Cohen-Solal, M.E. Osteoprotegerin inhibits cartilage degradation through an effect on trabecular bone in murine experimental osteoarthritis. Arthritis Rheum. 2008, 58, 2379–2386. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.A.; Bonnet, C.S.; Turner, E.L.; Wilson, D.; Situ, M.; McWilliams, D.F. Angiogenesis in the synovium and at the osteochondral junction in osteoarthritis. Osteoarthr. Cartil. 2007, 15, 743–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, T.; Sato, M.; Kobayashi, M.; Yokoyama, M.; Tani, Y.; Mochida, J. Bevacizumab, an anti-vascular endothelial growth factor antibody, inhibits osteoarthritis. Arthritis Res. Ther. 2014, 16, 427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdmann, R.; Ozden, C.; Weidmann, J.; Schultze, A. Targeting the Gremlin-VEGFR2 axis—A promising strategy for multiple diseases?: Targeting the Gremlin-VEGFR2 axis. J. Pathol. 2015, 236, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Salvat, C.; Pigenet, A.; Humbert, L.; Berenbaum, F.; Thirion, S. Immature murine articular chondrocytes in primary culture: A new tool for investigating cartilage. Osteoarthr. Cartil. 2005, 13, 243–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosset, M.; Berenbaum, F.; Thirion, S.; Jacques, C. Primary culture and phenotyping of murine chondrocytes. Nat. Protoc. 2008, 3, 1253–1260. [Google Scholar] [CrossRef]
- Fermor, B.; Weinberg, J.B.; Pisetsky, D.S.; Misukonis, M.A.; Banes, A.J.; Guilak, F. The effects of static and intermittent compression on nitric oxide production in articular cartilage explants. J. Orthop. Res. 2001, 19, 729–737. [Google Scholar] [CrossRef]
- Shibakawa, A.; Yudoh, K.; Masuko-Hongo, K.; Kato, T.; Nishioka, K.; Nakamura, H. The role of subchondral bone resorption pits in osteoarthritis: MMP production by cells derived from bone marrow. Osteoarthr. Cartil. 2005, 13, 679–687. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Lozano, M.-L.; Sudre, L.; van Eegher, S.; Citadelle, D.; Pigenet, A.; Lafage-Proust, M.-H.; Pastoureau, P.; De Ceuninck, F.; Berenbaum, F.; Houard, X. Gremlin-1 and BMP-4 Overexpressed in Osteoarthritis Drive an Osteochondral-Remodeling Program in Osteoblasts and Hypertrophic Chondrocytes. Int. J. Mol. Sci. 2022, 23, 2084. https://doi.org/10.3390/ijms23042084
Pérez-Lozano M-L, Sudre L, van Eegher S, Citadelle D, Pigenet A, Lafage-Proust M-H, Pastoureau P, De Ceuninck F, Berenbaum F, Houard X. Gremlin-1 and BMP-4 Overexpressed in Osteoarthritis Drive an Osteochondral-Remodeling Program in Osteoblasts and Hypertrophic Chondrocytes. International Journal of Molecular Sciences. 2022; 23(4):2084. https://doi.org/10.3390/ijms23042084
Chicago/Turabian StylePérez-Lozano, Maria-Luisa, Laure Sudre, Sandy van Eegher, Danièle Citadelle, Audrey Pigenet, Marie-Helène Lafage-Proust, Philippe Pastoureau, Frédéric De Ceuninck, Francis Berenbaum, and Xavier Houard. 2022. "Gremlin-1 and BMP-4 Overexpressed in Osteoarthritis Drive an Osteochondral-Remodeling Program in Osteoblasts and Hypertrophic Chondrocytes" International Journal of Molecular Sciences 23, no. 4: 2084. https://doi.org/10.3390/ijms23042084
APA StylePérez-Lozano, M.-L., Sudre, L., van Eegher, S., Citadelle, D., Pigenet, A., Lafage-Proust, M.-H., Pastoureau, P., De Ceuninck, F., Berenbaum, F., & Houard, X. (2022). Gremlin-1 and BMP-4 Overexpressed in Osteoarthritis Drive an Osteochondral-Remodeling Program in Osteoblasts and Hypertrophic Chondrocytes. International Journal of Molecular Sciences, 23(4), 2084. https://doi.org/10.3390/ijms23042084